
The Mechanisms of the Potential Probiotic Lactiplantibacillus plantarum against Cardiovascular Disease and the Recent Developments in its Fermented Foods



Abstract
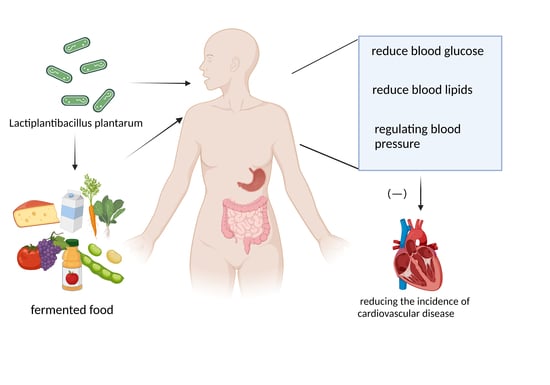
:
1. Introduction
2. Probiotic Potential of Lb. plantarum and Foods Fermented with It
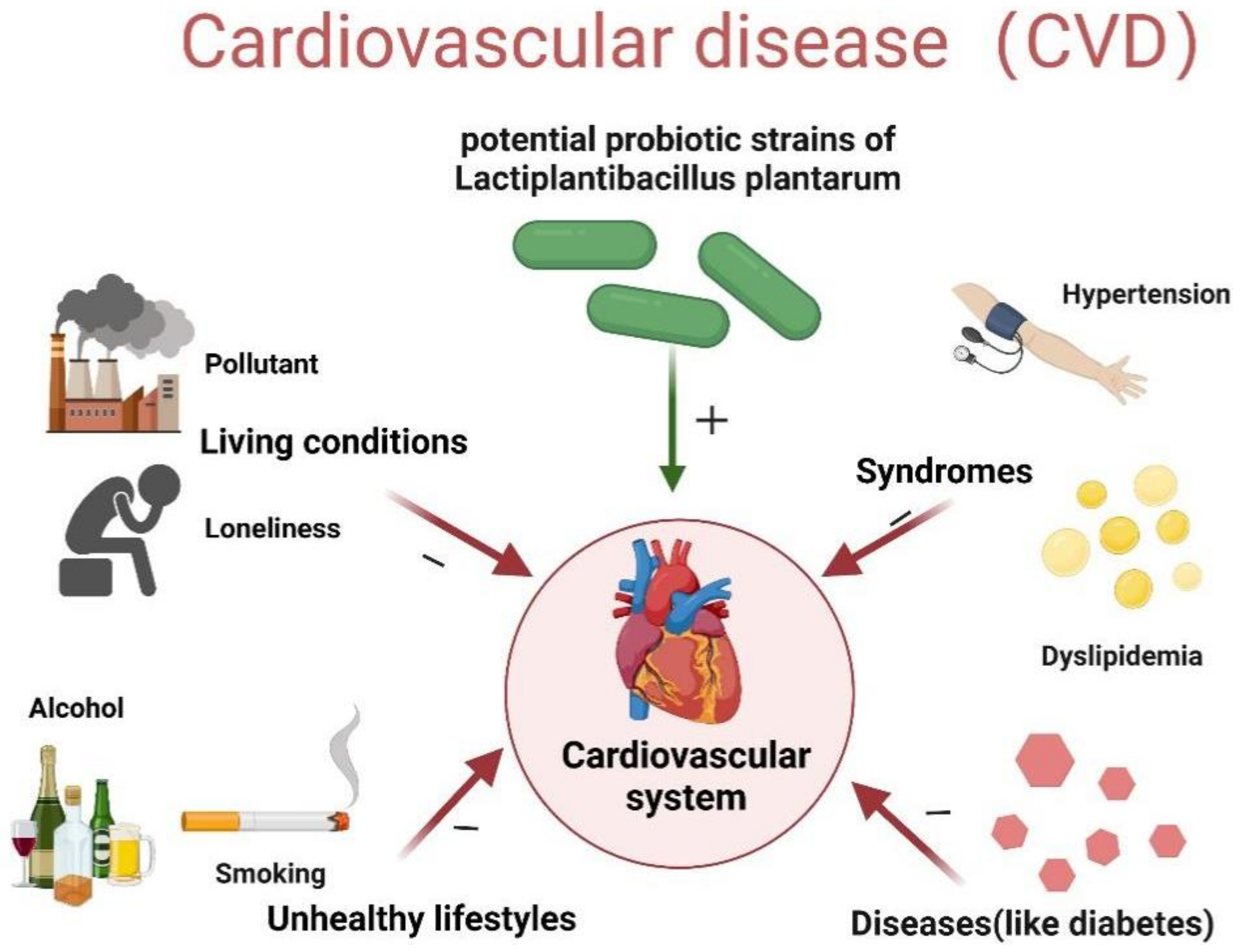
3. Possible Mechanism of Lb. plantarum against CVD
3.1. Mechanisms of Antioxidant Abilities by Lb. plantarum

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Lb. plantarum | Resource | In Vivo Studies(a) or In Vitro Studies(b) | Antioxidant Mechanism | Reference |
|---|---|---|---|---|
| Lb. plantarum CCFM10 | Different storage centers (edible fungi strains) | a | Increase levels of GSH, CAT, SOD, and TOC in serum of mice with oxidative damage | [35] |
| Lb. plantarum CCFM242 | Different storage centers (edible fungi strains) | a | Improve total antioxidant capacity of liver | [35] |
| Lb. plantarum RS15-3 | Different storage centers (edible fungi strains) | a | Increase levels of GSH, CAT, SOD, and TOC in serum of mice with oxidative damage | [35] |
| Lb. plantarum FEED8 | Intestinal tract of longevity elderly in Bama, Guangxi | a | Increase levels of glutathione (GSH) and other indicators | [34] |
| Lb. plantarum NCFM, ATCC 14917 and NDC 75017 | a | Increase SOD activity, GSH-Px activity, and T-AOC content in serum, brain, and liver, decreasing MDA content. | [26] | |
