
Recent Research on Fusarium Mycotoxins in Maize—A Review

 , , , , and
, , , , and
Abstract
1. Introduction
2. Classification
3. Occurrence
3.1. Free and Modified Mycotoxins
3.2. Emerging Mycotoxins
4. Toxicology
4.1. Free and Modified Mycotoxins
4.1.1. Trichothecenes and Their Modified Forms
4.1.2. ZEN and Its Modified Forms
4.1.3. Fumonisins and Their Modified Forms
4.2. Emerging Mycotoxins
5. Progress in Detoxification
5.1. Biological Detoxification of Maize
5.1.1. Trichoderma as Biological Detoxifying Organisms
5.1.2. Lactic Acid Bacteria as Biological Detoxifying Agent
5.1.3. Plant Secondary Metabolites as Biocontrol Detoxifying Agent
Essential Oils
Plant Extracts
5.2. Edible Mushrooms Source of Biological Detoxifying Agent
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Soto-Gómez, D.; Pérez-Rodríguez, P. Sustainable agriculture through perennial grains: Wheat, rice, maize, and other species. A review. Agric. Ecosyst. Environ. 2022, 325, 107747. [Google Scholar] [CrossRef]
- OECD-FAO. Agricultural Outlook 2022–2031; OECD-FAO: Rome, Italy; Paris, France, 2022. [Google Scholar] [CrossRef]
- Liliane, T.; Mutengwa, C. Factors Affecting Yield of Crops, Agronomy—Climate Change & Food Security; IntechOpen: Amanullah, Afghanistan, 2020; Available online: https://www.intechopen.com/books/agronomy-climate-change-food-security/factors-affecting-yield-of-crops (accessed on 14 September 2022).
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Foster, G.D. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Timmusk, S.; Nevo, E.; Ayele, F.; Noe, S.; Niinemets, Ü. Fighting Fusarium pathogens in the era of climate change: A Conceptual approach. Pathogens 2020, 9, 419. [Google Scholar] [CrossRef] [PubMed]
- Medina, Á.; González-Jartín, J.M.; Sainz, M.J. Impact of global warming on mycotoxins. Curr. Opin. Food Sci. 2017, 18, 76–81. [Google Scholar] [CrossRef]
- Chukwudi, U.P.; Kutu, F.R.; Mavengahama, S. Mycotoxins in maize and implications on food security: A Review. Agric. Rev. 2021, 140, 42–49. [Google Scholar] [CrossRef]
- Bouwer, G.A. Framework for effective Bt maize IRM Programs: Incorporation of lessons learned from Busseola fusca resistance development. Front. Bioeng. Biotechnol. 2020, 8, 717. [Google Scholar] [CrossRef]
- De La Campa, R.; Hooker, D.C.; Miller, J.D.; Schaafsma, A.W.; Hammond, B.G. Modeling effects of environment, insect damage, and Bt genotypes on fumonisin accumulation in maize in Argentina and Philippines. Mycopathologia 2005, 159, 539–552. [Google Scholar] [CrossRef]
- Vaughan, M.; Backhouse, D.; Del Ponte, E.M. Climate change impacts on the ecology of Fusarium graminearum species complex and susceptibility of wheat to Fusarium head blight: A review. World Mycotoxin J. 2016, 9, 685–700. [Google Scholar] [CrossRef]
- Carbas, B.; Simões, D.; Soares, A.; Freitas, A.; Ferreira, B.; Carvalho, A.R.F.; Silva, A.S.; Pinto, T.; Diogo, E.; Andrade, E.; et al. Occurrence of Fusarium spp. in maize grain harvested in Portugal and accumulation of related mycotoxins during storage. Foods 2021, 10, 375. [Google Scholar] [CrossRef]
- Commission of the European Communities. Commission Regulation EC (1881/2006). Off. J. Eur. Union 2006, 49, 5–24. [Google Scholar]
- Miedaner, T.; Boeven, A.L.G.C.; Gaikpa, D.S.; Kistner, M.B.; Grote, C.P. Genomics-assisted breeding for quantitative disease resistances in small-grain cereals and maize. Int. J. Mol. Sci. 2020, 21, 9717. [Google Scholar] [CrossRef] [PubMed]
- Stępień, Ł.; Gromadzka, K.; Chełkowski, J.; Basińska-Barczak, A.; Lalak-Kańczugowska, J. Diversity and mycotoxin production by Fusarium temperatum and Fusarium subglutinans as causal agents of pre-harvest Fusarium maize ear rot in Poland. J. Appl. Genet. 2019, 60, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Pfordt, A.; Ramos Romero, L.; Schiwek, S.; Karlovsky, P.; von Tiedemann, A. Impact of environmental conditions and agronomic practices on the prevalence of Fusarium species associated with ear-and stalk rot in maize. Pathogens 2020, 9, 236. [Google Scholar] [CrossRef] [PubMed]
- Lanubile, A.; Maschietto, V.; De Leonardis, S.; Battilani, P.; Paciolla, C.; Marocco, A. Defense responses to mycotoxin-producing fungi Fusarium proliferatum, F. subglutinans, and Aspergillus flavus in kernels of susceptible and resistant maize genotypes. Mol. Plant Microbe Interact. 2015, 28, 546–557. [Google Scholar] [CrossRef]
- Simões, D.; Diogo, E.; de Andrade, E. First report of Fusarium andiyazi presence in Portuguese maize kernels. Agriculture 2022, 12, 336. [Google Scholar] [CrossRef]
- Tillmann, M.; von Tiedemann, A.; Winter, M. Crop rotation effects on incidence and diversity of Fusarium species colonizing stem bases and grains of winter wheat. J. Plant Dis. Prot. 2017, 124, 121–130. [Google Scholar] [CrossRef]
- Galiano-Carneiro, A.L.; Kessel, B.; Presterl, T.; Gaikpa, D.S.; Kistner, M.B.; Miedaner, T. Multi-parent QTL mapping reveals stable QTL conferring resistance to Gibberella Ear Rot in maize. Euphytica 2021, 217, 2. [Google Scholar] [CrossRef]
- Rossi, V.; Scandolara, A.; Battilani, P. Effect of environmental conditions on spore production by Fusarium verticillioides, the causal agent of maize ear rot. Eur. J. Plant Pathol. 2009, 123, 159–169. [Google Scholar] [CrossRef]
- Castañares, E.; Martínez, M.; Cristos, D.; Rojas, D.; Lara, B.; Stenglein, S.; Dinolfo, M.I. Fusarium species and mycotoxin contamination in maize in Buenos Aires province, Argentina. Eur. J. Plant Pathol 2019, 155, 1265–1275. [Google Scholar] [CrossRef]
- Czembor, E.; Stępień, Ł.; Waśkiewicz, A. Effect of environmental factors on Fusarium species and associated mycotoxins in maize grain grown in Poland. PLoS ONE 2015, 10, e0133644. [Google Scholar] [CrossRef]
- Mshelia, P.L.; Selamat, J.; Samsudin, I.P.N.; Rafii, M.Y.; Mutalib, N.A.; Nordin, N.; Berthiller, F. Effect of temperature, water activity and carbon dioxide on fungal growth and mycotoxin production of acclimatised isolates of Fusarium verticillioides and F. graminearum. Toxins 2020, 12, 478. [Google Scholar] [CrossRef] [PubMed]
- Uwineza, P.A.; Urbaniak, M.; Bryła, M.; Stepien, Ł.; Modrzewska, M.; Waśkiewicz, A. In vitro effects of lemon balm extracts in reducing the growth and mycotoxins biosynthesis of Fusarium culmorum and F. proliferatum. Toxins 2022, 14, 355. [Google Scholar] [CrossRef] [PubMed]
- Waśkiewicz, A.; Beszterda, M.; Golinski, P. Occurrence of fumonisins in food—An interdisciplinary approach to the problem. Food Control 2012, 26, 491–499. [Google Scholar] [CrossRef]
- Ekwomadu, T.I.; Dada, T.A.; Nleya, N.; Gopane, R.; Sulyok, M.; Mwanza, M. Variation of Fusarium free, masked, and emerging mycotoxin metabolites in maize from agriculture regions of South Africa. Toxins 2020, 12, 149. [Google Scholar] [CrossRef]
- Zhang, J.; Abdelraheem, A.; Zhu, Y.; Elkins-Arce, H.; Dever, J.; Whitelock, D.; Wheeler, T.A. Studies of evaluation methods for resistance to Fusarium wilt race 4 (Fusarium oxysporum f. sp. vasinfectum) in cotton: Effects of cultivar, planting date, and inoculum density on disease progression. Front. Plant Sci. 2022, 13, 900131. [Google Scholar] [CrossRef]
- Shank, R.A.; Foroud, N.A.; Hazendonk, P.; Eudes, F.; Blackwell, B.A. Current and future experimental strategies for structural analysis of trichothecene mycotoxins—A prospectus. Toxins 2011, 3, 1518–1553. [Google Scholar] [CrossRef]
- Piec, J.; Pallez, M.; Beyer, M.; Vogelgsang, S.; Hoffmann, L.; Pasquali, M. The Luxembourg database of trichothecene type B F. graminearum and F. culmorum producers. Bioinformation 2016, 12, 1–3. [Google Scholar] [CrossRef]
- Ropejko, K.; Twarużek, M. Zearalenone and its metabolites—General overview, occurrence, and toxicity. Toxins 2021, 13, 35. [Google Scholar] [CrossRef]
- Hao, K.; Suryoprabowo, S.; Song, S.; Liu, L.; Kuang, H. Rapid detection of zearalenone and its metabolite in corn flour with the immunochromatographic test strip. Food Agric. Immunol. 2018, 29, 498–510. [Google Scholar] [CrossRef]
- Zhou, D.; Wang, X.; Chen, G.; Sun, S.; Yang, Y.; Zhu, Z.; Duan, C. The major Fusarium species causing maize ear and kernel rot and their toxigenicity in Chongqing, China. Toxins 2018, 10, 90. [Google Scholar] [CrossRef]
- Logrieco, A.; Mule, G.; Moretti, A.; Bottalico, A. Toxigenic Fusarium Species and Mycotoxins Associated with Maize Ear Rot in Europe. Mycotoxins in Plant Disease; Springer: Dordrecht, The Netherlands, 2002; pp. 597–609. [Google Scholar]
- Jestoi, M. Emerging Fusarium—Mycotoxins fusaproliferin, beauvericin, enniatins, and moniliformin—A review. Crit. Rev. Food Sci. Nutr. 2008, 48, 21–49. [Google Scholar] [CrossRef] [PubMed]
- Marin, S.; Magan, N.; Ramos, A.J.; Sanchis, V. Fumonisin-producing strains of Fusarium: A review of their ecophysiology. J. Food Prot. 2004, 67, 1792–1805. [Google Scholar] [CrossRef] [PubMed]
- Munkvold, G.P.; Arias, S.; Taschl, I.; Gruber-Dorninger, C. Mycotoxins in corn: Occurrence, impacts, and management. AACC Int. Press 2019, 235–287. [Google Scholar] [CrossRef]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-mycotoxin screening reveals the occurrence of 139 different secondary metabolites in feed and feed ingredients. Toxins 2013, 5, 504–523. [Google Scholar] [CrossRef] [PubMed]
- Zinedine, A.; Soriano, J.M.; Molto, J.C.; Manes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Pronk, M.E.J.; Schothorst, R.C.; van Egmond, H.P. Toxicology and Occurrence of Nivalenol, Fusarenon X, Diacetoxyscirpenol, Neosolaniol and 3- and 15-Acetylnivalenol: A Review of Six Trichothecenes; RIVM Report; RIVM: Utrecht, The Netherlands, 2002; p. 75. [Google Scholar]
- Rychlik, M.; Humpf, H.U.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Lorenz, N. Proposal of a comprehensive definition of modified and other forms of mycotoxins including “masked” mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [CrossRef] [PubMed]
- FDA. Guidance for Industry. Advisory Levels for Deoxynivalenol (DON) in Finished Wheat Products for Human Consumption and Grains and Grain By-Products Used for Animal Feed (June 29, 2010); U.S. Food and Drug Administration: Silver Spring, MD, USA, 2010. Available online: https://www.fda.gov/Food/GuidanceRegulation/GuidanceDocumentsRegulatoryInformation/ChemicalContaminantsMetalsNaturalToxinsPesticides/default.htm (accessed on 29 August 2018).
