
The Effects of Storage Temperature, Light Illumination, and Low-Temperature Plasma on Fruit Rot and Change in Quality of Postharvest Gannan Navel Oranges

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Storage Temperature
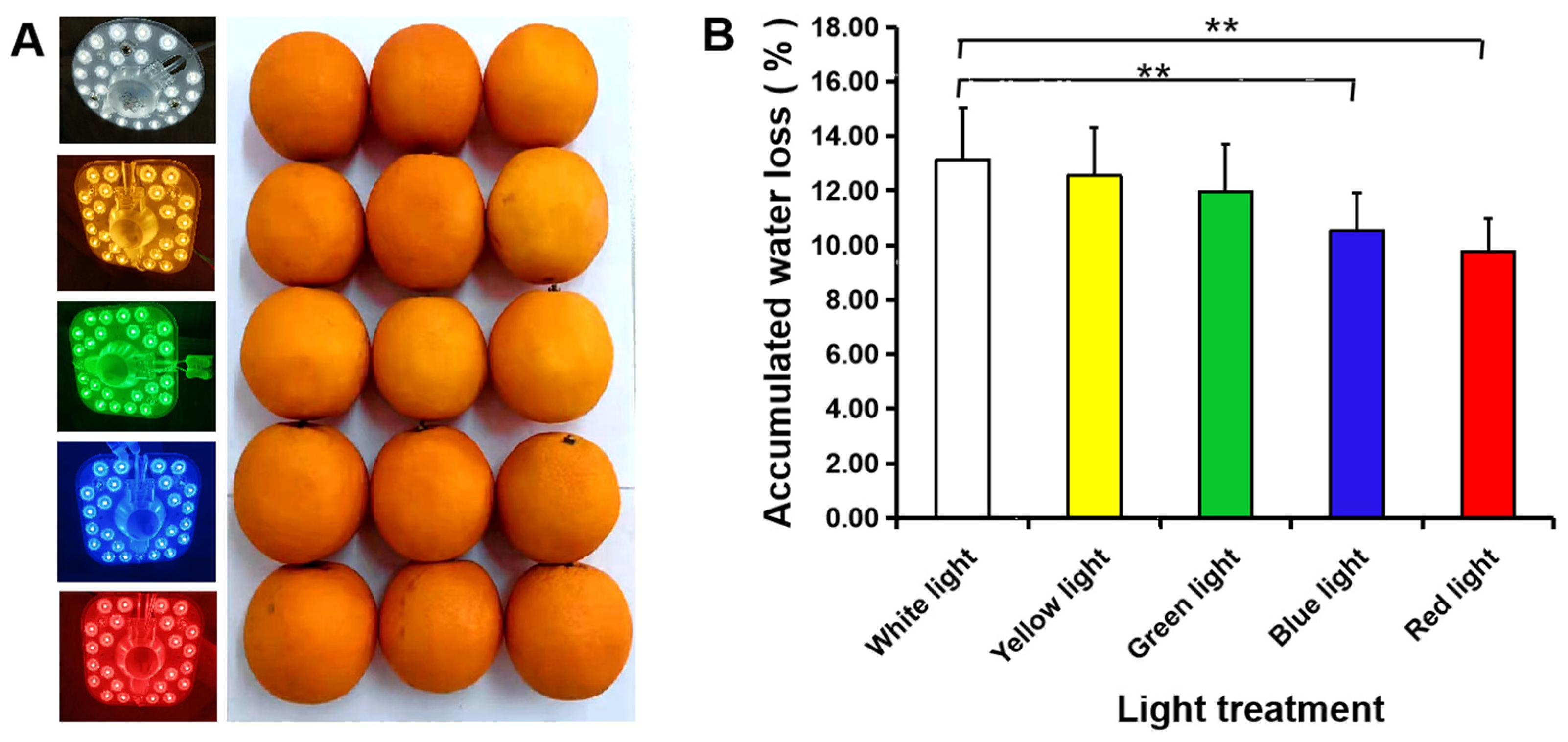
2.3. Light Treatment
2.4. Low-Temperature Plasma
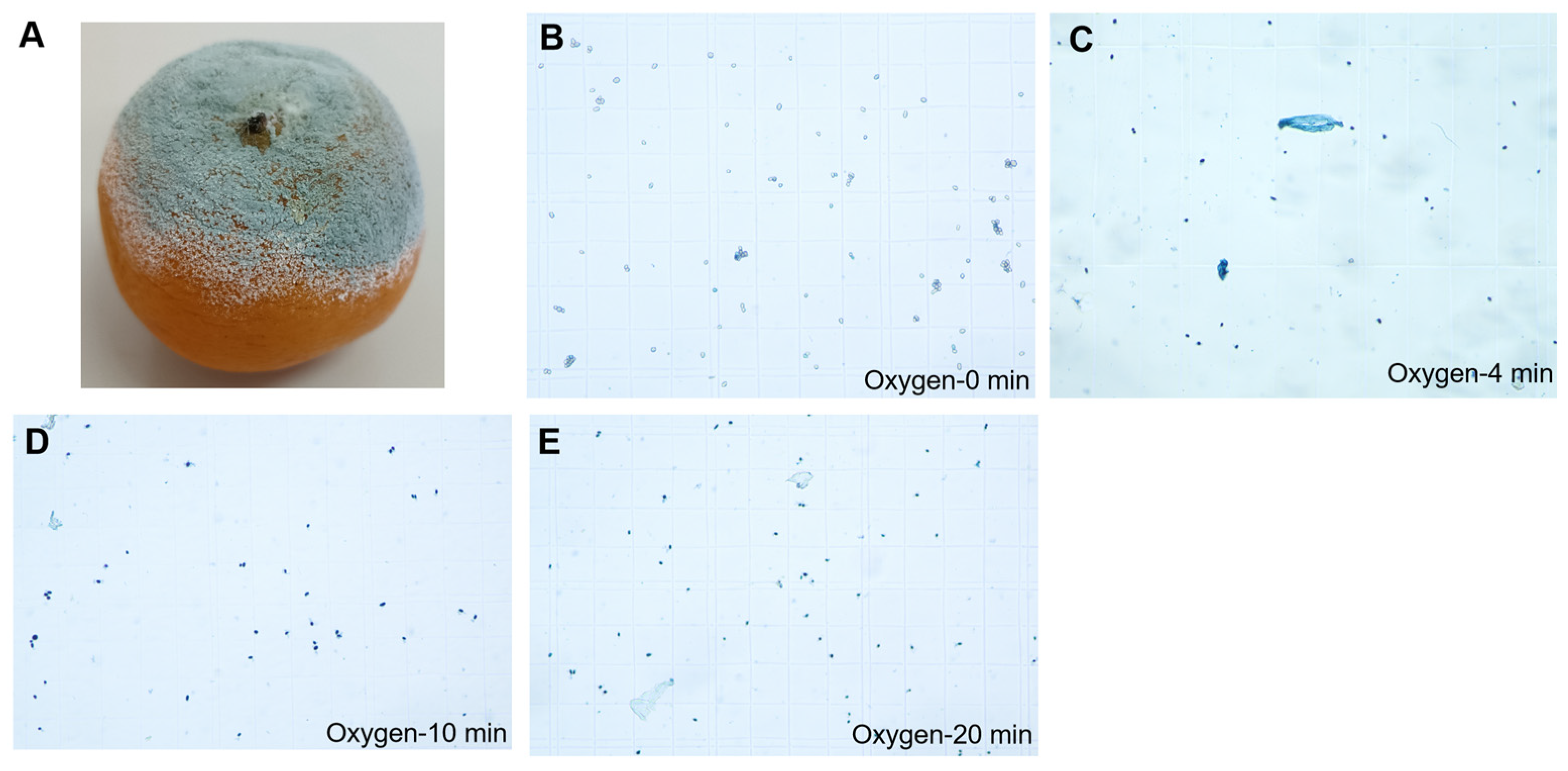
2.5. Fungal Infection
2.6. The Rupture of Pericarp Oil Cells
2.7. Firmness
2.8. Fruit Quality
2.9. Statistical Analysis
3. Results
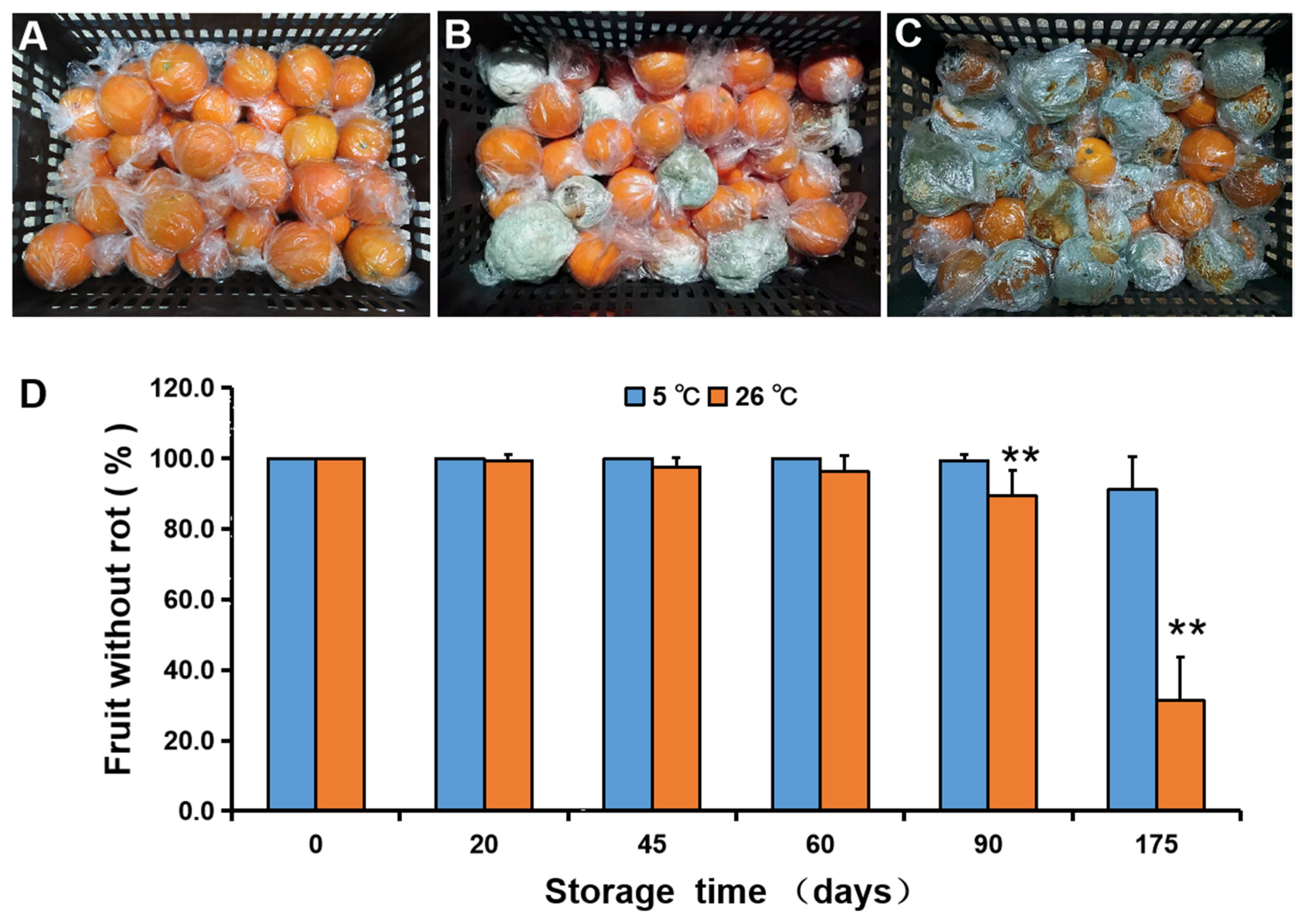
3.1. Fruit Rot
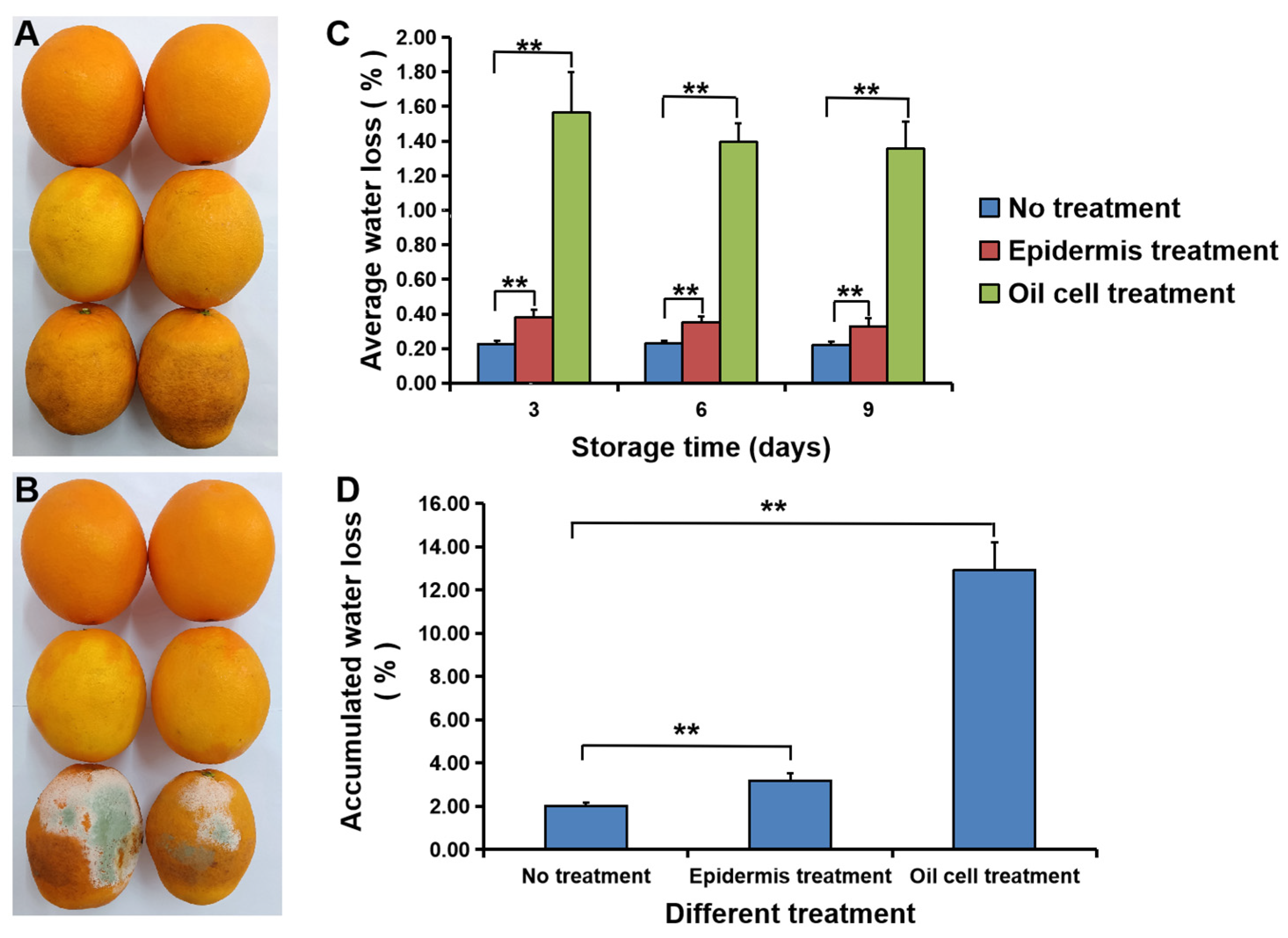
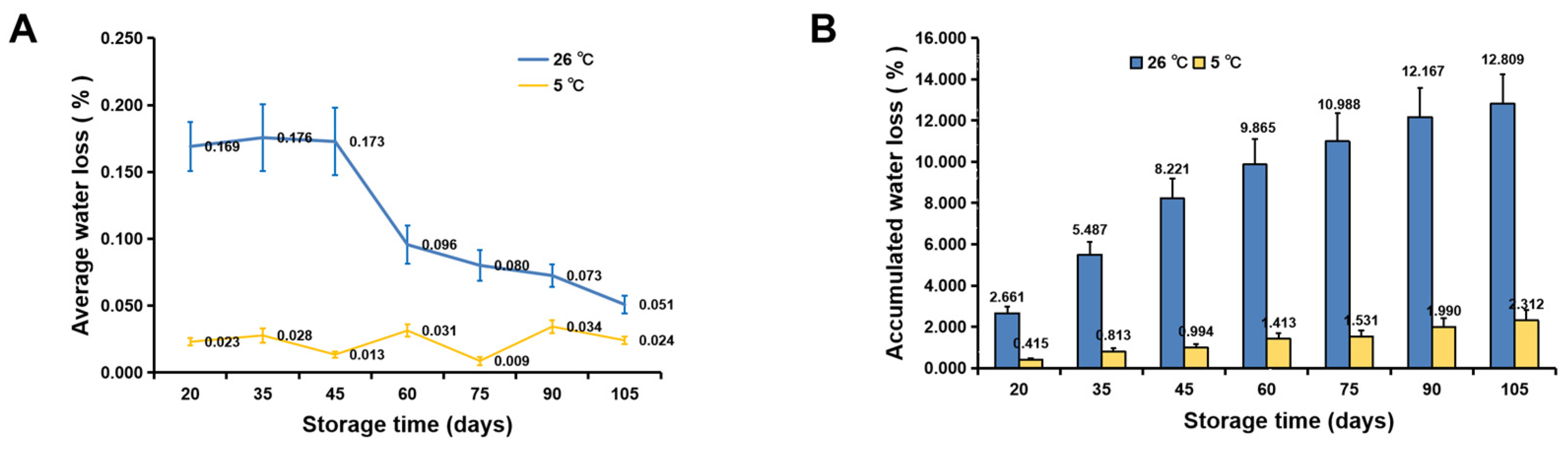
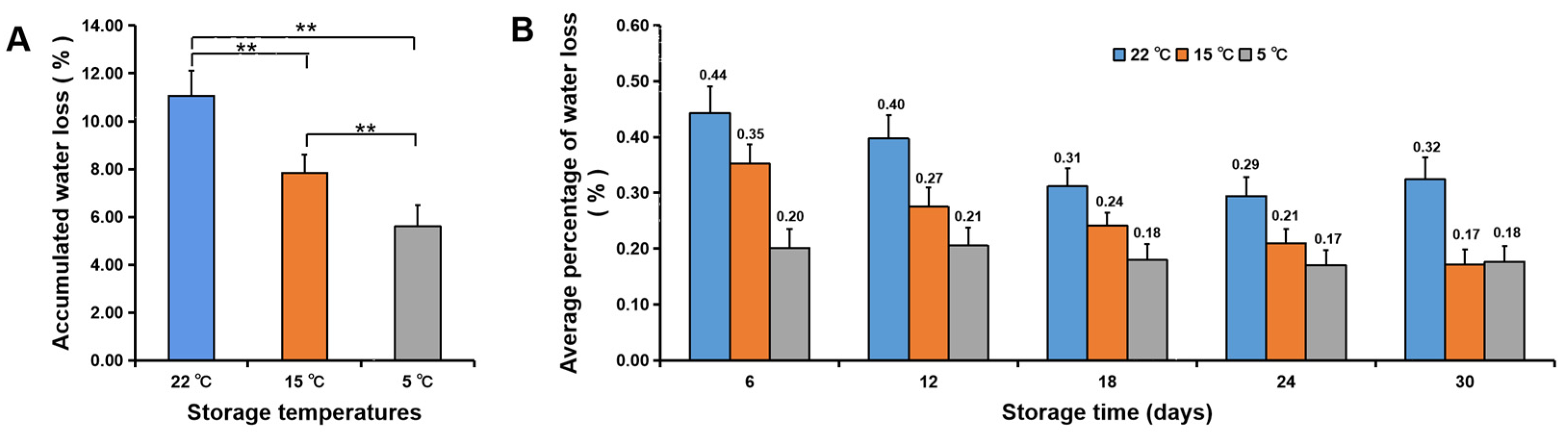
3.2. Water Loss
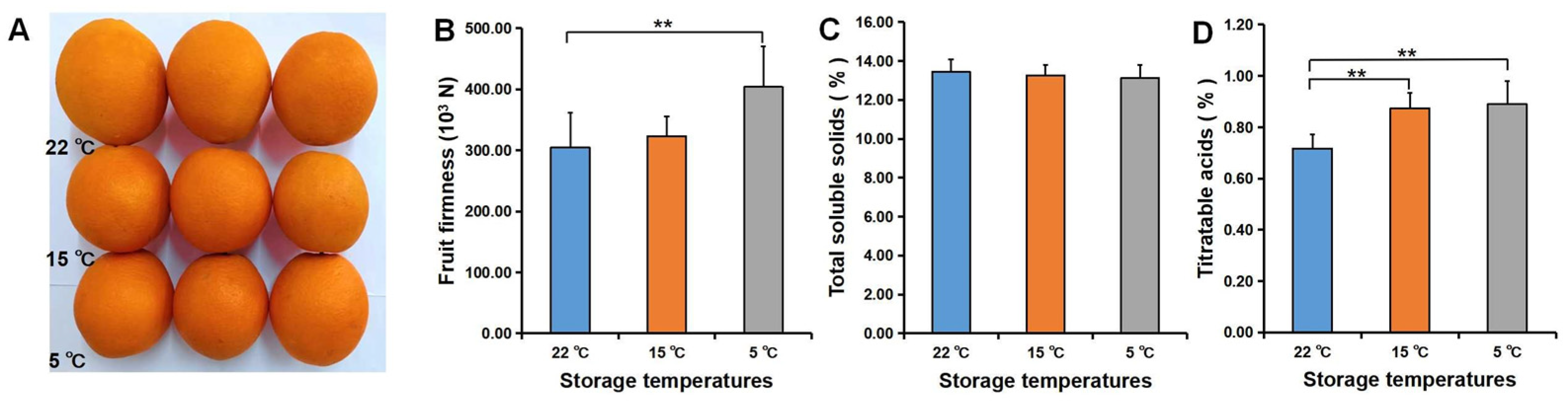
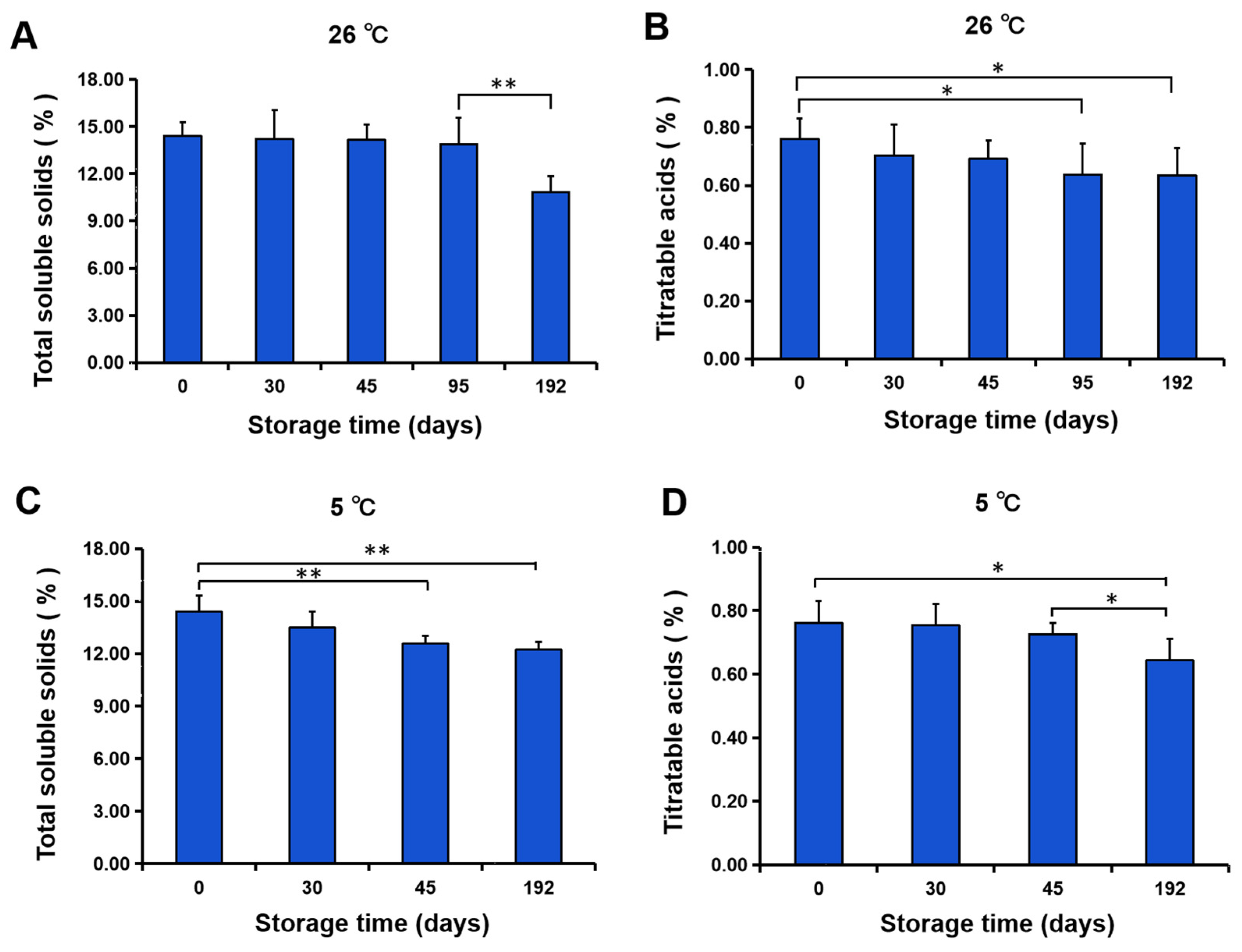
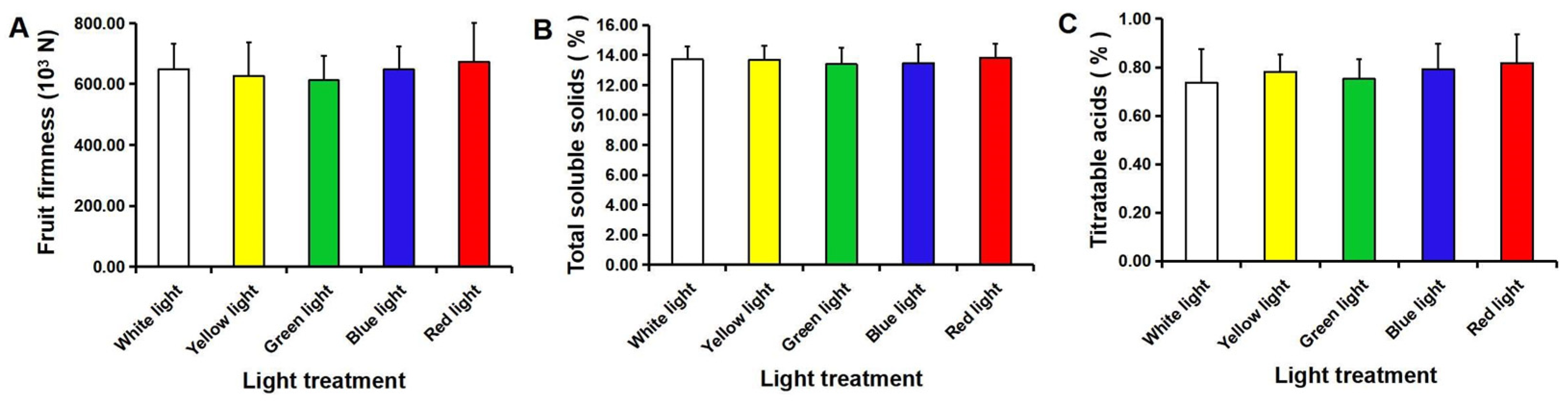
3.3. Fruit Quality
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Che, J.X.; Chen, X.M.; Ouyang, Q.L.; Tao, N.G. p-Anisaldehyde exerts its antifungal activity against Penicillium digitatum and Penicillium italicum by disrupting the cell wall integrity and membrane permeability. J. Microbiol. Biotechnol. 2019, 30, 878–884. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.L.; Fei, P.H.; Wang, Y.H.; Zheng, J.; Nie, J.Y.; Liao, W.Y.; Li, N.Z.; Gui, H.P.; Yan, H.J. Comparison of control effects of several chemicals on diseases of navel oranges during storage. South China Fruit 2015, 44, 38–40. [Google Scholar]
- Chen, C.Y.; Peng, X.; Chen, Y.H.; Wan, C.P.; Chen, J.Y. Screening of antifungal activity of extracts from 21 species of medicinal plants against Penicillium italicum of citrus fruit and the efficacy of Cynanchum atratum ethanol extracts for control. Acta Phytophylacica Sin. 2016, 43, 614–620. [Google Scholar]
