

Production of Corn Protein Hydrolysate with Glutamine-Rich Peptides and Its Antagonistic Function in Ulcerative Colitis In Vivo
Abstract
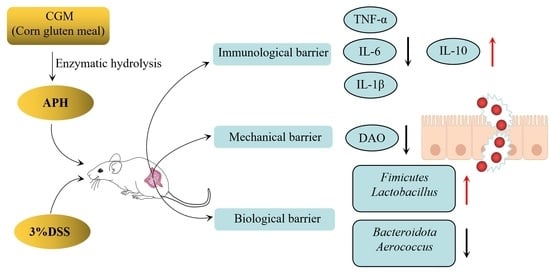
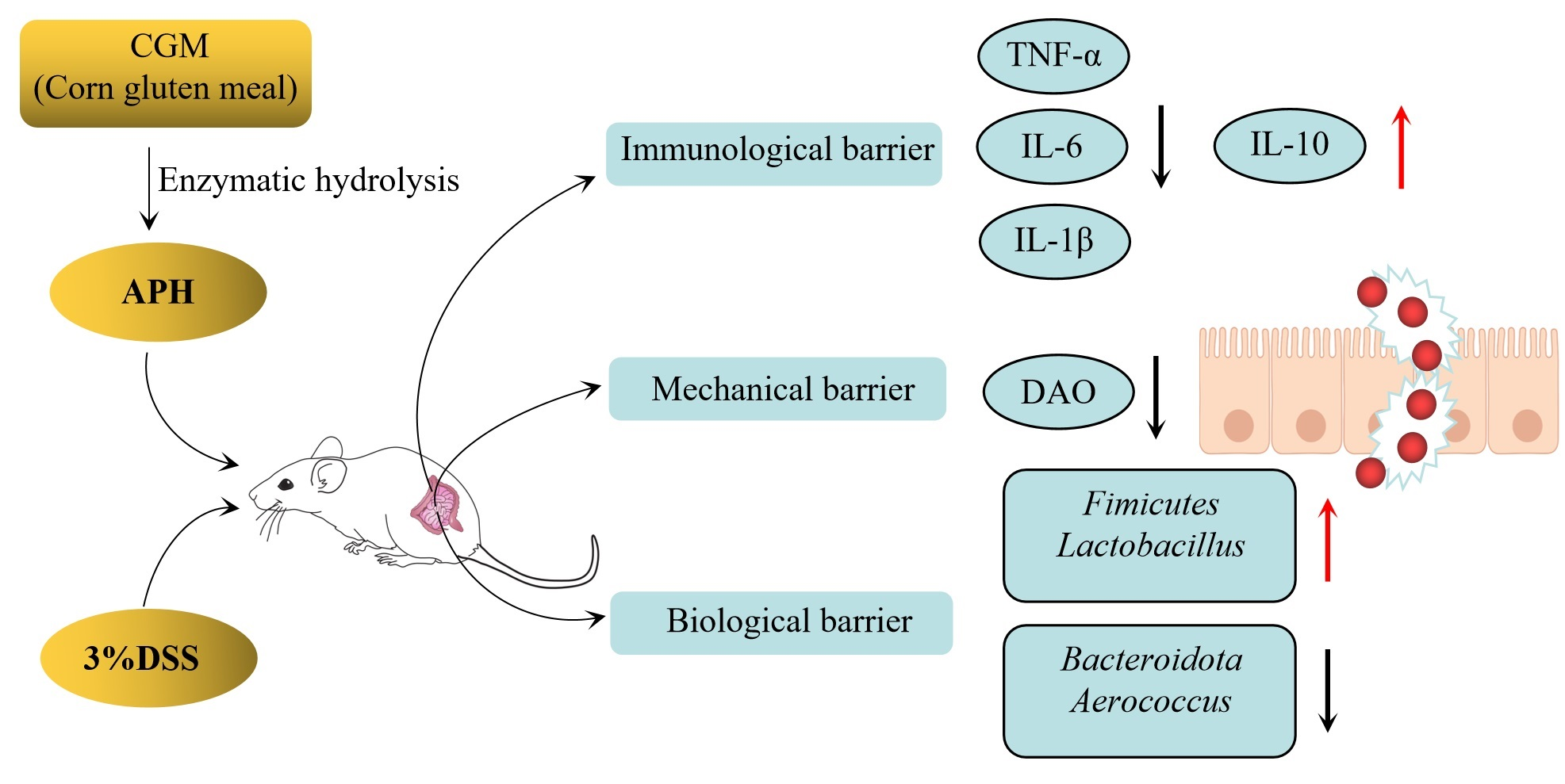
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of CGM Hydrolysates
2.3. Determination of Gln Content in Hydrolysates
2.4. Animals and Experimental Design
2.5. Histopathology
2.6. Determination of DAO, SOD, GSH-Px, and Inflammatory Cytokine Concentrations
2.7. 16S rRNA High Throughput Sequencing
2.8. Statistical Analysis
3. Results
3.1. Preparation of APH
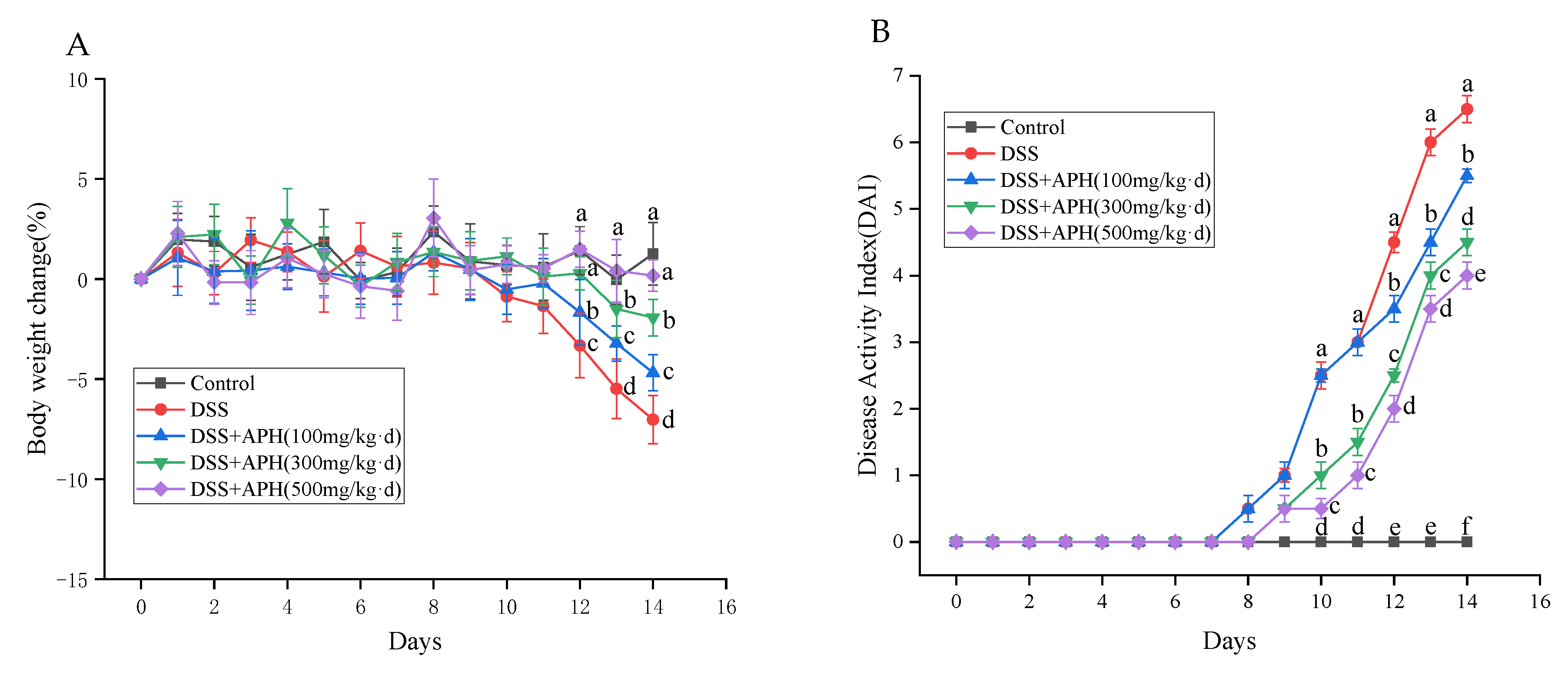
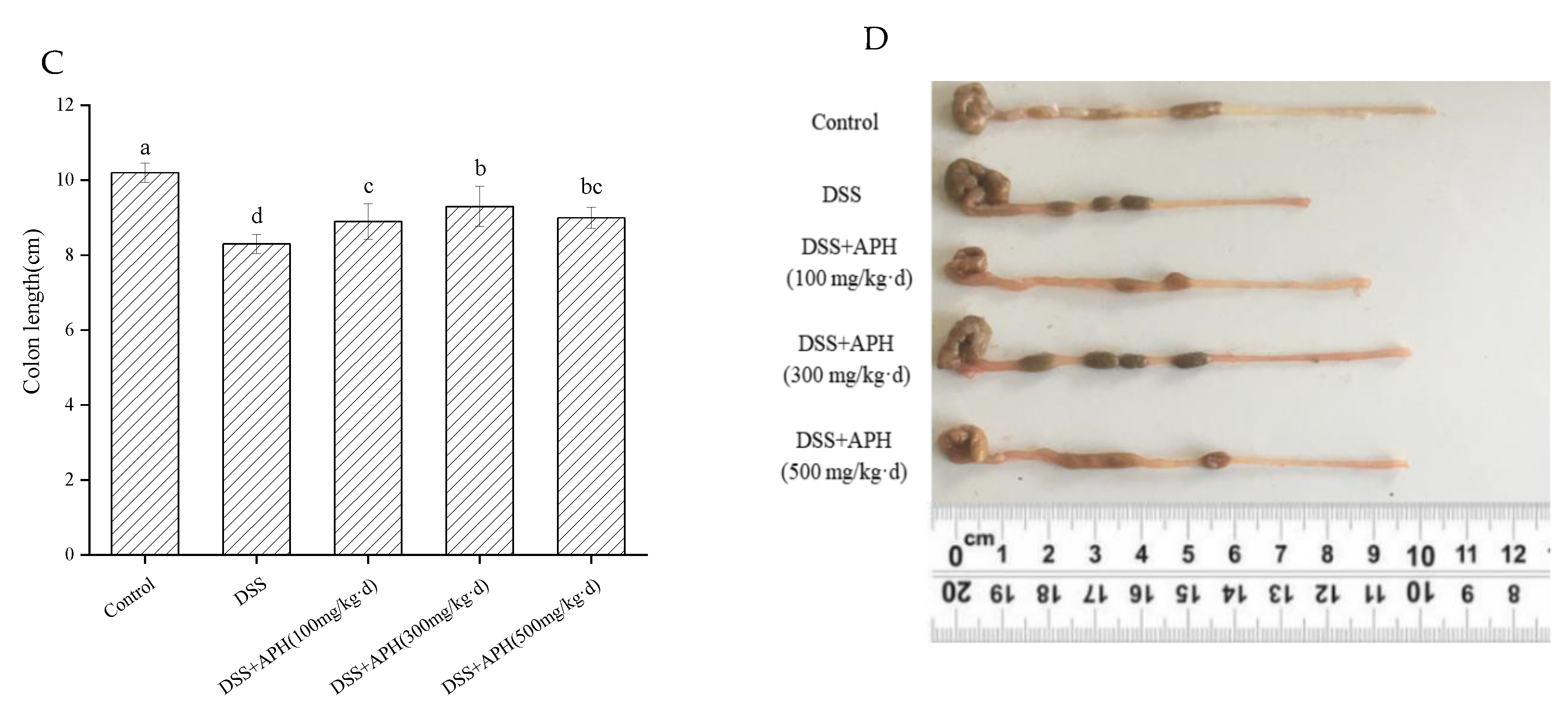
3.2. APH Alleviates Colitis Symptoms in Mice
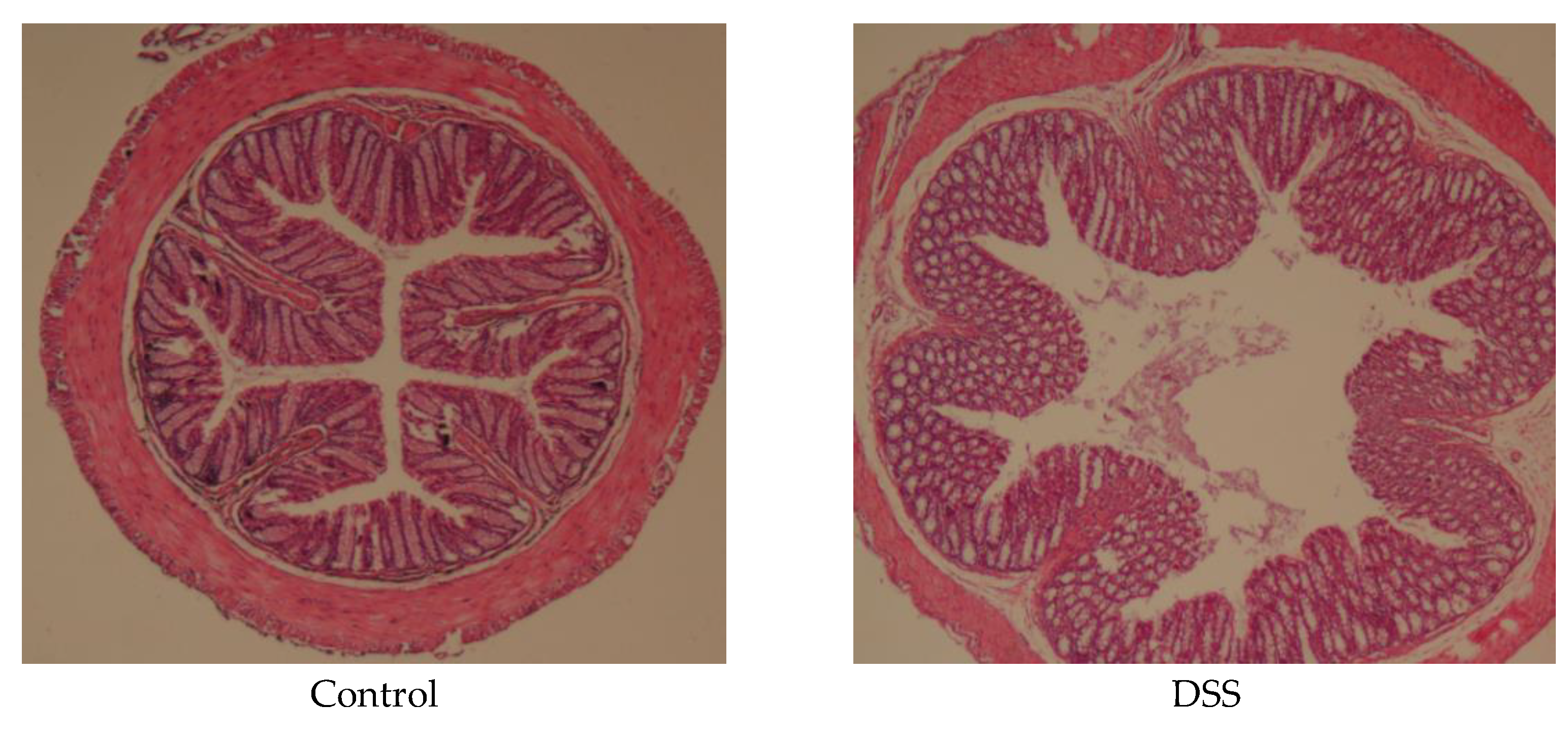
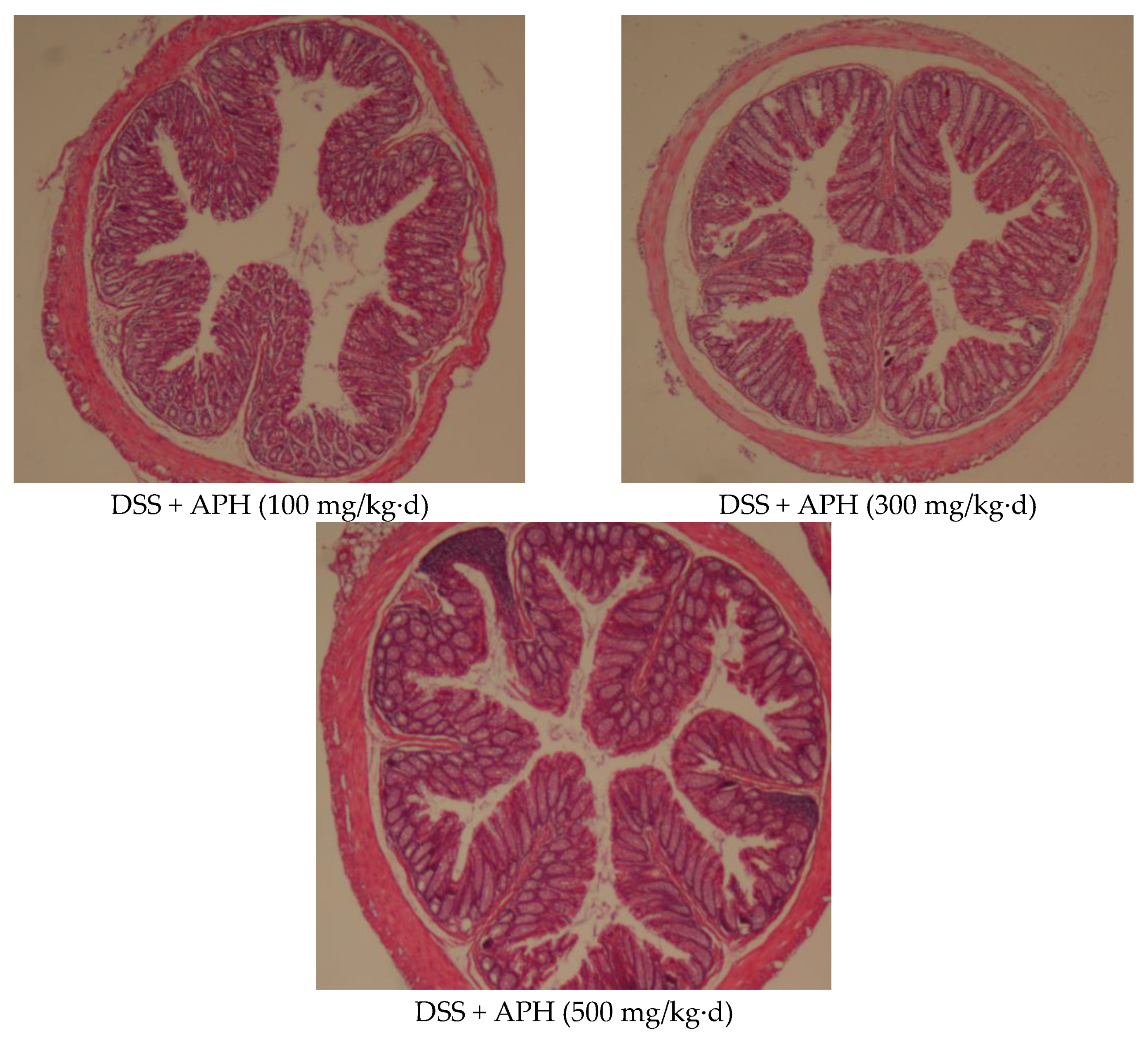
3.3. APH Alleviates Histopathological Changes in Mice
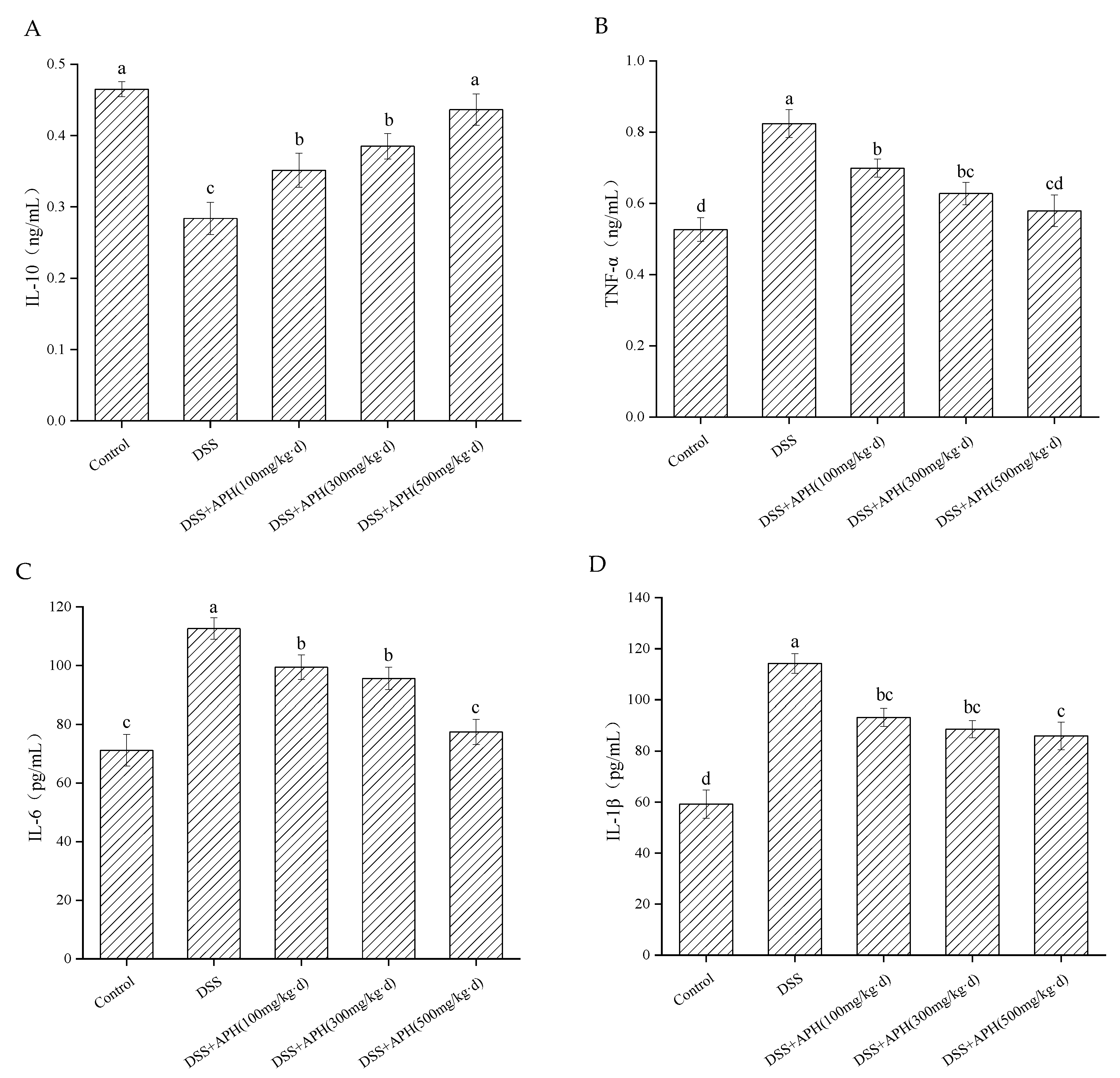
3.4. Effects of APH on the Inflammatory Cytokines in Mice
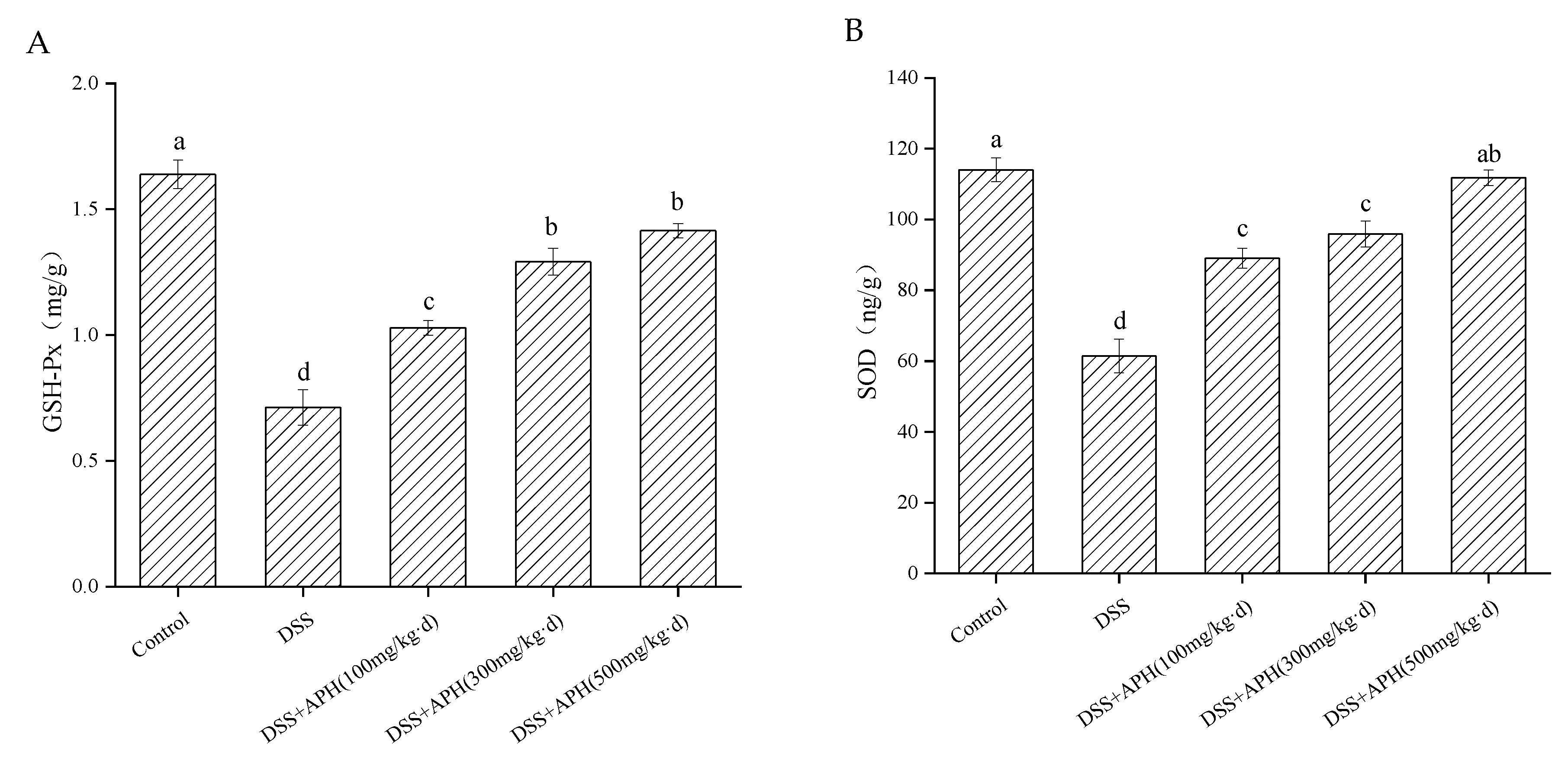
3.5. Effects of APH on Oxidative Stress-Related Indicators in Mice
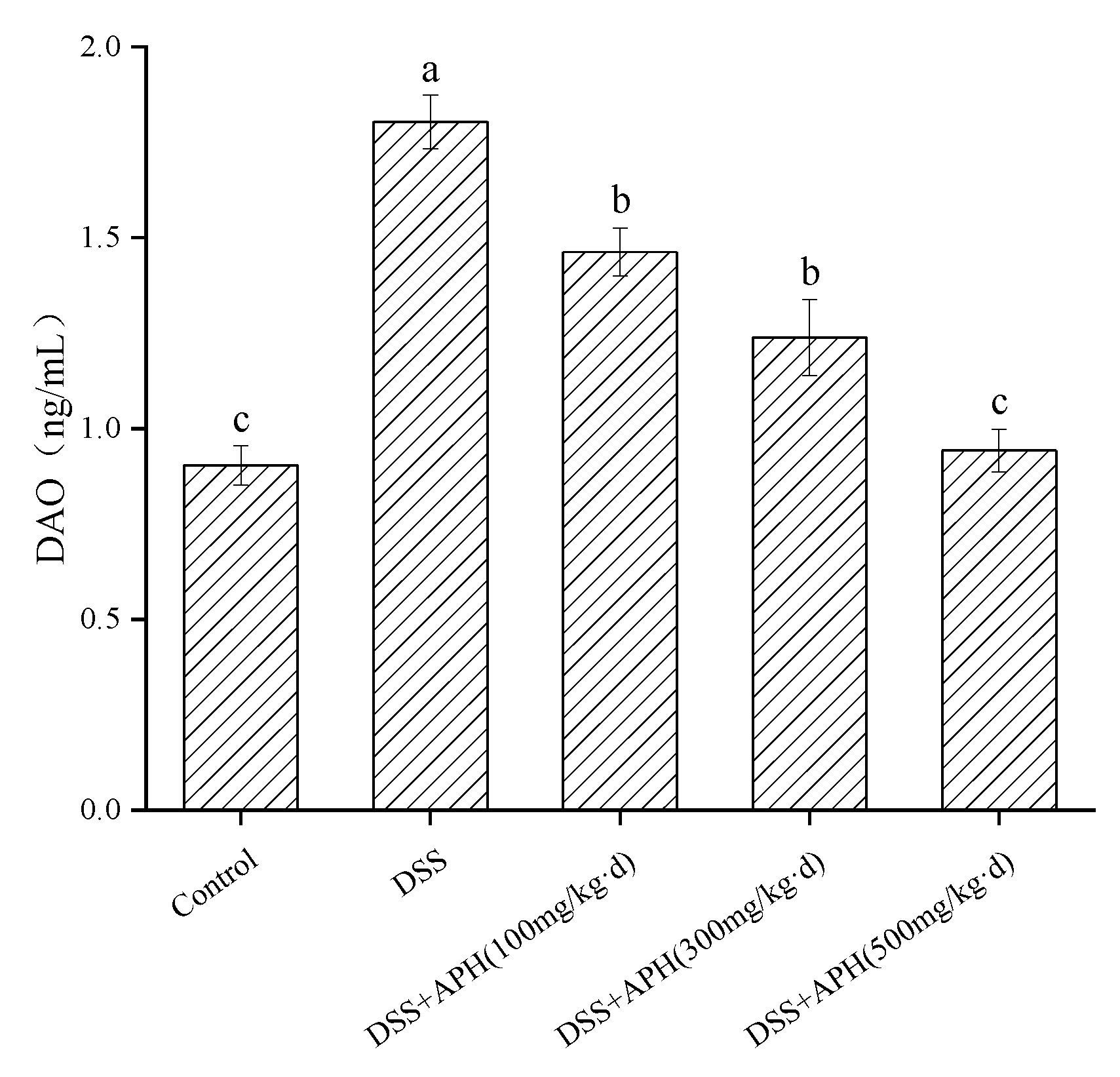
3.6. Effects of APH on Intestinal Permeability in Mice
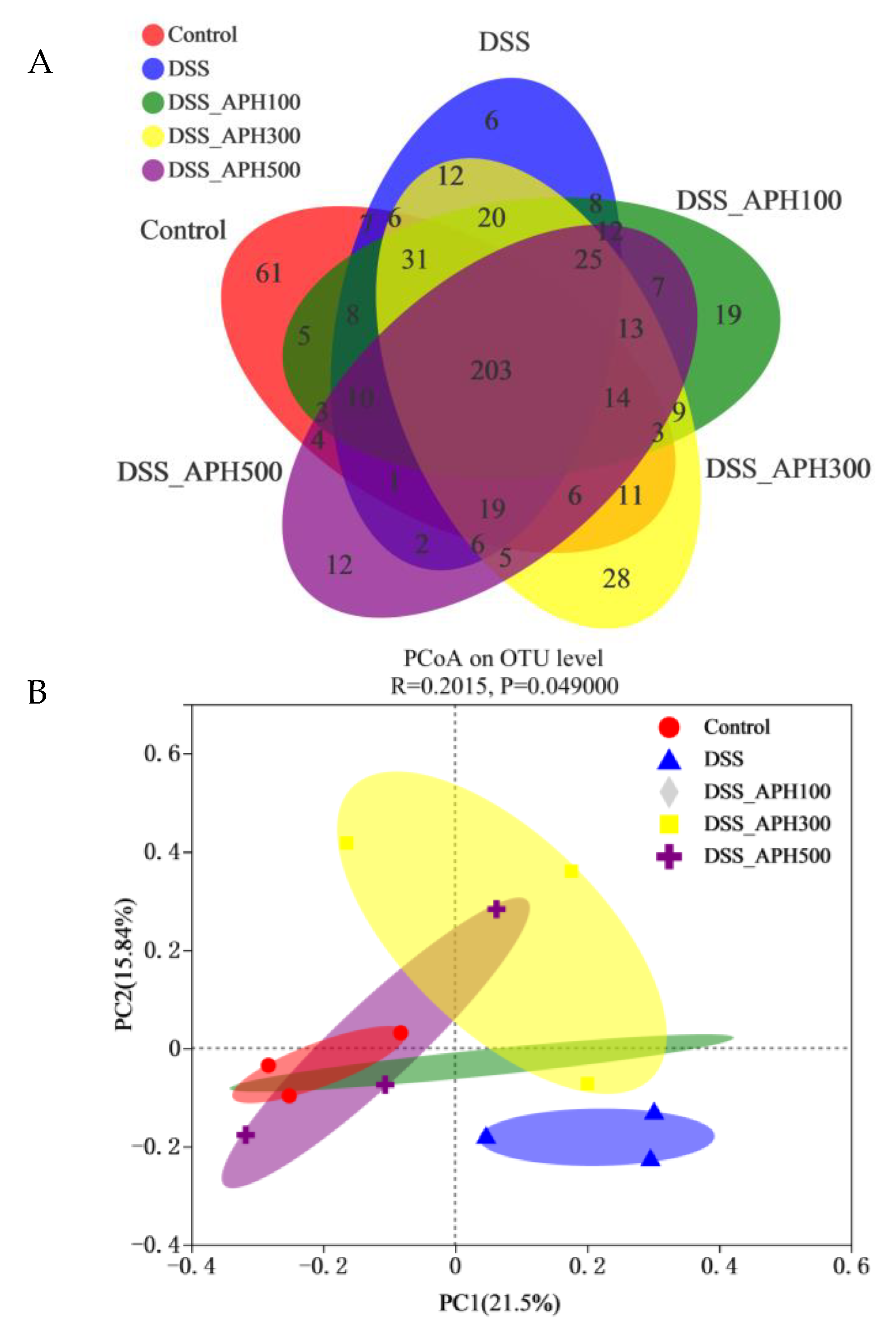
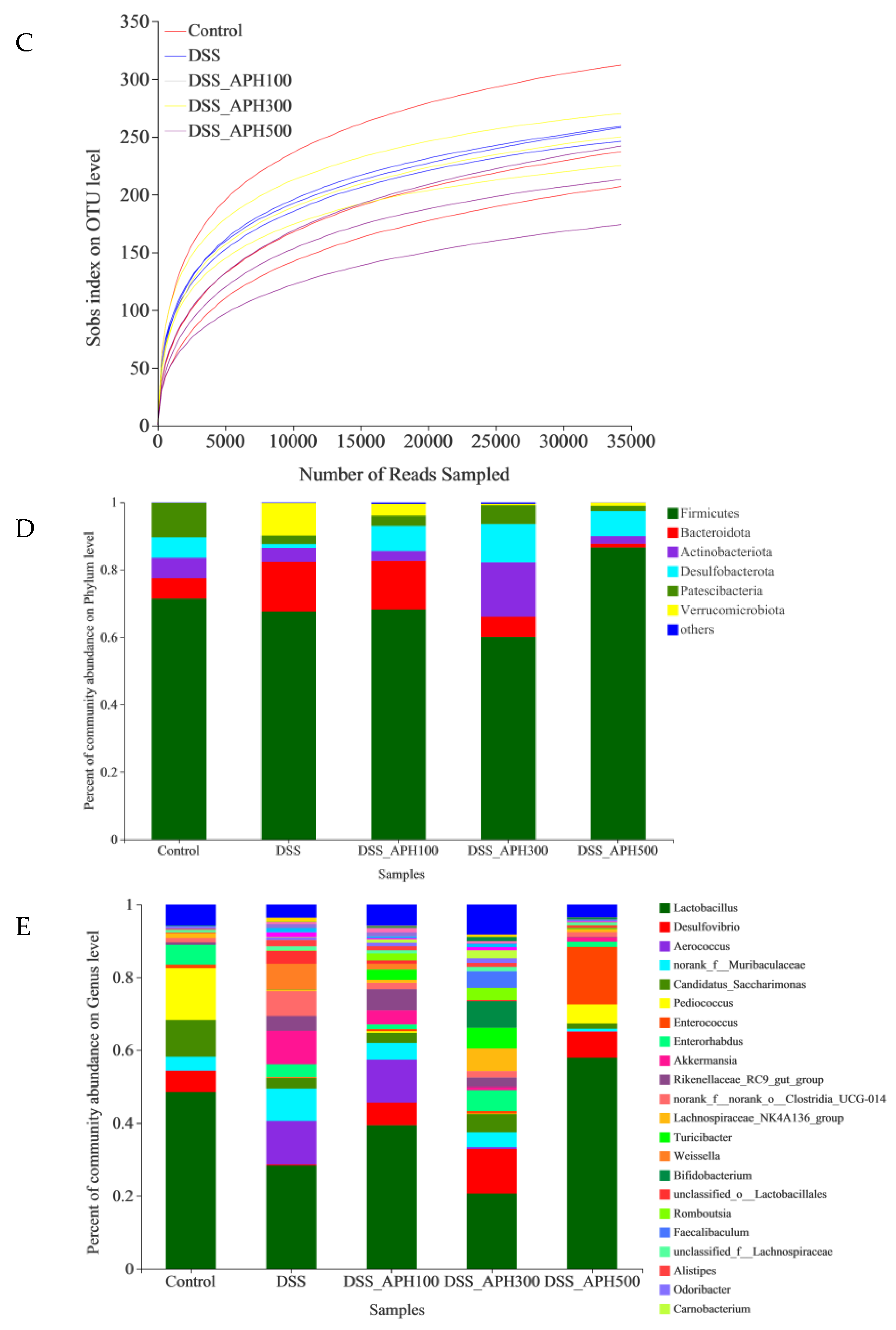
3.7. Effects of APH on Intestinal Microbiota in Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ungaro, R.; Mehandru, S.; Allen, P.B.; Biroulet, P.L.; Colombel, J.F. Ulcerative colitis. Lancet 2017, 389, 1756–1770. [Google Scholar] [CrossRef]
