
Macroalgae-Derived Multifunctional Bioactive Substances: The Potential Applications for Food and Pharmaceuticals

 , and
, and
Abstract
1. Introduction
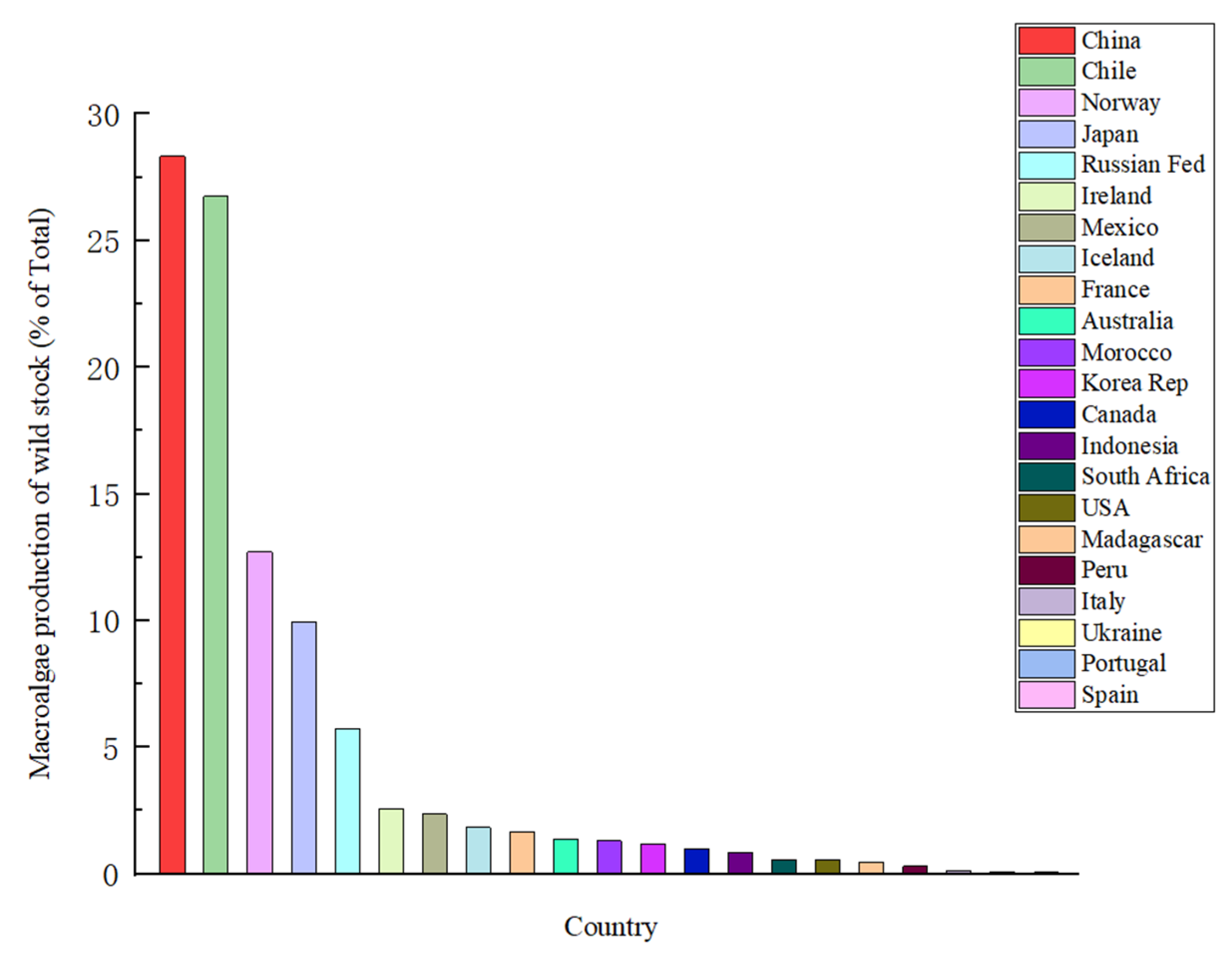
2. The Distribution Characteristics of Macroalgae
2.1. Distribution of Macroalgae in Intertidal Areas
2.2. Distribution of Macroalgae in Mangrove Areas
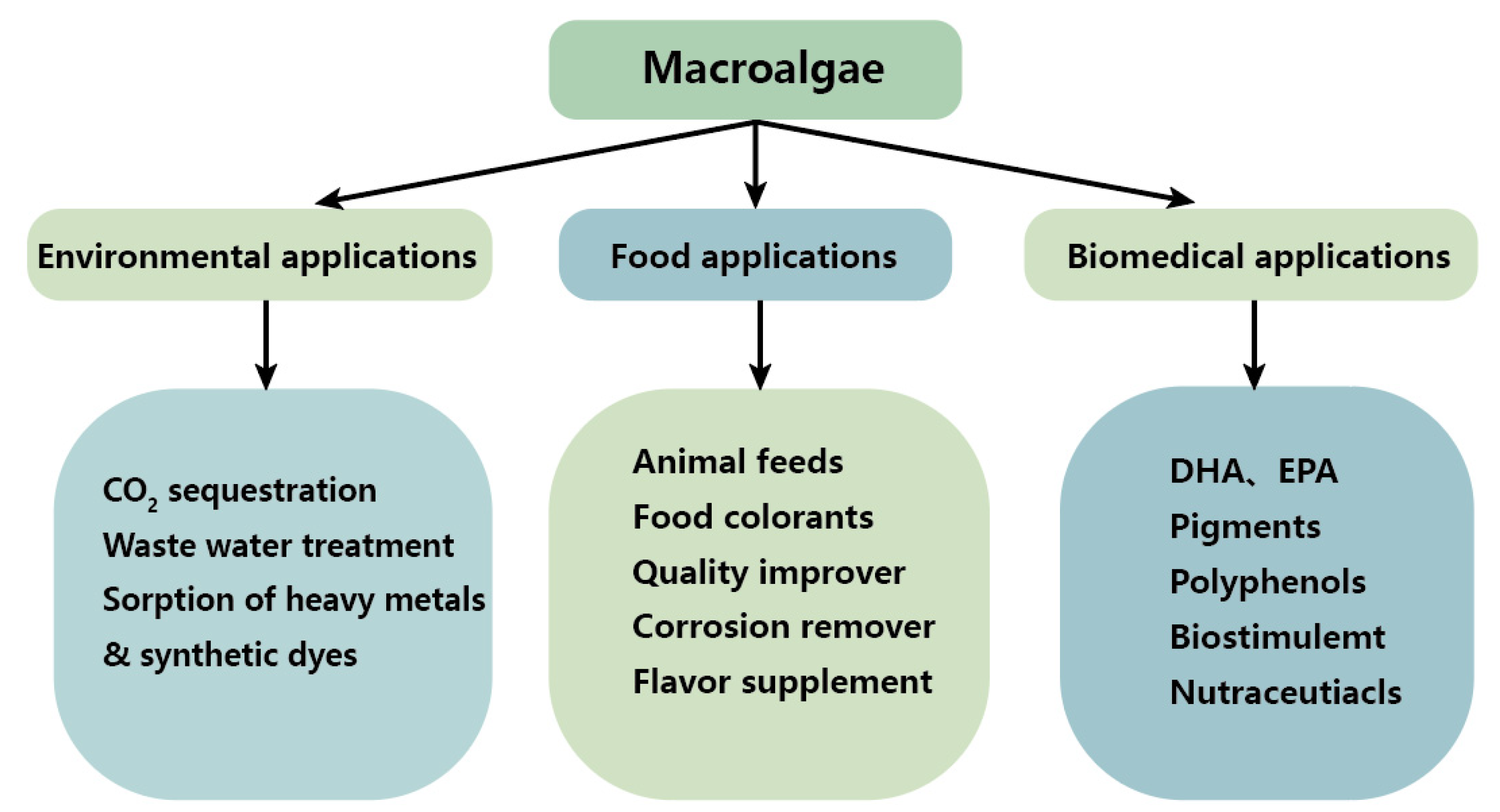
3. Nutritional Value of Macroalgae

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macroalgae | Photosynthetic Pigments | Photosynthetic Rate | Algae | Process | Process Conditions | Generated Products | References |
|---|---|---|---|---|---|---|---|
| Rhodophyta | Phycoerythrin | 20–1808.7 (μmol CO2/h) g/dry | Gracilaria corticata | SLE | Water, 12 h, 4 °C; UMFb, 200 kDa cut-off poly sulphone membrane | Pigments (R-PE and R-PC) Mineral-rich water | [22] |
| Gracilaria gracilis | Fast pyrolysis | 400, 500 and 600 °C, heating rate 50 °C, atmospheric pressure, inert atmosphere | Liquid | [23] | |||
| Kappa-phycus alvarezii | Thermal acid hydrolysis | 30–110 mM H2SO4, 30–120 min, 121 °C, pH 5.0 with NaOH | Agar | [24] | |||
| Gracilaria vermiculophylla | Water Extraction | Pre-treatment (5% NaOH, 80 °C, 2 h); neutralization (1.5% H2SO4, 2 h); water extraction at boiling tempera-ture (90 min), 50 g/L | Agar | [25] | |||
| Chlorophyta | Chlorophyll a, b, carotene and Xanthophyll | 30 to 1786 (μmol CO2/h) g/dry | Ulva fasciata | SLE | Water, 500 g/L, grinding | MRLE (mineral-rich liquid extract) | [26] |
| Ulva lactuca | SLE | 500 g/L, Water, crushing (biomass mechanically disintegrated into tiny particles) and filtration | Sap | [27] | |||
| Ulva lactuca | Lipids extraction | Lyophilization, cell destruction with a high-pressure homogenizer, sequen-tial solvent extraction (3 times) with Folch solution | Lipids | [28] | |||
| Ulva lactuca | SLE | 0.25 M NaOH (optimized alkali con-centration), 50 g/L, 60 °C, 1 h | Proteins | [29] | |||
| Phaeophyt | Fucoxanthin | 100–500 (μmol CO2/h) g/dry | Laminaria digitata | MAE | 0.1 M HCl, microwave irradiation, 15 min, 90 °C; Alginate removal by 2% CaCl2; Fucoidan precipitation by ethanol | Fucoidan | [30] |
| Laminaria digitata | Enzymatic hydrolysis | Enzymatic hydrolysis | Bioethanol | [31] | |||
| Sargassum muticum | Ethanolic extraction and SC-CO2 | Pilot plant extractor, 1 h, 50 °C, 10 and 35 MPa, 25 g CO2 min−1 | Fucoxanthin | [32] | |||
| Saccharina japonica | SLE | 0.5–4.0 wt% sodium carbonate, 100 g/L, 40–80 °C, 60–120 min | Alginate | [33] |
3.1. Food Additives
3.1.1. Flavor Supplements
3.1.2. Quality Improver
3.1.3. Preservatives and Nutritional Supplements
3.2. Animal Feeds
| Livestock | Seaweed Species | Extract | Add the Amount | Main Biological Effects in the Animals | References |
|---|---|---|---|---|---|
| Barramundi (Lates calcarifer) | Gracilaria pualvinata | Dry | 3%, 6%, and 9% | Increased levels of seaweed supplementation were negatively correlated with the serum triglycerides and cholesterol in the animals. | [55] |
| Buck and doe rabbits | Ulva lactuca | Fresh or dry | 1% and 2% | Seaweed supplementation improved the reproductive performance of rabbits by improving the semen fertility characteristics of bucks. | [56] |
| Female pigs | Seaweed extract (No species mentioned) | Fresh or dry | 180 mg laminarin and 340 mg fucoidan per kilogram feed | Seaweed extracts reduced the gene expression of pro-inflammatory cytokines in the colon of supplemented pigs after an experimental infection with Salmonella Typhimurium compared to animals receiving a basal diet. | [57] |
| Lohmann Lite hens | Chondrus crispus and Sarcodiotheca gaudichaudii | Fresh or dry | 2% and 4% | The incorporation of seaweed in the diet of hens reduced the negative effects on body weight and egg production of a challenge with Salmonella enteritidis compared to birds receiving a basal diet. | [58] |