- CFIA. RG-8 Regulatory Guidance. Contaminants in Feed (for- Merly RG-1, Chapter 7), Section 1: Mycotoxins in Livestock Feed; Canadian Food Inspection Agency (CFIA): Alberta, BC, USA, 2017; Available online: http://www.inspection.gc.ca/animals/feeds/regulatory-guidance/rg-8/eng/1347383943203/1347384015909 (accessed on 1 September 2022).
- FSANZ. Australia New Zealand Food Standards Code, 2017. Schedule 19, Maximum Levels of Contaminants and Natural Toxicants. Prepared by Food Standards Australia New Zealand (FSANZ) on 13 April 2017; Australian Government, Federal Register of Legislation: Canberra, ACT, Canada, 2017. Available online: https://www.legislation.gov.au/Details/F2017C00333 (accessed on 1 September 2022).
- FAMIC. Regulation Value of Pesticides, Heavy Metals and Myco-Toxins (Administrative Guideline) (Revised December 18, 2015); Food and Agricultural Materials Inspection Centre (FAMIC): Tokyo, Japan, 2015; Available online: http://www.famic.go.jp/ffis/feed/r_safety/r_feeds_%20safety22.html (accessed on 1 September 2022).
- Asam, S.; Habler, K.; Rychlik, M. Fusarium mycotoxins in Food. In Chemical Contaminants and Residues in Food; Woodhead Publishing: Amsterdam, The Netherlands, 2017. [Google Scholar]
- De Boevre, M.; Di Mavungu, J.D.; Maene, P.; Audenaert, K.; Deforce, D.; Haesaert, G.; De Saeger, S. Development and validation of an LC-MS/MS method for the simultaneous determination of deoxynivalenol, zearalenone, T-2-toxin and some masked metabolites in different cereals and cereal-derived food. Food Addit. Contam. 2012, 29, 819–835. [Google Scholar] [CrossRef]
- Ksieniewicz-Woźniak, E.; Bryła, M.; Michałowska, D.; Waśkiewicz, A.; Yoshinari, T. Transformation of selected Fusarium toxins and their masked forms during malting of various cultivars of wheat. Toxins 2021, 13, 866. [Google Scholar] [CrossRef]
- Bryła, M.; Roszko, M.; Szymczyk, K.; Jędrzejczak, R.; Obiedziński, M.W. Fumonisins and their masked forms in maize products. Food Control 2016, 59, 619–627. [Google Scholar] [CrossRef]
- Dall’Asta, C.; Galaverna, G.; Mangia, M.; Sforza, S.; Dossena, A.; Marchelli, R. Free and bound fumonisins in gluten-free food products. Mol. Nutr. Food Res. 2009, 53, 492–499. [Google Scholar] [CrossRef]
- Tan, H.; Hongyuan, Z.; Ting, G.; Jiaxin, L.; Chi, Z.; Shuo, W.; Yuhao, Z.; Liang, M. Zein structure and its hidden zearalenone: Effect of zein extraction methods. Food Chem. 2022, 374, 131563. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Li, Y.; Zhou, H.; Guo, T.; Zhou, Y.; Zhang, Y.; Ma, L. Temperature and pH levels: Key factors effecting hidden/free zearalenone during maize processing. Int. Food Res. J. 2022, 160, 111721. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Zhou, H.; Guo, T.; Zhang, Y.; Li, J.; Zhang, C.; Ma, L. Effect of temperature and pH on the conversion between free and hidden zearalenone in zein. Food Chem. 2021, 360, 130001. [Google Scholar] [CrossRef] [PubMed]
- Kovalsky, P.; Kos, G.; Nahrer, K.; Schwab, C.; Jenkins, T.; Schatzmayr, G.; Sulyok, M.; Krska, R. Co-occurrence of regulated, masked and emerging mycotoxins and secondary metabolites in finished feed and maize-an extensive survey. Toxins 2016, 8, 363. [Google Scholar] [CrossRef]
- Jajić, I.; Dudaš, T.; Krstović, S.; Krska, R.; Sulyok, M.; Bagi, F.; Savić, Z.; Guljaš, D.; Stankov, A. Emerging Fusarium mycotoxins fusaproliferin, beauvericin, enniatins, and moniliformin in Serbian maize. Toxins 2019, 11, 357. [Google Scholar] [CrossRef]
- Cole, R.J.; Kirksey, J.W.; Cutler, H.G.; Doupnik, B.L.; Peckham, J.C. Toxin from Fusarium moniliforme: Effects on plants and animals. Science 1973, 179, 1324–1326. [Google Scholar] [CrossRef]
- Orlando, B.; Grignon, G.; Vitry, C.; Kashefifard, K.; Valade, R. Fusarium species and enniatin mycotoxins in wheat, durum wheat, triticale and barley harvested in France. Mycotoxin Res. 2019, 35, 369–380. [Google Scholar] [CrossRef]
- Abdallah, M.F.; Girgin, G.; Baydar, T.; Krska, R.; Sulyok, M. Occurrence of multiple mycotoxins and other fungal metabolites in animal feed and maize samples from Egypt using LC-MS/MS. J. Sci. Food Agric. 2017, 97, 4419–4428. [Google Scholar] [CrossRef]
- Hu, L.; Liu, H.; Yang, J.; Wang, C.; Wang, Y.; Yang, Y.; Chen, X. Free and hidden fumonisins in raw maize and maize-based products from China. Food Addit. Contam. 2019, 12, 90–96. [Google Scholar] [CrossRef]
- Topi, D.; Babič, J.; Pavšič-Vrtač, K.; Tavčar-Kalcher, G.; Jakovac-Strajn, B. Incidence of Fusarium mycotoxins in wheat and maize from Albania. Molecules 2021, 26, 172. [Google Scholar] [CrossRef]
- Fusilier, K.; Chilvers, M.I.; Limay-Rios, V.; Singh, M.P. Mycotoxin co-occurrence in Michigan harvested maize grain. Toxins 2022, 14, 431. [Google Scholar] [CrossRef] [PubMed]
- Tarazona, A.; Gómez, J.V.; Jiménez, M.; Mateo, F.; Romera, D.; Mateo, E.M. Study on mycotoxin contamination of maize kernels in Spain. Food Control 2020, 118, 107370. [Google Scholar] [CrossRef]
- Hanvi, D.M.; Lawson-Evi, P.; De Boevre, M.; Goto, C.E.; De Saeger, S.; Eklu-Gadegbeku, K. Natural occurrence of mycotoxins in maize and sorghum in Togo. Mycotoxin Res. 2019, 35, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Reisinger, N.; Schürer-Waldheim, S.; Mayer, E.; Debevere, S.; Antonissen, G.; Sulyok, M.; Nagl, V. Mycotoxin Occurrence in Maize Silage-A Neglected Risk for Bovine Gut Health? Toxins 2019, 11, 577. [Google Scholar] [CrossRef] [PubMed]
- Anjorin, S.T.; Fapohunda, S.; Sulyok, M.; Krska, R. Natural co-occurrence of emerging and minor mycotoxins on maize grains from Abuja, Nigeria. Ann. Agric. Environ. Sci. 2016, 1, 21–29. [Google Scholar]
- Oliveira, M.S.; Rocha, A.; Sulyok, M.; Krska, R.; Mallmann, C.A. Natural mycotoxin contamination of maize (Zea mays L.) in the South region of Brazil. Food Control 2017, 73, 127–132. [Google Scholar] [CrossRef]
- Birr, T.; Jensen, T.; Preußke, N.; Sönnichsen, F.D.; De Boevre, M.; De Saeger, S.; Hasler, M.; Verreet, J.A.; Klink, H. Occurrence of Fusarium mycotoxins and their modified forms in forage maize cultivars. Toxins 2021, 13, 110. [Google Scholar] [CrossRef]
- Worku, A.F.; Merkuz, A.; Kalsa, K.K.; Subramanyam, B.; Habtu, N.G. Occurrence of mycotoxins in stored maize in Ethiopia. Ethiop. J. Agric. Sci. 2019, 29, 31–43. [Google Scholar]
- Bertuzzi, T.; Giorni, P.; Rastelli, S.; Vaccino, P.; Lanzanova, C.; Locatelli, S. Co-occurrence of moniliformin and regulated Fusarium toxins in maize and wheat grown in Italy. Molecules 2020, 25, 2440. [Google Scholar] [CrossRef]
- Zhao, Z.; Rao, Q.; Song, S.; Liu, N.; Han, Z.; Hou, J.; Wu, A. Simultaneous determination of major type B trichothecenes and deoxynivalenol-3-glucoside in animal feed and raw materials using improved DSPE combined with LC-MS/MS. J. Chromatogr. B 2014, 963, 75–82. [Google Scholar] [CrossRef]
- Vandicke, J.; De Visschere, K.; Croubels, S.; De Saeger, S.; Audenaert, K.; Haesaert, G. Mycotoxins in Flanders’ Fields: Occurrence and correlations with Fusarium species in whole-plant harvested maize. Microorganisms 2019, 7, 571. [Google Scholar] [CrossRef] [PubMed]
- Bryła, M.; Waśkiewicz, A.; Szymczyk, K.; Jędrzejczak, R. Effects of pH and temperature on the stability of fumonisins in maize products. Toxins 2017, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Zhou, H.; Guo, T.; Zhang, Y.; Ma, L. Zein-bound zearalenone: A hidden mycotoxin found in maize and maize-products. Food Control 2021, 124, 107903. [Google Scholar] [CrossRef]