| Lb. plantarum C88 | Traditional fermented milk tofu in Inner Mongolia | a | Regulation of intracellular antioxidant enzyme activity in oxidative damaged Caco-2 cells | [36] |
| Lb. plantarum Y-20 | Chopped pepper by natural fermentation | a | Protect the Caco-2 cells against H2O2 and induce oxidative stress by renewing the enzymatic and non-enzymatic antioxidant defense system. | [37] |
| Lb. plantarum JMCC0017 | Xinjiang traditional fermented dairy products | b | Metabolites have significant antioxidant activity | [38] |
| Lb. plantarum DM5 | Fermented beverage Marcha of Sikkim | a | Scavenge free radicals and superoxide anion | [27,28] |
| Lb. plantarum KM1 | Natural fermentation products | a | Scavenge hydroxyl radical, superoxide anion radical, and DPPH radical | [31] |
| Lb. plantarum AR501 | a | Scavenge hydroxyl radical, superoxide anion radical, and DPPH | [39] | |
| Lb. plantarum JM113 | healthy intestinal contents of Tibetan chicken | a | Change the expression levels of Phosphoglycerin kinase, α-glycerophosphate oxidase, pyruvate oxidase, and NADH peroxidase to alleviate oxidative stress. | [23] |
| Lb. plantarum NC8 | a | Up-regulation of antioxidant gene expression | [40] | |
| Lb. plantarum SM4. | kimchi and fermented with white Taraxacum coreanum | a | Increase the mRNA levels of Nrf2 and its corresponding downstream HO-1 gene | [41] |
| Lb. plantarum KCCP11226 | a | Regulation of Bcl-2 family members and activation of Bcl-2/Bax signaling pathway | [24,25] | |