- Peng, X.; Chen, Y.H.; Chen, J.Y. Antifungal mechanism of Cynanchum atratum Bunge extracts on Penicillium italicum and control effect on blue mold of navel oranges. Acta Phytopathol. Sin. 2017, 47, 398–405. [Google Scholar]
- Chen, C.Y.; Qi, W.W.; Peng, X.; Chen, J.Y.; Wan, C.P. Inhibitory effect of 7-Demethoxytylophorine on Penicillium italicum and its possible mechanism. Microorganisms 2019, 7, 36. [Google Scholar] [CrossRef]
- Liu, X.; Jiao, W.X.; Du, Y.M.; Chen, Q.M.; Su, Z.B.; Fu, M.R. Chlorine dioxide controls green mold caused by Penicillium digitatum in citrus fruit and the mechanism involved. J. Agric. Food Chem. 2020, 68, 13897–13905. [Google Scholar] [CrossRef]
- Qian, X.; Yang, Q.Y.; Zhang, Q.D.; Abdelhai, M.H.; Dhanasekaran, S.; Serwah, B.N.A.; Gu, N.; Zhang, H.Y. Elucidation of the initial growth process and the infection mechanism of Penicillium digitatum on postharvest citrus (Citrus reticulata Blanco). Microorganisms 2019, 7, 485. [Google Scholar] [CrossRef]
- Yang, Q.Y.; Qian, X.; Dhanasekaran, S.; Boateng, N.A.S.; Yan, X.L.; Zhu, H.M.; He, F.T.; Zhang, H.Y. Study on the infection mechanism of Penicillium Digitatum on postharvest citrus (Citrus reticulata Blanco) based on transcriptomics. Microorganisms 2019, 7, 672. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Sun, X.P.; Xu, Q.; Candelas, L.G.; Li, H.Y. The pH signaling transcription factor PacC is required for full virulence in Penicillium digitatum. Appl. Microbiol. Biotechnol. 2013, 97, 9087–9098. [Google Scholar] [CrossRef]