- Wang, W.; Zhu, Y.; Abraham, N.; Li, X.Z.; Kimber, M.; Zhou, T. The ribosome-binding mode of trichothecene mycotoxins rationalizes their structure—Activity relationships. Int. J. Mol. Sci. 2021, 22, 1604. [Google Scholar] [CrossRef]
- Aupanun, S.; Poapolathep, S.; Giorgi, M.; Imsilp, K.; Poapolathep, A. An overview of the toxicology and toxicokinetics of fusarenon-X, a type B trichothecene mycotoxin. J. Vet. Med. Sci. 2017, 79, 6–13. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Risks to human and animal health related to the presence of deoxynivalenol and its acetylated and modified forms in food and feed. EFSA J. 2017, 15, 4718. [Google Scholar] [CrossRef]
- Broekaert, N.; Devreese, M.; Demeyere, K.; Berthiller, F.; Michlmayr, H.; Varga, E.; Adam, G.; Meyer, E.; Croubels, S. Comparative in vitro cytotoxicity of modified deoxynivalenol on porcine intestinal epithelial cells. Food Chem. Toxicol. 2016, 95, 103–109. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, S.; Tan, Y.; Liu, N.; Wu, A. Individual and combined cytotoxic effects of co-occurring deoxynivalenol family mycotoxins on human gastric epithelial cells. Toxins 2017, 9, 96. [Google Scholar] [CrossRef]
- He, Y.; Yin, X.; Dong, J.; Yang, Q.; Wu, Y.; Gong, Z. Transcriptome analysis of Caco-2 cells upon the exposure of mycotoxin deoxynivalenol and its acetylated derivatives. Toxins 2021, 13, 167. [Google Scholar] [CrossRef]
- Springler, A.; Hessenberger, S.; Reisinger, N.; Kern, C.; Nagl, V.; Schatzmayr, G.; Mayer, E. Deoxynivalenol and its metabolite deepoxy-deoxynivalenol: Multi-parameter analysis for the evaluation of cytotoxicity and cellular effects. Mycotoxin Res. 2017, 33, 25–37. [Google Scholar] [CrossRef]
- Khaneghah, A.M.; Martins, L.M.; von Hertwig, A.M.; Bertoldo, R.; Sant’Ana, A.S. Deoxynivalenol and its masked forms: Characteristics, incidence, control and fate during wheat and wheat based products processing—A review. Trends Food Sci. Technol. 2018, 71, 13–24. [Google Scholar] [CrossRef]
- Payros, D.; Alassane-Kpembi, I.; Pierron, A.; Loiseau, N.; Pinton, P.; Oswald, I.P. Toxicology of deoxynivalenol and its acetylated and modified forms. Arch. Toxicol. 2016, 90, 2931–2957. [Google Scholar] [CrossRef] [PubMed]
- Juan-García, A.; Juan, C.; Tolosa, J.; Ruiz, M.J. Effects of deoxynivalenol, 3-acetyl-deoxynivalenol and 15-acetyl-deoxynivalenol on parameters associated with oxidative stress in HepG2 cells. Mycotoxin Res. 2019, 35, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Fan, M.; Chu, X.; Zhang, Y.; Rahman, S.U.; Jiang, Y.; Chen, X.; Zhu, D.; Feng, S.; Li, Y. Deoxynivalenol induces toxicity and apoptosis in piglet hippocampal nerve cells via the MAPK signaling pathway. Toxicon 2018, 155, 1–8. [Google Scholar] [CrossRef]
- Nagashima, H. Deoxynivalenol and nivalenol toxicities in cultured cells: A review of comparative studies. Food Saf. 2018, 6, 51–57. [Google Scholar] [CrossRef][Green Version]
- Nagashima, H.; Nakagawa, H.; Iwashita, K. Cytotoxic effects of nivalenol on HL60 cells. Mycotoxins 2006, 56, 65–70. [Google Scholar] [CrossRef]
- Nagashima, H.; Kushiro, M.; Nakagawa, H.; Iwashita, K. Comparison of antiproliferative effects of trichothecene mycotoxins, nivalenol and deoxynivalenol, in cultured cells. Rep. Natl. Food Res. Inst. 2012, 76, 29–32. [Google Scholar] [CrossRef]
- Thuvander, A.; Wikman, C.; Gadhasson, I. In vitro exposure of human lymphocytes to trichothecenes: Individual variation in sensitivity and effects of combined exposure on lymphocyte function. Food Chem. Toxicol. 1999, 37, 639–648. [Google Scholar] [CrossRef]
- Minervini, F.; Fornelli, F.; Flynn, K.M. Toxicity and apoptosis induced by the mycotoxins nivalenol, deoxynivalenol and fumonisin B1 in a human erythroleukemia cell line. Toxicol. Vitr. 2004, 18, 21–28. [Google Scholar] [CrossRef]
- Aupanun, S.; Phuektes, P.; Poapolathep, S.; Alassane-Kpembi, I.; Oswald, I.P.; Poapolathep, A. Individual and combined cytotoxicity of major trichothecenes type B, deoxynivalenol, nivalenol, and fusarenon-X on Jurkat human T cells. Toxicon 2019, 160, 29–37. [Google Scholar] [CrossRef]
- Aupanun, S.; Poapolathep, S.; Phuektes, P.; Giorgi, M.; Zhang, M.; Oswald, I.P.; Poapolathep, A. Individual and combined mycotoxins deoxynivalenol, nivalenol, and fusarenon-X induced apoptosis in lymphoid tissues of mice after oral exposure. Toxicon 2019, 165, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Schwartz-Zimmermann, H.E.; Binder, S.B.; Hametner, C.; Miró-Abella, E.; Schwarz, C.; Michlmayr, H.; Berthiller, F. Metabolism of nivalenol and nivalenol-3-glucoside in rats. Toxicol. Lett. 2019, 306, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Janik, E.; Niemcewicz, M.; Podogrocki, M.; Ceremuga, M.; Stela, M.; Bijak, M. T-2 Toxin—The Most Toxic Trichothecene Mycotoxin: Metabolism, Toxicity, and Decontamination Strategies. Molecules 2021, 26, 6868. [Google Scholar] [CrossRef]
- Ji, F.; He, D.; Olaniran, A.O.; Mokoena, M.P.; Xu, J.; Shi, J. Occurrence, toxicity, production and detection of Fusarium mycotoxin: A review. Food Prod. Process. Nutr. 2019, 1, 6. [Google Scholar] [CrossRef]
- Taroncher, M.; Rodríguez-Carrasco, Y.; Ruiz, M.J. Interactions between T-2 toxin and its metabolites in HepG2 cells and in silico approach. Food Chem. Toxicol. 2021, 148, 111942. [Google Scholar] [CrossRef] [PubMed]
- Arcella, D.; Gergelova, P.; Innocenti, M.L.; Steinkellner, H. Human and animal dietary exposure to T-2 and HT-2. EFSA J. 2017, 15, 4972. [Google Scholar] [CrossRef]
- Taroncher, M.; Rodríguez-Carrasco, Y.; Ruiz, M.-J. T-2 toxin and its metabolites: Characterization, cytotoxic mechanisms and adaptive cellular response in human hepatocarcinoma (HepG2) cells. Food Chem. Toxicol. 2020, 145, 111654. [Google Scholar] [CrossRef] [PubMed]
- Ling, A.; Sun, L.W.; Guo, W.B.; Sun, S.Y.; Yang, J.H.; Zhao, Z.H. Individual and combined cytotoxic effects of T-2 toxin and its four metabolites on porcine Leydig cells. Food Chem. Toxicol. 2020, 139, 111277. [Google Scholar] [CrossRef]
- Tran, V.N.; Viktorova, J.; Augustynkova, K.; Jelenova, N.; Dobiasova, S.; Rehorova, K.; Fenclova, M.; Stranska-Zachariasova, M.; Vitek, L.; Hajslova, J. In silico and in vitro studies of mycotoxins and their cocktails; Their toxicity and its mitigation by silibinin pre-treatment. Toxins 2020, 12, 148. [Google Scholar] [CrossRef]
- Marin, D.E.; Taranu, I.; Burlacu, R.; Tudor, D.S. Effects of zearalenone and its derivatives on the innate immune response of swine. Toxicon 2010, 56, 956–963. [Google Scholar] [CrossRef]
- Zhang, J.; Sheng, K.; Wu, W.; Zhang, H. Anorectic responses to T-2 toxin, HT-2 toxin, diacetoxyscirpenol and neosolaniol correspond to plasma elevations of neurotransmitters 5-hydroxytryptamine and substance P. Ecotoxicol. Environ. Saf. 2018, 161, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.L.; Sun, X.F.; Feng, Y.Z.; Li, B.; Li, Y.P.; Yang, F.; Nyachoti, C.M.; Shen, W.; Sun, S.D.; Li, L. Zearalenone exposure impairs ovarian primordial follicle formation via down-regulation of Lhx8 expression in vitro. Toxicol. Appl. Pharmacol. 2017, 317, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Kuca, K.; Nepovimova, E.; Wu, W. Type A trichothecene diacetoxyscirpenol-induced emesis corresponds to secretion of peptide YY and serotonin in mink. Toxins 2020, 12, 419. [Google Scholar] [CrossRef] [PubMed]
- Broekaert, N.; Devreese, M.; De Boevre, M.; De Saeger, S.; Croubels, S. T-2 toxin-3α-glucoside in broiler chickens: Toxicokinetics, absolute oral bioavailability, and in vivo hydrolysis. J. Agric. Food Chem. 2017, 65, 4797–4803. [Google Scholar] [CrossRef]