- Mak, W.Y.; Zhao, M.; Ng, S.C.; Burisch, J. The epidemiology of inflammatory bowel disease: East meets west. J. Gastroenterol. Hepatol. 2020, 35, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Philipsen, L.; Reddycherla, A.V.; Hartig, R.; Gumz, J.; Kästle, M.; Kritikos, A.; Poltorak, M.P.; Prokazov, Y.; Turbin, E. De novo phosphorylation and conform -ational opening of the tyrosine kinase Lck act in concert to initiate T cell receptor signaling. Sci. Signal. 2017, 10, eaaf4736. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.C.; Li, S.; Cao, H.Y.; Shen, P.; Liu, J.X.; Fu, Y.H.; Cao, Y.G.; Zhang, N.S. The protective role of phloretin against dextran sulfate sodium-induced ulcerative colitis in mice. Food Funct. 2019, 10, 422–431. [Google Scholar] [CrossRef]
- Krupoves, A.; Mack, D.R.; Seidman, E.G.; Deslandres, C.; Bucionis, V.; Amre, D.K. Immediate and long-term outcomes of corticosteroid therapy in pediatric Crohn’s disease patients. Inflamm. Bowel Dis. 2011, 17, 954–962. [Google Scholar] [CrossRef]
- Mir, F.A.; Juboori, A.; Bragg, J.D.; Tahan, V. Autoimmune hemolytic anemia associated with infliximab infusion in ulcerative colitis. North. Clin. Istanb. 2018, 5, 64–66. [Google Scholar]
- Lu, Y.J.; Wang, J.; Soladoye, O.P.; Aluko, R.E.; Fu, Y.; Zhang, Y.H. Preparation, receptors, bioactivity and bioavailability of γ-glutamyl peptides: A comprehensive review. Trends Food Sci. Technol. 2021, 113, 301–314. [Google Scholar] [CrossRef]
- Wang, B.; Wu, G.Y.; Zhou, Z.G.; Dai, Z.L.; Sun, Y.L. Glutamine and intestinal barrier function. Amino Acids 2014, 47, 2143–2154. [Google Scholar] [CrossRef]
- Tian, J.; He, G.; Mai, K.S.; Liu, C.D.; Zhou, H.H.; Wen, H. Dietary Ala-Gln ameliorated growth suppression and intestinal injury induced by soya saponin in zebrafish. Aquaculture 2020, 529, 735748. [Google Scholar] [CrossRef]