| Nile tilapia (Oreochiromisniloticx) | Ulva sp. (mixture of Ulva rigida 5% and 10% and ulva lactaca) | Fresh or dry | 5% and 10% | Seaweed meal increased total carotenoid contents in the skin and enhanced immune responses in supplemented fish. Seaweed supplementation showed no effects on the growth performance or the organoleptic properties of the flesh with respect to fish receiving a basal diet. | [59] |
| Red tilapia (Oreochronis miloticus) | Fermented Enteromorpha prolifera | Dry | 1%, 2%, 3%, 4% and 5% | Fermented seaweed had positive effects on the growth performance, the activity of the digestive enzymes and the immunity of the fish from the supplemented group with respect to control. The recommended optimum level of inclusion of this algal product in the diet of fish ranged between 3.7% and 4.19%. | [60] |
4. The potential Medicinal Applications of Macroalgae
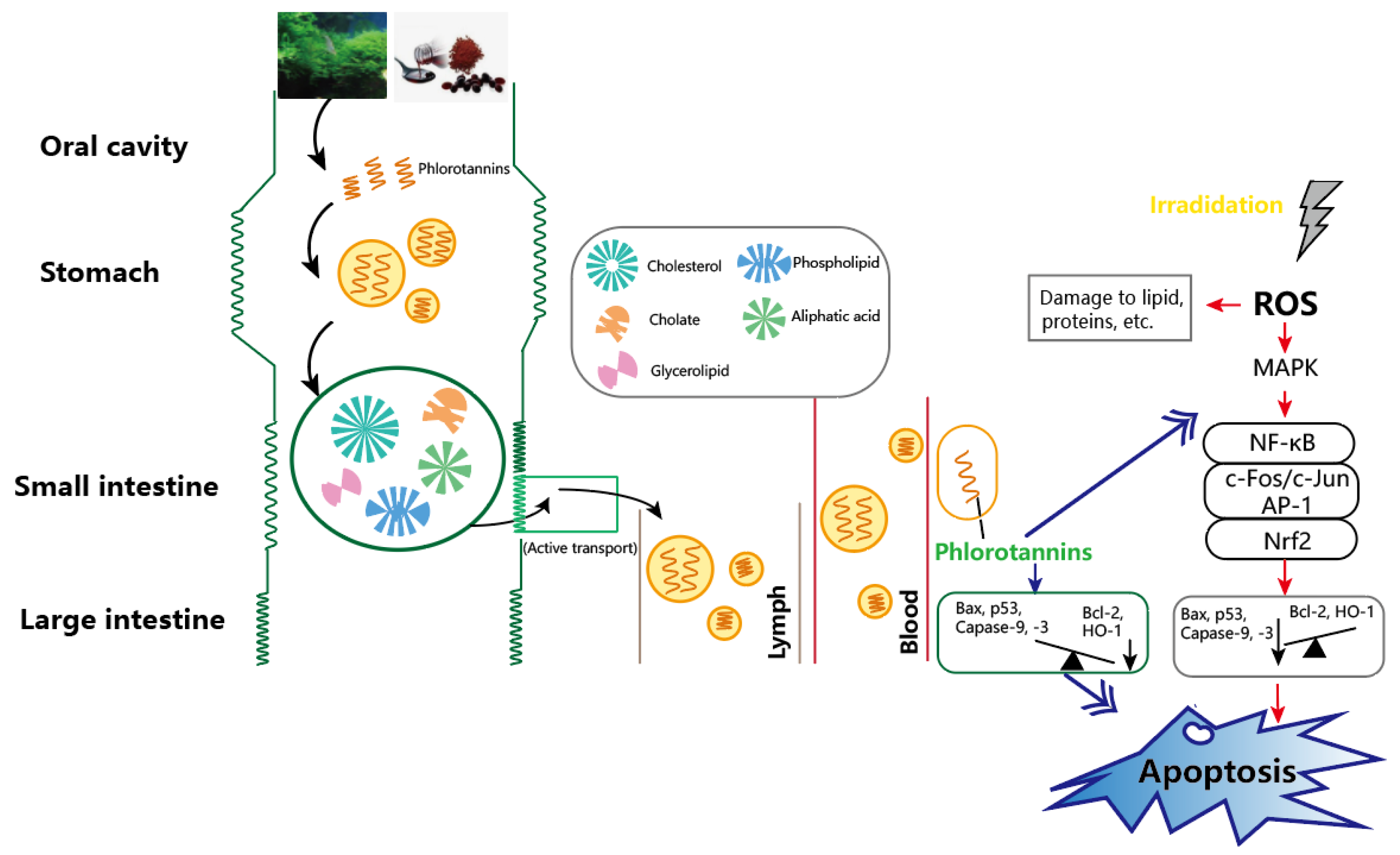
4.1. Polyphenols
4.2. Polysaccharides
4.3. Peptides and Other Substances
5. Treatment of Wastewater and Waste Gas with Macroalgae and Their Potential Applications
5.1. Wastewater Treatment with Macroalgae
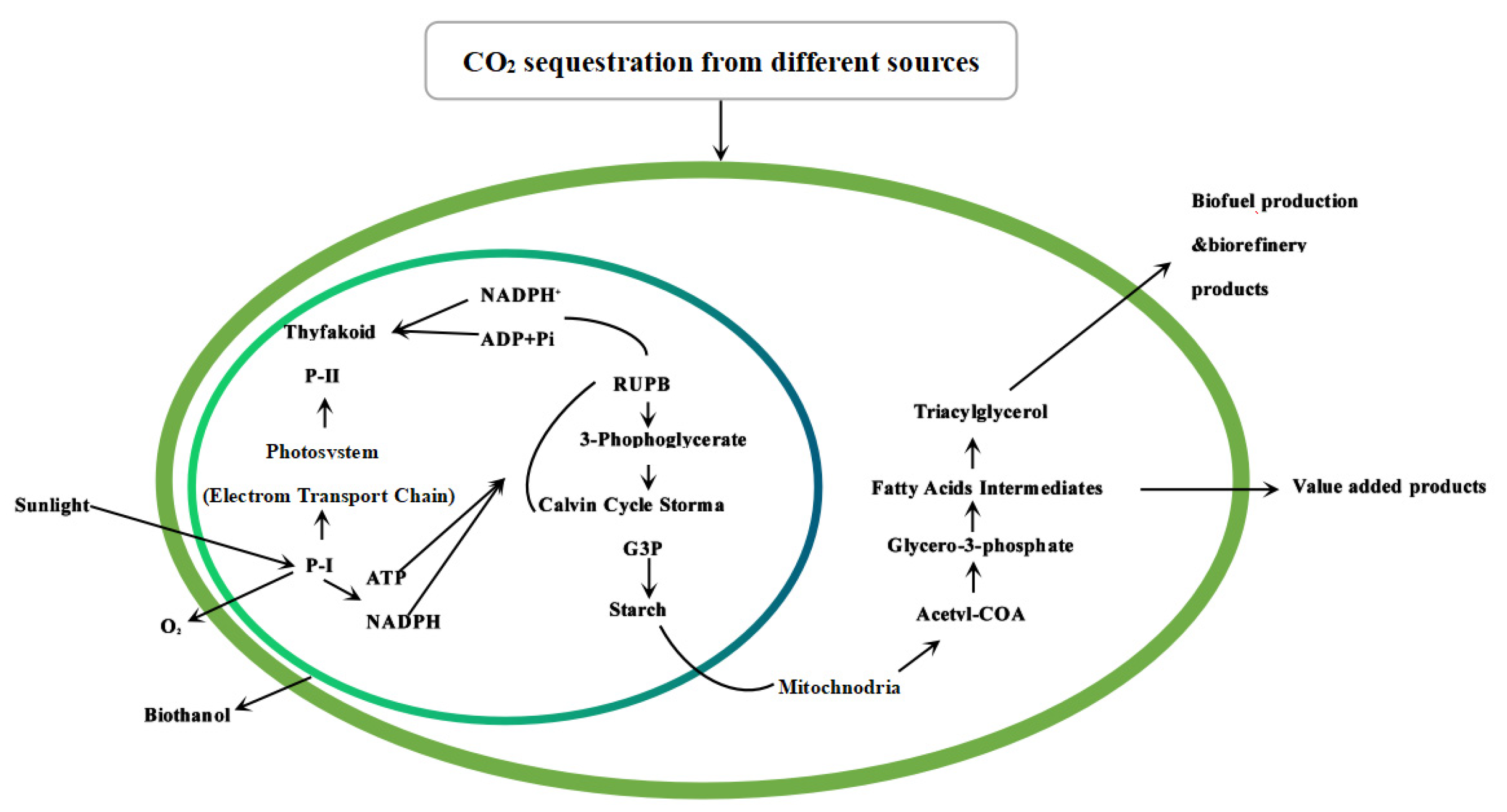
5.2. Carbon Sequestration with Macroalgae and Their Resource Utilization
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ścieszka, S.; Klewicka, E. Algae in food: A general review. Ctit. Rev. Food Sci. 2019, 59, 3538–3547. [Google Scholar] [CrossRef] [PubMed]
- Barzkar, N.; Jahromi, S.T.; Poorsaheli, H.B.; Vianello, F. Metabolites from marine microorganisms, micro, and macroalgae: Immense scope for pharmacology. Mar. Drugs 2019, 17, 464. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.T.; Fan, X.L.; Zhao, X.X.; Fu, S.F.; He, S.; Manasa, M.R.K.; Guo, R.B. Effects of organic loading rate on biogas production from macroalgae: Performance and microbial community structure. Bioresour. Technol. 2017, 235, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Bhagea, R.; Puchooa, D. Isolation and characterisation of microalgae from University of Mauritius farm for bioethanol production. Asian, J. Microbiol. Biotechnol. Environ. Sci. 2015, 17, 1065–1070. [Google Scholar]
- Marques, S.M.; Américo-Pinheiro, J.H.P. Algas como bioindicadores da qualidade da água. Rev. Científica ANAP Brasil 2017, 10, 19. [Google Scholar] [CrossRef]
- Lubchenco, J.; Haugan, P.M.; Pangestu, M.E. Five priorities for a sustainable ocean economy. Nature 2020, 588, 30–32. [Google Scholar] [CrossRef]
- Chung, I.K.; Beardall, J.; Mehta, S.; Sahoo, D.; Stojkovic, S. Using marine macroalgae for carbon sequestration:Acritical appraisal. J. Appl. Phycol. 2011, 23, 877–886. [Google Scholar] [CrossRef]
- Chopin, T.; Tacon, A.G. Importance of seaweeds and extractive species in global aquaculture production. Rev. Fish Sci. Aquac. 2021, 29, 139–148. [Google Scholar] [CrossRef]
- Charoensiddhi, S.; Lorbeer, A.J.; Franco, C.M.M.; Su, P.; Conlon, M.A.; Zhang, W. Process and economic feasibility for the production of functional food from the brown alga Ecklonia radiate. Algal Res. 2018, 29, 80–91. [Google Scholar] [CrossRef]
- Koyande, A.K.; Chew, K.W.; Manickam, S.; Chang, J.S.; Show, P.L. Emerging algal nanotechnology for high-value compounds: A direction to future food production. Trends Food Sci. Tech. 2021, 116, 290–302. [Google Scholar] [CrossRef]
- Yong, J.J.J.Y.; Chew, K.W.; Khoo, K.S.; Show, P.L.; Chang, J.S. Prospects and development of algal-bacterial biotechnology in environmental management and protection. Biotechnol. Adv. 2021, 47, 107684. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.D.; Kannah, R.Y.; Kumar, G.; Sivashanmugam, P.; Banu, J.R. A novel energetically efficient combinative microwave pretreatment for achieving profitable hydrogen production from marine macro algae (Ulva Reticul). Bioresour. Technol. 2020, 301, 122759. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Gong, J.; Lou, W.; Zou, D. Photosynthetic behaviors in response to intertidal zone and algal mat density in Ulva lactuca (Chlorophyta) along the coast of Nan’ao Island, Shantou, China. Environ. Sci. Pollut. Res. 2019, 26, 13346–13353. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Debognies, A.; Zhang, Q.; Zhang, Z.; Zhou, Z.; Zhang, J.; Qian, H. Effects of ofloxacin on the structure and function of freshwater microbial communities. Aquat. Toxicol. 2022, 244, 106084. [Google Scholar] [CrossRef]