- Tatay, E.; Espín, S.; García-Fernández, A.J.; Ruiz, M.J. Oxidative damage and disturbance of antioxidant capacity by zearalenone and its metabolites in human cells. Toxicol. Vitr. 2017, 45, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Salah-Abbès, J.B.; Belgacem, H.; Ezzdini, K.; Abdel-Wahhab, M.A.; Abbès, S. Zearalenone nephrotoxicity: DNA fragmentation, apoptotic gene expression and oxidative stress protected by Lactobacillus plantarum MON03. Toxicon 2020, 175, 28–35. [Google Scholar] [CrossRef]
- Pierzgalski, A.; Bryła, M.; Kanabus, J.; Modrzewska, M.; Podolska, G. Updated review of the toxicity of selected Fusarium toxins and their modified forms. Toxins 2021, 13, 768. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Appropriateness to set a group health-based guidance value for zearalenone and its modified forms. EFSA J. 2016, 14, e04425. [Google Scholar] [CrossRef]
- Karaman, E.F.; Zeybel, M.; Ozden, S. Evaluation of the epigenetic alterations and gene expression levels of HepG2 cells exposed to zearalenone and α-zearalenol. Toxicol. Lett. 2020, 326, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Taranu, I.; Burlacu, R.; Manda, G.; Motiu, M.; Neagoe, I.; Dragomir, C.; Stancu, M.; Calin, L. Effects of zearalenone and its derivatives on porcine immune response. Toxicol. Vitr. 2011, 25, 1981–1988. [Google Scholar] [CrossRef]
- Dellafiora, L.; Perotti, A.; Galaverna, G.; Buschini, A.; Dall’Asta, C. On the masked mycotoxin zearalenone-14-glucoside. Does the mask truly hide? Toxicon 2016, 111, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Cirlini, M.; Barilli, A.; Galaverna, G.; Michlmayr, H.; Adam, G.; Berthiller, F.; Dall’Asta, C. Study on the uptake and deglycosylation of the masked forms of zearalenone in human intestinal Caco-2 cells. Food Chem. Toxicol. 2016, 98, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Pistol, G.C.; Bulgaru, C.V.; Taranu, I. Cytotoxic and inflammatory effects of individual and combined exposure of HepG2 cells to zearalenone and its metabolites. Naunyn Schmiedebergs Arch. Pharmacol. 2019, 392, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Luongo, D.; Severino, L.; Bergamo, P.; De Luna, R.; Lucisano, A.; Rossi, M. Interactive effects of fumonisin B1 and α-zearalenol on proliferation and cytokine expression in Jurkat T cells. Toxicol. Vitr. 2006, 20, 1403–1410. [Google Scholar] [CrossRef]
- Ben Salem, I.; Boussabbeh, M.; Da Silva, J.P.; Guilbert, A.; Bacha, H.; Abid-Essefi, S.; Lemaire, C. SIRT1 protects cardiac cells against apoptosis induced by zearalenone or its metabolites α- and β-zearalenol through an autophagy-dependent pathway. Toxicol. Appl. Pharmacol. 2017, 314, 82–90. [Google Scholar] [CrossRef]
- Steinkellner, H.; Binaglia, M.; Dall’Asta, C.; Gutleb, A.C.; Metzler, M.; Oswald, I.P.; Parent-Massin, D.; Alexander, J. Combined hazard assessment of mycotoxins and their modified forms applying relative potency factors: Zearalenone and t2/ht2 toxin. Food Chem. Toxicol. 2019, 131, 110599. [Google Scholar] [CrossRef]
- Liu, L.; Xie, M.; Wei, D. Biological detoxification of mycotoxins: Current status and future advances. Int. J. Mol. Sci. 2022, 23, 1064. [Google Scholar] [CrossRef]
- Bittner, G.D.; Denison, M.S.; Yang, C.Z.; Stoner, M.A.; He, G. Chemicals having estrogenic activity can be released from some bisphenol a-free, hard and clear, thermoplastic resins. Environ. Health 2014, 13, 103. [Google Scholar] [CrossRef]
- Tatay, E.; Espín, S.; García-Fernández, A.-J.; Ruiz, M.-J. Estrogenic activity of zearalenone, α-zearalenol and β-zearalenol assessed using the E-screen assay in MCF-7 cells. Toxicol. Mech. Methods 2018, 28, 239–242. [Google Scholar] [CrossRef]
- Eze, U.A.; Huntriss, J.; Routledge, M.N.; Gong, Y.Y.; Connolly, L. The effect of individual and mixtures of mycotoxins and persistent organochloride pesticides on oestrogen receptor transcriptional activation using in vitro reporter gene assays. Food Chem. Toxicol. 2019, 130, 68–78. [Google Scholar] [CrossRef]
- Dellafiora, L.; Ruotolo, R.; Perotti, A.; Cirlini, M.; Galaverna, G.; Cozzini, P.; Buschini, A.; Dall’Asta, C. Molecular insights on xenoestrogenic potential of zearalenone-14-glucoside through a mixed in vitro/in silico approach. Food Chem. Toxicol. 2017, 108, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Foryst-Ludwig, A.; Clemenz, M.; Hohmann, S.; Hartge, M.; Sprang, C.; Frost, N.; Krikov, M.; Bhanot, S.; Barros, R.; Morani, A.; et al. Metabolic actions of estrogen receptor beta (ERβ) are mediated by a negative cross-talk with PPARγ. PLoS Genet. 2008, 4, e1000108. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Wang, L.; Ji, H.; Fang, Y.; Lei, P.; Zhang, X.; Jin, L.; Sun, D.; Dong, H. Toxic mechanism and biological detoxification of fumonisins. Toxins 2022, 14, 182. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.W. Fumonisins. In Veterinary Toxicology; Gupta, R.C., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2018; pp. 1003–1018. [Google Scholar]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Knutsen, H.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B. Appropriateness to set a group health-based guidance value for fumonisins and their modified forms. EFSA J. 2018, 16, 5172. [Google Scholar]
- Wangia-Dixon, R.N.; Nishimwe, K. Molecular toxicology and carcinogenesis of fumonisins: A review. J. Environ. Sci. Health Part C 2021, 39, 44–67. [Google Scholar] [CrossRef]
- Riley, R.T.; Merrill, A.H. Ceramide synthase inhibition by fumonisins: A perfect storm of perturbed sphingolipid metabolism, signaling, and disease. J. Lipid Res. 2019, 60, 1183–1189. [Google Scholar] [CrossRef]
- Singh, M.P.; Kang, S.C. Endoplasmic reticulum stress-mediated autophagy activation attenuates fumonisin b1 induced hepatotoxicity in vitro and in vivo. Food Chem. Toxicol. 2017, 110, 371–382. [Google Scholar] [CrossRef]
- Arumugam, T.; Pillay, Y.; Ghazi, T.; Nagiah, S.; Abdul, N.S.; Chuturgoon, A.A. Fumonisin B1-induced oxidative stress triggers Nrf2-mediated antioxidant response in human hepatocellular carcinoma (HepG2) cells. Mycotoxin Res. 2019, 35, 99–109. [Google Scholar] [CrossRef]
- Sheik Abdul, N.; Marnewick, J.L. Fumonisin B1-induced mitochondrial toxicity and hepatoprotective potential of rooibos: An update. J. Appl. Toxicol. 2020, 40, 1602–1613. [Google Scholar] [CrossRef]
- Molina-Pintor, I.B.; Rojas-Garcia, A.E.; Medina-Dias, I.M.; Barrón-Vivanco, B.S.; Bernal-Hernández, Y.Y.; Ortega-Cervantes, L.; Ramos, A.J.; Herrera-Moreno, J.F.; Gonzáles-Ariaz, C.A. An update of geneotoxic and epigenetic studies of fumonisin B1. World Mycotoxin J. 2021, 15, 57–72. [Google Scholar] [CrossRef]
- Pinhão, M.; Tavares, A.M.; Loureiro, S.; Louro, H.; Alvito, P.; Silva, M.J. Combined cytotoxic and genotoxic effects of ochratoxin A and fumonisin B1 in human kidney and liver cell models. Toxicol. Vitr. 2020, 68, 104949. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Jia, B.; Liu, N.; Yu, D.; Zhang, S.; Wu, A. Fumonisin B1 triggers carcinogenesis via HDAC/PI3K/Akt signalling pathway in human esophageal epithelial cells. Sci. Total Environ. 2021, 787, 147405. [Google Scholar] [CrossRef] [PubMed]
- Ahangarkani, F.; Rouhi, S.; Gholamour Azizi, I. A review on incidence and toxicity of fumonisins. Toxin. Rev. 2014, 33, 95–100. [Google Scholar] [CrossRef]
- Cao, C.; Xian, R.; Lin, F.; Li, X.; Li, X.; Qiang, F.; Li, X. Fumonisin B1 induces hepatotoxicity in mice through the activation of oxidative stress, apoptosis and fibrosis. Chemosphere 2022, 296, 133910. [Google Scholar] [CrossRef]
