
Efficient Utilization of Fruit Peels for the Bioproduction of D-Allulose and D-Mannitol

Abstract
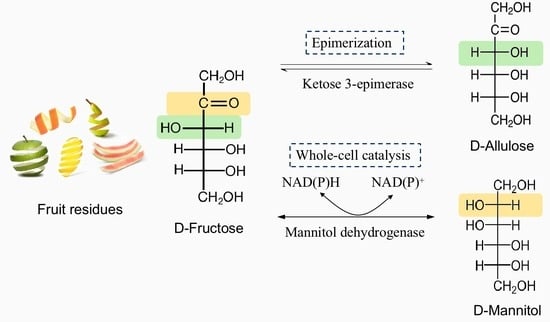
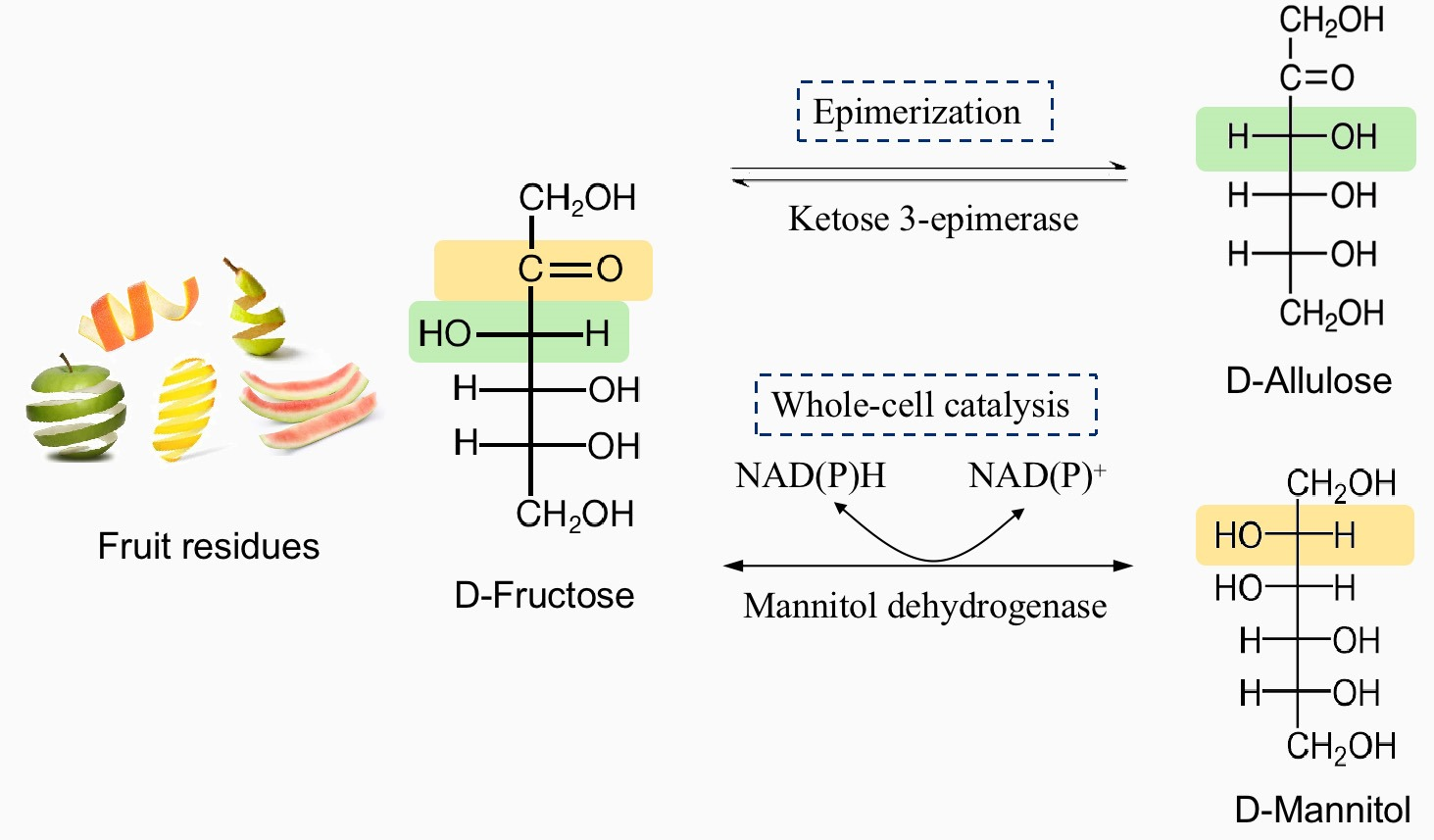
1. Introduction
2. Materials and Methods
2.1. Strains, Plasmid, Reagents, and Materials
2.2. Heterogeneous Expression of KEase in E. coli
2.3. Purification of KEase
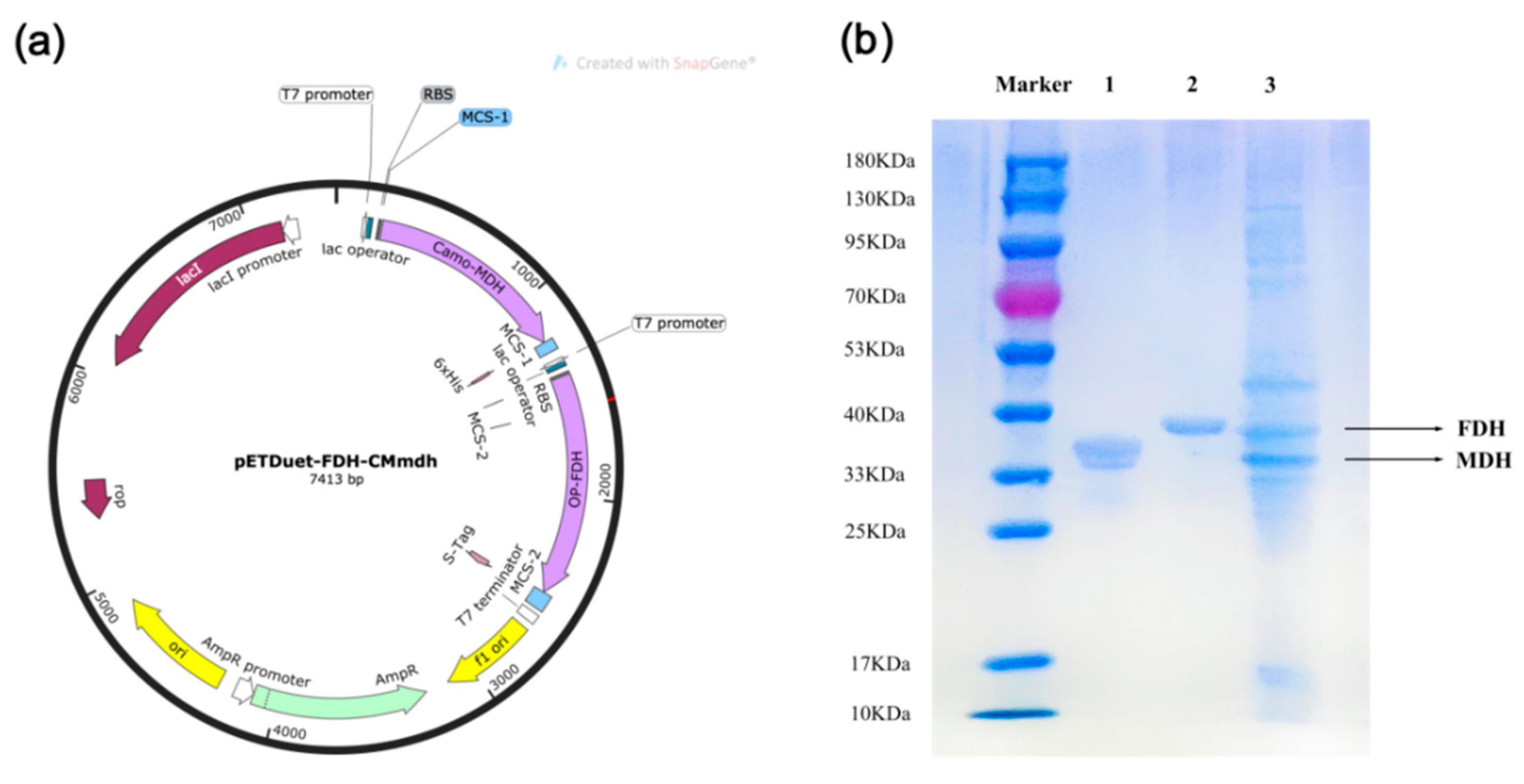
2.4. Construction of the MDH-FDH Co-Expression System
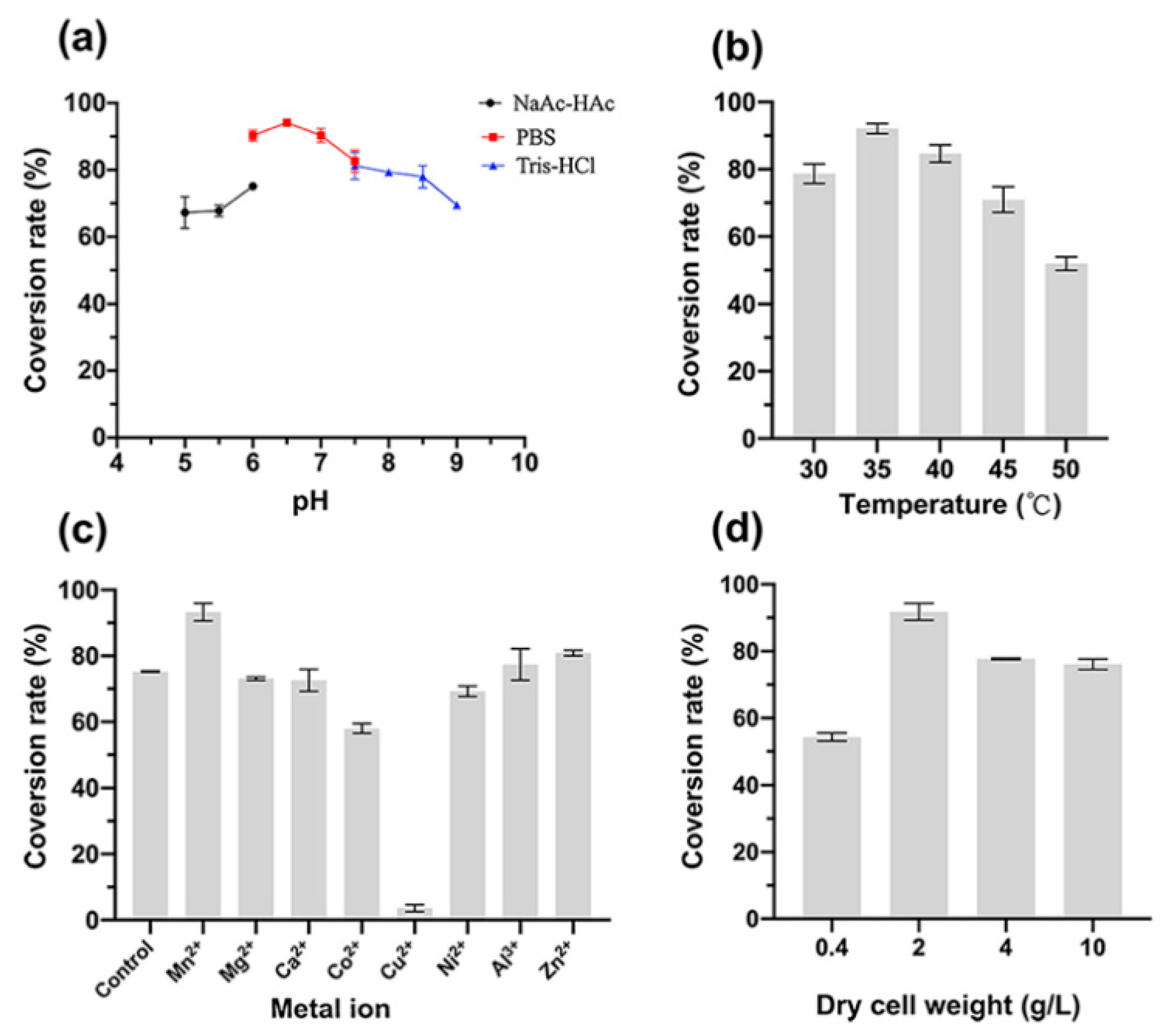
2.5. Optimization of Whole-Cell Transformation Conditions
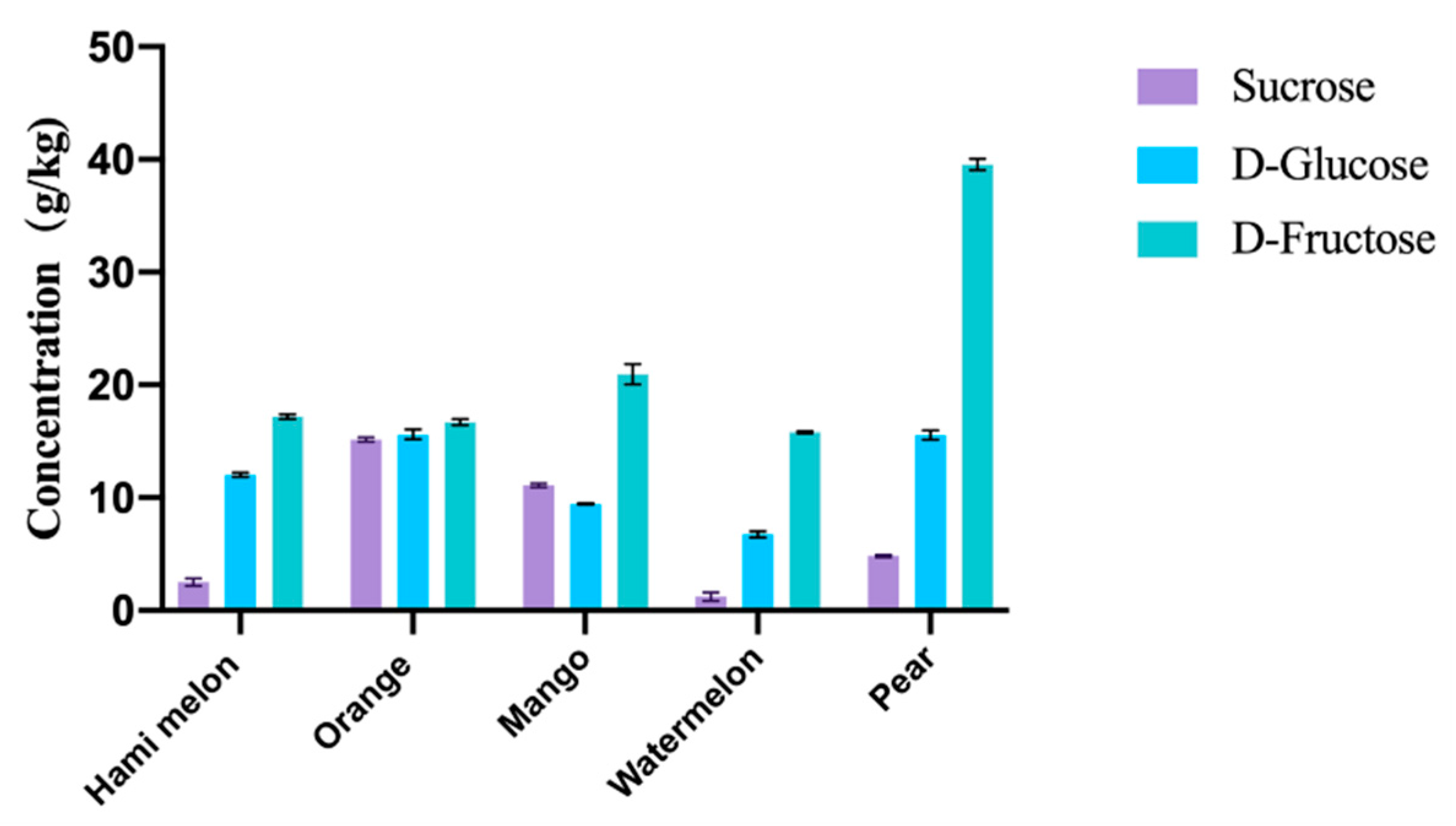
2.6. Preparation and Sugar Content Analysis of Fruit Residues
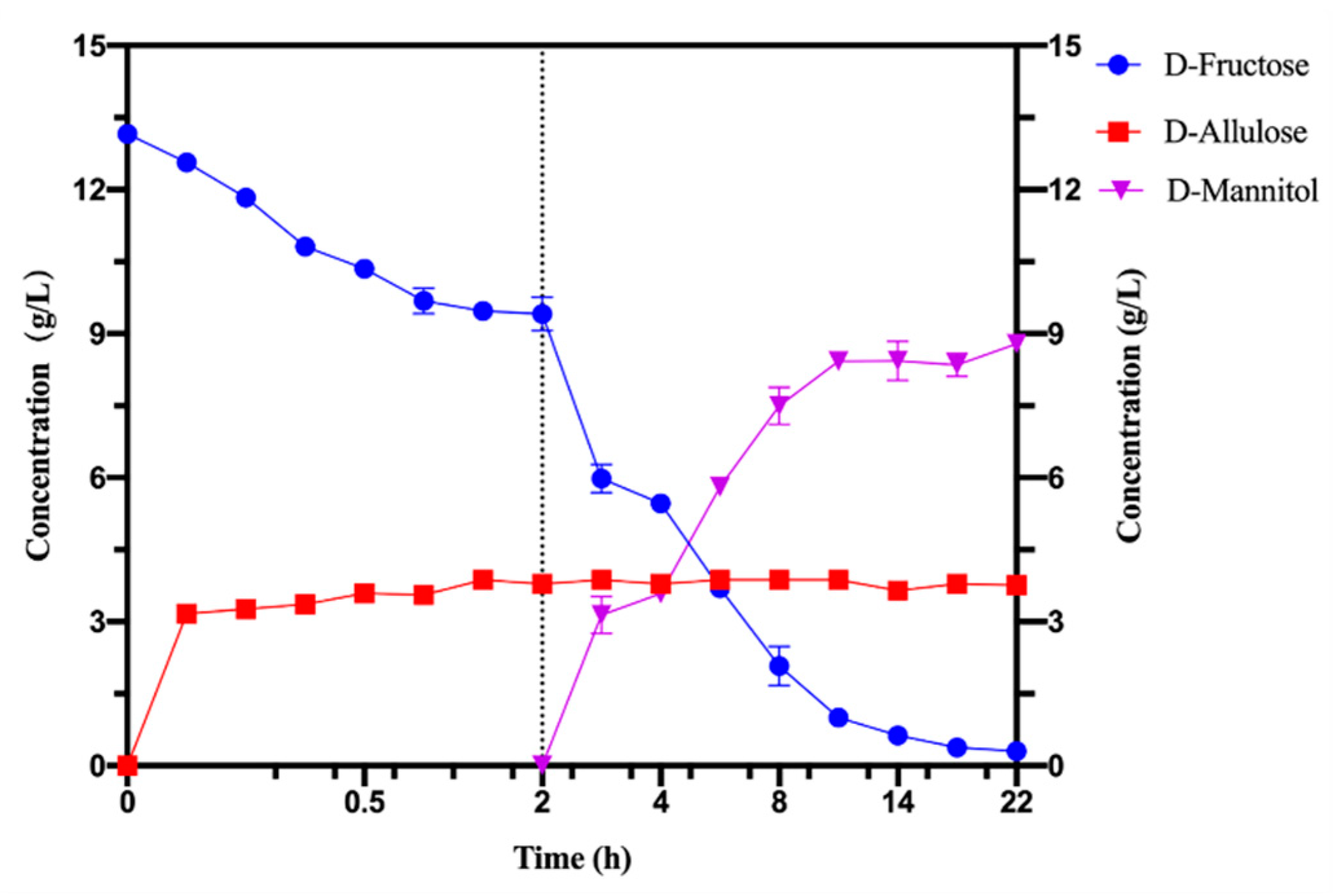
2.7. Production of D-Allulose and D-Mannitol from Fruit Residues
3. Results
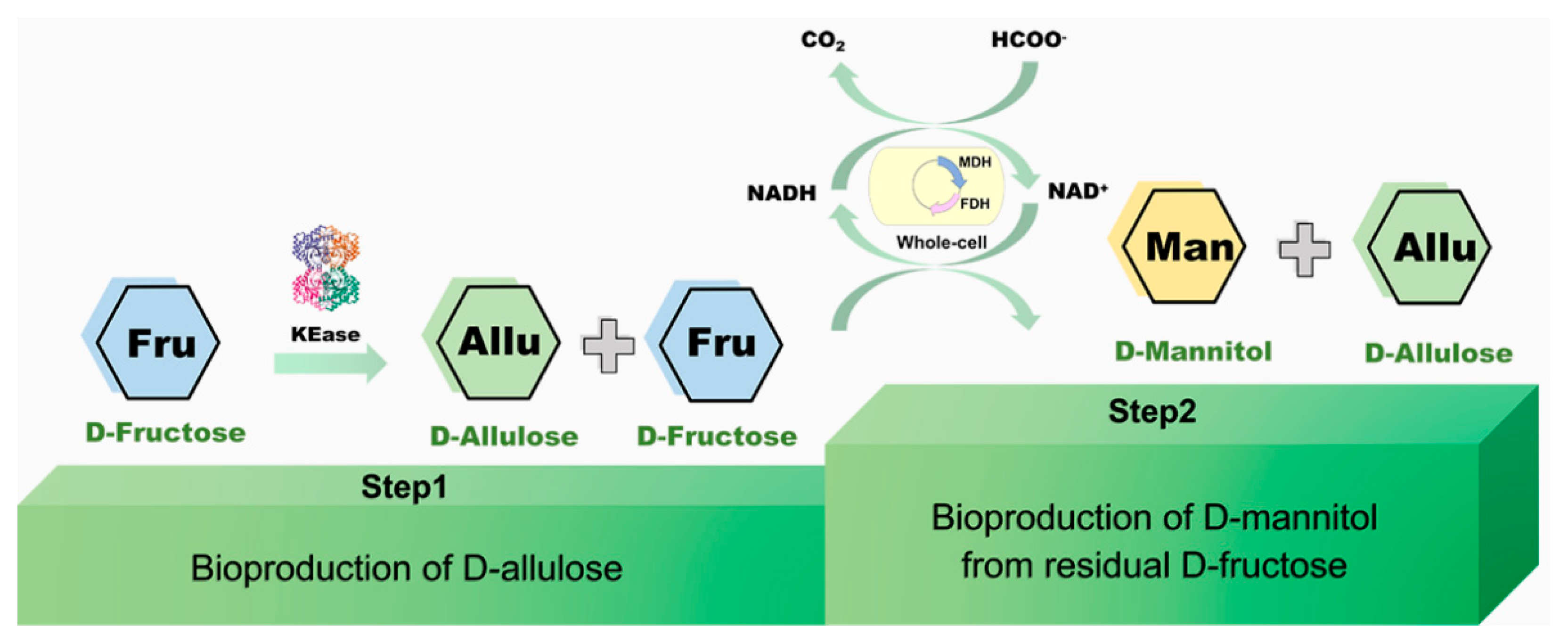
3.1. Construction of Two-Stage Strategy Biosynthesis of D-Allulose and D-Mannitol
3.2. Optimization of Whole-Cell Transformation Conditions
3.3. Sugar Content of Fruit Residues
3.4. Bioconversion of D-Allulose and D-Mannitol from Pear Peels
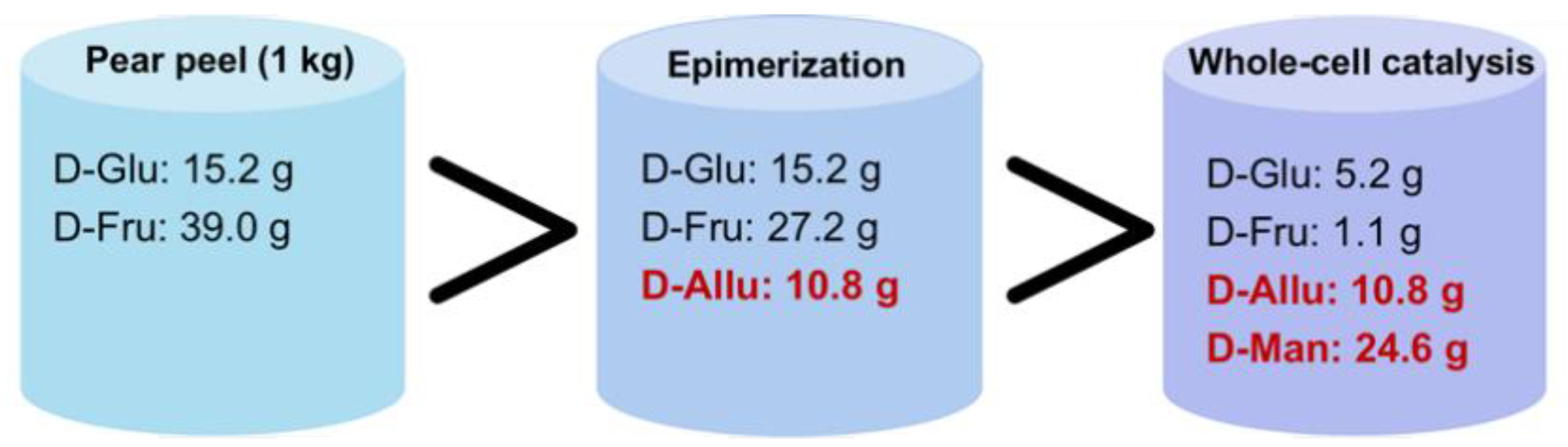
3.5. Overall Mass Balance
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khaltaev, N.; Axelrod, S. Global Trends in diabetes-related mortality with regard to lifestyle modifications, risk factors, and affordable management: A preliminary analysis. Chronic Dis. Transl. Med. 2021, 7, 182–189. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Kearsley, M.W. Sweeteners and Sugar Alternatives in Food Technology, 2nd ed.; Wiley-Blackwell: Chichester, UK; Ames, IA, USA, 2012. [Google Scholar]