- Betancourtt, C.; Zapata, J.; Latorre, N.; Anguita, C.; Castañeda, F.; Meynard, A.; Contreras-Porcia, L. Variación espacio-temporal en la composición del ensamble de macroalgas del intermareal rocoso de Maitencillo, Valparaíso, costa central de Chile. Rev. Biol. Mar. Oceanog. 2018, 53, 105–117. [Google Scholar] [CrossRef]
- Bertolini, C. Can secondary species maintain a primary role? Consistent inter-regional effects of understory algae on diversity. Mar. Biodivers. 2019, 49, 841–849. [Google Scholar] [CrossRef]
- Andrades, R.; Machado, F.S.; Reis-Filho, J.A.; Macieira, R.M.; Giarrizzo, T. Intertidal biogeographic subprovinces: Local and regional factors shaping fish assemblages. Front. Mar. Sci. 2018, 5, 412. [Google Scholar] [CrossRef]
- Gómez, I.; Navarro, N.P.; Huovinen, P. Bio-optical and physiological patterns in Antarctic seaweeds: A functional trait based approach to characterize vertical zonation. Prog. Oceanogr. 2019, 174, 17–27. [Google Scholar] [CrossRef]
- Wang, H.M.D.; Li, X.C.; Lee, D.J.; Chang, J.S. Potential biomedical applications of marine algae. Bioresour. Technol. 2017, 244, 1407–1415. [Google Scholar] [CrossRef]
- Son, A.R.; Park, C.S.; Park, K.R.; Kim, B.G. Amino acid digestibility in plant protein sources fed to growing pigs. Asian Austral. J. Anim. 2019, 32, 1745. [Google Scholar] [CrossRef]
- Sumayya, S.; Murugan, K. Phytochemical screening, RP-HPLC and FTIR analysis of Kappaphycus alvarezii (Doty) Doty EX PC Silva: Macro red algae. J. Pharmacogn Phytochem. 2017, 6, 325–330. [Google Scholar]
- Baghel, R.S.; Trivedi, N.; Reddy, C.R.K. A simple process for recovery of a stream of products from marine macroalgal biomass. Bioresource Technol. 2016, 203, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Francavilla, M.; Manara, P.; Kamaterou, P.; Monteleone, M.; Zabaniotou, A. Cascade approach of red macroalgae Gracilaria gracilis sustainable valorization by extraction of phycobiliproteins and pyrolysis of residue. Bioresource Technol. 2015, 184, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Masarin, F.; Cedeno, F.R.P.; Chavez, E.G.S.; de Oliveira, L.E.; Gelli, V.C.; Monti, R. Chemical analysis and biorefinery of red algae Kappa-phycus alvarezii for efficient production of glucose from residue of carrageenan extraction process. Biotechnol. Biofuels 2016, 9, 122. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, R.; Banerjee, S.; Arora, A. Recent advances in the integrated biorefinery concept for the valorization of algal biomass through sustainable routes. Biofuel Bioprod. Bior. 2021, 15, 879–898. [Google Scholar] [CrossRef]
- Trivedi, N.; Baghel, R.S.; Bothwell, J.; Gupta, V.; Reddy, C.R.K.; Lali, A.M.; Jha, B. An integrated process for the extraction of fuel and chemicals from marine macroalgal biomass. Sci. Rep. 2016, 6, 30728. [Google Scholar] [CrossRef] [PubMed]
- Gajaria, T.K.; Suthar, P.; Baghel, R.S.; Balar, N.B.; Sharnagat, P.; Mantri, V.A.; Reddy, C.R.K. Integration of protein extraction with a stream of byproducts from marine macroalgae: A model forms the basis for marine bioeconomy. Bioresource Technol. 2017, 243, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Masri, M.A.; Jurkowski, W.; Shaigani, P.; Haack, M.; Mehlmer, N.; Brück, T. A waste-free, microbial oil centered cyclic bio-refinery approach based on flexible macroalgae biomass. Appl. Energy 2018, 224, 1–12. [Google Scholar] [CrossRef]
- Mhatre, A.; Gore, S.; Mhatre, A.; Trivedi, N.; Sharma, M.; Pandit, R.; Anil, A.; Lali, A. Effect of multiple products extraction on bio-methane potential of marine macrophytic green alga Ulva lactuca. Renew Energy 2019, 132, 742–751. [Google Scholar] [CrossRef]
- Alvarado-Morales, M.; Gunnarsson, I.B.; Fotidis, I.A.; Vasilakou, E.; Lyberatos, G.; Angelidaki, I. Laminaria digitata as a potential carbon source for succinic acid and bioenergy production in a biorefinery perspective. Algal Res. 2015, 9, 126–132. [Google Scholar] [CrossRef]