- Chen, D.; Förster, H.; Adaskaveg, J.E. Natamycin, a biofungicide for managing major postharvest fruit decays of citrus. Plant Dis. 2021, 105, 1408–1414. [Google Scholar] [CrossRef]
- Yang, Y.C.; Li, K.; Liu, C.X.; Cheng, F.; Liu, C.; Quan, W.J.; Xue, Y.H.; Zou, K.; Liu, S.P. Sanxiapeptin, a linear pentapeptide from Penicillium oxalicum, inhibited the growth of citrus green mold. Food Chem. 2022, 366, 130541. [Google Scholar] [CrossRef] [PubMed]
- Montesinos-Herrero, C.; Del Río, M.Á.; Rojas-Argudo, C.; Palou, L. Short exposure to high CO2 and O2 at curing temperature to control postharvest diseases of citrus fruit. Plant Dis. 2012, 96, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Smilanick, J.L.; Mansour, M.F. Influence of temperature and humidity on survival of Penicillium digitatum and Geotrichum citri-aurantii. Plant Dis. 2007, 91, 990–996. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.S.; Zhang, M. Advances in the study of the effect of temperature shock treatments on storage quality of postharvest fruit and vegetables. Food Sci. 2016, 37, 230–238. [Google Scholar]
- Lin, L.P.; Ye, M.; Chen, Y.L.; Zheng, F.Y.; Wu, G.P. Research of the preservation effect on postharvest Gannan navel oranges by hot water treatment. Food Ferment. Ind. 2017, 43, 252–256, 265. [Google Scholar]
- Yun, Z.; Gao, H.; Liu, P.; Liu, S.; Luo, T.; Jin, S.; Xu, Q.; Xu, J.; Cheng, Y.; Deng, X. Comparative proteomic and metabolomic profiling of citrus fruit with enhancement of disease resistance by postharvest heat treatment. BMC Plant Biol. 2013, 13, 44. [Google Scholar] [CrossRef]
