

Monosaccharide Composition and In Vitro Activity to HCT-116 Cells of Purslane Polysaccharides after a Covalent Chemical Selenylation
 ,
,
Abstract
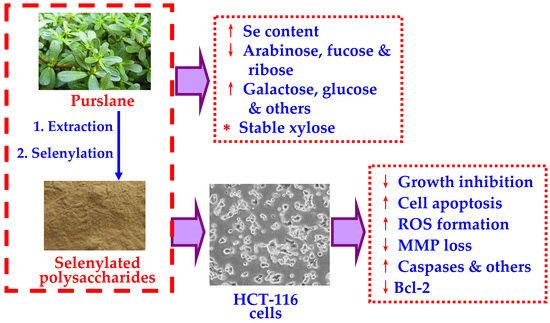
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Cell Line and Cell Culture
2.3. Preparation and Chemical Selenylation of P. oleracea Polysaccharides
2.4. Determination of Ash, Total Saccharide and Se Contents
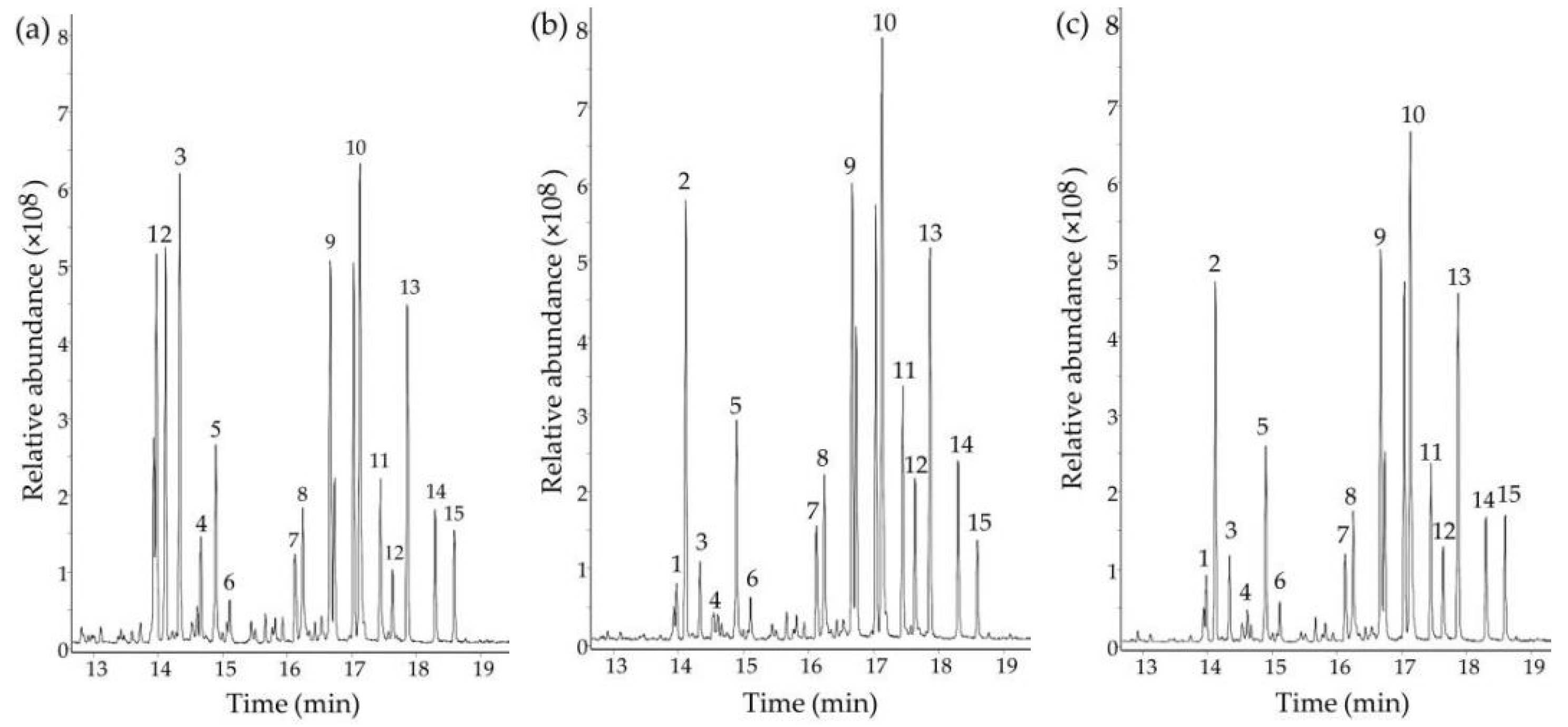
2.5. Determination of Monosaccharide Composition
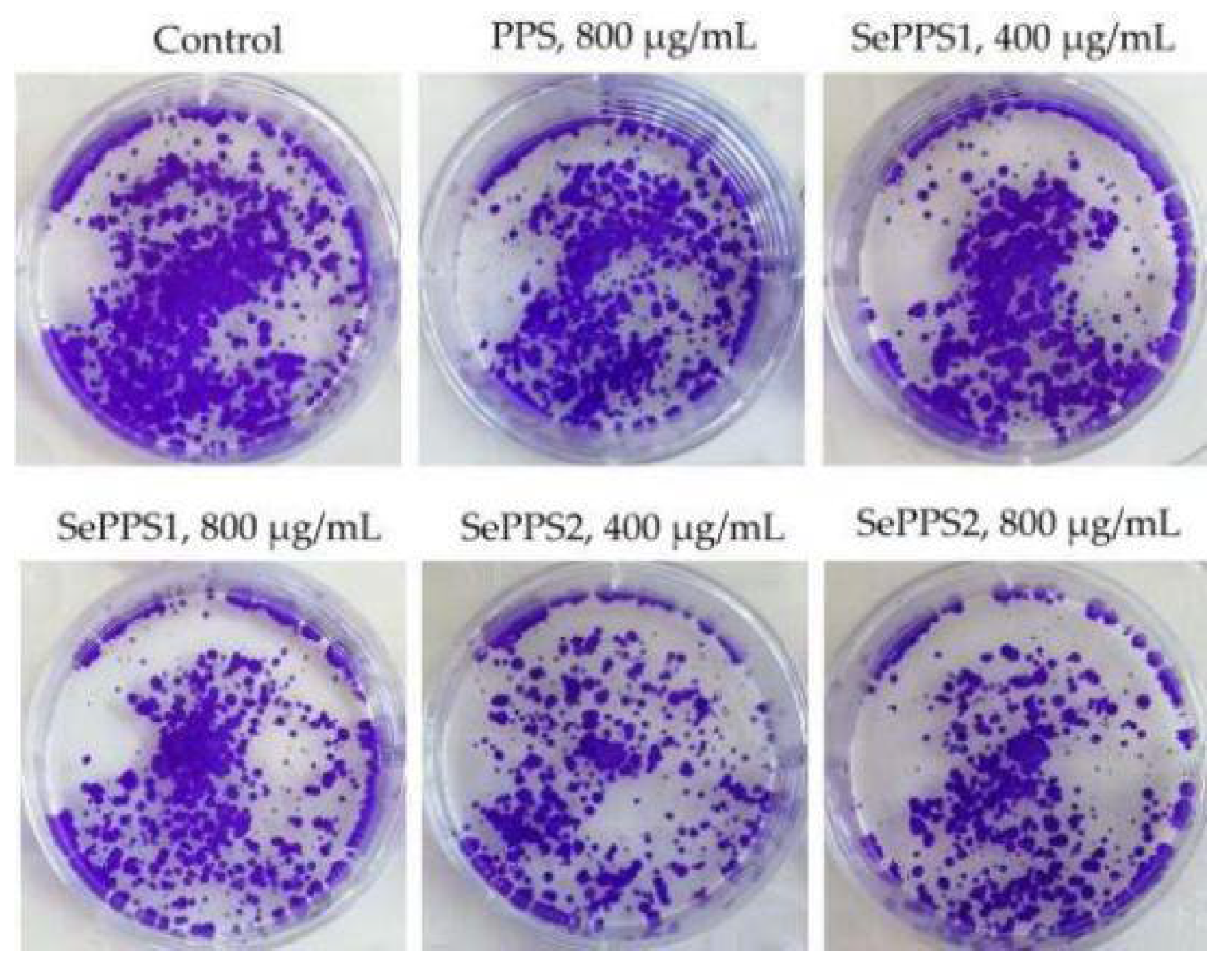
2.6. Assay of Growth Inhibition and Colony Formation
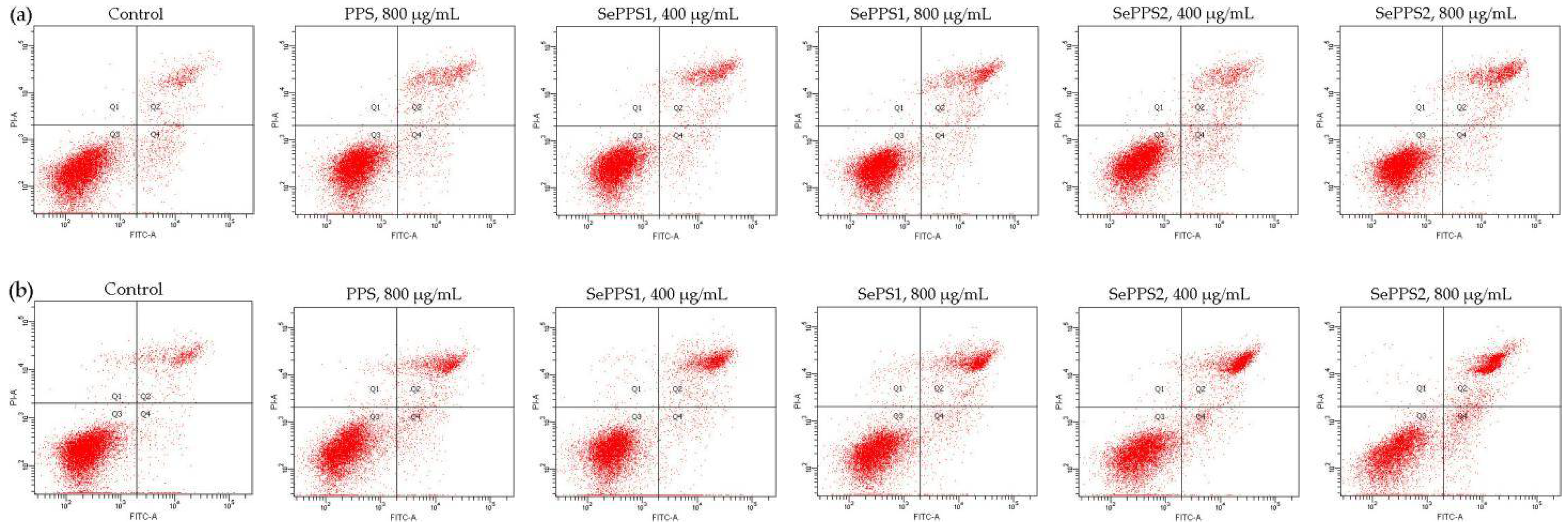
2.7. Assay of Cell Apoptosis by Flow Cytometry
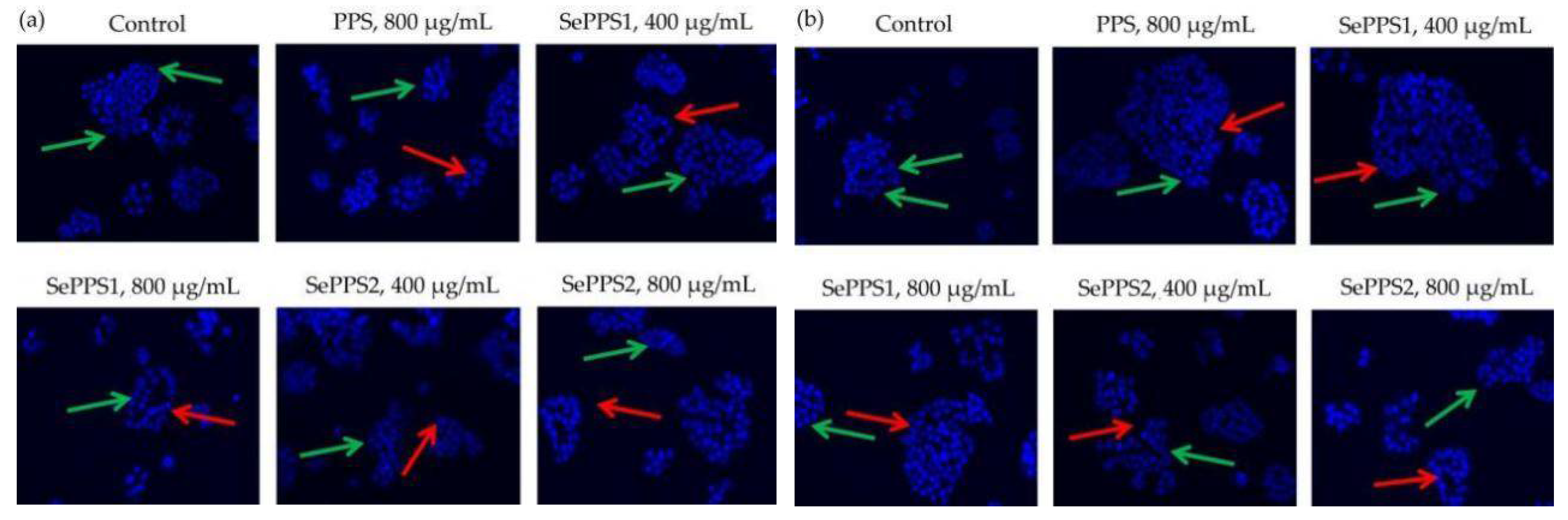
2.8. Hoechst 33258 Staining
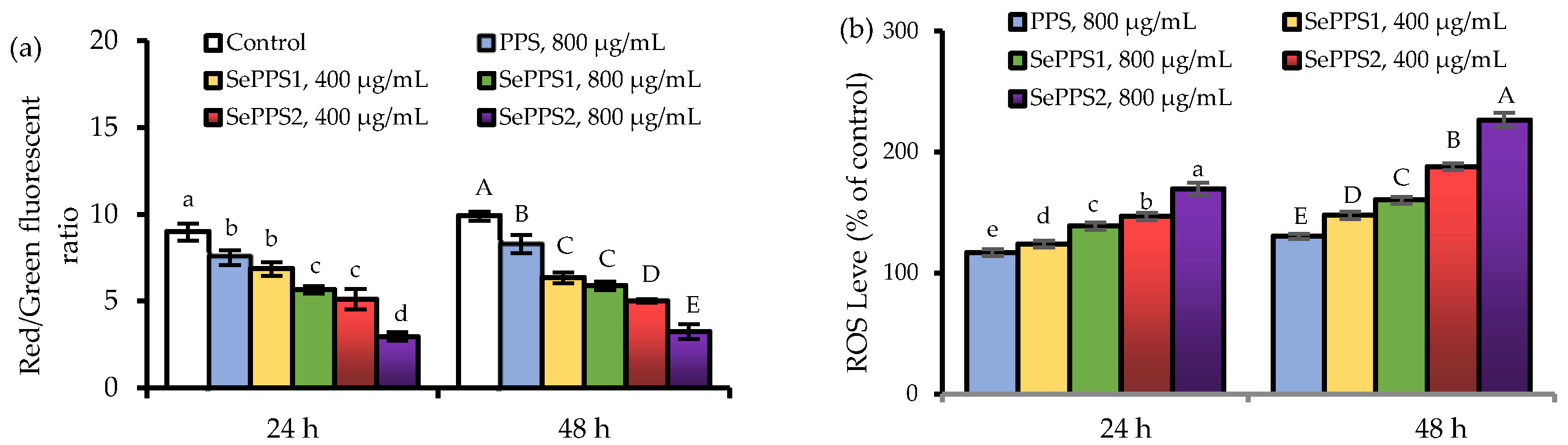
2.9. Assay of Intracellular ROS
2.10. Assay of Mitochondrial Membrane Potential Loss
2.11. Assay of Gene Expression Using RT-PCR Assay
2.12. Western-Blotting Analysis
2.13. Statistical Analysis
3. Results
3.1. Several Chemical Features of the Prepared Polysaccharide Samples
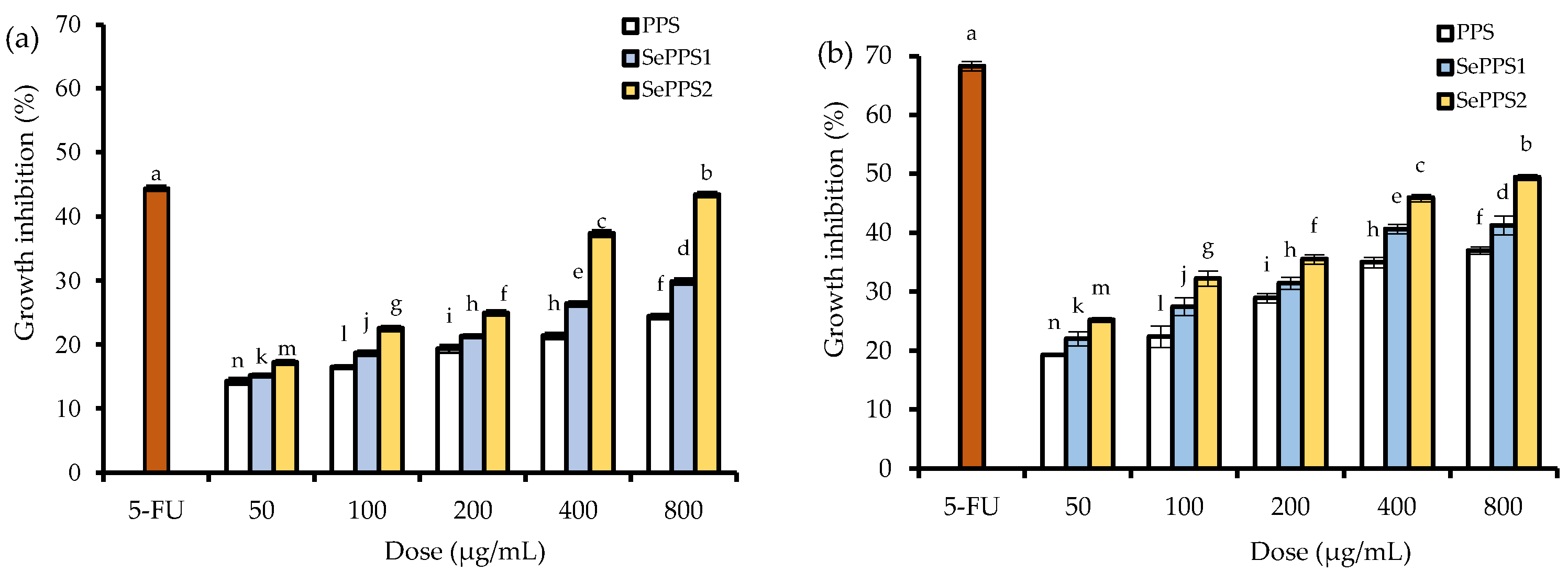
3.2. The Growth Inhibitory Effect of the Polysaccharide Samples on HCT-116 Cells
3.3. Effects of the Polysaccharide Samples on MMP Loss and ROS Formation
3.4. Apoptosis Induction of the Polysaccharide Samples towards HCT-116 Cells
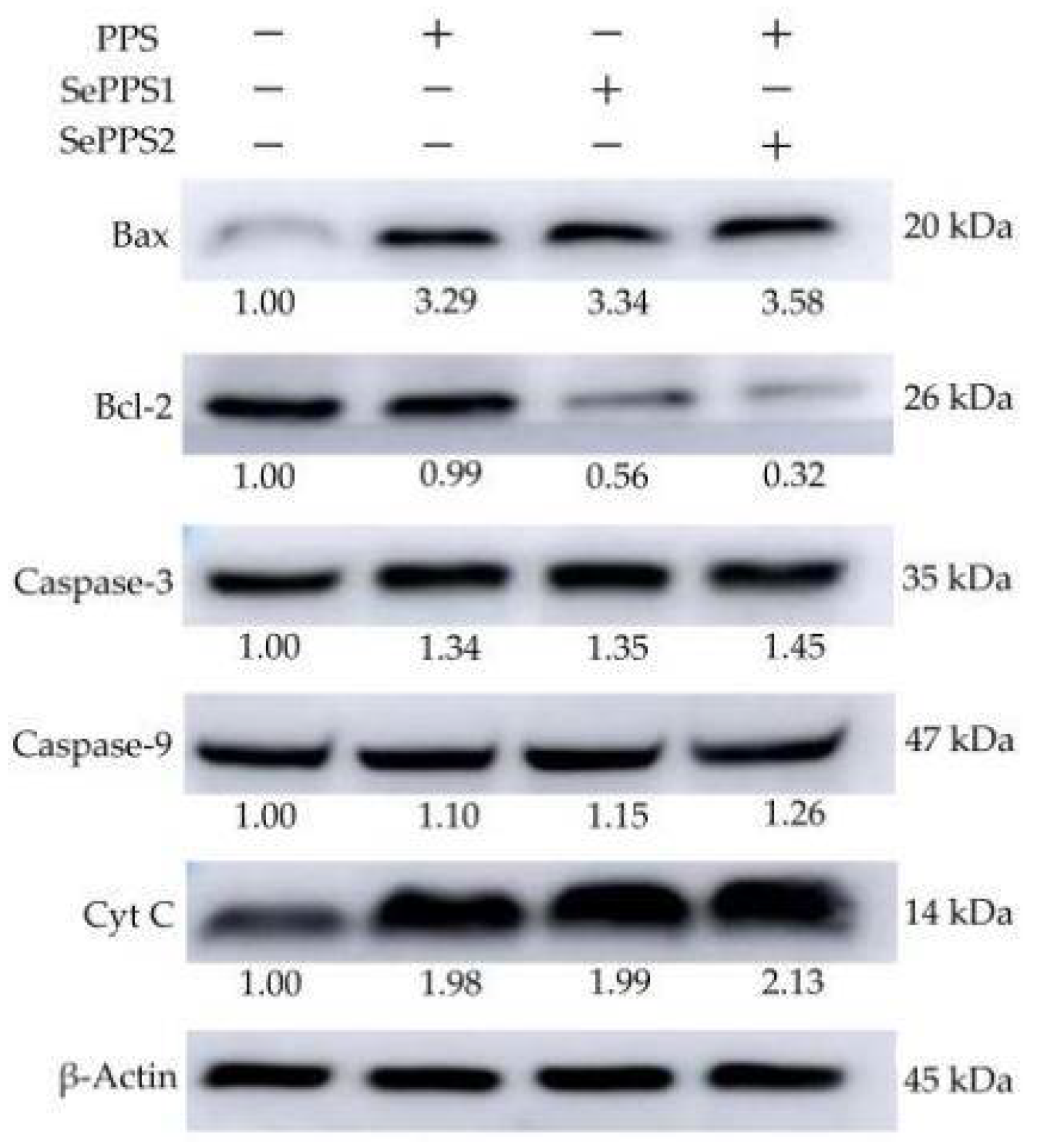
3.5. Effects of the Polysaccharide Samples on Gene and Protein Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Phillips, K.M.; Haytowitz, D.B.; Pehrsson, P.R. Implications of two different methods for analyzing total dietary fiber in foods for food composition databases. J. Food Compos. Anal. 2019, 84, e103253. [Google Scholar] [CrossRef]
- Jones, J.M.; García, C.G.; Braun, H.J. Perspective: Whole and refined grains and health-evidence supporting “make half your grains whole”. Adv. Nutr. 2020, 11, 492–506. [Google Scholar] [CrossRef]
- Dong, C.X.; Hayashi, K.; Lee, J.B.; Hayashi, T. Characterization of structures and antiviral effects of polysaccharides from Portulaca oleracea L. Chem. Pharm. Bull. 2010, 58, 507–510. [Google Scholar] [CrossRef]