- Yu, S.; Jia, B.; Liu, N.; Yu, D.; Wu, A. Evaluation of the Individual and combined toxicity of fumonisin mycotoxins in human gastric epithelial cells. Int. J. Mol. Sci. 2020, 21, 5917. [Google Scholar] [CrossRef]
- Gu, M.J.; Han, S.E.; Hwang, K.; Mayer, E.; Reisinger, N.; Schatzmayr, D.; Park, B.-C.; Han, S.H.; Yun, C.-H. Hydrolyzed fumonisin B1 induces less inflammatory responses than fumonisin B1 in the co-culture model of porcine intestinal epithelial and immune cells. Toxicol. Lett. 2019, 305, 110–116. [Google Scholar] [CrossRef]
- Fraeyman, S.; Croubels, S.; Devreese, M.; Antonissen, G. Emerging Fusarium and Alternaria mycotoxins: Occurrence, toxicity and toxicokinetics. Toxins 2017, 9, 228. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Novak, B.; Nagl, V.; Berthiller, F. Emerging mycotoxins: Beyond traditionally determined food contaminants. J. Agric. Food Chem. 2017, 65, 7052–7070. [Google Scholar] [CrossRef]
- De Sá, S.V.M.; Monteiro, C.; Fernandes, J.O.; Pinto, E.; Faria, M.A.; Cunha, S.C. Emerging mycotoxins in infant and children foods: A review. Crit. Rev. Food Sci. Nutr. 2021. [Google Scholar] [CrossRef]
- Mallebrera, B.; Prosperini, A.; Font, G.; Ruiz, M.J. In vitro mechanisms of beauvericin toxicity: A review. Food Chem. Toxicol. 2018, 111, 537–545. [Google Scholar] [CrossRef]
- Wu, Q.; Patocka, J.; Kuca, K. Beauvericin, a Fusarium mycotoxin: Anticancer activity, mechanisms, and human exposure risk assessment. Mini Rev. Med. Chem. 2019, 19, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Agahi, F.; Álvarez-Ortega, N.; Font, G.; Juan-García, A.; Juan, C. Oxidative stress, glutathione, and gene expression as key indicators in SH-SY5Y cells exposed to zearalenone metabolites and beauvericin. Toxicol. Lett. 2020, 334, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Cimbalo, A.; Alonso-Garrido, M.; Font, G.; Frangiamone, M.; Manyes, L. Transcriptional changes after enniatins A, A1, B and B1 ingestion in rat stomach, liver, kidney and lower intestine. Foods 2021, 10, 1630. [Google Scholar] [CrossRef] [PubMed]
- Fremy, J.M.; Alassane-Kpembi, I.; Oswald, I.P.; Cottrill, B.; Van Egmond, H.P. A review on combined effects of moniliformin and co-occurring Fusarium toxins in farm animals. World Mycotoxin J. 2019, 12, 281–291. [Google Scholar] [CrossRef]
- Pierron, A.; Neves, M.; Puel, S.; Lippi, Y.; Soler, L.; Miller, J.D.; Oswald, I.P. Intestinal toxicity of the new type A trichothecenes, NX and 3ANX. Chemosphere 2021, 288, 132415. [Google Scholar] [CrossRef] [PubMed]
- Woelflingseder, L.; Warth, B.; Vierheilig, I.; Schwartz-Zimmermann, H.; Hametner, C.; Nagl, V.; Novak, B.; Šarkanj, B.; Berthiller, F.; Adam, G. The Fusarium metabolite culmorin suppresses the in vitro glucuronidation of deoxynivalenol. Arch. Toxicol. 2019, 93, 1729–1743. [Google Scholar] [CrossRef]
- Jarolim, K.; Wolters, K.; Woelflingseder, L.; Pahlke, G.; Beisl, J.; Puntscher, H.; Braun, D.; Sulyok, M.; Warth, B.; Marko, D. The secondary Fusarium metabolite aurofusarin induces oxidative stress, cytotoxicity and genotoxicity in human colon cells. Toxicol. Lett. 2018, 284, 170–183. [Google Scholar] [CrossRef]
- Arumugam, T.; Ghazi, T.; Abdul, N.S.; Chuturgoon, A.A. A review on the oxidative effects of the fusariotoxins: Fumonisin B1 and fusaric acid. Toxicology 2021, 19, 181–190. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Control strategies: Prevention and detoxification in foods. Foods 2020, 86, 137. [Google Scholar] [CrossRef]
- Sujayasree, O.J.; Chaitanya, A.K.; Bhoite, R.; Pandiselvam, R.; Kothakota, A.; Gavahian, M.; Mousavi Khaneghah, A. Ozone: An advanced oxidation technology to enhance sustainable food consumption through mycotoxin degradation. Ozone Sci. Eng. 2022, 44, 17–37. [Google Scholar] [CrossRef]
- Wielogorska, E.; Ahmed, Y.; Meneely, J.; Graham, W.G.; Elliott, C.T.; Gilmore, B.F. A holistic study to understand the detoxification of mycotoxins in maize and impact on its molecular integrity using cold atmospheric plasma treatment. Food Chem. 2019, 301, 125281. [Google Scholar] [CrossRef] [PubMed]
- Alabouvette, C.; Olivain, C.; Migheli, Q.; Steinberg, C. Microbiological control of soil-borne phytopathogenic fungi with special emphasis on wilt-inducing Fusarium oxysporum. New Phytol. 2009, 184, 529–544. [Google Scholar] [CrossRef]
- da Cruz Cabral, L.; Fernández Pinto, V.; Patriarca, A. Application of plant derived compounds to control fungal spoilage and mycotoxin production in foods. Int. J. Food Microbiol. 2013, 166, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sui, Y.; Wisniewski, M.; Droby, S.; Liu, Y. Review: Utilization of antagonistic yeasts to manage postharvest fungal diseases of fruit. Int. J. Food Microbiol. 2013, 167, 153–160. [Google Scholar] [CrossRef]
- Guan, S.; Zhou, T.; Yin, Y.; Xie, M.; Ruan, Z.; Young, J. Microbial strategies to control aflatoxins in food and feed. World Mycotoxin J. 2011, 4, 413–424. [Google Scholar] [CrossRef]
- Abdallah, M.F.; De Boevre, M.; Landschoot, S.; De Saeger, S.; Haesaert, G.; Audenaert, K. Fungal endophytes control Fusarium graminearum and reduce trichothecenes and zearalenone in maize. Toxins 2018, 10, 493. [Google Scholar] [CrossRef]
- Abdallah, M.F.; Ameye, M.; De Saeger, S.; Audenaert, K.; Haesaert, G. Biological Control of Mycotoxigenic Fungi and Their Toxins: An Update for the Pre-Harvest Approach. Mycotoxins-Impact and Management Strategies; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Błaszczyk, L.; Basińska-Barczak, A.; Ćwiek-Kupczyńska, H.; Gromadzka, K.; Popiel, D.; Stępień, Ł. Suppressive effect of Trichoderma spp. on toxigenic Fusarium species. Pol. J. Microbiol. 2017, 66, 85. [Google Scholar] [CrossRef]
- Pellan, L.; Ahmeth, C.; Dieye, T.; Durand, N.; Fontana, A.; Strub, C.; Schorr-Galindo, S. Biocontrol agents: Toolbox for the screening of weapons against mycotoxigenic Fusarium. J. Fungi 2021, 7, 446. [Google Scholar] [CrossRef]
- Soler, S.; Rey, M.; Ait-lahsen, H.; Monte, E.; Llobell, A. Biocontrol fungus Trichoderma harzianum. Society 2001, 67, 5833–5839. [Google Scholar] [CrossRef]
- Ren, X.; Branà, M.T.; Haidukowski, M.; Gallo, A.; Zhang, Q.; Logrieco, A.F.; Li, P.; Zhao, S.; Altomare, C. Potential of Trichoderma spp. for biocontrol of aflatoxin-producing Aspergillus flavus. Toxins 2022, 14, 86. [Google Scholar] [CrossRef]
- Zin, N.A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci. 2020, 65, 168–178. [Google Scholar] [CrossRef]
- Tian, Y.; Tan, Y.; Liu, N.; Yan, Z.; Liao, Y.; Chen, J.; De Saeger, S.; Yang, H.; Zhang, Q.; Wu, A. Detoxification of deoxynivalenol via glycosylation represents novel insights on antagonistic activities of Trichoderma when confronted with Fusarium graminearum. Toxins 2016, 8, 335. [Google Scholar] [CrossRef] [PubMed]
- He, A.L.; Jia, L.I.U.; Wang, X.H.; Zhang, Q.G.; Wei, S.; Jie, C. Soil application of Trichoderma asperellum GDFS1009 granules promotes growth and resistance to Fusarium graminearum in maize. J. Integr. Agric. 2019, 18, 599–606. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Dou, K.; Lu, Z.; Wang, X.; Li, Y.; Chen, J. Enhanced biocontrol activity of cellulase from Trichoderma harzianum against Fusarium graminearum through activation of defense-related genes in maize. Physiol. Mol. Plant Pathol. 2018, 103, 130–136. [Google Scholar] [CrossRef]