- Tan, B.; Liu, H.L.; He, G.Z.; Xiao, H.; Xiao, D.F.; Liu, Y.H.; Wu, J.P.; Fang, J.; Yin, Y.L. Alanyl-glutamine but not glycyl-glutamine improved the proliferation of enterocytes as glutamine substitution in vitro. Amino Acids 2017, 49, 2023–2031. [Google Scholar] [CrossRef]
- Gioia, L.D.; Cuq, B.; Guilbert, S. Effect of hydrophilic plasticizers on thermomechanical properties of corn gluten meal. Cereal Chem. 1998, 75, 514–519. [Google Scholar] [CrossRef]
- Wang, X.J.; Liu, X.L.; Zheng, X.Q.; Qu, Y.; Shi, Y.G. Preparation of corn glycopeptides and evaluation of their antagonistic effects on alcohol-induced liver injury in rats. J. Funct. Foods 2020, 66, 103776. [Google Scholar] [CrossRef]
- Guo, D.J.; Zhang, Y.; Zhao, J.J.; He, H.; Hou, T. Selenium-biofortified corn peptides: Attenuating concanavalin A-induced liver injury and structure characterization. J. Trace Elem. Med. Biol. 2019, 51, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Liao, W.; Qi, X.; Yu, W.L.; Wu, J.P. Identification of immunomodulatory peptides from zein hydrolysates. Eur. Food Res. Technol. 2020, 246, 931–937. [Google Scholar] [CrossRef]
- Zheng, X.Q.; Liu, X.L.; Wang, X.J.; Lin, J.; Li, D. Production of hydrolysate with antioxidative activity by enzymatic hydrolysis of extruded corn gluten. Appl. Microbiol. Biot. 2006, 73, 763–770. [Google Scholar] [CrossRef]
- Liu, X.L.; Zheng, X.Q.; Song, Z.L.; Liu, X.F.; Kopparapu, N.K.; Wang, X.J.; Zheng, Y.J. Preparation of enzymatic pretreated corn gluten meal hydrolysate and in vivo evaluation of its antioxidant activity. J. Funct. Foods 2015, 18, 1147–1157. [Google Scholar] [CrossRef]