- López-Legarda, X.; Arboleda-Echavarría, C.; Parra-Saldívar, R.; Rostro-Alanis, M.; Alzate, J.F.; Villa-Pulgarín, J.A.; Segura-Sánchez, F. Biotechnological production, characterization and in vitro antitumor activity of polysaccharides from a native strain of Lentinus crinitus. Int. J. Biol. Macromol. 2020, 164, 3133–3144. [Google Scholar] [CrossRef]
- Wang, W.S.; Song, X.L.; Gao, Z.; Zhao, H.J.; Wang, X.X.; Liu, M.; Jia, L. Anti-hyperlipidemic, antioxidant and organic protection effects of acidic-extractable polysaccharides from Dictyophora indusiata. Int. J. Biol. Macromol. 2019, 129, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.G.; Shen, Z.J.; Chen, X.P. Evaluation of free radicals scavenging and immunity-modulatory activities of Purslane polysaccharides. Int. J. Biol. Macromol. 2009, 45, 448–452. [Google Scholar] [CrossRef]
- Zeng, P.J.; Guo, Z.H.; Zeng, X.; Hao, C.; Zhang, Y.R.; Zhang, M.; Liu, Y.; Li, H.; Li, J.; Zhang, L. Chemical, biochemical, preclinical and clinical studies of Ganoderma lucidum polysaccharide as an approved drug for treating myopathy and other diseases in China. J. Cell. Mol. Med. 2018, 22, 3278–3297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.R.; Jiang, Y.F.; Zhang, M.; Zhang, L.J. Ganoderma sinense polysaccharide: An adjunctive drug used for cancer treatment. Prog. Mol. Biol. Transl. Sci. 2019, 163, 165–177. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Zhou, Y.X.; Zheng, Z.M.; Tang, W.Q.; Song, M.Z.; Wang, J.L.; Wang, K.P. Lentinan inhibited colon cancer growth by inducing endoplasmic reticulum stress-mediated autophagic cell death and apoptosis. Carbohydr. Polym. 2021, 267, e118154. [Google Scholar] [CrossRef]
- Hu, H.G.; Zhao, Q.L.; Pang, Z.C.; Xie, J.G.; Lin, L.G.; Yao, Q.S. Optimization extraction, characterization and anticancer activities of polysaccharides from mango pomace. Int. J. Biol. Macromol. 2018, 117, 1314–1325. [Google Scholar] [CrossRef]
- Zhao, R.; Zhang, T.; Ma, B.L.; Li, X. Antitumor activity of Portulaca oleracea L. polysaccharide on HeLa cells through inducing TLR4/NF-kappaB signaling. Nutr. Cancer 2017, 69, 131–139. [Google Scholar] [CrossRef]