- Queb-González, D.B.; Lopez-Malo, A.; Sosa-Morales, M.E.; Villa-Rojas, R. Postharvest heat treatments to inhibit Penicillium digitatum growth and maintain quality of Mandarin (Citrus reticulata blanco). Heliyon 2020, 6, e03166. [Google Scholar] [CrossRef]
- Ballester, A.R.; Lafuente, M.T. LED blue light-induced changes in phenolics and ethylene in citrus fruit: Implication in elicited resistance against Penicillium digitatum infection. Food Chem. 2017, 218, 575–583. [Google Scholar] [CrossRef]
- Lafuente, M.T.; Alférez, F.; González-Candelas, L. Light-emitting diode blue light alters the ability of Penicillium digitatum to infect citrus fruit. Photochem. Photobiol. 2018, 94, 1003–1009. [Google Scholar] [CrossRef]
- Chen, J.Y.; Shen, Y.; Chen, C.; Wan, C. Inhibition of key citrus postharvest fungal strains by plant extracts in vitro and in vivo: A review. Plants 2019, 8, 26. [Google Scholar] [CrossRef]
- Costa, J.H.; Wassano, C.I.; Angolini, C.F.F.; Scherlach, K.; Hertweck, C.; Pacheco, F.T. Antifungal potential of secondary metabolites involved in the interaction between citrus pathogens. Sci. Rep. 2019, 9, 18647. [Google Scholar] [CrossRef] [PubMed]
- Chen, O.; Yi, L.H.; Deng, L.L.; Ruan, C.Q.; Zeng, K.F. Screening antagonistic yeasts against citrus green mold and the possible biocontrol mechanisms of Pichia galeiformis (BAF03). J. Sci. Food Agric. 2020, 100, 3812–3821. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, Y.X.; Xu, Y.; Lu, H.L.; Sang, Y.L.; Li, J.P.; Xu, R.; Wang, Q.; Li, Y.Y.; Wang, X.Q.; et al. Insights into the fungicidal activity of low-temperature plasma against the pathogen of navel orange fruit mildew. In Proceedings of the 16th Annual Conference of China Electrotechnical Society; Liang, X., Li, Y., He, J., Yang, Q., Eds.; Lecture Notes in Electrical Engineering; Springer: Singapore, 2022; Volume 890, pp. 834–843. [Google Scholar]