| Lb. plantarum ZLP001 | b | Produce white T. coreanum fermented product which shows higher bioactive properties of oxidation resistance | [42] | |
| Lb. plantarum KLDS1.0202 | Cheddar cheese | b | Produce C-30 carotenoid 4,4′-diaponeurosporene. | [43,44] |
| Lb. plantarum isolated from traditional sourdough | Traditional Sourdough | a | Supplementation of Lb. plantarum ZLP001 increases the concentration of superoxide dismutase (p < 0.05), glutathione peroxidase (p < 0.01), and catalase in serum (p < 0.10), while decreasing the concentration of malondialdehyde (p < 0.05). | [45] |
| Lb. plantarum NJAU-01 | jinhua ham | The strain not only itself has a certain antioxidant activity, but also promotes the decomposition of protein of Cheddar cheese | [29,30] |
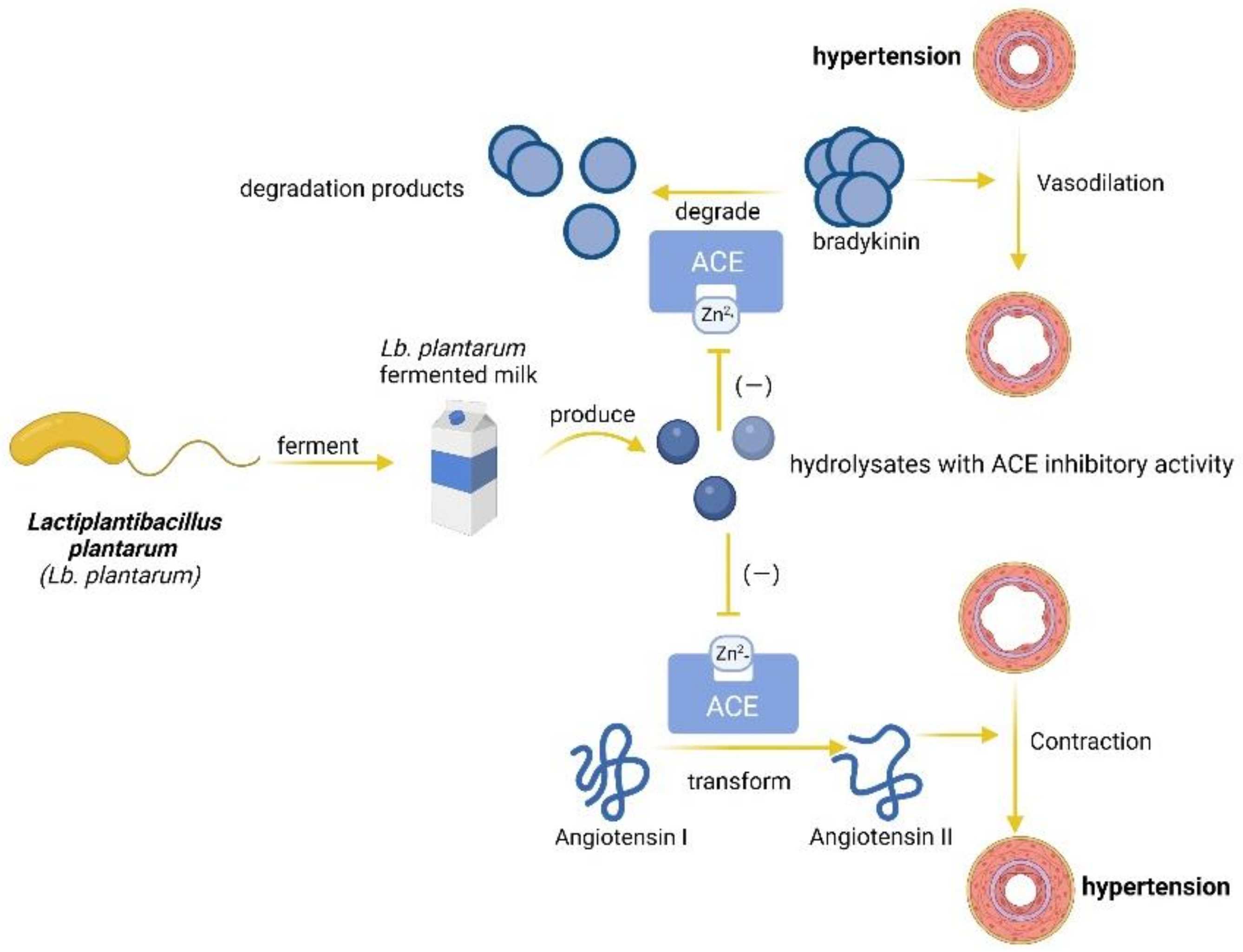
3.2. Mechanisms of Blood Pressure Lowering by Lb. plantarum
| Lactiplantibacillus plantarum | Fermented Food | Fermentation Condition | ACE Inhibition Effect | References |
|---|---|---|---|---|
| Lb. plantarum L69 together with Directed Vat Set starter containing Lactobacillus bulgaricus and Streptococcus thermophilus (1:1) | skim milk powder | 4.5 h, 42 °C, pH = 4.65 | Lb. plantarum L69 contributed to ACEI substances production compared with single Directed Vat Set starter, with ACEI activity reaching 87.14%. | [66] |
| Lb. plantarum SPS109 | whey beverage | 72 h, 35 °C, pH = 5.5 | ACEI activity = 25.70 ± 1.20% | [67] |