- Hou, X.; Hansen, J.H.; Bjerre, A.B. Integrated bioethanol and protein production from brown seaweed Laminaria digitata. Bioresource Technol. 2015, 197, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Balboa, E.; Moure, A.; Domínguez, H. Valorization of Sargas-sum muticum biomass according to the biorefinery concept. Mar. Drugs 2015, 13, 3745–3760. [Google Scholar] [CrossRef] [PubMed]
- Ryu, H.J.; Oh, K.K. Combined de-algination process as a fractionation strategy for valorization of Brown Macroalga Saccharina japonica. Appl. Biochem. Biotech. 2017, 182, 238–249. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Chung, H.; Kim, Y.S. Effects of enzymatic modification of wheat protein on the formation of pyrazines and other volatile components in the Maillard reaction. Food Chem. 2012, 131, 1248–1254. [Google Scholar] [CrossRef]
- Laohakunjit, N.; Selamassakul, O.; Kerdchoechuen, O. Seafood-like flavour obtained from the enzymatic hydrolysis of the protein by-products of seaweed (Gracilaria sp.). Food Chem. 2014, 158, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Leandro, A.; Pereira, L.; Gonçalves, A.M. Diverse applications of marine macroalgae. Mar. Drugs. 2019, 18, 17. [Google Scholar] [CrossRef]
- Cikoš, A.M.; Jerković, I.; Molnar, M.; Šubarić, D.; Jokić, S. New trends for macroalgal natural products applications. Nat. Pro. Res. 2021, 35, 1180–1191. [Google Scholar] [CrossRef]
- Khalil, H.P.S.; Lai, T.K.; Tye, Y.Y.; Rizal, S.; Chong, E.W.N.; Yap, S.W.; Paridah, M.T. A review of extractions of seaweed hydrocolloids: Properties and applications. Express Polym. Lett. 2018, 12, 296–317. [Google Scholar] [CrossRef]
- Kączkowski, J. Mikrobiologiczny rozkład alkaloidów tropanowych [Microbiological decomposition of tropane alkaloids]. Act. Soc. Bot. Pol. 2017. [Google Scholar] [CrossRef]
- Pereira, L.; Van, D.V.F. Portuguese carrageenophytes: Carrageenan composition and geographic distribution of eight species (Gigartinales, Rhodophyta). Carbohyde Polym. 2011, 84, 614–623. [Google Scholar] [CrossRef]
- Dobrinčić, A.; Balbino, S.; Zorić, Z.; Pedisić, S.; Bursać Kovačević, D.; Elez Garofulić, I.; Dragović-Uzelac, V. Advanced technologies for the extraction of marine brown algal polysaccharides. Mar. Drugs 2020, 18, 168. [Google Scholar] [CrossRef] [PubMed]
- Alba, K.; Kontogiorgos, V. Seaweed polysaccharides (agar, alginate carrageenan). In Encyclopedia of Food Chemistry; Elsevier: Amsterdam, The Netherlands, 2018; pp. 240–250. [Google Scholar]
- Polat, S.; Trif, M.; Rusu, A.; Šimat, V.; Čagalj, M.; Alak, G.; Özogul, F. Recent advances in industrial applications of seaweeds. Crit. Rev. Food Sci. 2021, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Roohinejad, S.; Koubaa, M.; Barba, F.J.; Saljoughian, S.; Amid, M.; Greiner, R. Application of seaweeds to develop new food products with enhanced shelf-life, quality and health-related beneficial properties. Food Res. Int. 2017, 99, 1066–1083. [Google Scholar] [CrossRef] [PubMed]
- Leandro, A.; Pacheco, D.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M. Seaweed’s bioactive candidate compounds to food industry and global food security. Life 2020, 10, 140. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.A.; Félix, R.; Pais, A.C.; Rocha, S.M.; Silvestre, A.J. The quest for phenolic compounds from macroalgae: A review of extraction and identification methodologies. Biomolecules 2019, 9, 847. [Google Scholar] [CrossRef]
- Xue, M.; Ji, X.; Xue, C.; Liang, H.; Ge, Y.; He, X.; Zhang, L. Caspase-dependent and caspase-independent induction of apoptosis in breast cancer by fucoidan via the PI3K/AKT/GSK3β pathway in vivo and in vitro. Biom. Pharmacother. 2017, 94, 898–908. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K.; Saeid, A. Plant growth biostimulants, dietary feed supplements and cosmetics formulated with supercritical CO2 algal extracts. Molecules 2017, 22, 66. [Google Scholar] [CrossRef]
- Wang, F.; Shen, H.; Liu, T.; Yang, X.; Yang, Y.; Guo, Y. Formation of pyrazines in Maillard model systems: Effects of structures of lysine-containing dipeptides/tripeptides. Foods 2021, 10, 273. [Google Scholar] [CrossRef]