- Katharina, S.K.; Peter, S.; Peter, F. Quantitative analyses of glutamine in peptides and proteins. J. Agric. Food Chem. 1996, 44, 1808–1811. [Google Scholar]
- Chen, G.; Ran, X.; Li, B.; Li, Y.H.; He, D.W.; Huang, B.H.; Fu, S.P.; Liu, J.X.; Wang, W. Sodium Butyrate Inhibits Inflammation and Maintains Epithelium Barrier Integrity in a TNBS-induced Inflammatory Bowel Disease Mice Model. EBioMedicine 2018, 30, 317–325. [Google Scholar] [CrossRef]
- Song, W.; Li, Y.; Zhang, X.; Wang, Z. Potent anti-inflammatory activity of polysac charides extracted from Blidingia minima and their effect in a mouse model of inflammatory bowel disease. J. Funct. Foods 2019, 61, 103494. [Google Scholar] [CrossRef]
- Higaki-Sato, N.; Sato, K.; Esumi, Y.; Okumura, T.; Yoshikawa, H.; Tanaka-Kuwajima, C.; Kurata, A.; Ohtsuki, K. Isolation and Identification of Indigestible Pyroglutamyl Peptides in an Enzymatic Hydrolysate of Wheat Gluten Prepared on an Industrial Scale. J. Agric. Food Chem. 2003, 51, 8–13. [Google Scholar] [CrossRef]
- Mannheim, A.; Cheryan, M. Water soluble zein by enzymatic modifi-cation in organic solvents. Cereal Chem. 1993, 70, 115–121. [Google Scholar]
- Roth, E. Nonnutritive Effects of Glutamine. J. Nutr. 2008, 138, 2025S–2031S. [Google Scholar] [CrossRef]
- Zhou, N.; Yao, Y.; Wu, N.; Du, H.Y.; Xu, M.S.; Zhao, Y.; Tu, Y.G. VF-4 and DR-8 Derived from Salted Egg White Inhibit Inflammatory Activity via NF-κB/PI3K-Akt/MAPK Signal Transduction Pathways in HT-29 Cells Induced by TNF-α. Mol. Nutr. Food Res. 2022, 66, 2100682. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Cui, Y.; Liu, P.; Zhao, Y.; Wang, L.; Liu, Y.; Xie, J. Small Peptides Isolated from Enzymatic Hydrolyzate of Fermented Soybean Meal Promote Endothelium-Independent Vasorelaxation and ACE Inhibition. J. Agric. Food Chem. 2017, 65, 10844–10850. [Google Scholar] [CrossRef]