- Yun, Z.; Jin, S.; Ding, Y.D.; Wang, Z.; Gao, H.J.; Pan, Z.L.; Xu, J.; Cheng, Y.X.; Deng, X.X. Comparative transcriptomics and proteomics analysis of citrus fruit, to improve understanding of the effect of low temperature on maintaining fruit quality during lengthy post-harvest storage. J. Exp. Bot. 2012, 63, 2873–2893. [Google Scholar] [CrossRef] [PubMed]
- De Ramón-Carbonell, M.; López-Pérez, M.; González-Candelas, L.; Sánchez-Torres, P. PdMFS1 transporter contributes to Penicilliun digitatum fungicide resistance and fungal virulence during citrus fruit infection. J. Fungi 2019, 5, 100. [Google Scholar] [CrossRef]
- Harries, E.; Gandía, M.; Carmona, L.; Marcos, J.F. The Penicillium digitatum protein O-mannosyltransferase Pmt2 is required for cell wall integrity, conidiogenesis, virulence and sensitivity to the antifungal peptide PAF26. Mol. Plant Pathol. 2015, 16, 748–761. [Google Scholar] [CrossRef]
- Besil, N.; Pérez-Parada, A.; Cesio, V.; Varela, P.; Rivas, F.; Heinzen, H. Degradation of imazalil, orthophenylphenol and pyrimethanil in Clementine mandarins under conventional postharvest industrial conditions at 4 °C. Food Chem. 2016, 194, 1132–1137. [Google Scholar] [CrossRef]
- Cheng, Y.J.; Qin, W.X.; Zhao, X.; Jin, M.M.; Zhang, M. Preservation properties of ‘Tarocco’ oranges with intermittent heating treatment on temperature-shift transportation. Food Ferment. Ind. 2016, 42, 196–203. [Google Scholar]