- Kishida, K.; Martinez, G.; Iida, T.; Yamada, T.; Ferraris, R.P.; Toyoda, Y. D-Allulose is a substrate of glucose transporter type 5 (GLUT5) in the small intestine. Food Chem. 2019, 277, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.-M.; Hyun Lee, J.; Youl Kim, D.; Hwang, S.-H.; Hong, Y.-H.; Kim, S.-B.; Jin Lee, S.; Hye Park, C. Dietary D-psicose reduced visceral fat mass in high-fat diet-induced obese rats. J. Food Sci. 2012, 77, H53–H58. [Google Scholar] [CrossRef] [PubMed]
- Iida, T.; Yamada, T.; Hayashi, N.; Okuma, K.; Izumori, K.; Ishii, R.; Matsuo, T. Reduction of abdominal fat accumulation in rats by 8-week ingestion of a newly developed sweetener made from high fructose corn syrup. Food Chem. 2013, 138, 781–785. [Google Scholar] [CrossRef]
- Hossain, A.; Yamaguchi, F.; Matsuo, T.; Tsukamoto, I.; Toyoda, Y.; Ogawa, M.; Nagata, Y.; Tokuda, M. Rare sugar D-allulose: Potential role and therapeutic monitoring in maintaining obesity and type 2 diabetes mellitus. Pharmacol. Ther. 2015, 155, 49–59. [Google Scholar] [CrossRef]
- Hossain, A.; Tsukamoto, I.; Yamaguchi, F.; Hirata, Y.; Dong, Y.; Kamitori, K.; Sui, L.; Machiko, N.; Masaki, U.; Kazuyuki, N.; et al. Intestinal absorption, organ distribution, and urinary excretion of the rare sugar D-psicose. Drug Des. Dev. Ther. 2014, 8, 1955–1964. [Google Scholar] [CrossRef][Green Version]
- Zhang, W.; Chen, D.; Chen, J.; Xu, W.; Chen, Q.; Wu, H.; Guang, C.; Mu, W. D-Allulose, a versatile rare sugar: Recent biotechnological advances and challenges. Crit. Rev. Food Sci. Nutr. 2021, 1–19. [Google Scholar] [CrossRef]
- Niibo, M.; Kanasaki, A.; Iida, T.; Ohnishi, K.; Ozaki, T.; Akimitsu, K.; Minamino, T. D-Allulose Protects against Diabetic Nephropathy Progression in Otsuka Long-Evans Tokushima Fatty Rats with Type 2 Diabetes. PLoS ONE 2022, 17, e0263300. [Google Scholar] [CrossRef]
- Liu, B.; Gou, Y.; Tsuzuki, T.; Yamada, T.; Iida, T.; Wang, S.; Banno, R.; Toyoda, Y.; Koike, T. D-Allulose improves endurance and recovery from exhaustion in male C57BL/6J mice. Nutrients 2022, 14, 404. [Google Scholar] [CrossRef]