- Song, Y.; Yang, Y.; Zhang, Y.Y.; Duan, L.S.; Zhou, C.; Ni, Y.Y.; Liao, X.J.; Li, Q.H.; Hu, X.S. Effect of acetylation on antioxidant and cytoprotective activity of polysaccharides isolated from pumpkin (Cucurbita pepo, lady godiva). Carbohydr. Polym. 2013, 98, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zong, M.H.; Lou, W.Y. Effect of acetylation modification on the emulsifying and antioxidant properties of polysaccharide from Millettia speciosa Champ. Food Hydrocoll. 2022, 124, e107217. [Google Scholar] [CrossRef]
- Liu, X.F.; Wang, X.Q.; Xu, X.F.; Zhang, X.W. Purification, antitumor and anti-inflammation activities of an alkali-soluble and carboxymethyl polysaccharide CMP33 from Poria cocos. Int. J. Biol. Macromol. 2019, 127, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.H.; Li, T.Y.; Lu, C.J.; Xu, H.P. Selenium-containing polymers: Perspectives toward diverse applications in both adaptive and biomedical materials. Macromolecules 2018, 51, 7435–7455. [Google Scholar] [CrossRef]
- Liu, J.; Chen, X.; Yue, C.G.; Hou, R.; Chen, J.; Lu, Y.; Li, X.P.; Li, R.J.; Lu, C.; Gao, Z.Z.; et al. Effect of selenylation modification on immune-enhancing activity of Atractylodes macrocephala polysaccharide. Int. J. Biol. Macromol. 2015, 72, 1435–1440. [Google Scholar] [CrossRef]
- Gong, F.Y.; Li, F.L.; Zhang, L.L.; Li, J.; Zhang, Z.; Wang, G.G. Hypoglycemic effects of crude polysaccharide from Purslane. Int. J. Mol. Sci. 2009, 10, 880–888. [Google Scholar] [CrossRef]
- Fiorito, S.; Epifano, F.; Preziuso, F.; Taddeo, V.A.; Genovese, S. Selenylated plant polysaccharides: A survey of their chemical and pharmacological properties. Phytochemistry 2018, 153, 1–10. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis; Association of Official Analytical Chemists (AOAC): Arlington, VA, USA, 2000. [Google Scholar]
- Ma, F.Y.; Zhang, S.Y.; Yao, Y.N.; Wen, Y.R.; Hu, W.P.; Zhang, J.; Liu, X.H.; Bell, A.E.; Tikkanen-Kaukanen, C. Chemical components and emulsification properties of mucilage from Dioscorea opposita Thunb. Food Chem. 2017, 228, 315–322. [Google Scholar] [CrossRef]
- Shand, C.A.; Balsam, M.; Hillier, S.J.; Hudson, G.; Newman, G.; Arthur, J.R.; Nicol, F. Aqua regia extractable selenium concentrations of some Scottish topsoils measured by ICP-MS and the relationship with mineral and organic soil components. J. Sci. Food Agric. 2010, 90, 972–980. [Google Scholar] [CrossRef]