| Lb. plantarum previously isolated and identified from Chiapas double cream cheese | reconstituted whole milk. | 48 h, 37 °C, pH = 9 | 59.3 ± 1.6% ACEI activity | [68] |
| Lb. plantarum K25 together with yogurt starter L. delbrueckiissp. bulgaricus and Streptococcus thermophilus) | yogurt | 37 °C, pH = 4.5 ± 0.5, then storing in 4 °C for 21 days | Lb. plantarum K25 significantly raising the ACEI ratio in contrast to fermentation with the starter only, with the estimation of ACEI activity = 49.3% | [69] |
| Lb. plantarum BG 112 | soymilk containing okara flour | 32 h | 50% ACEI activity | [70] |
| Lb. Plantarum which NCBI Accession is KF806535 | soy milk | 24 h, 37 °C, 100 rpm | ACEI activity (in-vitro) of peptides all above 70% | [71] |
| Lb. plantarum (TISTR 858) | eggshell membranes | 30 °C, 120 rpm | ACE-inhibition corresponding to 49.3% with the concentration of the protein hydrolysates obtained after fermentation up to 2 mg/mL | [72] |
| Lb. plantarum 70810 | navy bean milk | 2 h, 31 °C | 50% inhibiting concentration (IC50) = 109 ± 5.1μg protein/ml | [73] |
| Lb. plantarum B1-6 | navy bean milk | 3 h, 37 °C | IC50 = 101 ± 2.2 μg protein/mL, in vitro gastrointestinal simulation IC50 = 21 ± 2.1 μg protein/ml | [73] |
| Lb. plantarum (NCDO 1193) | freeze-dried camu-camu powder and soymilk combination | 37 °C, 72 h | 94.0 ± 1.0% ACEI activity | [74] |