- Ogata, H.; Hibi, T. Cytokine and anti-cytokine therapies for inflammatory bowel disease. Curr. Pharm. Des. 2003, 9, 1107–1113. [Google Scholar] [CrossRef]
- Mudter, J.; Neurath, M.F. IL-6 signaling in inflammatory bowel disease: Pathophysiological role and clinical relevance. Inflamm. Bowel Dis. 2007, 13, 1016–1023. [Google Scholar] [CrossRef]
- Billiet, T.; Rutgeerts, P.; Ferrante, M.; Assche, G.V.; Vermeire, S. Targeting TNF-α for the treatment of inflammatory bowel disease. Expert Opin. Biol. Ther. 2014, 14, 75–101. [Google Scholar]
- Puleston, J.; Cooper, M.; Murch, S. A distinct subset of chemokines dominates the mucosal chemokine response in inflammatory bowel disease. Aliment. Pharm. Ther. 2005, 21, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.Z.; Zhang, S.Y.; Yuan, L.Y.; Yang, Y.Y.; Lin, S.Y. Glutamine and methionine targeted pulsed electric field treatment for enhanced immune activity in pine nut Gln-Trp-Phe-Met peptides. Int. J. Food Sci. Tech. 2020, 55, 2954–2961. [Google Scholar] [CrossRef]
- Fillmann, H.; Kretzmann, N.A.; San-Miguel, B.; Llesuy, S.; Marroni, N.; González-Gallego, J.; Tunón, M.J. Glutamine inhibits over-expression of pro-inflammatory genes and down-regulates the nuclear factor kappaB pathway in an experimental model of colitis in the rat. Toxicology 2007, 236, 217–226. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Zhao, Y.; Yao, Y.; Xu, M.S.; Du, H.Y.; Wu, N.; Tu, Y.G. Isolation and identification of peptides from simulated gastrointestinal digestion of preserved egg white and their anti-inflammatory activity in TNF-α-induced Caco-2 cells. J. Nutr. Biochem. 2019, 63, 44–53. [Google Scholar] [CrossRef]
