
Effect of Autochthonous Lactic Acid Bacteria-Enhanced Fermentation on the Quality of Suancai


Abstract

1. Introduction
2. Materials and Methods
2.1. Suancai Preparation and Sampling
2.2. Physical and Chemical Analysis
2.3. DNA Extraction and High-Throughput Sequencing
2.4. Sequence Analysis
2.5. GC-MS Analysis
2.6. Free Amino Acid Analysis
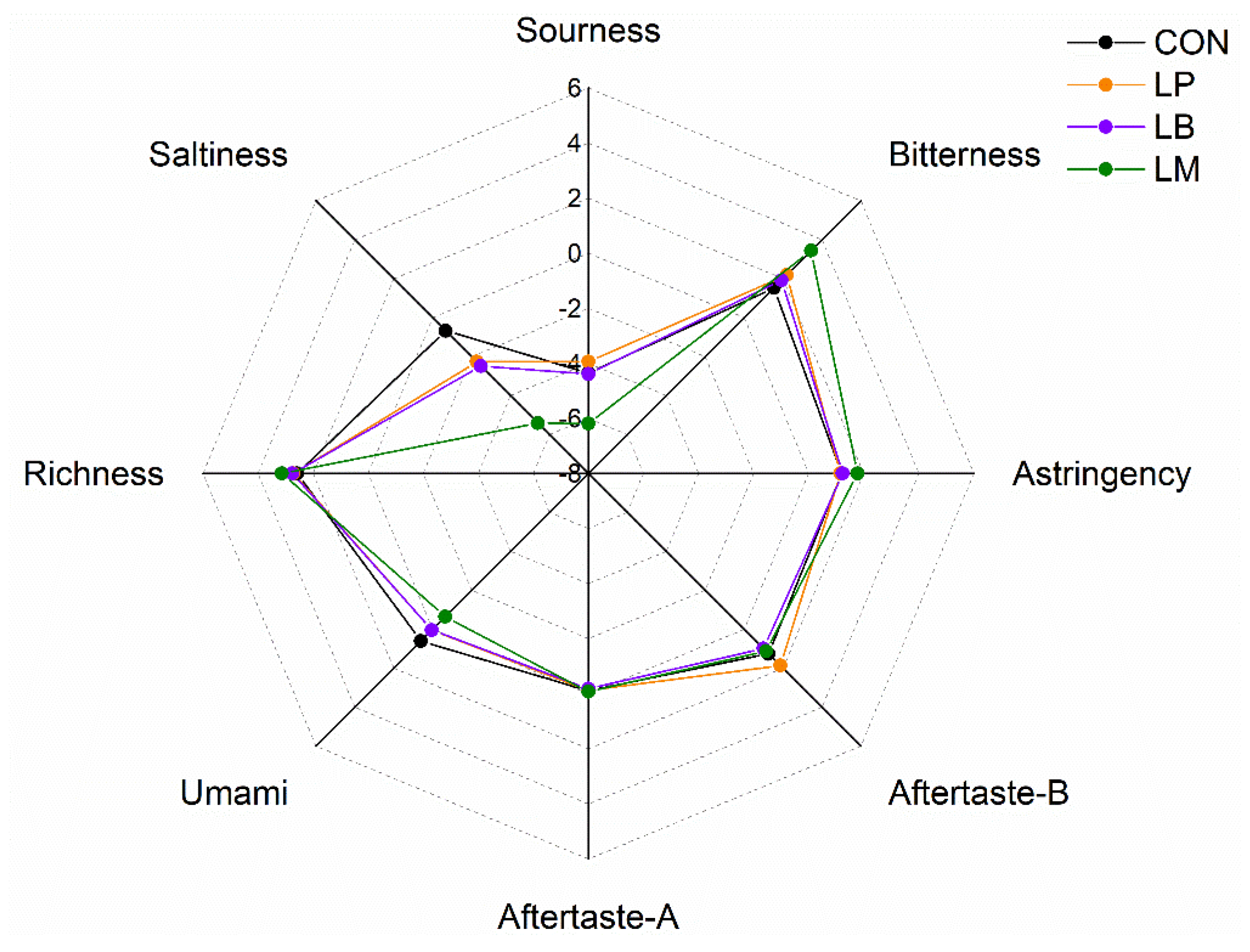
2.7. E-Tongue Analysis
2.8. Statistical Analysis
3. Results and Discussion
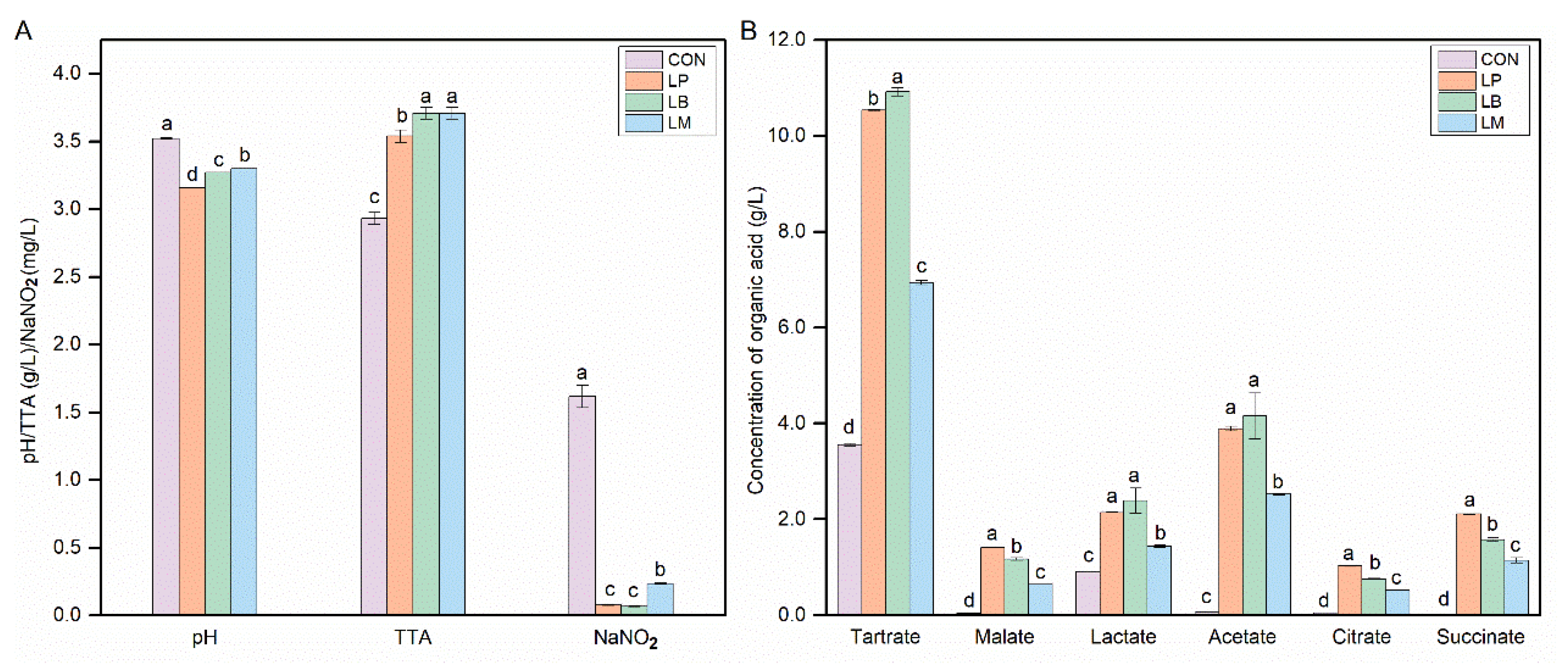
3.1. The Physicochemical Properties in Suancai with Different Starters
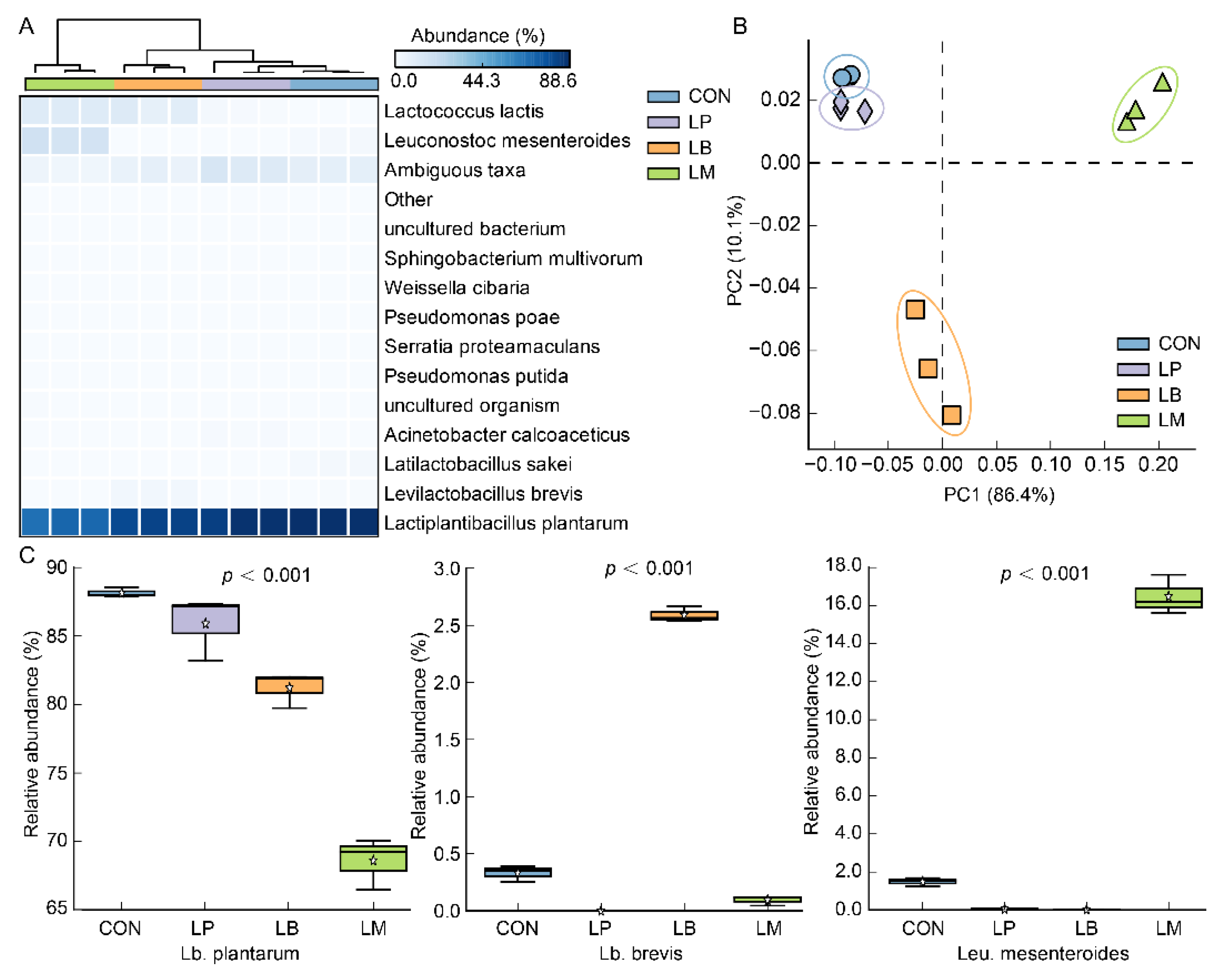
3.2. The Diversity of Microbial Community in Suancai with Different Starters
3.3. The Composition of the Bacterial Community in Suancai with Different Starters
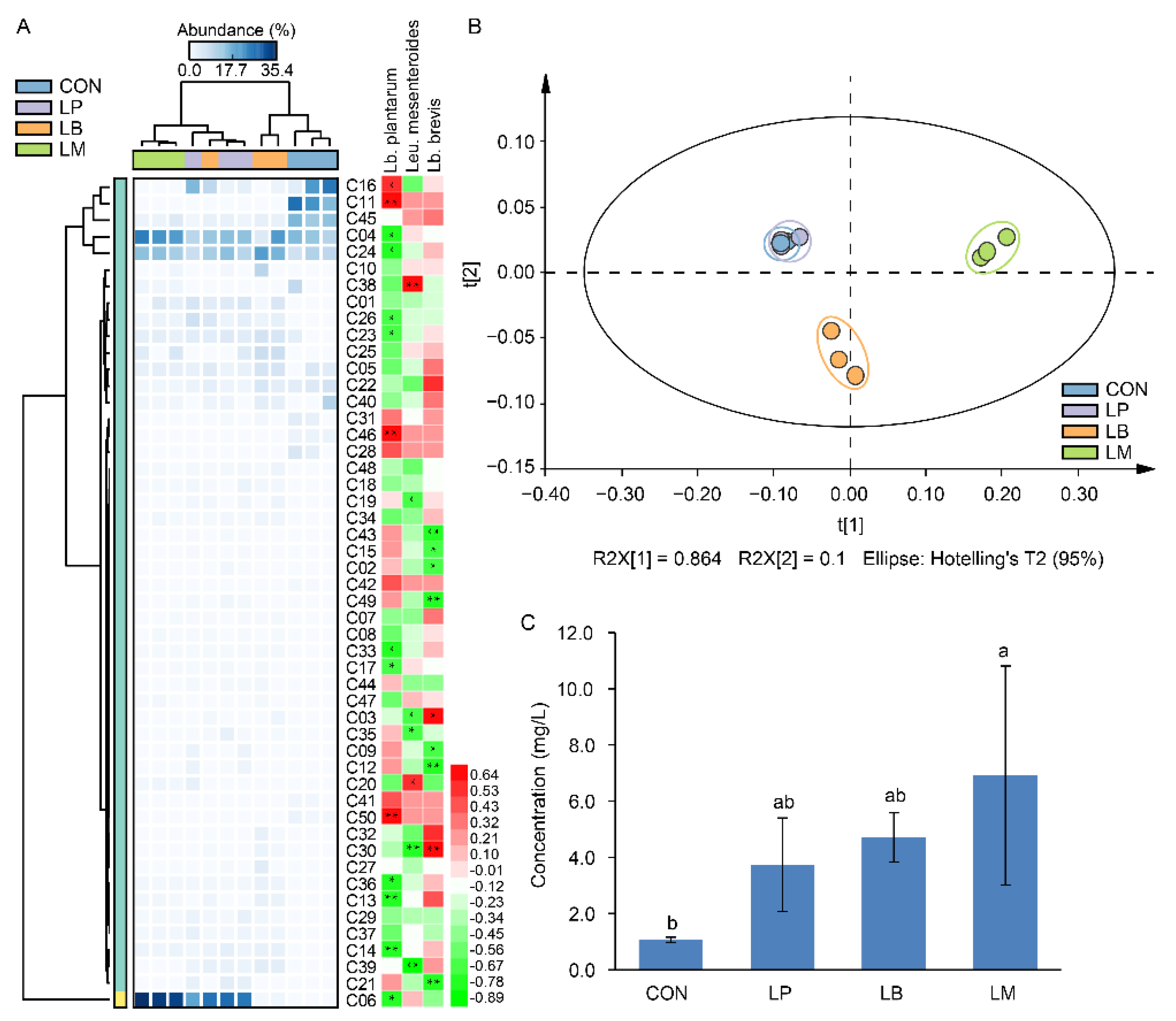
3.4. Volatile Compound Analysis in Suancai with Different Starters
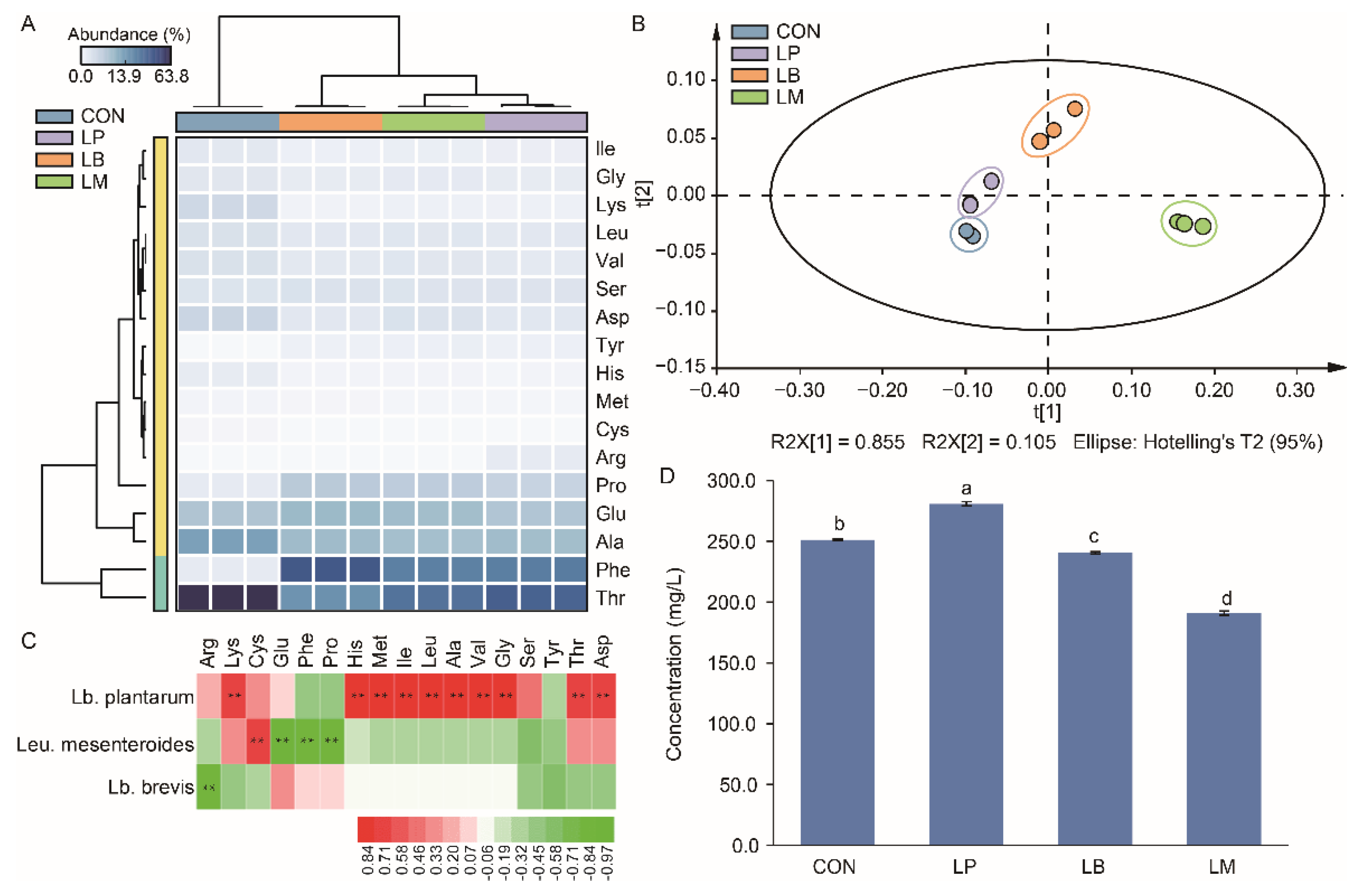
3.5. Changes of the Free Amino Acid in Suancai with Different Starters
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liang, H.; He, Z.; Wang, X.; Song, G.; Chen, H.; Lin, X.; Ji, C.; Zhang, S. Bacterial profiles and volatile flavor compounds in commercial Suancai with varying salt concentration from Northeastern China. Food Res. Int. 2020, 137, 109384. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Huang, T.; Huang, C.; Jim, H.; Peng, Z.; Xie, M.; Xiong, T. The microbial communities and flavour compounds of Jiangxi yancai, Sichuan paocai and Dongbei Suancai: Three major types of traditional Chinese fermented vegetables. LWT—Food Sci. Technol. 2020, 121, 108865. [Google Scholar] [CrossRef]