- Kulshreshtha, G.; Hincke, M.T.; Prithiviraj, B.; Critchley, A. A review of the varied uses of macroalgae as dietary supplements in selected poultry with special reference to laying hen and broiler chickens. J. Mar. Sci. Eng. 2020, 8, 536. [Google Scholar] [CrossRef]
- Gumus, R.E.C.E.P.; Gelen, S.U.; Koseoglu, S.; Ozkanlar, S.; Ceylan, Z.G.; Imik, H. The effects of fucoxanthin dietary inclusion on the growth performance, antioxidant metabolism and meat quality of broilers. Braz. J. Poultry Sci. 2018, 20, 487–496. [Google Scholar] [CrossRef]
- Mandal, A.B.; Biswas, A.; Mir, N.A.; Tyagi, P.K.; Kapil, D.; Biswas, A.K. Effects of dietary supplementation of Kappaphycus alvarezii on productive performance and egg quality traits of laying hens. J. Appl. Phycol. 2019, 31, 2065–2072. [Google Scholar] [CrossRef]
- Angell, A.R.; Angell, S.F.; de Nys, R.; Paul, N.A. Seaweed as a protein source for mono-gastric livestock. Trends Food Sci. Tech. 2016, 54, 74–84. [Google Scholar] [CrossRef]
- Wan, A.H.; Davies, S.J.; Soler-Vila, A.; Fitzgerald, R.; Johnson, M.P. Macroalgae as a sustainable aquafeed ingredient. Rev. Aquacult. 2019, 11, 458–492. [Google Scholar] [CrossRef]
- Morshedi, V.; Nafisi Bahabadi, M.; Sotoudeh, E.; Azodi, M.; Hafezieh, M. Nutritional evaluation of Gracilaria pulvinata as partial substitute with fish meal in practical diets of barramundi (Lates calcarifer). J. Appl. Phycol. 2018, 30, 619–628. [Google Scholar] [CrossRef]
- Okab, A.B.; Samara, E.M.; Abdoun, K.A.; Rafay, J.; Ondruska, L.; Parkanyi, V.; Lukac, N. Effects of dietary seaweed (Ulva lactuca) supplementation on the reproductive performance of buck and doe rabbits. J. Appl. Anim. Res. 2013, 41, 347–355. [Google Scholar] [CrossRef]
- Bouwhuis, M.A.; McDonnell, M.J.; Sweeney, T.; Mukhopadhya, A.; O’Shea, C.J.; O’Doherty, J.V. Seaweed extracts and galacto-oligosaccharides improve intestinal health in pigs following Salmonella Typhimurium challenge. Animal 2017, 11, 1488–1496. [Google Scholar] [CrossRef]
- Kulshreshtha, G.; Rathgeber, B.; Stratton, G.; Thomas, N.; Evans, F.; Critchley, A.; Prithiviraj, B. Feed supplementation with red seaweeds, Chondrus crispus and Sarcodiotheca gaudichaudii, affects performance, egg quality, and gut microbiota of layer hens. Poultry Sci. 2014, 93, 2991–3001. [Google Scholar] [CrossRef]
- Valente, L.M.; Araújo, M.; Batista, S.; Peixoto, M.J.; Sousa-Pinto, I.; Brotas, V.; Rema, P. Carotenoid deposition, flesh quality and immunological response of Nile tilapia fed increasing levels of IMTA-cultivated Ulva spp. J. Appl. Phycol. 2016, 28, 691–701. [Google Scholar] [CrossRef]
- Yang, H.; Li, Z.B.; Chen, Q.; Li, W.J.; Sun, Y.Z.; Lu, J. Effect of fermented Enteromopha prolifera on the growth performance, digestive enzyme activities and serum non-specific immunity of red tilapia (Oreochromis mossambicus× Oreochromis niloticus). Aquac. Res. 2016, 47, 4024–4031. [Google Scholar] [CrossRef]
- Manivasagan, P.; Oh, J. Marine polysaccharide-based nanomaterials as a novel source of nanobiotechnological applications. Int. J. Biol. Macromol. 2016, 82, 315–327. [Google Scholar] [CrossRef]
- Thepot, V.; Campbell, A.H.; Rimmer, M.A.; Paul, N.A. Meta-analysis of the use of seaweeds and their extracts as immunostimulants for fish: A systematic review. Rev. Aquacult. 2021, 13, 907–933. [Google Scholar] [CrossRef]
- Besednova, N.N.; Andryukov, B.G.; Zaporozhets, T.S.; Kryzhanovsky, S.P.; Kuznetsova, T.A.; Fedyanina, L.N.; Zvyagintseva, T.N. Algae polyphenolic compounds and modern antibacterial strategies: Current achievements and immediate prospects. Biomedicines 2020, 8, 342. [Google Scholar] [CrossRef] [PubMed]