- Guha, S.; Majumder, K. Structural-features of food-derived bioactive peptides with anti-inflammatory activity: A brief review. J. Food Biochem. 2018, 43, e12531. [Google Scholar] [CrossRef] [PubMed]
- Chalamaiah, M.; Yu, W.L.; Wu, J.P. Immunomodulatory and anticancer protein hydrolysates (peptides) from food proteins: A review. Food Chem. 2018, 245, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Marcela, G.M.; Blanca, H.L.; Jose, M.S.; Rosalva, M.E.; Cristina, M.V. Peptides derived from in vitro gastrointestinal digestion of germinated soybean proteins inhibit human colon cancer cells proliferation and inflammation. Food Chem. 2018, 242, 75–82. [Google Scholar]
- Pravda, J. Radical induction theory of ulcerative colitis. World J. Gastroenterol. 2005, 11, 2371–2384. [Google Scholar] [CrossRef]
- Tanida, S.; Mizoshita, T.; Mizushima, T. Involvement of oxidative stress and mucosal addressin cell adhesion molecule-1 (MAdCAM-1) in inflammatory bowel disease. J. Clin. Biochem. Nutr. 2011, 48, 112–116. [Google Scholar] [CrossRef]
- Naito, Y.J.; Takagi, T.; Yoshikawa, T. Neutrophil-Dependent Oxidative Stress in Ulcerative Colitis. J. Clin. Biochem. Nutr. 2007, 41, 18–26. [Google Scholar] [CrossRef]
- Liang, Q.F.; Chalamaiah, M.; Ren, X.F.; Ma, H.L.; Wu, J.P. Identification of New Anti-inflammatory Peptides from Zein Hydrolysate after Simulated Gastrointestinal Digestion and Transport in Caco-2 Cells. J. Agric. Food Chem. 2018, 66, 1114–1120. [Google Scholar] [CrossRef]