- Satora, P.; Skotniczny, M.; Strnad, S.; Piechowicz, W. Chemical composition and sensory quality of sauerkraut produced from different cabbage varieties. LWT—Food Sci. Technol. 2021, 136, 110325. [Google Scholar] [CrossRef]
- Liang, H.; He, Z.; Wang, X.; Song, G.; Chen, H.; Lin, X.; Ji, C.; Li, S. Effects of salt concentration on microbial diversity and volatile compounds during Suancai fermentation. Food Microbiol. 2020, 91, 103537. [Google Scholar] [CrossRef]
- He, Z.; Wang, X.; Chen, H.; Lin, X.; Ji, C.; Li, S.; Liang, H. Effects of different temperatures on bacterial diversity and volatile flavor compounds during the fermentation of Suancai, a traditional fermented vegetable food from northeastern China. LWT—Food Sci. Technol. 2020, 118, 108773. [Google Scholar] [CrossRef]
- Park, S.E.; Seo, S.H.; Kim, E.J.; Na, C.S.; Son, H.S. Effects of different fermentation temperatures on metabolites of Kimchi. Food Biosci. 2018, 23, 100–106. [Google Scholar] [CrossRef]
- Guan, Q.Q.; Huang, T.; Peng, F.; Huang, J.Q.; Liu, Z.G.; Peng, Z.; Xie, M.Y.; Xiong, T. The microbial succession and their correlation with the dynamics of flavor compounds involved in the natural fermentation of suansun, a traditional Chinese fermented bamboo shoots. Food Res. Int. 2022, 157, 111216. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.D.; Chen, G.; Tang, Y.; Ming, J.Y.; Huang, R.Q.; Li, J.Y.; Ye, M.Z.; Fan, Z.Y.; Chi, Y.L.; Zhang, Q.S.; et al. Study of bacterial community succession and reconstruction of the core lactic acid bacteria to enhance the flavor of paocai. Int. J. Food Microbiol. 