- Tsuzuki, T.; Suzuki, R.; Kajun, R.; Yamada, T.; Iida, T.; Liu, B.; Koike, T.; Toyoda, Y.; Negishi, T.; Yukawa, K. Combined effects of exercise training and D-allulose intake on endurance capacity in mice. Physiol. Rep. 2022, 10, e15297. [Google Scholar] [CrossRef]
- Rakhat, Y.; Kaneko, K.; Wang, L.; Han, W.; Seino, Y.; Yabe, D.; Yada, T. D-Allulose inhibits ghrelin-responsive, glucose-sensitive and neuropeptide Y neurons in the arcuate nucleus and central injection suppresses appetite-associated food intake in mice. Nutrients 2022, 14, 3117. [Google Scholar] [CrossRef] [PubMed]
- O’Charoen, S.; Hayakawa, S.; Matsumoto, Y.; Ogawa, M. Effect of D-psicose used as sucrose replacer on the characteristics of meringue. J. Food Sci. 2014, 79, E2463–E2469. [Google Scholar] [CrossRef]
- Hadipernata, M.; Ogawa, M.; Hayakawa, S. Effect of D-allulose on rheological properties of chicken breast sausage. Poult. Sci. 2016, 95, 2120–2128. [Google Scholar] [CrossRef] [PubMed]
- Izumori, K. Izumoring: A strategy for bioproduction of all hexoses. J. Biotechnol. 2006, 124, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yu, S.; Zhang, T.; Jiang, B.; Mu, W. Recent advances in D-allulose: Physiological functionalities, applications, and biological production. Trends Food Sci. Technol. 2016, 54, 127–137. [Google Scholar] [CrossRef]
- Hu, M.; Li, M.; Jiang, B.; Zhang, T. Bioproduction of D-allulose: Properties, applications, purification, and future perspectives. Compr. Rev. Food Sci. Food Saf. 2021, 20, 6012–6026. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Gu, L.; Cheng, C.; Ma, J.; Xin, F.; Liu, J.; Wu, H.; Jiang, M. Recent advances in microbial production of mannitol: Utilization of low-cost substrates, strain development and regulation strategies. World J. Microbiol. Biotechnol. 2018, 34, 41. [Google Scholar] [CrossRef]