- Prajapati, V.D.; Maheriya, P.M.; Jani, G.K.; Solanki, H.K. Carrageenan: A natural seaweed polysaccharide and its applica-tions. Carbohyd Polym. 2014, 105, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Zhang, Q.; Qi, H.; Zhang, H.; Niu, X.; Xu, Z.; Li, Z. Degradation of porphyran from Porphyra haitanensis and theantioxidant activities of the degraded porphyrans with different molecular weight. Int. J. Biol. Macromol. 2006, 38, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Liu, Y.; Wang, J.; Wu, S.; Geng, L.; Sui, Z.; Zhang, Q. Antihypertensive effects of two novel angiotensin I-converting enzyme (ACE) inhibitory peptides from Gracilariopsis lemaneiformis (Rhodophyta) in spontaneously hypertensive rats (SHRs). Mar. Drugs 2018, 16, 299. [Google Scholar] [CrossRef]
- Sudhakar, M.P.; Kumar, B.R.; Mathimani, T.; Arunkumar, K. A review on bioenergy and bioactive compounds from microalgae and macroalgae-sustainable energy perspective. J. Clean. Prod. 2019, 228, 1320–1333. [Google Scholar] [CrossRef]
- Wang, J.; Hu, S.; Nie, S.; Yu, Q.; Xie, M. Reviews on mechanisms of in vitro antioxidant activity of polysaccharides. Oxidative Med. Cell. Longev. 2016, 2016, 5692852. [Google Scholar] [CrossRef]
- Filote, C.; Santos, S.C.; Popa, V.I.; Botelho, C.; Volf, I. Biorefinery of marine macroalgae into high-tech bioproducts: A review. Environ. Chem. Lett. 2021, 19, 969–1000. [Google Scholar] [CrossRef]
- Alagan, V.; Valsala, R.N.; Rajesh, K.D. Bioactive chemical constituent analysis, in vitro antioxidant and antimicrobial activity of whole plant methanol extracts of Ulva lactuca Linn. Br. J. Pharm. Res. 2017, 15, 1–14. [Google Scholar] [CrossRef]
- Ahmed, A.B.A.; Adel, M.; Talati, A.; Kumar, M.S.; Abdulrahim, K.; Abdulhameed, M.M. Seaweed polysaccharides and their production and applications. In Seaweed Polysaccharides: Isolation, Biological and Biomedical Applications; Elsevier: Amsterdam, The Netherlands, 2017; pp. 369–382. [Google Scholar]
- Chakraborty, K.; Joseph, D.; Praveen, N.K. Antioxidant activities and phenolic contents of three red seaweeds (Division: Rhodophyta) harvested from the Gulf of Mannar of Peninsular India. J. Food Sci. Tech. 2015, 52, 1924–1935. [Google Scholar] [CrossRef] [PubMed]
- Cotas, J.; Leandro, A.; Monteiro, P.; Pacheco, D.; Figueirinha, A.; Gonçalves, A.M.; Pereira, L. Seaweed phenolics: From extraction to applications. Mar. Drugs 2020, 18, 384. [Google Scholar] [CrossRef] [PubMed]
- Rufchaei, R.; Nedaei, S.; Hoseinifar, S.H.; Hassanpour, S.; Golshan, M.; Sayad Bourani, M. Improved growth performance, serum and mucosal immunity, haematology and antioxidant capacity in pikeperch (Sander lucioperca) using dietary water hyacinth (Eichhornia crassipes) leaf powder. Aquac. Res. 2021, 52, 2194–2204. [Google Scholar] [CrossRef]
- Zhang, Y.; Shen, Y.; Zhang, H.; Wang, L.; Zhang, H.; Qian, H.; Qi, X. Isolation, purification and identification of two antioxidant peptides from water hyacinth leaf protein hydrolysates (WHLPH). Eur. Food Res. Technol. 2018, 244, 83–96. [Google Scholar] [CrossRef]
- Sanjeewa, K.A.; Jeon, Y.J. Edible brown seaweeds: A review. J. Food Bioactives. 2018, 2, 37–50. [Google Scholar] [CrossRef]
- Kulshreshtha, G.; Burlot, A.S.; Marty, C.; Critchley, A.; Hafting, J.; Bedoux, G.; Prithiviraj, B. Enzyme-assisted extraction of bioactive material from Chondrus crispus and Codium fragile and its effect on herpes simplex virus (HSV-1). Mar. Drugs 2015, 13, 558–580. [Google Scholar] [CrossRef]
- Dassarma, B.; Nandi, D.K.; Gangopadhyay, S.; Samanta, S. Hepatoprotective effect of food preservatives (butylated hydroxyanisole, butylated hydroxytoluene) on carbon tetrachloride-induced hepatotoxicity in rat. Toxicol. Rep. 2018, 5, 31–37. [Google Scholar] [CrossRef]
- Zhang, Y.; Zeng, Y.; Men, Y.; Zhang, J.; Liu, H.; Sun, Y. Structural characterization and immunomodulatory activity of exopolysaccharides from submerged culture of Auricularia auricula-judae. Int. J. Biol. Macromol. 2018, 115, 978–984. [Google Scholar] [CrossRef]
- Zammuto, V.; Rizzo, M.G.; Spanò, A.; Spagnuolo, D.; Di Martino, A.; Morabito, M.; Gugliandolo, C. Effects of crude polysaccharides from marine macroalgae on the adhesion and biofilm formation of Pseudomonas aeruginosa and Staphylococcus aureus. Algal Res. 2022, 63, 102646. [Google Scholar] [CrossRef]