2022, 375, 109702. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Xiong, T.; Peng, Z.; Liu, C.; Huang, T.; Yu, H.; Xie, M. Correlation between microbiota and flavours in fermentation of Chinese Sichuan Paocai. Food Res. Int. 2018, 114, 123–132. [Google Scholar] [CrossRef]
- Du, R.; Song, G.; Zhao, D.; Sun, J.; Ping, W.; Ge, J. Lactobacillus casei starter culture improves vitamin content, increases acidity and decreases nitrite concentration during sauerkraut fermentation. Int. J. Food Sci. Tech. 2018, 53, 1925–1931. [Google Scholar] [CrossRef]
- Song, G.; He, Z.; Wang, X.; Zhao, M.; Cao, X.; Lin, X.; Ji, C.; Zhang, S.; Liang, H. Improving the quality of Suancai by inoculating with Lactobacillus plantarum and Pediococcus pentosaceus. Food Res. Int. 2021, 148, 110581. [Google Scholar] [CrossRef]
- Han, Y.Q.; Wang, C.; Zhang, X.L.; Li, X.; Gao, Y. Characteristic volatiles analysis of Dongbei Suancai across different fermentation stages based on HS-GC-IMS with PCA. J. Food Sci. 2022, 87, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Hu, W.; Xiu, Z.; Jiang, A.; Yang, X.; Sarengaowa; Ji, Y.; Guan, Y.; Feng, K. Comparison of northeast sauerkraut fermentation between single lactic acid bacteria strains and traditional fermentation. Food Res. Int. 2020, 137, 109553. [Google Scholar] [CrossRef]