- Rice, T.; Zannini, E.; Arendt, E.K.; Coffey, A. A review of polyols-biotechnological production, food applications, regulation, labeling and health effects. Crit. Rev. Food Sci. Nutr. 2020, 60, 2034–2051. [Google Scholar] [CrossRef]
- Dai, Y.; Meng, Q.; Mu, W.; Zhang, T. Recent advances in the applications and biotechnological production of mannitol. J. Funct. Foods 2017, 36, 404–409. [Google Scholar] [CrossRef]
- Saha, B.C.; Racine, F.M. Biotechnological production of mannitol and its applications. Appl. Microbiol. Biotechnol. 2011, 89, 879–891. [Google Scholar] [CrossRef]
- Zhang, W.; Li, H.; Zhang, T.; Jiang, B.; Zhou, L.; Mu, W. Characterization of a D-psicose 3-epimerase from Dorea Sp. CAG317 with an acidic PH optimum and a high specific activity. J. Mol. Catal. B Enzym. 2015, 120, 68–74. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Hossain, G.S.; Li, J.; Shin, H.; Chen, R.R.; Du, G.; Liu, L.; Chen, J. Bioconversion of L-glutamic acid to α-ketoglutaric acid by an immobilized whole-cell biocatalyst expressing L-amino acid deaminase from Proteus Mirabilis. J. Biotechnol. 2014, 169, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Chen, J.; Liu, X.; Guang, C.; Zhang, W.; Mu, W. Biochemical identification of a hyperthermostable L-Ribulose 3-Epimerase from Labedella endophytica and its application for D-allulose bioconversion. Int. J. Biol. Macromol. 2021, 189, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhu, Y.; Jiang, X.; Zhang, W.; Guang, C.; Mu, W. One-pot production of D-allulose from inulin by a novel identified thermostable exoinulinase from Aspergillus Piperis and Dorea sp. D-allulose 3-epimerase. Process Biochem. 2020, 99, 87–95. [Google Scholar] [CrossRef]