- Liu, C.C.; Wan, H.J.; Yang, Y.X.; Ye, Q.J.; Zhou, G.Z.; Wang, X.R.; Ahammed, G.J.; Cheng, Y. Post-Harvest LED Light Irradiation Affects Firmness, Bioactive Substances, and Amino Acid Compositions in Chili Pepper (Capsicum annum L.). Foods 2022, 11, 2712. [Google Scholar] [CrossRef]
- Pristijono, P.; Bowyer, M.C.; Papoutsis, K.; Scarlett, C.J.; Vuong, Q.V.; Stathopoulos, C.E.; Golding, J.B. Improving the storage quality of Tahitian limes (Citrus latifolia) by pre-storage UV-C irradiation. J. Food Sci. Technol. 2019, 56, 1438–1444. [Google Scholar] [CrossRef]
- Zhao, S.S.; Han, X.Y.; Liu, B.; Guan, W.Q.; Dai, Q.Y.; Li, J.H.; Zhang, R. Mechanism analysis on the action of alternating magnetic field in inhibiting chilling injury of bananas. Jiangsu J. Agric. Sci. 2021, 37, 739–745. [Google Scholar]
- Ding, Y.D.; Chang, J.W.; Ma, Q.L.; Chen, L.L.; Liu, S.; Jin, S.Z.; Han, J.W.; Xu, R.W.; Zhu, A.D.; Guo, J.; et al. Network analysis of postharvest senescence process in citrus fruit revealed by transcriptomic and metabolomic profiling. Plant Physiol. 2015, 168, 357–376. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.H.; Zhang, J.; Yang, L.Z.; Wang, X.Y.; Fu, F.H.; Wang, R.R.; Zhang, Q.; Shan, Y. Changes in cuticle components and morphology of ‘Satsuma’ mandarin (Citrus unshiu) during ambient storage and their potential role on Penicillium digitatum infection. Molecules 2020, 25, 412. [Google Scholar] [CrossRef] [PubMed]