- Xiao, M.; Peng, Z.; Liu, C.; Huang, T.; Yu, H.; Xie, M. Exploring the typical flavours formation by combined with metatranscriptomics and metabolomics during Chinese Sichuan paocai fermentation. LWT—Food Sci. Technol. 2022, 153, 112474. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Reyon, D.; Tsai, S.Q.; Khayter, C.; Foden, J.A.; Sander, J.D.; Joung, J.K. FLASH assembly of TALENs for high-throughput genome editing. Nat. Biotechnol. 2012, 30, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.X.; Yan, X.; Shang, H.; Ji, C.F.; Zhang, S.F.; Liang, H.P.; Chen, Y.X.; Lin, X.P. Screening of Lactiplantibacillus plantarum with high stress tolerance and high esterase activity and their effect on promoting protein metabolism and flavor formation in Suanzhayu, a Chinese fermented fish. Foods 2022, 11, 1932. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, G.; Shen, W.; Wang, Y.; Zhang, W.; Chi, Y. Microbial safety and sensory quality of instant low-salt Chinese paocai. Food Control 2016, 59, 575–580. [Google Scholar] [CrossRef]
- Endo, A.; Maeno, S.; Liu, S.Q. Lactic acid bacteria: Leuconostoc spp. In Encyclopedia of Dairy Sciences, 3rd ed.; McSweeney, P.L.H., McNamara, J.P., Eds.; Academic Press: Oxford, USA, 2022; pp. 226–232. [Google Scholar] [CrossRef]
- Huang, Y.; Jia, X.; Yu, J.; Chen, Y.; Liu, D.; Liang, M. Effect of different lactic acid bacteria on nitrite degradation, volatile profiles, and sensory quality in Chinese traditional paocai. LWT—Food Sci. Technol. 2021, 147, 111597. [Google Scholar] [CrossRef]