- Yu, S.; Zhu, L.; Zhou, C.; An, T.; Zhang, T.; Jiang, B.; Mu, W. Promising properties of a formate dehydrogenase from a methanol-assimilating yeast Ogataea Parapolymorpha DL-1 in His-tagged form. Appl. Microbiol. Biotechnol. 2014, 98, 1621–1630. [Google Scholar] [CrossRef]
- Xu, W.; Lu, F.; Wu, H.; Zhang, W.; Guang, C. Identification of a highly thermostable mannitol 2-ehydrogenase from Caldicellulosiruptor Morganii Rt8.B8 and its application for the preparation of D-mannitol. Process Biochem. 2020, 96, 194–201. [Google Scholar] [CrossRef]
- Song, Y.; Oh, C.; Bae, H.-J. Simultaneous production of bioethanol and value-added D-psicose from Jerusalem artichoke (Helianthus Tuberosus L.) tubers. Bioresour. Technol. 2017, 244, 1068–1072. [Google Scholar] [CrossRef]
- Narayan Patel, S.; Singh, V.; Sharma, M.; Sangwan, R.S.; Singhal, N.K.; Singh, S.P. Development of a thermo-stable and recyclable magnetic nanobiocatalyst for bioprocessing of fruit processing residues and D-allulose synthesis. Bioresour. Technol. 2018, 247, 633–639. [Google Scholar] [CrossRef]
- Kaup, B.; Bringer-Meyer, S.; Sahm, H. Metabolic engineering of Escherichia Coli: Construction of an efficient biocatalyst for D-mannitol formation in a whole-cell biotransformation. Appl. Microbiol. Biotechnol. 2004, 64, 333–339. [Google Scholar] [CrossRef]
- Fontes, C.P.M.L.; Honorato, T.L.; Rabelo, M.C.; Rodrigues, S. Kinetic study of mannitol production using cashew apple juice as substrate. Bioprocess Biosyst. Eng. 2009, 32, 493–499. [Google Scholar] [CrossRef]
- Patel, S.N.; Sharma, M.; Lata, K.; Singh, U.; Kumar, V.; Sangwan, R.S.; Singh, S.P. Improved operational stability of D-psicose 3-epimerase by a novel protein engineering strategy, and D-psicose production from fruit and vegetable residues. Bioresour. Technol. 2016, 216, 121–127. [Google Scholar] [CrossRef]