- Li, Y.; Sun, R.; Yu, J.; Saravanakumar, K.; Chen, J. Antagonistic and biocontrol potential of Trichoderma asperellum ZJSX5003 against the maize stalk rot pathogen Fusarium graminearum. Indian J. Microbiol. 2016, 56, 318–327. [Google Scholar] [CrossRef]
- Tian, Y.; Tan, Y.; Yan, Z.; Liao, Y.; Chen, J.; De Boevre, M.; De Saeger, S.; Wu, A. Antagonistic and detoxification potentials of Trichoderma isolates for control of zearalenone (ZEN) producing Fusarium graminearum. Front. Microbiol. 2018, 8, 2710. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Sachadyn-Król, M.; Varzakas, T. Lactic acid bacteria as antibacterial agents to extend the shelf life of fresh and minimally processed fruits and vegetables: Quality and safety aspects. Microorganisms 2020, 8, 952. [Google Scholar] [CrossRef]
- Smaoui, S.; Agriopoulou, S.; D’Amore, T.; Tavares, L.; Khaneghah, A.M. The control of Fusarium growth and decontamination of produced mycotoxins by lactic acid bacteria. Crit. Rev. Food Sci. Nutr. 2022. [Google Scholar] [CrossRef]
- Li, J.; Wang, W.; Chen, S.; Shao, T.; Tao, X.; Yuan, X. Effect of lactic acid bacteria on the fermentation quality and mycotoxins concentrations of corn silage infested with mycotoxigenic fungi. Toxins 2021, 13, 699. [Google Scholar] [CrossRef]
- Mokoena, M.P.; Chelule, P.K.; Gqaleni, N. Reduction of fumonisin B1 and zearalenone by lactic acid bacteria in fermented maize meal. J. Food Prot. 2005, 68, 2095–2099. [Google Scholar] [CrossRef]
- Franco, T.S.; Garcia, S.; Hirooka, E.Y.; Ono, Y.S.; dos Santos, J.S. Lactic acid bacteria in the inhibition of Fusarium graminearum and deoxynivalenol detoxification. J. Appl. Microbiol. 2011, 111, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Niderkorn, V.; Morgavi, D.P.; Aboab, B.; Lemaire, M.; Boudra, H. Cell wall component and mycotoxin moieties involved in the binding of fumonisin B1 and B2 by lactic acid bacteria. J. Appl. Microbiol. 2009, 106, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Niderkorn, V.; Morgavi, D.P.; Pujos, E.; Tissandier, A.; Boudra, H. Screening of fermentative bacteria for their ability to bind and biotransform deoxynivalenol, zearalenone and fumonisins in an in vitro simulated corn silage model. Food Addit. Contam. 2007, 24, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.C.; Pante, G.C.; Centenaro, B.M.; Almeida, R.T.R.D.; Pilau, E.J.; Dias Filho, B.P.; Machinski Junior, M. Antifungal and antimycotoxigenic effects of Zingiber officinale, Cinnamomum zeylanicum and Cymbopogon martinii essential oils against Fusarium verticillioides. Food Addit. Contam. Part A 2020, 37, 1531–1541. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.C.; Endo, E.H.; de Souza, M.R.; Zanqueta, E.B.; Polonio, J.C.; Pamphile, J.A.; Ueda-Nakamura, T.; Nakamura, C.V.; Filho, B.P.D.; de Abreu Filho, B.A. Bioactivity of essential oils in the control of Alternaria alternata in dragon fruit (Hylocereus undatus Haw.). Ind. Crops Prod. 2017, 97, 101–109. [Google Scholar] [CrossRef]
- Mirza Alizadeh, A.; Golzan, S.A.; Mahdavi, A.; Dakhili, S.; Torki, Z.; Hosseini, H. Recent advances on the efficacy of essential oils on mycotoxin secretion and their mode of action. Crit. Rev. Food Sci. Nutr. 2022, 62, 4726–4751. [Google Scholar] [CrossRef]
- Gwiazdowska, D.; Marchwińska, K.; Juś, K.; Uwineza, P.A.; Gwiazdowski, R.; Waśkiewicz, A.; Kierzek, R. The concentration-dependent effects of essential oils on the growth of Fusarium graminearum and mycotoxins biosynthesis in wheat and maize grain. Appl. Sci. 2022, 12, 473. [Google Scholar] [CrossRef]
- Perczak, A.; Gwiazdowska, D.; Gwiazdowski, R.; Juś, K.; Marchwińska, K.; Waśkiewicz, A. The inhibitory potential of selected essential oils on Fusarium spp. growth and mycotoxins biosynthesis in maize seeds. Pathogens 2020, 9, 23. [Google Scholar] [CrossRef]
- López, A.G.; Theumer, M.G.; Zygadlo, J.A.; Rubinstein, H.R. Aromatic plants essential oils activity on Fusarium verticillioides fumonisin B1 production in corn grain. Mycopathologia 2004, 158, 343–349. [Google Scholar] [CrossRef]
- Pante, G.C.; Castro, J.C.; Lini, R.S.; Romoli, J.C.Z.; Almeida, R.T.R.D.; Garcia, F.P.; Machinski, M. Litsea cubeba essential oil: Chemical profile, antioxidant activity, cytotoxicity, effect against Fusarium verticillioides and fumonisins production. J. Environ. Sci. Health A 2021, 56, 387–395. [Google Scholar] [CrossRef]
- Perczak, A.; Juś, K.; Marchwińska, K.; Gwiazdowska, D.; Waśkiewicz, A.; Goliński, P. Degradation of zearalenone by essential oils under in vitro conditions. Front. Microbiol. 2016, 7, 1224. [Google Scholar] [CrossRef] [PubMed]
- Xing, F.; Hua, H.; Selvaraj, J.N.; Yuan, Y.; Zhao, Y.; Zhou, L.; Liu, Y. Degradation of fumonisin B1 by cinnamon essential oil. Food Control 2014, 38, 37–40. [Google Scholar] [CrossRef]
- Makhuvele, R.; Naidu, K.; Gbashi, S.; Thipe, V.C.; Adebo, O.A.; Njobeh, P.B. The use of plant extracts and their phytochemicals for control of toxigenic fungi and mycotoxins. Heliyon 2020, 6, e05291. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Ramos, A.J.; Sanchis, V.; Marín, S. Effect of Equisetum arvense and Stevia rebaudiana extracts on growth and mycotoxin production by Aspergillus flavus and Fusarium verticillioides in maize seeds as affected by water activity. Int. J. Food Microbiol. 2012, 153, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Seepe, H.A.; Lodama, K.E.; Sutherland, R.; Nxumalo, W.; Amoo, S.O. In vivo antifungal activity of South African medicinal plant extracts against Fusarium pathogens and their phytotoxicity evaluation. Plants 2020, 9, 1668. [Google Scholar] [CrossRef]
- Ferruz, E.; Loran, S.; Herrera, M.; Gimenez, I.; Bervis, N.; Barcena, C.; Carramiñana, J.J.; Juan, T.; Herrera, A.; Ariño, A. Inhibition of Fusarium growth and mycotoxin production in culture medium and in maize kernels by natural phenolic acids. J. Food Prot. 2016, 79, 1753–1758. [Google Scholar] [CrossRef]
- Montibus, M.; Vitrac, X.; Coma, V.; Loron, A.; Pinson-Gadais, L.; Ferrer, N.; Verdal-Bonnin, M.N.; Gabaston, J.; Waffo-Téguo, P.; Richard-Forget, F.; et al. Screening of wood/forest and vine by-products as sources of new drugs for sustainable strategies to control Fusarium graminearum and the production of mycotoxins. Molecules 2021, 26, 405. [Google Scholar] [CrossRef]
- Savoie, J.M.; Mata, G.; Atanasova Penichon, V.; Foulonge-Oriol, M. Using mushroom-forming fungi in preventing and reducing mycotoxins in cereal products. Sci. Fungorum 2019, 49, e1256. [Google Scholar] [CrossRef]
- El-Ramady, H.; Abdalla, N.; Fawzy, Z.; Badgar, K.; Llanaj, X.; Törős, G.; Hajdú, P.; Eid, Y.; Prokisch, J. Green biotechnology of oyster mushroom (Pleurotus ostreatus L.): A sustainable strategy for myco-remediation and bio-fermentation. Sustainability 2022, 14, 3667. [Google Scholar] [CrossRef]
- Mihai, R.A.; Melo Heras, E.J.; Florescu, L.I.; Catana, R.D. The edible gray oyster fungi Pleurotus ostreatus (Jacq. ex Fr.) P. Kumm a potent waste consumer, a biofriendly species with antioxidant activity depending on the growth substrate. J. Fungi 2022, 8, 274. [Google Scholar] [CrossRef]
- Branà, M.T.; Cimmarusti, M.T.; Haidukowski, M.; Logrieco, A.F.; Altomare, C. Bioremediation of aflatoxin B1-contaminated maize by king oyster mushroom (Pleurotus eryngii). PLoS ONE 2017, 12, e0182574. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.W.; Pryor, B.M. Degradation of aflatoxin B1 from naturally contaminated maize using the edible fungus Pleurotus ostreatus. AMB Express 2017, 7, 110. [Google Scholar] [CrossRef] [PubMed]