- Zhu, Y.; Guo, L.; Yang, Q. Partial replacement of nitrite with a novel probiotic Lactobacillus plantarum on nitrate, color, biogenic amines and gel properties of Chinese fermented sausages. Food Res. Int. 2020, 137, 109351. [Google Scholar] [CrossRef]
- Yang, Y.; Fan, Y.; Li, T.; Yang, Y.; Zeng, F.; Wang, H.; Suo, H.; Song, J.; Zhang, Y. Microbial composition and correlation between microbiota and quality-related physiochemical characteristics in chongqing radish paocai. Food Chem. 2022, 369, 130897. [Google Scholar] [CrossRef]
- Liang, H.; Chen, H.; Zhang, W.; Yu, C.; Ji, C.; Lin, X. Investigation on microbial diversity of industrial Zhacai paocai during fermentation using high-throughput sequencing and their functional characterization. LWT—Food Sci. Technol. 2018, 91, 460–466. [Google Scholar] [CrossRef]
- Liu, Z.G.; Xiao, M.Y.; Xu, Y.Z.; Li, D.Y.; Zhu, W.H.; Huang, T.; Peng, F.; Guan, Q.Q.; Peng, Z.; Xie, M.Y.; et al. Effect of homo- and hetero-fermentative lactic acid bacteria on physicochemical properties, amino acid, and volatile flavor compounds during paocai fermentation by pure culture. J. Food Process Pres. 2022, e17052. [Google Scholar] [CrossRef]
- Wang, D.; Chen, G.; Tang, Y.; Li, H.; Shen, W.; Wang, M.; Liu, S.; Qin, W.; Zhang, Q. Effects of temperature on paocai bacterial succession revealed by culture-dependent and culture-independent methods. Int. J. Food Microbiol. 2020, 317, 108463. [Google Scholar] [CrossRef]
- Abiad, M.G.; Ajjour, C.; Kassaify, Z.; Iskandar, C.F.; Hamzeh, R.; Hassan, H.F. Preliminary physiological, phenotypic, and probiotic characterization of Lactobacillus strains isolated from Anbaris-traditional Lebanese fermented dairy product. Int. J. Food Prop. 2022, 25, 1266–1278. [Google Scholar] [CrossRef]