- Song, Y.; Nguyen, Q.A.; Wi, S.G.; Yang, J.; Bae, H.-J. Strategy for dual production of bioethanol and D-psicose as value-added products from cruciferous vegetable residue. Bioresour. Technol. 2017, 223, 34–391. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Patel, S.N.; Lata, K.; Singh, U.; Krishania, M.; Sangwan, R.S.; Singh, S.P. A novel approach of integrated bioprocessing of cane molasses for production of prebiotic and functional bioproducts. Bioresour. Technol. 2016, 219, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Men, Y.; Zhu, P.; Zhu, Y.; Zeng, Y.; Yang, J.; Sun, Y. The development of low-calorie sugar and functional jujube food using biological transformation and fermentation coupling technology. Food Sci. Nutr. 2019, 7, 1302–1310. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zhang, W.; Wu, H.; Guang, C.; Mu, W. Mannitol: Physiological functionalities, determination methods, biotechnological production, and applications. Appl. Microbiol. Biotechnol. 2020, 104, 6941–6951. [Google Scholar] [CrossRef]
- Zhu, P.; Zeng, Y.; Chen, P.; Men, Y.; Yang, J.; Yue, X.; Zhang, J.; Zhu, Y.; Sun, Y. A one-pot two-enzyme system on the production of high value-added D-allulose from Jerusalem artichoke tubers. Process Biochem. 2020, 88, 90–96. [Google Scholar] [CrossRef]
- Yang, J.; Tian, C.; Zhang, T.; Ren, C.; Zhu, Y.; Zeng, Y.; Men, Y.; Sun, Y.; Ma, Y. Development of food-grade expression system for D-allulose 3-epimerase preparation with tandem isoenzyme genes in Corynebacterium Glutamicum and its application in conversion of cane molasses to D-allulose. Biotechnol. Bioeng. 2019, 116, 745–756. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raw Material | Method | Production | Reference |
|---|---|---|---|