- He, Y.Z.; Li, Z.R.; Tan, F.J.; Liu, H.; Zhu, M.; Yang, H.B.; Bi, G.; Wan, H.L.; Wang, J.Q.; Xu, R.W.; et al. Fatty acid metabolic flux and lipid peroxidation homeostasis maintain the biomembrane stability to improve citrus fruit storage performance. Food Chem. 2019, 292, 314–324. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.L.; Ding, Y.D.; Chang, J.W.; Sun, X.H.; Zhang, L.; Wei, Q.J.; Cheng, Y.J.; Chen, L.L.; Xu, J.; Deng, X.X. Comprehensive insights on how 2,4-dichlorophenoxyacetic acid retards senescence in post-harvest citrus fruit using transcriptomic and proteomic approaches. J. Exp. Bot. 2014, 65, 61–74. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, H.; Zhu, F.; Xu, R.; Cheng, Y. Transcript profiles analysis of citrus aquaporins in response to fruit water loss during storage. Plant Biol. 2021, 23, 819–830. [Google Scholar] [CrossRef]
- Chen, M.; Xiang, M.L.; Chen, Y.H.; Liu, Y.; Gao, Y.; Liu, B.; Chen, J.Y. Effects of storage time and logistics temperature on shelf-life and fruit quality of ‘Newhall’ navel oranges. Acta Agric. Univ. Jiangxiensis 2014, 6, 1223–1228. [Google Scholar]
- Hauswirth, A.; Köhler, R.; Ten Bosch, L.; Avramidis, G.; Gerhard, C. Spectroscopic investigation of the impact of cold plasma treatment at atmospheric pressure on sucrose and glucose. Foods 2022, 11, 2786. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Soluble Solids | Titratable Acids | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | Air | Oxygen | Control | Air | Oxygen | |||||
| 0 min | 3 min | 6 min | 3 min | 6 min | 0 min | 3 min | 6 min | 3 min | 6 min | |
| 1 | 10.8 | 13.3 | 11.9 | 11.9 | 10.2 | 0.67 | 0.91 | 0.76 | 0.73 | 0.62 |
| 2 | 13.3 | 13.4 | 12.8 | 11.8 | 11.8 | 0.81 | 0.95 | 0.86 | 0.74 | 0.73 |
| 3 | 12.6 | 10.6 | 12.5 | 11.6 | 10.7 | 0.75 | 0.57 | 0.6 | 0.64 | 0.68 |