- Jing, S.; Chai, W.; Guo, G.; Zhang, X.; Dai, J.; Yan, L.J. Comparison of antioxidant and antiproliferative activity between Kunlun Chrysanthemum flowers polysaccharides (KCCP) and fraction PII separated by column chromatography. J. Chromatogr. B. 2016, 1019, 169–177. [Google Scholar] [CrossRef]
- Dedhia, N.; Marathe, S.J.; Singhal, R.S. Food polysaccharides: A review on emerging microbial sources, bioactivities, nanoformulations and safety considerations. Carbohyd. Polym. 2022, 287, 119355. [Google Scholar] [CrossRef] [PubMed]
- Ponce, N.M.; Flores, M.L.; Pujol, C.A.; Becerra, M.B.; Navarro, D.A.; Córdoba, O.; Stortz, C.A. Fucoidans from the phaeophyta Scytosiphon lomentaria: Chemical analysis and antiviral activity of the galactofucan component. Carbohyd. Res. 2019, 478, 18–24. [Google Scholar] [CrossRef]
- Han, Y.; Wu, J.; Liu, T.; Hu, Y.; Zheng, Q.; Wang, B.; Li, X. Separation, characterization and anticancer activities of a sulfated polysaccharide from Undaria pinnatifida. Int. J. Biol. Macromol. 2016, 83, 42–49. [Google Scholar] [CrossRef]
- Wen, Z.S.; Xiang, X.W.; Jin, H.X.; Guo, X.Y.; Liu, L.J.; Huang, Y.N.; Qu, Y.L. Composition and anti-inflammatory effect of polysaccharides from Sargassum horneri in RAW264. 7 macrophages. Int. J. Biol. Macromol. 2016, 88, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.Y.; Zhou, W.J.; Wang, H.; Guo, G.L.; Su, Z.X.; Pu, Y.F. Antialgal compounds with antialgal activity against the common red tide microalgae from a green algae Ulva pertusa. Ecotox. Environ. Safe. 2018, 157, 61–66. [Google Scholar] [CrossRef]
- Karadeniz, F.; Lee, S.G.; Oh, J.H.; Yu, G.H.; Jang, M.S.; Seo, Y.; Kong, C.S. Solvent-partitioned fractions from Ishige okamurae extract inhibit MMP-2 and MMP-9 activities in human fibrosarcoma cells in vitro. J. Appl. Phycol. 2018, 30, 121–127. [Google Scholar] [CrossRef]
- Foseid, L.; Natvik, I.; Devle, H.; Ekeberg, D. Identification of fatty acids in fractionated lipid extracts from Palmaria palmata, Alaria esculenta and Saccharina latissima by off-line SPE GC-MS. J. Appl. Phycol. 2020, 32, 4251–4262. [Google Scholar] [CrossRef]
- Yosief, H.O.; Sarker, M.I. Naturally Derived Fatty Acid Based Antibacterial Agents. In Conversion of Renewable Biomass into Bioproducts; American Chemical Society: Washington, DC, USA, 2021; pp. 91–117. [Google Scholar]
- Miyashita, K.; Beppu, F.; Hosokawa, M.; Liu, X.; Wang, S. Nutraceutical characteristics of the brown seaweed carotenoid fucoxanthin. Arch. Biochem. Biophys. 2020, 686, 108364. [Google Scholar] [CrossRef]
- Li, J.; Guo, C.; Wu, J. Astaxanthin in liver health and disease: A potential therapeutic agent. Drug. Des. Dev. Ther. 2020, 14, 2275. [Google Scholar] [CrossRef]
- Aoi, W.; Naito, Y.; Sakuma, K.; Kuchide, M.; Tokuda, H.; Maoka, T.; Yoshikawa, T. Astaxanthin limits exercise-induced skeletal and cardiac muscle damage in mice. Antioxid. Redox. Sign. 2003, 5, 139–144. [Google Scholar] [CrossRef]
- Cuellar-Bermudez, S.P.; Aguilar-Hernandez, I.; Cardenas-Chavez, D.L.; Ornelas-Soto, N.; Romero-Ogawa, M.A.; Parra-Saldivar, R. Extraction and purification of high-value metabolites from microalgae: Essential lipids, astaxanthin and phycobiliproteins. Microb. Biotechnol. 2015, 8, 190–209. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, S.; Singh, Y.; Khattar, J.I.S.; Singh, D.P. Phycobiliprotein production by a novel cold desert cyanobacterium Nodularia sphaerocarpa PUPCCC 420.1. J. Appl. Phycol. 2017, 29, 1819–1827. [Google Scholar] [CrossRef]
- Kong, W.; Kong, J.; Ma, J.; Lyu, H.; Feng, S.; Wang, Z.; Shen, B. Chlorella vulgaris cultivation in simulated wastewater for the biomass production, nutrients removal and CO2 fixation simultaneously. J. Environ. Manage 2021, 284, 112070. [Google Scholar] [CrossRef] [PubMed]
- Sebeia, N.; Jabli, M.; Ghith, A.; Elghoul, Y.; Alminderej, F.M. Production of cellulose from Aegagropila Linnaei macro-algae: Chemical modification, characterization and application for the bio-sorptionof cationic and anionic dyes from water. Int. J. Biol. Macromol. 2019, 135, 152–162. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Silva, L.; de Oliveira Carvalho, J.; de Sousa Bezerra, R.D.; da Silva, M.S.; Ferreira, F.J.L.; Osajima, J.A.; da Silva Filho, E.C. Potential of cellulose functionalized with carboxylic acid as biosorbent for the removal of cationic dyes in aqueous solution. Molecules 2018, 23, 743. [Google Scholar] [CrossRef] [PubMed]