| Fruit and vegetable residues | N-terminal SUMO fusion of A. tumefaciens KEase as the biocatalyst. | The conversion yield of D-allulose reached 25–35% in fruit and vegetable residues. | [33] |
| Cane molasses | Dextransucrase from L. mesenteroides MTCC 10,508 to produce prebiotic oligosaccharides, SUMO fusion of A. tumefaciens KEase to synthesize D-allulose. | 124 g oligosaccharides (DP3-DP6) and 37 g D-allulose were obtained from 1 kg cane molasses. | [35] |
| Cruciferous vegetable residues | Residues was hydrolyzed into D-glucose and D-fructose by cellulase at first; then, D-glucose was fermented to bioethanol by yeast, while D-fructose was converted to D-allulose by KEase. | 49.4 g D-allulose and 166.7 g bioethanol were obtained from 1 kg dry weight cabbage (320.0 g glucose, 142.0 g fructose), 42.6 g D-allulose and 163.8 g bioethanol were obtained after separation. | [34] |
| Jerusalem artichoke tubers | Jerusalem artichoke was first hydrolyzed into D-glucose and D-fructose by exoinulinase; then, D-glucose was fermented to bioethanol by yeast, while D-fructose was converted to D-allulose by KEase. | 173.9 g D-allulose and 180.3 g bioethanol were obtained from 1 kg dry weight jerusalem artichoke (151.4 g glucose, 564.7 g fructose), 137.8 g D-allulose and 148.3 g bioethanol were obtained after separation. | [29] |
| Fruit residues | Using covalently immobilized KEase onto functionalized iron oxide magnetic nanoparticles as catalyst. | Immobilized enzyme is stable and still has 80% activity after 60 days of storage at 4 °C; and still has 90% activity after catalyzing ten cycles, and the conversion rate in kinnow and apple pomace is about 20%. | [30] |
| Inulin | A one-pot two-enzyme reaction system with Dorea sp. KEase and Aspergillus piperis exoinulinase. | 21.4 g/L D-allulose was obtained from 100 g/L inulin. | [26] |