| Lb. plantarum 69 | goat milk | 35 °C, CaCl2 concentration of 0.07%, and Tween-80 concentration of 0.04% | 88.91%ACEI activity | [51] |
| Lb. plantarum KU15003 | yogurt | fermentation was terminated when the pH reached 4.4 ± 0.1. | IC50 = 0.68 mg/mL | [75] |
| Lb. plantarum KU15031 (T3) | yogurt | fermentation was terminated when the pH reached 4.4 ± 0.1. | IC50 = 0.79 mg/mL | [75] |
| Lb. plantarum NK181 (T4) | yogurt | fermentation was terminated when the pH reached 4.4 ± 0.1. | IC50 = 0.48 mg/mL | [75] |
| Lb. plantarum QS670 | milk | 37 °C, 48 h | IC50 = 1.26 mg/mL | [57] |
3.3. Mechanisms of Lipid Lowering by Lb. plantarum
3.3.1. Lowering Cholesterol through Signaling Pathways
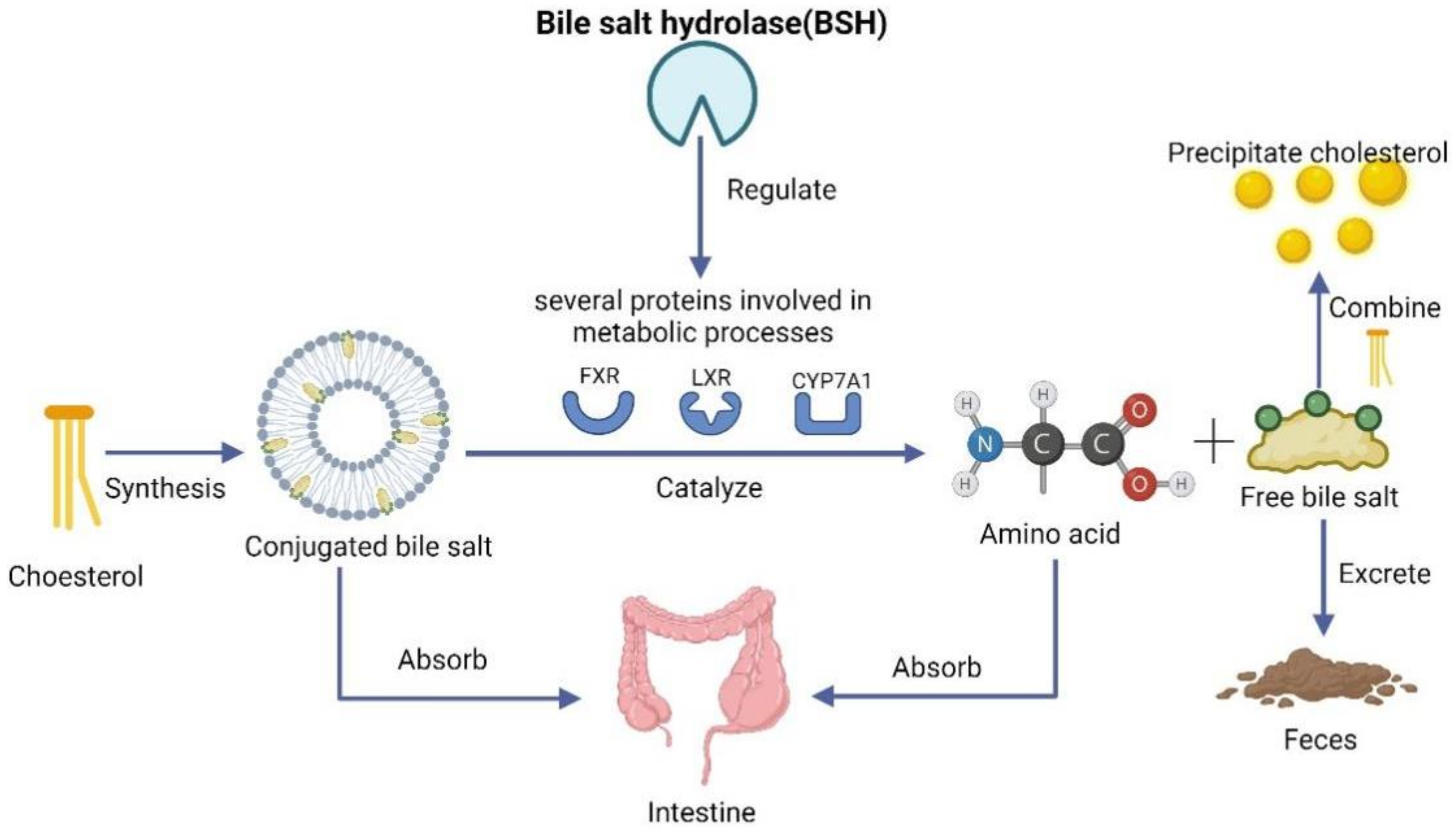
3.3.2. Lowering Cholesterol through Bile Acids
3.3.3. Lowering Cholesterol through Intestinal Flora
3.3.4. Lowering Cholesterol through Conjugated Linoleic Acid (CLA) Isomerase
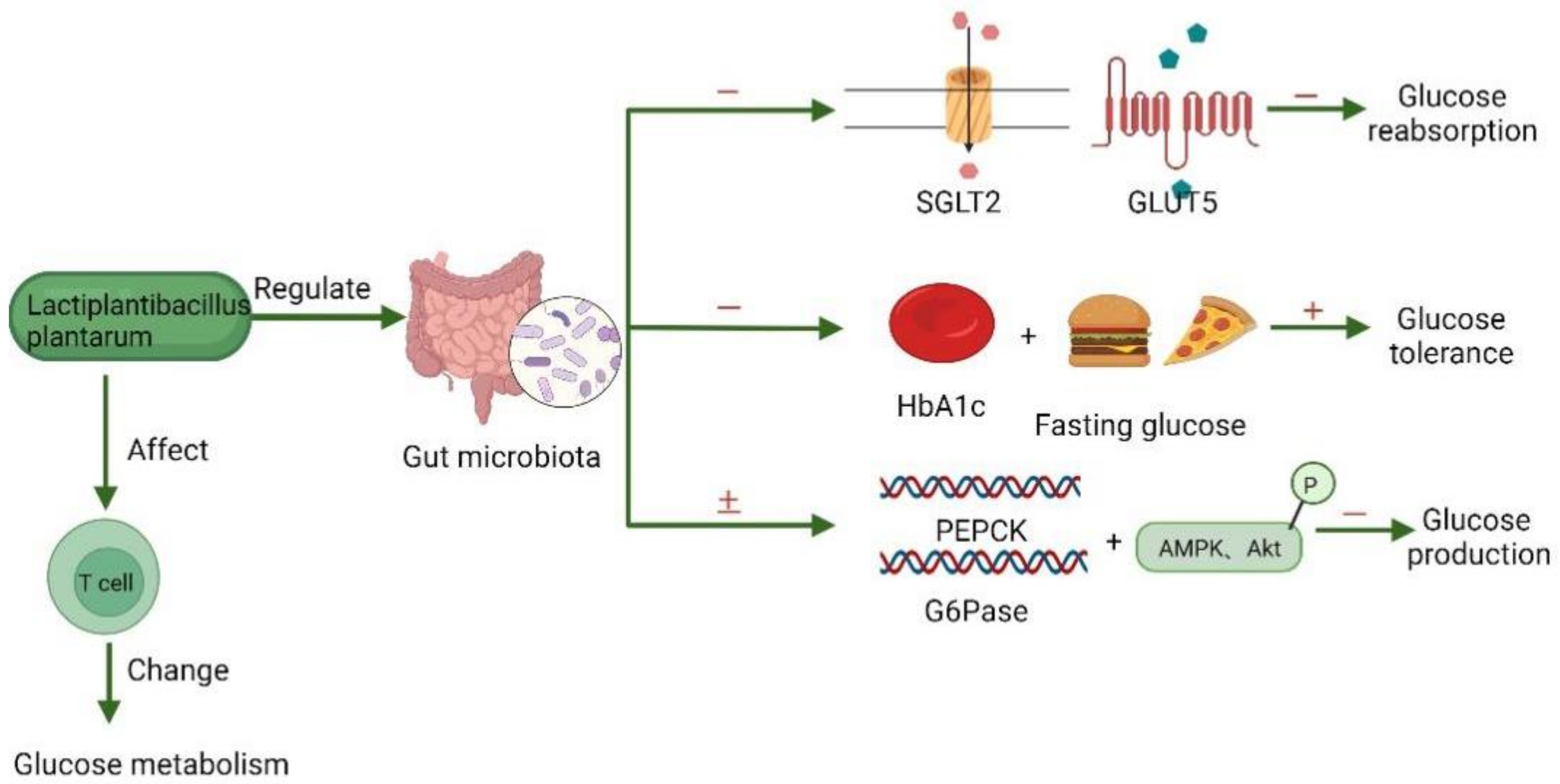
3.4. Mechanisms of Glucose Lowering by Lb. plantarum
4. Recent Developments in Lb. plantarum-Fermented Food
4.1. Fermented Food with Antioxidant Function
4.2. Fermented Food with Cholesterol-Lowering Function
4.3. Fermented Food with Blood Pressure Lowering Function
4.4. Fermented Food with Hypoglycemic Function
4.5. Other Applications of Lb. plantarum
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Cardiovascular Diseases (CVDs). 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 11 June 2021).