- Motomura, M.; Toyomasu, T.; Mizuno, K.; Shinozawa, T. Purification and characterization of an aflatoxin degradation enzyme from Pleurotus ostreatus. Microbiol. Res. 2003, 158, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Alberts, J.F.; Gelderblom, W.C.A.; Botha, A.; van Zyl, W.H. Degradation of aflatoxin B1 by fungal laccase enzymes. Int. J. Food Microbiol. 2009, 135, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Yehia, R.S. Aflatoxin detoxification by manganese peroxidase purified from Pleurotus ostreatus. Braz. J. Microbiol. 2014, 45, 127–133. [Google Scholar] [CrossRef]
- Merel, D.; Savoie, J.M.; Mata, G.; Salmones, D.; Ortega, C.; Atanasova, V.; Chéreau, S.; Monribot-Villanueva, J.L.; Guerrero-Analco, J.A. Methanolic extracts from cultivated mushrooms affect the production of fumonisins b and fusaric acid by Fusarium verticillioides. Toxins 2020, 12, 366. [Google Scholar] [CrossRef]


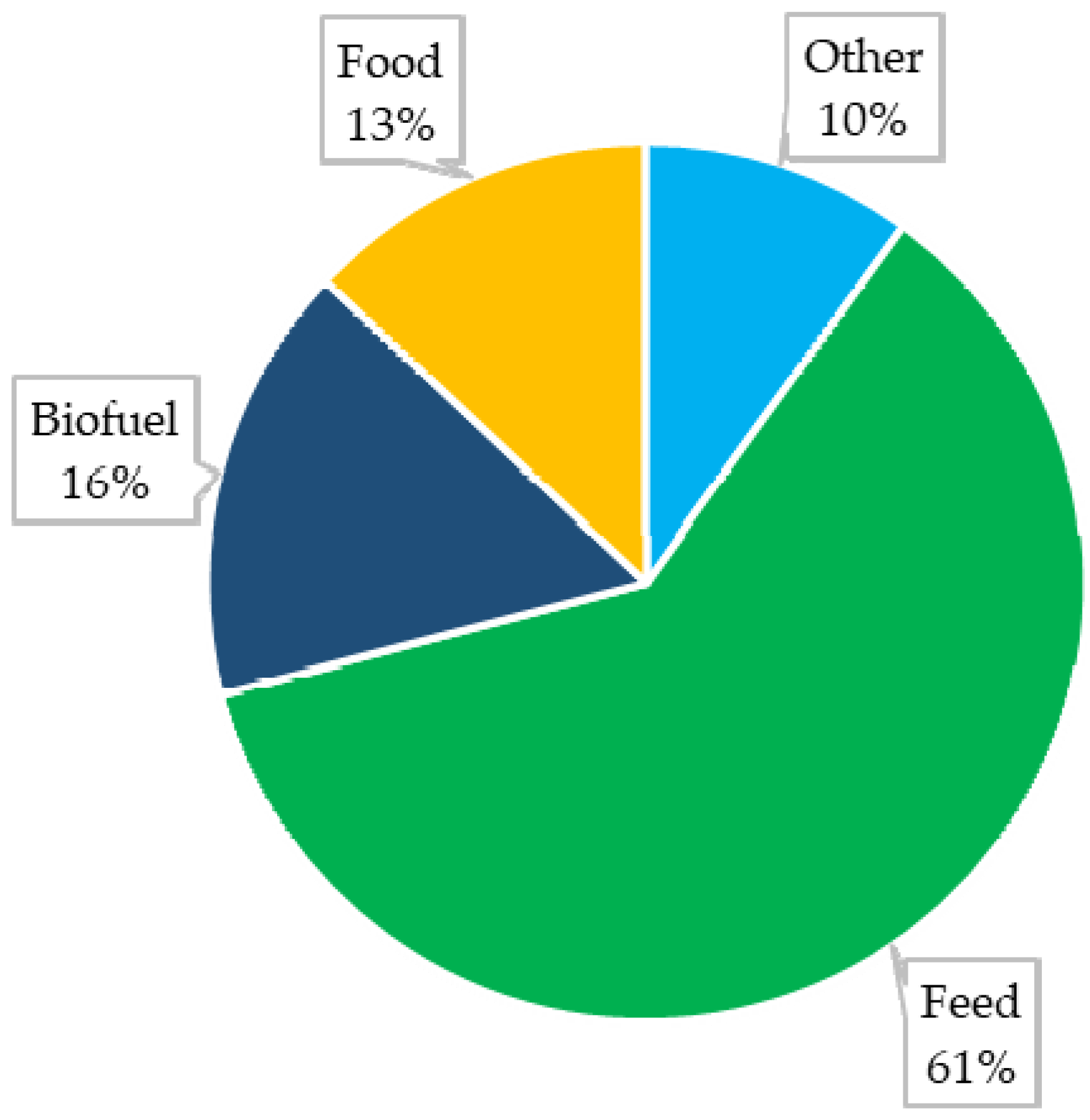{kind=link}
| Mycotoxin | Abbreviation | Molar Mass [g/mol] | Produced by: | Source |
|---|---|---|---|---|
| Fumonisin B1 | FB1 | 721.83 | F. dlamini F. globosum F. nygamai F. oxysporum F. proliferatum F. subglutinans F. temperatum F. thapsinum F. verticillioides | [26,32,33,34,35,36,37] |
| Fumonisin B2 | FB2 | 705.83 | ||
| Fumonisin B3 | FB3 | 705.83 | ||
| Fumonisin B4 | FB4 | 689.83 | ||
| Fumonisin A1 | FA1 | 763.90 | ||
| Zearalenone | ZEN | 318.36 | F. cerealis F. culmorum F. equiseti F. graminearum F. heterosporum F. meridionale F. semitectum | [26,32,33,36,37,38] |
| HT-2 toxin | HT-2 | 424.48 | F. acuminatum F. armeniacum F. langsethiae F. poae F. sporotrichioides | [26,33,36,37] |
| T-2 toxin | T-2 | 466.52 | F. acuminatum F. armeniacum F. equiseti F. langsethiae F. poae F. sambucinum F. sporotrichioides | [26,33,36,37] |
| Deoxynivalenol | DON | 296.31 | F. boothii F. culmorum F. graminearum F. meridionale | [26,32,33,36,37] |
| Nivalenol | NIV | 312.32 | F. cerealis F. cortaderiae F. culmorum F. equiseti F. graminearum F. meridionale F. poae | [26,32,33,36,37,39] |
| 15-acetyl-deoxynivalenol and 3-acetyldeoxynivalenol | 15- and 3-AcDON | 338.35 | F. culmorum F. graminearum F. meridionale | [32,33,37] |
| Mycotoxin | Abbreviation | Molar Mass [g/mol] | Type of Modification | Source |
|---|---|---|---|---|
| Deoxynivalenol 3-β-D -glucoside | DON-3G | 458.46 | plant | [26,33,37] |
| Zearalenone-14-sulphate | ZEN-14S | 415.46 | fungi | [26,37] |
| α-Zearalenol | α-ZOL | 320.39 | fungi or/and plant | [33,37] |
| β-Zearalenol | β-ZOL | 320.39 | fungi or/and plant | [33,37] |
| Hydrolysed fumonisin B1 | HFB1 | 405.61 | fungi | [26,37,48] |
| Mycotoxin | Abbreviation | Molar Mass [g/mol] | Produced by: | Source |
|---|---|---|---|---|
| Beauvericin | BEA | 783.96 | F. acuminatum F. anthophilum F. armeniacum F. avenaceum F. dlamini F. equiseti F. globosum F. nygamai F. oxysporum F. proliferatum F. poae F. sambucinum F. semitectum F. sporotrichioides F. subglutianans F. temperatum F. tricinctum F. verticillioides | [26,33,36,37] |
| Culmorin | CUL | 238.37 | F. culmorum F. cerealis F. graminearum F. langsethiae F. poae F. sporotrichioides F. venenatum | [26,36,37] |
| 15Hydroxyculmorin | 15h-CUL | 254.36 | F. culmorum F. graminearum F. poae | [26,37] |
| 5Hydroxyculmorin | 5h-CUL | 254.36 | F. culmorum F. graminearum F. poae | [26] |
| Monoacetoxyscirpenol | MAS | 324.40 | F. acuminatum F. equiseti F. sporotrichioides F. poae | [26,33,37] |
| Diacetoxyscirpenol | DAS | 366.41 | F. acuminatum F. armeniacum F. boothii F. cerealis F. cortaderiae F. culmorum F. equiseti F. graminearum F. meridionale F. sambucinum F. semitectum F. sporotrichoides F. poae | [26,33,36,37] |
| Neosolaniol | NEO | 382.40 | F. acuminatum F. poae F. sambucinum | [26,33,37] |
| Moniliformin | MON | 120.04 | F. acuminatum F. avenaceum F. chlamydosporum F. culmorum F. dlamini F. equiseti F. fusariodes F. oxysporum F. proliferatum F. semitectum F. sporotrichioides F. subglutians F. temperatum F. thapsinum F. tricinctum | [26,33,34,36,37] |
| Enniatin A | ENN A | 681.90 | F. acuminatum F. avenaceum F. chlamydosporum F. culmorum F. moniliforme F. nivale F. oxysporum F. poae F. proliferatum F. roseumF. solani F. sporotrichioides F. tritinctum | [26,36,37] |
| Enniatin A1 | ENN A1 | 667.90 | ||
| Enniatin B | ENN B | 639.82 | ||
| Enniatin B1 | ENN B1 | 653.85 | ||
| Enniatin B2 | ENN B2 | 625.80 | ||
| Aurofusarin | Au-FU | 570.50 | F. acuminatum F. avenaceum F. cerealis F. culmorum F. graminearum F. poae F. sambucinum F. sporotrichiodes F. tricinctum | [26,36,37] |
| Bikaverin | BKV | 382.3 | F. antholphilum F. bulbigenum F. dlamini F. fujikuroi F. moniliforme F. napiformi F. nygamai F. oxysporum F. proliferatum F. solani F. subglutitans F. verticillioides | [26,37] |
| Butenolide | BUT | 156.14 | F. cerealis F. culmorum F. graminearum F. poae F. sporotrichioides F. verticillioides | [26,36,37] |
| Equisetin | EQ | 373.49 | F. equiseti F. semitectum | [37] |
| Fusaric acid | FA | 179.22 | F. cerealis F. oxysporum F. proliferatum F. solani F. sublutinans F. thapsinum F. verticillioides | [26,36,37] |
| Siccanol | SCN | 402.58 | F. graminearum | [26,37] |
| Fusaproliferin | FUS | 444.60 | F. fujikuroi F. globosum F. proliferatum F. subglutinans F. temperatum | [33,36,37] |
| Mycotoxin | Positive Samples (%) | Min-Max [µg/kg] | Origin | References |
|---|---|---|---|---|
| FB1 | 121/123 (98%) | 12.6–8908 | South Africa | [26] |
| 40/79 (51%) | 68–2453 * | Egypt | [57] | |
| 9/12 (75%) | <LOQ–49 ** | China | [58] | |
| 34/45 (76%) | 59.9–9873 | Albania | [59] | |
| 79/90 (88%) | <LOD–45,145.82 | Michigan (USA) | [60] | |
| 70 (71.4%) | 1080 *** | Spain | [61] | |
| 48/55 (88%) | 101–1838 | West Africa (Togo) | [62] | |
| 55/158 (34.8%) | 60–553 * | Europe (various countries) | [63] **** | |
| FB2 | 112/123 (91%) | 7.9–3383 | South Africa | [26] |
| 14/79 (18%) | 4.7–386 * | Egypt | [57] | |
| 27/45 (60%) | 105–9218 | Albania | [59] | |
| 0/12 (0%) | <LOD | China | [58] | |
| 76/90 (84%) | <LOD–22,538.63 | Michigan (USA) | [60] | |
| 55/98 (56.1%) | 1306 *** | Spain | [61] | |
| 30/55 (55%) | 45–586.4 | West Africa (Togo) | [62] | |
| 46/158 (89.1%) | 20.4–133 * | Europe (various countries) | [63] **** | |
| FB3 | 98/123 (80%) | <LOQ–990 | South Africa | [26] |
| 6/79 (8%) | 16.8–286 * | Egypt | [57] | |
| 72/90 (80%) | <LOD–17,972.72 | Michigan (USA) | [60] | |
| 14/55 (26%) | 43–185.6 | West Africa (Togo) | [62] | |
| FB4 | 101/123 (82%) | <LOQ–1014 | South Africa | [26] |
| 29/30 (96%) | 461–2716 * | Nigeria | [64] | |
| FB1 + FB2 | 148/148 (100%) | 62.4–6 | Brasil | [65] |
| 6274 | ||||
| 34/45 (76%) | 59.9–16,970 | Albania | [59] | |
| FA1 | 66/123 (54%) | <LOQ–51.4 | South Africa | [26] |
| HFB1 ***** | 9/12 (75%) | 144–7226 ** | China | [58] |
| HFB2 ***** | 0/12 (0%) | <LOD | China | [58] |
| ZEN | 41/123 (33%) | <LOQ–146 | South Africa | [26] |
| 5/6 (83%) | <LOD–1071 | Belgium | [46] | |
| 10/79 (13%) | 3.4–184 * | Egypt | [57] | |
| 107/158 (67.7%) | 15.2–1670 * | Europe (various countries) | [63] **** | |
| 2/45 (4.4%) | 218–263 | Albania | [59] | |
| 76/90 (84%) | 0.56–4148.75 | Michigan (USA) | [60] | |
| 24/98 (24.5%) | 110.3 *** | Spain | [61] | |
| 1/55 (2%) | 79–79 | West Africa (Togo) | [62] | |
| 115/120 (96%) | <LOD–3910 | Germany | [66] | |
| 42 (73.6%) | 1.4–444.6 | Brasil | [65] | |
| 13/23 (56.52%) | <LOD–163.58 | China | [52] | |
| ZEN-14Glc | 1/6 (17%) | 274–274 | Belgium | [46] |
| ZEN-14S | 1/6 (17%) | 51–51 | Belgium | [46] |
| α-ZEL | 6/6 (100%) | 22–262 | Belgium | [46] |