- Cekanaviciute, E.; Yoo, B.B.; Runia, T.F. Gut bacteria from multiple sclerosis patients modulate human T cells and exacerbate symptoms in mouse models. Natl. Acad. Sci. 2017, 114, 10713–10718. [Google Scholar] [CrossRef]
- Bedoui, S.; Velkoska, E.; Bozinovski, S.; Jones, J.E.; Anderson, G.P.; Morris, M.J. Unaltered TNF-alpha production by macrophages and monocytes in diet-induced obesity in the rat. J. Inflamm. 2005, 2, 2. [Google Scholar] [CrossRef][Green Version]
- Zhang, H.L.; Wang, Y.; Su, Y.C.; Fang, X.D.; Guo, W.J. The alleviating effect and mechanism of Bilobalide on ulcerative colitis. Food Funct. 2021, 12, 6226–6239. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.K.; Han, Y.H.; Gu, M.; Song, M.Y.; Wu, X.; Li, Z.Z.; Li, F.; Goulette, T.; Xiao, H. Dietary cranberry suppressed colonic inflammation and alleviated gut microbiota dysbiosis in dextran sodium sulfate-treated mice. Food Funct. 2019, 10, 6331–6341. [Google Scholar] [CrossRef]
- Liu, Z.J.; Liu, F.; Wang, W.; Sun, C.B.; Gao, D.; Ma, J.G.; Hussain, M.A.; Xu, C.; Jiang, Z.M.; Hou, J.C. Study of the alleviation effects of a combination of Lactobacillus rhamnosus and inulin on mice with colitis. Food Funct. 2020, 11, 3823–3837. [Google Scholar] [CrossRef] [PubMed]
- Powell, N.; Walker, A.W.; Stolarczyk, E.; Canavan, J. The transcription factor T-bet regulates intestinal inflammation mediated by interleukin-7 receptor innate lymphoid cells. Immunity 2012, 37, 674–684. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.A.; Yu, H.; Albesiano, E. A sensitive stool diagnostic assay to study enterotoxigenic bacteroides fragilis in inflammatory bowel disease and colitis-associated cancer. Gastroenterology 2011, 140, S360. [Google Scholar] [CrossRef]