- Shengshou, H. Report on Cardiovascular Health and Diseases Burden in China: An Updated Summary of 2020. Chin. Circ. J. 2021, 36, 521–545. [Google Scholar]
- Andersson, C.; Vasan, R.S. Epidemiology of cardiovascular disease in young individuals. Nat. Rev. Cardiol. 2017, 15, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Khanal, M.K.; Bhandari, P.; Dhungana, R.R.; Gurung, Y.; Rawal, L.B.; Pandey, G.; Bhandari, M.; Devkota, S.; de Courten, M.; de Courten, B. Poor glycemic control, cardiovascular disease risk factors and their clustering among patients with type 2 diabetes mellitus: A cross-sectional study from Nepal. PLoS ONE 2022, 17, e0271888. [Google Scholar] [CrossRef] [PubMed]
- Ciumărnean, L.; Milaciu, M.V.; Negrean, V.; Orășan, O.H.; Vesa, S.C.; Sălăgean, O.; Iluţ, S.; Vlaicu, S.I. Cardiovascular Risk Factors and Physical Activity for the Prevention of Cardiovascular Diseases in the Elderly. Int. J. Environ. Res. Public Health 2021, 19, 207. [Google Scholar] [CrossRef]
- Barbaresko, J.; Rienks, J.; Nöthlings, U. Lifestyle Indices and Cardiovascular Disease Risk: A Meta-analysis. Am. J. Prev. Med. 2018, 55, 555–564. [Google Scholar] [CrossRef]
- Powell-Wiley, T.M.; Baumer, Y.; Baah, F.O.; Baez, A.S.; Farmer, N.; Mahlobo, C.T.; Pita, M.A.; Potharaju, K.A.; Tamura, K.; Wallen, G.R. Social Determinants of Cardiovascular Disease. Circ. Res. 2022, 130, 782–799. [Google Scholar] [CrossRef]
- Habeeb, E.; Aldosari, S.; Saghir, S.A.; Cheema, M.; Momenah, T.; Husain, K.; Omidi, Y.; Rizvi, A.S.A.; Akram, M.; Ansari, R.A. Role of environmental toxicants in the development of hypertensive and cardiovascular diseases. Toxicol. Rep. 2022, 9, 521–533. [Google Scholar] [CrossRef]
- Fenster, K.; Freeburg, B.; Hollard, C.; Wong, C.; Laursen, R.R.; Ouwehand, A.C. The Production and Delivery of Probiotics: A Review of a Practical Approach. Microorganisms 2019, 7, 83. [Google Scholar] [CrossRef]
- Rasika, D.; Vidanarachchi, J.; Luiz, S.; Azeredo, D.; Cruz, A.; Ranadheera, C. Probiotic Delivery through Non-Dairy Plant-Based Food Matrices. Agriculture 2021, 11, 599. [Google Scholar] [CrossRef]
- Wu, W.; Wang, L.; Zhao, J.; Zhang, H.; Chen, W. Research progress on physiological characteristics and health. Food Ferment. Ind. 2019, 45, 1–13. [Google Scholar]
- Oh, Y.; Kim, T.; Moon, H.; Lee, S.; Lee, S.; Ji, G.; Hwang, K. Lactobacillus plantarum PMO 08 as a Probiotic Starter Culture for Plant-Based Fermented Beverages. Molecules 2020, 25, 5056. [Google Scholar] [CrossRef] [PubMed]
- Kimoto-Nira, H.; Moriya, N.; Nogata, Y.; Sekiyama, Y.; Toguchi, Y. Fermentation of Shiikuwasha (Citrus depressa Hayata) pomace by lactic acid bacteria to generate new functional materials. Int. J. Food Sci. Technol. 2018, 54, 688–695. [Google Scholar] [CrossRef]
- Han, S.; Lu, Y.; Xie, J.; Fei, Y.; Zheng, G.; Wang, Z.; Liu, J.; Lv, L.; Ling, Z.; Berglund, B.; et al. Probiotic Gastrointestinal Transit and Colonization After Oral Administration: A Long Journey. Front. Cell. Infect. Microbiol. 2021, 11, 609722. [Google Scholar] [CrossRef]
- Ghatani, K.; Tamang, B. Assessment of probiotic characteristics of lactic acid bacteria isolated from fermented yak milk products of Sikkim, India: Chhurpi, Shyow, and Khachu. Food Biotechnol. 2017, 31, 210–232. [Google Scholar] [CrossRef]