- Nwoba, E.G.; Moheimani, N.R.; Ubi, B.E.; Ogbonna, J.C.; Vadiveloo, A.; Pluske, J.R.; Huisman, J.M. Macroalgae culture to treat anaerobic digestion piggery effluent (ADPE). Bioresour. Technol. 2017, 227, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.; Madill, M.; Champagne, P. Use of freshwater macroalgae Spirogyra sp. for the treatment of municipal wastewaters and biomass production for biofuel applications. Biomass. Bioenerg. 2018, 111, 213–223. [Google Scholar] [CrossRef]
- Yang, X.; Liu, L.; Yin, Z.; Wang, X.; Wang, S.; Ye, Z. Quantifying photosynthetic performance of phytoplankton based on photosynthesis–irradiance response models. Environ. Sci. Eur. 2020, 32, 24. [Google Scholar] [CrossRef]
- Singh, S.P.; Singh, P. Effect of temperature and light on the growth of algae species: A review. Rene. Sust. Energ. Rev. 2015, 50, 431–444. [Google Scholar] [CrossRef]
- Tala, F.; Penna-Díaz, M.A.; Luna-Jorquera, G.; Rothäusler, E.; Thiel, M. Daily and seasonal changes of photobiological responses in floating bull kelp Durvillaea antarctica (Chamisso) Hariot (Fucales: Phaeophyceae). Phycologia 2017, 56, 271–283. [Google Scholar] [CrossRef]




| Macroalgae | Polysaccharides | Polyphenols | Peptides | Biological Activity | Applications | References |
|---|---|---|---|---|---|---|
| Chlorophyta | Ulvan | Alkaloids | NbrbcS1-1, NbrbcS1-2 | Anti-tumour, anti-oxidant anti-thrombolytic immunal modulation, anti-influenza, and anti-coagulant | Tissue engineering | [61,62,63] |
| Rhodophyta | Carrageenan | Terpenoids | FQIN [M(O)] CILR, TGAPCR | Anti-coagulant, plateletaggregation inhibition, anti-viral, anti-tumuor activity | Anti-oxidant, drug delivery | [61,63,64] |
| Porphyran | Phenolic compounds | Degraded and untreated porphyran possesses scavenging-free radical activity and functions as a reducing power | Cytotoxic, drug delivery | [61,63,65,66] | ||
| Agar | Rutin | Extracted for gelling and stabilising capabilities, anti-inflammatory | Anti-oxidant, drug delivery | [62,63] | ||
| Phaeophyceae | Alginate | Catechol | Aβ peptides | Commercial alginate salts have immunal modulation properties | Drug delivery, anti-fungal, anti-tumour | [61,62,63] |
| Laminarin | Hesperidin | Anti-tumour, anti-inflammatory, immune-stimulatory, anti-coagulant and anti-oxidant activity | [63,67] | |||
| Fucoidan | Fucoxanthins | Anti-viral, anti-tumor, immune-stimulatory, anti-oxidant, anti-inflammatory, anti-coagulantand, anti-thrombotic activity. Ability to reduce cholesterol, triglyceride and LDL-C, andincrease HDL-C. Gastric protection (e.g., antiulcer, anti-adhesion for Helicobacter pyroli), protection against urinary tract, kidney and liver diseases. | [63,67,68] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, J.; Qi, M.; Chen, H.; Zhou, C.; Ruan, R.; Yan, X.; Cheng, P. Macroalgae-Derived Multifunctional Bioactive Substances: The Potential Applications for Food and Pharmaceuticals. Foods 2022, 11, 3455. https://doi.org/10.3390/foods11213455
Guo J, Qi M, Chen H, Zhou C, Ruan R, Yan X, Cheng P. Macroalgae-Derived Multifunctional Bioactive Substances: The Potential Applications for Food and Pharmaceuticals. Foods. 2022; 11(21):3455. https://doi.org/10.3390/foods11213455
Chicago/Turabian StyleGuo, Jiameng, Mei Qi, Hongyu Chen, Chengxu Zhou, Roger Ruan, Xiaojun Yan, and Pengfei Cheng. 2022. "Macroalgae-Derived Multifunctional Bioactive Substances: The Potential Applications for Food and Pharmaceuticals" Foods 11, no. 21: 3455. https://doi.org/10.3390/foods11213455
APA StyleGuo, J., Qi, M., Chen, H., Zhou, C., Ruan, R., Yan, X., & Cheng, P. (2022). Macroalgae-Derived Multifunctional Bioactive Substances: The Potential Applications for Food and Pharmaceuticals. Foods, 11(21), 3455. https://doi.org/10.3390/foods11213455