- Liu, D.; Tang, W.; Yin, J.Y.; Nie, S.P.; Xie, M.Y. Monosaccharide composition analysis of polysaccharides from natural sources: Hydrolysis condition and detection method development. Food Hydrocoll. 2021, 116, e106641. [Google Scholar] [CrossRef]
- Pan, G.F.; Xie, Z.W.; Huang, S.X.; Tai, Y.L.; Cai, Q.S.; Jiang, W.; Sun, J.M.; Yuan, Y. Immune-enhancing effects of polysaccharides extracted from Lilium lancifolium Thunb. Int. Immunopharmacol. 2017, 52, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.L.; Liang, Y.T.; Wu, K.; Wang, C.X.; Zhang, T.; Peng, R.; Zou, F.D. Repressing PDCD4 activates JNK/ABCG2 pathway to induce chemoresistance to fluorouracil in colorectal cancer cells. Ann. Transl. Med. 2021, 9, e114. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Hu, J.H.; Li, M.; Zhu, S.Y.; Guo, S.J.; Guo, H.Y.; Wang, T.; Zang, Y.D.; Zang, J.; Wang, J.L. The role of Se content in improving anti-tumor activities and its potential mechanism for selenized Artemisia sphaerocephala polysaccharides. Food Funct. 2021, 12, 2058–2074. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Su, H.G.; Xu, Y.; Bao, T.; Zheng, X.D. Protective effect of wild raspberry (Rubus hirsutus Thunb.) extract against acrylamide-induced oxidative damage is potentiated after simulated gastrointestinal digestion. Food Chem. 2016, 196, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Che, Y.; Tian, Y.; Chen, R.; Xia, L.; Liu, F.; Su, Z.L. IL-22 ameliorated cardiomyocyte apoptosis in cardiac ischemia/reperfusion injury by blocking mitochondrial membrane potential decrease, inhibiting ROS and cytochrome C. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, e166171. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Quezada, R.; González-Castañeda, G.; Bahena, I.; Domínguez, A.; Domínguez-López, P.; Casas, E.; Betancourta, M.; Casillasa, F.; Rodríguezd, J.J.; Álvarezd, L.; et al. Effect of perfluorohexane sulfonate on pig oocyte maturation, gap-junctional intercellular communication, mitochondrial membrane potential and DNA damage in cumulus cells in vitro. Toxicol. Vitr. 2021, 70, e105011. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Zhao, L.; Xie, Y.H.; Chen, Z.; Yang, S.F.; Yin, B.; Li, G.M.; Gou, H.J.; Lin, S.Q.; Wu, J.Q. Antiviral activity of Portulaca oleracea L. extracts against porcine epidemic diarrhea virus by partial suppression on myd88/NF-kappab activation in vitro. Microb. Pathog. 2021, 154, e104832. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.Z.; Zhang, C.; Tian, W.J.; Liu, K.H.; Hou, R.R.; Yue, C.J.; Wu, Y.; Wang, D.Y.; Liu, J.G.; Hu, Y.L.; et al. The antioxidative and hepatoprotective effects comparison of Chinese angelica polysaccharide (CAP) and selenizing CAP (sCAP) in CCl4 induced hepatic injury mice. Int. J. Biol. Macromol. 2017, 97, 46–54. [Google Scholar] [CrossRef]
- Al-Wraikat, M.; Hou, C.; Zhao, G.T.; Lu, H.; Zhang, H.K.; Lei, Y.A.; Ali, Z.S.; Li, J.K. Degraded polysaccharide from Lycium barbarum L. leaves improve wheat dough structure and rheology. LWT-Food Sci. Technol. 2021, 145, e111372. [Google Scholar] [CrossRef]
- Guo, Y.L.; Wei, Y.; Cai, Z.X.; Hou, B.C.; Zhang, H.B. Stability of acidified milk drinks induced by various polysaccharide stabilizers: A review. Food Hydrocoll. 2021, 118, e106814. [Google Scholar] [CrossRef]
- Xia, T.; Liu, C.S.; Hu, Y.N.; Luo, Z.Y.; Chen, F.L.; Yuan, L.X.; Tan, X.M. Coix seed polysaccharides alleviate type 2 diabetes mellitus via gut microbiota-derived short-chain fatty acids activation of IGF1/PI3K/AKT signaling. Food Res. Int. 2021, 150, e110717. [Google Scholar] [CrossRef] [PubMed]
- Geng, Z.H.; Huang, L.; Song, M.B.; Song, Y.M. Cardiovascular effects in vitro of a polysaccharide from Salvia miltiorrhiza. Carbohydr. Polym. 2015, 121, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Padmanaban, D.; Samuel, A.; Sahayanathan, G.J.; Raja, K.; Chinnasamy, A. Anticancer effect of marine bivalves derived polysaccharides against human cancer cells. Biocatal. Agric. Biotechnol. 2022, 39, e102240. [Google Scholar] [CrossRef]
- Yuan, B.; Yang, X.Q.; Kou, M.; Lu, C.Y.; Wang, Y.Y.; Peng, J.; Chen, P.; Jiang, J.H. Selenylation of polysaccharide from the sweet potato and evaluation of antioxidant, antitumor, and antidiabetic activities. J. Agric. Food Chem. 2017, 65, 605–617. [Google Scholar] [CrossRef]