- Wang, G.; Chen, Y.; Fei, S.; Xie, C.; Xia, Y.; Ai, L. Colonisation with endogenous Lactobacillus reuteri R28 and exogenous Lactobacillus plantarum AR17-1 and the effects on intestinal inflammation in mice. Food Funct. 2021, 12, 2481–2488. [Google Scholar] [CrossRef] [PubMed]
- Galdino, I.K.C.P.D.O.; Oliveira, M.M.; Oliveira, A.T.; da Silva, G.M.; de Oliveira, T.A.; dos Santos, K.M.O.; Egito, A.S.D.; Buriti, F.C.A. Fermentative behavior of native lactobacilli in goat milk and their survival under in vitro simulated gastrointestinal conditions. LWT 2020, 135, 109905. [Google Scholar] [CrossRef]
- Flach, J.; van der Waal, M.; Nieuwboer, M.V.D.; Claassen, H.; Larsen, O.F.A. The underexposed role of food matrices in probiotic products: Reviewing the relationship between carrier matrices and product parameters. Crit. Rev. Food Sci. Nutr. 2017, 58, 2570–2584. [Google Scholar] [CrossRef]
- Sharma, R.; Mokhtari, S.; Jafari, S.M.; Sharma, S. Barley-based probiotic food mixture: Health effects and future prospects. Crit. Rev. Food Sci. Nutr. 2021, 1–15. [Google Scholar] [CrossRef]
- Zhu, W.; Lyu, F.; Naumovski, N.; Ajlouni, S.; Ranadheera, C.S. Functional Efficacy of Probiotic Lactobacillus sanfranciscensis in Apple, Orange and Tomato Juices with Special Reference to Storage Stability and In Vitro Gastrointestinal Survival. Beverages 2020, 6, 13. [Google Scholar] [CrossRef]
- De Oliveira, P.M.; de Castro Leite Junior, B.R.; Martins, E.M.F.; Martins, M.L.; Vieira, N.R.; de Barros, F.A.R.; Cristianini, M.; de Almeida Costa, N.; Ramos, A.M. Mango and carrot mixed juice: A new matrix for the vehicle of probiotic lactobacilli. J. Food Sci. Technol. 2020, 58, 98–109. [Google Scholar] [CrossRef]
- Baldwin, C.M.; Bootzin, R.R.; Schwenke, D.C.; Quan, S.F. Antioxidant nutrient intake and supplements as potential moderators of cognitive decline and cardiovascular disease in obstructive sleep apnea. Sleep Med. Rev. 2005, 9, 459–476. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, L.; Duan, Y.; Yang, X. Antioxidant activity of Lactobacillus plantarum JM113 in vitro and its protective effect on broiler chickens challenged with deoxynivalenol1. J. Anim. Sci. 2017, 95, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Seo, D.-H.; Park, Y.-S.; Cha, I.-T.; Seo, M.-J. Isolation of Lactobacillus plantarum subsp. plantarum Producing C30 Carotenoid 4,4′-Diaponeurosporene and the Assessment of Its Antioxidant Activity. J. Microbiol. Biotechnol. 2019, 29, 1925–1930. [Google Scholar] [CrossRef]
- Kim, M.; Jung, D.-H.; Seo, D.-H.; Park, Y.-S.; Seo, A.M.-J. 4,4′-Diaponeurosporene from Lactobacillus plantarum subsp. plantarum KCCP11226: Low Temperature Stress-Induced Production Enhancement and In Vitro Antioxidant Activity. J. Microbiol. Biotechnol. 2021, 31, 63–69. [Google Scholar] [CrossRef]
- Liu, S.; Jiang, C.Y. Antioxidant activity of three strains of lactic acid bacteria and investigate the antioxidant mechanism primarily. In Proceedings of the 9th International Symposium on Lactic Acid Bacteria and Health, Tianjing, China, 21–23 May 2014; pp. 68–69. [Google Scholar]
- Das, D.; Goyal, A. Potential probiotic attributes and antagonistic activity of an indigenous isolate Lactobacillus plantarum DM5 from an ethnic fermented beverage “Marcha” of North Eastern Himalayas. Int. J. Food Sci. Nutr. 2014, 65, 335–344. [Google Scholar] [CrossRef]
- Das, D.; Goyal, A. Antioxidant activity and γ-aminobutyric acid (GABA) producing ability of probiotic Lactobacillus plantarum DM5 isolated from Marcha of Sikkim. LWT Food Sci. Technol. 2015, 61, 263–268. [Google Scholar] [CrossRef]