- Torres, S.; Verón, H.; Contreras, L.; Isla, M.I. An overview of plant-autochthonous microorganisms and fermented vegetable foods. Food Sci. Hum. Well. 2020, 9, 112–123. [Google Scholar] [CrossRef]
- Hashemi, S.M.B.; Jafarpour, D. Fermentation of bergamot juice with Lactobacillus plantarum strains in pure and mixed fermentations: Chemical composition, antioxidant activity and sensorial properties. LWT—Food Sci. Technol. 2020, 131, 109803. [Google Scholar] [CrossRef]
- Wu, Y.; Li, S.; Tao, Y.; Li, D.; Han, Y.; Show, P.L.; Wen, G.; Zhou, J. Fermentation of blueberry and blackberry juices using Lactobacillus plantarum, Streptococcus thermophilus and Bifidobacterium bifidum: Growth of probiotics, metabolism of phenolics, antioxidant capacity in vitro and sensory evaluation. Food Chem. 2021, 348, 129083. [Google Scholar] [CrossRef]
- Cavanagh, D.; Fitzgerald, G.F.; McAuliffe, O. From field to fermentation: The origins of Lactococcus lactis and its domestication to the dairy environment. Food Microbiol. 2015, 47, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Sabatini, N.; Mucciarella, M.R.; Marsilio, V. Volatile compounds in uninoculated and inoculated table olives with Lactobacillus plantarum (Olea europea L., cv. Moresca and Kalamata). LWT—Food Sci. Technol. 2008, 41, 2017–2022. [Google Scholar] [CrossRef]
- Agerbirk, N.; Olsen, C.E. Glucosinolate structures in evolution. Phytochemistry 2012, 77, 16–45. [Google Scholar] [CrossRef] [PubMed]