- He, N.W.; Shi, X.L.; Zhao, Y.; Tian, L.G.; Wang, D.Y.; Yang, X.B. Inhibitory effects and molecular mechanisms of selenium-containing tea polysaccharides on human breast cancer MCF-7 cells. J. Agric. Food Chem. 2013, 61, 579–588. [Google Scholar] [CrossRef]
- Liao, K.N.; Bian, Z.D.; Xie, D.G.; Peng, Q. A selenium-modified ginseng polysaccharide promotes the apoptosis in human promyelocytic leukemia (HL-60) cells via a mitochondrial-mediated pathway. Biol. Trace Elem. Res. 2017, 177, 64–71. [Google Scholar] [CrossRef]
- Moura, A.F.; Lima, K.S.B.; Sousa, T.S.; Marinho-Filho, J.D.B.; Pessoa, C.; Silveira, E.R.; Pessoab, O.D.L.; Costa-Lotufoa, L.V.; Moraes, M.O.; Araújo, A.J. In vitro antitumor effect of a lignan isolated from Combretum fruticosum, trachelogenin, in HCT-116 human colon cancer cells. Toxicol. Vitr. 2018, 47, 129–136. [Google Scholar] [CrossRef]
- Cakir, B.; Tunali-Akbay, T. Potential anticarcinogenic effect of goat milk-derived bioactive peptides on HCT-116 human colorectal carcinoma cell line. Anal. Biochem. 2021, 622, e114166. [Google Scholar] [CrossRef]
- Wang, J.L.; Li, Q.Y.; Bao, A.J.; Liu, X.R.; Zeng, J.Y.; Yang, X.P.; Yao, J.; Zhang, J.; Lei, Z.Q. Synthesis of selenium-containing Artemisia sphaerocephala polysaccharides: Solution conformation and anti-tumor activities in vitro. Carbohydr. Polym. 2016, 152, 70–78. [Google Scholar] [CrossRef]
- Yuan, C.F.; Wang, C.D.; Wang, J.J.; Kumar, V.; Anwar, F.; Xiao, F.X.; Mushtaq, G.; Liu, Y.F.; Kamal, M.A.; Yuan, D. Inhibition on the growth of human MDA-MB-231 breast cancer cells in vitro and tumor growth in a mouse xenograft model by Se-containing polysaccharides from Pyracantha fortuneana. Nutr. Res. 2016, 36, 1243–1254. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Kumari, P.; Ahmad, M. Apigenin attenuates edifenphos-induced toxicity by modulating ROS-mediated oxidative stress, mitochondrial dysfunction and caspase signal pathway in rat liver and kidney. Pestic. Biochem. Physiol. 2019, 159, e163172. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.M.; Wu, Q.; Yu, G.N.; Li, L.; Zhao, X.H.; Huang, X.T.; Mei, W.J. Polypyridyl Ruthenium(II) complex-induced mitochondrial membrane potential dissipation activates DNA damage-mediated apoptosis to inhibit liver cancer. Eur. J. Med. Chem. 2019, 164, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Kale, J.; Osterlund, E.J.; Andrews, D.W. BCL-2 family proteins: Changing partners in the dance towards death. Cell Death Differ. 2018, 25, 65–80. [Google Scholar] [CrossRef]
- Zeriouh, W.; Nani, A.; Belarbi, M.; Dumont, A.; de Rosny, C.; Aboura, I.; Aboura, I.; Ghanemi, F.Z.; Murtaza, B.; Patoli, D.; et al. Phenolic extract from oleaster (Olea europaea var. Sylvestris) leaves reduces colon cancer growth and induces caspase-dependent apoptosis in colon cancer cells via the mitochondrial apoptotic pathway. PLoS ONE 2017, 12, e0170823. [Google Scholar] [CrossRef]
- Guo, F.; Yang, F.; Zhu, Y.H. Scutellarein from Scutellaria barbata induces apoptosis of human colon cancer HCT116 cells through the ROS-mediated mitochondria-dependent pathway. Nat. Prod. Res. 2019, 33, 2372–2375. [Google Scholar] [CrossRef]
- Li, P.; Lin, Y.; Chen, Y.L. Brassica juncea polysaccharides induce apoptosis of colorectal cancer cells via mitochondrial- and caspase-dependent apoptosis pathways. Trop. J. Pharm. Res. 2021, 20, 2333–2338. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′-3′) | |
|---|---|---|
| Bax | Forward Reverse | CAGTTTGCTGGCAAAGTAGAAA CGAACTGGACAGTAACATGGAG |
| Bcl-2 | Forward Reverse | GACTTCGCCGAGATGTCCAG GAACTCAAAGAAGGCCACAATC |
| Caspase-3 | Forward Reverse | CTGAATGTTTCCCTGAGGTTTG CCAAAGATCATACATGGAAGCG |
| Caspase-9 | Forward Reverse | CTGCTGCGTGGTGGTCATTCTC TCGACCGACACAGGGCATCC |
| Cytochrome C | Forward Reverse | GCCAATAAGAACAAAGGCATCA TTAAGTCTGCCCTTTCTTCCTT |
| β-Actin | Forward Reverse | AACACCCCAGCCATGTACG ATGTCACGCACGATTTCCC |
| Monosaccharide | PPS (%) | SePPS1 (%) | SePPS2 (%) |
|---|---|---|---|
| Arabinose | 12.69 | 2.04 | 2.60 |
| Galactose | 12.33 | 17.29 | 17.00 |
| Fructose | 2.46 | 4.44 | 4.62 |
| Fucose | 2.16 | 0.86 | 0.63 |
| Galacturonic acid | 7.56 | 9.29 | 9.30 |
| Glucose | 21.92 | 27.77 | 28.39 |
| Glucuronic acid | 2.88 | 4.46 | 4.19 |
| Mannose | 2.46 | 2.92 | 2.67 |
| Ribose | 10.13 | 1.62 | 2.29 |
| Rhamnose | 9.06 | 10.73 | 10.04 |