| Jerusalem artichoke | A one-pot two-enzyme reaction system with Ruminococcus sp. KEase and Bacillus velezensis exoinulinase. | The final high-fructose syrup contained 10.4, 29.2, and 10.3 g/L D-glucose, D-fructose, and D-allulose. | [38] |
| Cane molasses | Integrated expression of 3 KEases in C. glutamicum and immobilized the cells of KEases and invertase to catalyze cane molasses for two-step reaction. | 61.2 g/L D-allulose was obtained from cane molasses (300.0 g/L sucrose, 16.5 g/L glucose, 69.0 g/L fructose). | [35] |
| Jujube | D-glucose isomerase and KEases converted D-glucose and D-fructose into D-allulose, Pediococcus pentosaceus PC-5 and Lactobacillus plantarum M were employed to increase bioactivities and flavor volatiles components. | 110 g/L D-allulose was obtained from jujube juice (352 g/L glucose, 360 g/L fructose), 100 mg/L gamma-aminobutyric acid was obtained after fermentation. | [39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Chen, J.; Xu, W.; Zhang, W.; Chen, Y.; Mu, W. Efficient Utilization of Fruit Peels for the Bioproduction of D-Allulose and D-Mannitol. Foods 2022, 11, 3613. https://doi.org/10.3390/foods11223613
Li J, Chen J, Xu W, Zhang W, Chen Y, Mu W. Efficient Utilization of Fruit Peels for the Bioproduction of D-Allulose and D-Mannitol. Foods. 2022; 11(22):3613. https://doi.org/10.3390/foods11223613
Chicago/Turabian StyleLi, Jin, Jiajun Chen, Wei Xu, Wenli Zhang, Yeming Chen, and Wanmeng Mu. 2022. "Efficient Utilization of Fruit Peels for the Bioproduction of D-Allulose and D-Mannitol" Foods 11, no. 22: 3613. https://doi.org/10.3390/foods11223613
APA StyleLi, J., Chen, J., Xu, W., Zhang, W., Chen, Y., & Mu, W. (2022). Efficient Utilization of Fruit Peels for the Bioproduction of D-Allulose and D-Mannitol. Foods, 11(22), 3613. https://doi.org/10.3390/foods11223613