- Dan, T.; Chen, H.; Li, T.; Tian, J.; Ren, W.; Zhang, H.; Sun, T. Influence of Lactobacillus plantarum P-8 on fermented milk favor and storage stability. Front. Microbiol. 2019, 9, 3133. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Zhang, C.H.; Liu, F.S.; Jin, Z.Y.; Xia, X.L. Ecological succession and functional characteristics of lactic acid bacteria in traditional fermented foods. Crit. Rev. Food Sci. 2022. [Google Scholar] [CrossRef]
- Liu, M.; Nauta, A.; Francke, C.; Siezen, R. Comparative genomics of enzymes in flavor-forming pathways from amino acids in lactic acid bacteria. Appl. Environ. Microb. 2008, 74, 4590–4600. [Google Scholar] [CrossRef]
- Freiding, S.; Ehrmann, M.A.; Vogel, R.F. Comparison of different IlvE aminotransferases in Lactobacillus sakei and investigation of their contribution to aroma formation from branched chain amino acids. Food Microbiol. 2012, 29, 205–214. [Google Scholar] [CrossRef]
- Pan, X.; Zhang, S.; Xu, X.; Lao, F.; Wu, J. Volatile and non-volatile profiles in jujube pulp co-fermented with lactic acid bacteria. LWT—Food Sci. Technol. 2022, 154, 112772. [Google Scholar] [CrossRef]
- Zhao, D.; Tang, J.; Ding, X. Correlation between flavour compounds and sensory properties of Potherb Mustard (Brassica juncea, Coss.) pickle. Food Sci. Technol. Int. 2007, 13, 423–435. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples 1 | CON | LP | LB | LM |
|---|---|---|---|---|
| Chao1 | 20.61 ± 5.29 a | 34.60 ± 2.45 a | 29.38 ± 4.20 a | 31.70 ± 13.17 a |