| Xylose | 1.05 | 1.13 | 1.09 |
| Cell Group | Dose Level (μg/mL) | Total Apoptotic Cells (%) | |
|---|---|---|---|
| 24 h | 48 h | ||
| Control | None | 10.7 ± 0.2 | 9.5 ± 0.3 |
| PPS | 800 | 14.8 ± 0.8 | 20.2 ± 0.5 |
| SePPS1 | 400 | 16.8 ± 0.4 | 23.3 ± 0.3 |
| 800 | 17.2 ± 0.2 | 24.3 ± 0.4 | |
| SePPS2 | 400 | 18.1 ± 0.5 | 31.9 ± 0.7 |
| 800 | 20.5 ± 1.1 | 35.9 ± 0.9 | |
| Gene | Polysaccharide Sample and Used Dose Level (μg/mL) | ||||
|---|---|---|---|---|---|
| PPS, 800 | SePPS1, 400 | SePPS1, 800 | SePPS2, 400 | SePPS2, 800 | |
| Bax | 1.28 ± 0.02 | 1.58 ± 0.03 | 1.85 ± 0.04 | 1.89 ± 0.05 | 2.30 ± 0.01 |
| Bcl-2 | 0.96 ± 0.02 | 0.94 ± 0.03 | 0.82 ± 0.01 | 0.67 ± 0.03 | 0.61 ±0.01 |
| Caspase-3 | 1.37 ± 0.02 | 1.47 ± 0.03 | 1.75 ± 0.02 | 2.35 ± 0.03 | 2.39 ± 0.04 |
| Caspase-9 | 1.06 ± 0.02 | 1.18 ± 0.02 | 1.65 ± 0.01 | 1.78 ± 0.03 | 2.40 ± 0.03 |
| Cytochrome C | 1.04 ± 0.01 | 1.28 ± 0.02 | 1.36 ± 0.04 | 1.57 ± 0.02 | 1.64 ± 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.-Y.; Guan, Q.-Y.; Lin, Y.-R.; Zhao, J.-R.; Wang, L.; Zhang, Q.; Liu, H.-F.; Zhao, X.-H. Monosaccharide Composition and In Vitro Activity to HCT-116 Cells of Purslane Polysaccharides after a Covalent Chemical Selenylation. Foods 2022, 11, 3748. https://doi.org/10.3390/foods11233748
Li L-Y, Guan Q-Y, Lin Y-R, Zhao J-R, Wang L, Zhang Q, Liu H-F, Zhao X-H. Monosaccharide Composition and In Vitro Activity to HCT-116 Cells of Purslane Polysaccharides after a Covalent Chemical Selenylation. Foods. 2022; 11(23):3748. https://doi.org/10.3390/foods11233748
Chicago/Turabian StyleLi, Ling-Yu, Qing-Yun Guan, Ya-Ru Lin, Jun-Ren Zhao, Li Wang, Qiang Zhang, Hong-Fang Liu, and Xin-Huai Zhao. 2022. "Monosaccharide Composition and In Vitro Activity to HCT-116 Cells of Purslane Polysaccharides after a Covalent Chemical Selenylation" Foods 11, no. 23: 3748. https://doi.org/10.3390/foods11233748
APA StyleLi, L.-Y., Guan, Q.-Y., Lin, Y.-R., Zhao, J.-R., Wang, L., Zhang, Q., Liu, H.-F., & Zhao, X.-H. (2022). Monosaccharide Composition and In Vitro Activity to HCT-116 Cells of Purslane Polysaccharides after a Covalent Chemical Selenylation. Foods, 11(23), 3748. https://doi.org/10.3390/foods11233748