| 4 | 13.0 | 11.9 | 11.4 | 11.9 | 13.1 | 0.73 | 0.74 | 0.53 | 0.65 | 0.71 |
| 5 | 11.6 | 12.0 | 11.0 | 11.7 | 11.2 | 0.85 | 0.76 | 0.67 | 0.75 | 0.65 |
| 6 | 12.1 | 11.6 | 12.2 | 10.8 | 12.4 | 0.77 | 0.65 | 0.67 | 0.69 | 0.72 |
| Mean | 12.23 | 12.13 | 11.97 | 11.62 | 11.57 | 0.76 | 0.76 | 0.68 | 0.70 | 0.69 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Li, Y.; Xu, Y.; Sang, Y.; Mei, S.; Xu, C.; Yu, X.; Pan, T.; Cheng, C.; Zhang, J.; et al. The Effects of Storage Temperature, Light Illumination, and Low-Temperature Plasma on Fruit Rot and Change in Quality of Postharvest Gannan Navel Oranges. Foods 2022, 11, 3707. https://doi.org/10.3390/foods11223707
Sun Y, Li Y, Xu Y, Sang Y, Mei S, Xu C, Yu X, Pan T, Cheng C, Zhang J, et al. The Effects of Storage Temperature, Light Illumination, and Low-Temperature Plasma on Fruit Rot and Change in Quality of Postharvest Gannan Navel Oranges. Foods. 2022; 11(22):3707. https://doi.org/10.3390/foods11223707
Chicago/Turabian StyleSun, Ying, Yuanyuan Li, Yu Xu, Yali Sang, Siyi Mei, Chaobin Xu, Xingguo Yu, Taoyu Pan, Chen Cheng, Jun Zhang, and et al. 2022. "The Effects of Storage Temperature, Light Illumination, and Low-Temperature Plasma on Fruit Rot and Change in Quality of Postharvest Gannan Navel Oranges" Foods 11, no. 22: 3707. https://doi.org/10.3390/foods11223707
APA StyleSun, Y., Li, Y., Xu, Y., Sang, Y., Mei, S., Xu, C., Yu, X., Pan, T., Cheng, C., Zhang, J., Jiang, Y., & Gao, Z. (2022). The Effects of Storage Temperature, Light Illumination, and Low-Temperature Plasma on Fruit Rot and Change in Quality of Postharvest Gannan Navel Oranges. Foods, 11(22), 3707. https://doi.org/10.3390/foods11223707