| Goods coverage | 0.99 ± 0.00 a | 0.99 ± 0.00 a | 0.99 ± 0.00 a | 0.99 ± 0.00 a |
| Shannon | 0.75 ± 0.03 c | 1.00 ± 0.11 b | 1.11 ± 0.04 b | 1.48 ± 0.09 a |
| Observed species | 16.00 ± 2.33 b | 28.57 ± 0.76 a | 24.23 ± 3.48 ab | 25.93 ± 6.05 a |
| Simpson | 0.22 ± 0.01 c | 0.26 ± 0.03 c | 0.33 ± 0.01 b | 0.49 ± 0.02 a |
| PD whole tree | 1.39 ± 0.14 b | 2.56 ± 0.09 a | 2.14 ± 0.26 ab | 2.57 ± 0.68 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, X.; Zhao, M.; Zou, S.; Li, Z.; Wu, Y.; Ji, C.; Chen, Y.; Dong, L.; Zhang, S.; Liang, H. Effect of Autochthonous Lactic Acid Bacteria-Enhanced Fermentation on the Quality of Suancai. Foods 2022, 11, 3310. https://doi.org/10.3390/foods11213310
Cao X, Zhao M, Zou S, Li Z, Wu Y, Ji C, Chen Y, Dong L, Zhang S, Liang H. Effect of Autochthonous Lactic Acid Bacteria-Enhanced Fermentation on the Quality of Suancai. Foods. 2022; 11(21):3310. https://doi.org/10.3390/foods11213310
Chicago/Turabian StyleCao, Xinying, Mingwei Zhao, Sibo Zou, Zhigao Li, Yuzheng Wu, Chaofan Ji, Yingxi Chen, Liang Dong, Sufang Zhang, and Huipeng Liang. 2022. "Effect of Autochthonous Lactic Acid Bacteria-Enhanced Fermentation on the Quality of Suancai" Foods 11, no. 21: 3310. https://doi.org/10.3390/foods11213310
APA StyleCao, X., Zhao, M., Zou, S., Li, Z., Wu, Y., Ji, C., Chen, Y., Dong, L., Zhang, S., & Liang, H. (2022). Effect of Autochthonous Lactic Acid Bacteria-Enhanced Fermentation on the Quality of Suancai. Foods, 11(21), 3310. https://doi.org/10.3390/foods11213310