| 71/120 (59%) | <LOD–423 | Germany | [66] | |
| β-ZEL | 4/6 (67%) | 12–103 | Belgium | [46] |
| 38/120 (32%) | <LOD–203 | Germany | [66] | |
| DON | 61/123 (50%) | 8.2–1380 | South Africa | [26] |
| 11/150(7.3%) | 270–1980 | Ethiopia | [67] | |
| 6/6 (100%) | 411–5245 | Belgium | [46] | |
| 6/79 (8%) | 311–807 * | Egypt | [57] | |
| 11/45 (24%) | 110–798 | Albania | [59] | |
| 120/120 (100%) | <LOQ–10,972 | Germany | [66] | |
| 107/158 (67.7%) | 303–3060 * | Europe (various countries) | [63] **** | |
| 87/90 (97%) | <LOD–20,475 | Michigan (USA) | [60] | |
| 81/81 (100%) | 44–614 | Northen Italy (Pidemont) | [68] | |
| 63/81 (78%) | <LOD–216 | Northen Italy (Lombardy) | ||
| 72/81 (89%) | <LOD–83 | Northen Italy (Vento) | ||
| 31/98 (31.6%) | 180 *** | Spain | [61] | |
| 33 (58%) | 41.6–1008 | Brasil | [65] | |
| DON-3G | 65/123 (53%) | 2.43–112 | South Africa | [26] |
| 120/120 (100%) | 95–3038 | Germany | [66] | |
| 4/79 (5%) | <LOQ–47.5 * | Egypt | [57] | |
| 8/10 (80%) | 14–121 | China | [69] | |
| 78/90 (87%) | <LOD–6266.49 | Michigan (USA) | [60] | |
| 40/158 (25.5%) | 17.1–129 * | Europe (various countries) | [63] **** | |
| 3-AcDON | 6/6 (100%) | 63–643 | Belgium | [46] |
| 29/90 (32%) | <LOD–63.04 | Michigan (USA) | [60] | |
| 5/98 (5.1%) | 63.9 *** | Spain | [61] | |
| 2/79 (3%) | <LOQ | Egypt | [57] | |
| 108/257 (42%) | 0–1046.8 * | Belgium | [70] | |
| 15-AcDON | 6/6 (100%) | 61–792 | Belgium | [46] |
| 66/90 (73.3%) | <LOD–1787.6 | Michigan (USA) | [60] | |
| 112/257 (43%) | 0–819.3 * | Belgium | [70] | |
| NIV | 14/123 (11%) | 7.7–35.7 | South Africa | [26] |
| 10/30 (33.3%) | 5.1–50.8 * | Nigeria | [64] | |
| 8/79 (10%) | 1.6–142 * | Egypt | [57] | |
| 10/30 (33.3%) | 5.1–50.8 | Nigeria | [64] | |
| T-2 | 1/123 (0.8%) | 148–148 | South Africa | [26] |
| 1/45 (2.2%) | 106–106 * | Albania | [59] | |
| 17/90 (19%) | <LOD–156.65 | Michigan (USA) | [60] | |
| 5/98 (5.1%) | 8.6 *** | Spain | [61] | |
| 6/158 (3.8%) | 2.55–4.08 * | Europe (various countries) | [63] **** | |
| HT-2 toxin | 1/123 (0.8%) | 40.2–40.2 | South Africa | [26] |
| 5/98 (5.1%) | 11.7 *** | Spain | [61] | |
| 4/90 (4%) | <LOD–276.74 | Michigan (USA) | [60] |
| Mycotoxin | Positive Samples (%) | Min–Max [µg/kg] | Origin | References |
|---|---|---|---|---|
| MON | 120/123 (98%) | <LOQ–1130 | South Africa | [26] |
| 26/29 (89.7%) | 15.3–1450 | Serbia (South-Backa) | [54] | |
| 21/21 (100%) | 5.06–850 | Serbia (South-Banat) | ||
| 3/6 (50%) | 34.8–405 | Serbia (West-Backa) | ||
| 12/12 (100%) | 7.18–1228 | Serbia (Middle-Banat) | ||
| 5/5 (100%) | 3.03–3856 | Serbia (Srem) | ||
| 18/79 (23%) | 1.6–142 * | Egypt | [57] | |
| 54/90 (60%) | <LOD–1160.35 | Michigan (USA) | [60] | |
| 81/81 (100%) | 93–751 | Northen Italy (Pidemont) | [68] | |
| 81/81 (100%) | 592–4800 | Northen Italy (Lombardy) | ||
| 81/81 (100%) | 8–1613 | Northen Italy (Vento) | ||
| 20/30 (66%) | 25.3–1387 * | Nigeria | [64] | |
| BEA | 107/123 (87%) | <LOQ–142 | South Africa | [26] |
| 20/21 (95.2%) | 0.41–129 | Serbia (South-Banat) | [54] | |
| 26/29 (89.7%) | 0.10–111 | Serbia (South-Backa) | ||
| 11/12 (91.7%) | 0.23–49.7 | Serbia (Middle-Banat) | ||
| 4/5 (80%) | 0.27–136 | Serbia (Srem) | ||
| 3/6 (50%) | 0.03–18.2 | Serbia (West-Backa) | ||
| 30/30 (100%) | 2.5–329 * | Nigeria | [64] | |
| 80/90 (89%) | 1.04–7446.21 | Michigan (USA) | [60] | |
| 50/79 (63%) | 0.64–72 * | Egypt | [57] | |
| FUS | 16/21 (76.2%) | 85.4–1121 | Serbia (South-Banat) | [54] |
| 11/12 (91.7%) | 450–1738 | Serbia (Middle-Banat) | ||
| 3/5 (60%) | 312–4488 | Serbia (Srem) | ||
| 22/29 (75.9%) | 91.3–4687 | Serbia (South-Backa) | ||
| 1/6 (16.7%) | 12,272–12,272 | Serbia (West-Backa) | ||
| 2/30 (7.1%) | 0.3–1.3 | Nigeria | [64] | |
| DAS | 2/123 (1.7%) | 4.4–5.0 | South Africa | [26] |
| 9/55 (17%) | 2.2–3 | West Africa (Togo) | [62] | |
| 12/30 (40%) | 5.9–6.59 * | Nigeria | [64] | |
| 22/257 (8.6%) | 0–14.9 * | Belgium | [70] | |
| MAS | 1/123 (0.8%) | 20.9–20.9 | South Africa | [26] |
| 4/158 (2.5%) | 9.91–32.9 * | Europe (various countries) | [63] ** | |
| NEO | 1/123 (0.8%) | 4.5–4.5 | South Africa | [26] |
| ENN A | 3/29 (10.3%) | 0.12–0.47 | Serbia (South-Backa) | [54] |
| 2/12 (16.7%) | 0.41–17.1 | Serbia (Middle-Banat) | ||
| 1/6 (16.7%) | 0.49 | Serbia (West-Backa) | ||
| 50/90 (56%) | <LOD–21.84 | Michigan (USA) | [60] | |
| ENN A1 | 4/90 (4%) | <LOD–27.28 | Michigan (USA) | [60] |
| 3/29 (10.3%) | 0.13–0.44 | Serbia (South-Backa) | [54] | |
| 3/12 (4.8%) | 0.11–27.4 | Serbia (Middle-Banat) | ||
| 1/21 (4.8%) | 0.59 | Serbia (South-Banat) | ||
| ENN B | 47/90 (52%) | <LOD–2.34 | Michigan (USA) | [60] |
| 1/21 (4.8%) | 7.55 | Serbia (South-Banat) | [54] | |
| 2/12 (16.7%) | 0.08–1.52 | Serbia (Middle-Banat) | ||
| 93/257 (36.2%) | 46.2–1984.9 * | Belgium | [70] | |
| ENN B1 | 6/90 (7%) | <LOD–7.94 | Michigan (USA) | [60] |
| 1/21 (4.8%) | 4.89 | Serbia (South-Banat) | [54] | |
| 1/6 (16.7%) | 0.22 | Serbia (West-Backa) | ||
| 2/12 (16.7%) | 0.20–16.3 | Serbia (Middle-Banat) | ||
| CUL | 18/123 (15%) | 13.3–465 | South Africa | [26] |
| 125/158 (79.1%) | 190–6680 | Europe (various countries) | [63] ** | |
| BUT | 35/123 (28%) | <LOQ–214 | South Africa | [26] |
| 30/158 (19%) | 28.9–583 | Europe (various countries) | [63] | |
| FA | 24/123 (20%) | 57–195 | South Africa | [26] |
| 35//158 (22.2%) | 229–4120 | Europe (various countries) | [63] | |
| Bikaverin | 82/123 (67%) | <LOQ–651 | South Africa | [26] |
| 42/158 (26.6%) | 20.3–415 | Europe (various countries) | [63] ** | |
| 15Hydroxyculmorin | 49/123 (40%) | <LOQ–2022 | South Africa | [26] |
| 84/158 (53.2%) | 229–1670 | Europe (various countries) | [63] | |
| 5Hydroxyculmorin | 18/123 (15%) | <LOQ–578 | South Africa | [26] |
| 19/158 (12%) | 571–1480 | Europe (various countries) | [63] ** | |
| Apicidin | 2/123 (1.6%) | 2.9–15.4 | South Africa | [26] |
| 79/123 (50%) | 9.49–175 | Europe (various countries) | [63] ** | |
| Aurofusarin | 89/123 (72%) | <LOQ–5470 | South Africa | [26] |
| Epiequisetin | 19/123 (15%) | <LOQ–18.9 | ||
| Equisetin | 30/123 (24%) | <LOQ–129 | ||
| Fusarinolic acid | 24/123 (20%) | <LOQ–3422 | ||
| Acuminatum B | 12/123 (9.8%) | <LOQ–219 | ||
| Acuminatum C | 7/123 (5.7%) | <LOQ–204 | ||
| Chlamydospordiol | 2/123 (1.7%) | 2.1–5.1 | ||
| Chlamydosporol | 1/123 (0.8%) | 87.0–26.9 | ||
| Chrysogin | 48/123 (30%) | <LOQ–7.7 | ||
| Siccanol | 91/123 (74%) | 34.6–252 | ||
| Fusapyron | 38/123 (31%) | <LOQ–18.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bryła, M.; Pierzgalski, A.; Zapaśnik, A.; Uwineza, P.A.; Ksieniewicz-Woźniak, E.; Modrzewska, M.; Waśkiewicz, A. Recent Research on Fusarium Mycotoxins in Maize—A Review. Foods 2022, 11, 3465. https://doi.org/10.3390/foods11213465
Bryła M, Pierzgalski A, Zapaśnik A, Uwineza PA, Ksieniewicz-Woźniak E, Modrzewska M, Waśkiewicz A. Recent Research on Fusarium Mycotoxins in Maize—A Review. Foods. 2022; 11(21):3465. https://doi.org/10.3390/foods11213465
Chicago/Turabian StyleBryła, Marcin, Adam Pierzgalski, Agnieszka Zapaśnik, Pascaline Aimee Uwineza, Edyta Ksieniewicz-Woźniak, Marta Modrzewska, and Agnieszka Waśkiewicz. 2022. "Recent Research on Fusarium Mycotoxins in Maize—A Review" Foods 11, no. 21: 3465. https://doi.org/10.3390/foods11213465
APA StyleBryła, M., Pierzgalski, A., Zapaśnik, A., Uwineza, P. A., Ksieniewicz-Woźniak, E., Modrzewska, M., & Waśkiewicz, A. (2022). Recent Research on Fusarium Mycotoxins in Maize—A Review. Foods, 11(21), 3465. https://doi.org/10.3390/foods11213465