- Liu, J.H.; He, Z.Y.; Ma, N.; Chen, Z.Y. Beneficial Effects of Dietary Polyphenols on High-Fat Diet-Induced Obesity Linking with Modulation of Gut Microbiota. J. Agric. Food Chem. 2020, 68, 33–47. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Weight Loss (%) | Stool Consistency | Occult/Gross Bleeding |
|---|---|---|---|
| 0 | 0 | Normal | Normal |
| 1 | 1–5 | - | - |
| 2 | 6–10 | Loose | Hemoccult positive |
| 3 | 11–15 | - | - |
| 4 | >16 | Diarrhea | Gross bleeding |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, Y.; Liu, X.; Wang, J.; Ma, Y.; Zheng, X. Production of Corn Protein Hydrolysate with Glutamine-Rich Peptides and Its Antagonistic Function in Ulcerative Colitis In Vivo. Foods 2022, 11, 3359. https://doi.org/10.3390/foods11213359
Jing Y, Liu X, Wang J, Ma Y, Zheng X. Production of Corn Protein Hydrolysate with Glutamine-Rich Peptides and Its Antagonistic Function in Ulcerative Colitis In Vivo. Foods. 2022; 11(21):3359. https://doi.org/10.3390/foods11213359
Chicago/Turabian StyleJing, Yan, Xiaolan Liu, Jinyu Wang, Yongqiang Ma, and Xiqun Zheng. 2022. "Production of Corn Protein Hydrolysate with Glutamine-Rich Peptides and Its Antagonistic Function in Ulcerative Colitis In Vivo" Foods 11, no. 21: 3359. https://doi.org/10.3390/foods11213359
APA StyleJing, Y., Liu, X., Wang, J., Ma, Y., & Zheng, X. (2022). Production of Corn Protein Hydrolysate with Glutamine-Rich Peptides and Its Antagonistic Function in Ulcerative Colitis In Vivo. Foods, 11(21), 3359. https://doi.org/10.3390/foods11213359