- Yang, B. Study on the Mechanism of Lactobacillus Plantarum NJAU-01 to Relieve Oxidative Stress in Mice. BMC Microbiol. 2021, 21, 182. [Google Scholar]
- Chen, L. Research on the Antioxidant Ability and Mechanism of Lactobacillus Plantarum NJAU-01; Yangzhou University: Yangzhou, China, 2020. [Google Scholar]
- Tian, Y.; Wang, Y.; Zhang, N.; Xing, X.; Xiao, M.; Wang, Y. Screening of Antioxidant Lactic Acid Bacteria and Its Antioxidant Mechanism. In Proceedings of the 16th International Symposium on Probiotics and Health, Jiangsu, China, 25 May 2021; pp. 42–43. [Google Scholar]
- Liu, B.; Xiao, X.; Zhou, X.; Zhou, J.; Lan, L.; Long, X.; Pan, Y.; Du, M.; Zhao, X. Effects of Lactobacillus plantarum CQPC01-fermented soybean milk on activated carbon-induced constipation through its antioxidant activity in mice. Food Sci. Nutr. 2019, 7, 2068–2082. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Long, L.; Sun, J.; Bu, L.; Cao, J.; Luo, Y.; Liu, H.; Wu, Y.; Meng, X. Exploring the antioxidant effect of Lactobacillus plantarum SCS2 on mice with type 2 diabetes. J. Food Biochem. 2021, 45, e13781. [Google Scholar] [CrossRef]
- Yan, L.; Wang, X.; Jing, T.; Liu, Z.; Liu, L. Antioxidant effect of Lactobacillus plantarum FEED8 on mice. China Dairy Cattle 2020, 59–63. [Google Scholar] [CrossRef]
- Zhao, J. Evaluation of Antioxidative Abilities of Lactobacillus Plantarum and the Antioxidative Mechanisms. J. Microbiol. Biotechnol. 2018, 28, 1052–1060. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, D.; Zhao, Y.; Zhang, X.; Huang, L. Protective effect of Lactobacillus plantarum C88 on H2O2-induced oxidative damage in Caco-2 cells. Sci. Agric. Sin. 2013, 46, 606–613. [Google Scholar]
- Huang, C. Studies on the Isolation, Identification of Lactobacillus Plantarum Y-20 and the Antioxidant Activity of Its Exopolysaccharides; Hunan Agricultural University: Hunan, China, 2019. [Google Scholar]
- Xun, Y.; Zhang, D.; Feng, L.; Xue, Y.; Wang, S.; Zhu, H. Study on the Antioxidant Effect of Lactobacillus plantarum JMCC0017; China Association for Science and Technology: Beijing, China, 2019; pp. 137–138. [Google Scholar]
- Lv, F.; Wang, G.; Xia, Y.; Zhang, H.; Ai, L.; Xiong, Z. Antioxidant Effect and Mechanism of Lactobacillus plantarum AR501; China Association for Science and Technology: Beijing, China, 2019; pp. 40–41. [Google Scholar]
- Wang, M. Preliminary Study on the Anti-oxidative Stress Effect of Lactobacillus Plantarum and Its Mechanism. Master’s Thesis, Jilin Agricultural University, Jilin, China, 2019. [Google Scholar]
- Hong, J.W.; Park, H.Y.; Kim, J.H.; Yeom, S.H.; Kim, J.W. Antioxidation and Anti-photoaging Effects of White Taraxacum Coreanum Extract by Lactobacillus plantarum. J. Korea Acad. Ind. Coop. Soc. 2021, 22, 554–562. [Google Scholar] [CrossRef]
- Wang, J.; Ji, H.F.; Wang, S.X.; Zhang, D.Y.; Liu, H.; Shan, D.C.; Wang, Y.M. Lactobacillus plantarum ZLP001: In vitro Assessment of Antioxidant Capacity and Effect on Growth Performance and Antioxidant Status in Weaning Piglets. Asian Australas. J. Anim. Sci. 2012, 25, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Liu, B.; Sun, X.; Li, X.; Li, D. Study on antioxidant activities of Lactobacillus plantarum and Lactobacillus casei. China Dairy Ind. 2018, 46, 4–8. [Google Scholar]
- Li, D.; Li, X.; Han, X.; Li, L.; Wang, L.; Zhang, X.; Wang, Y. Antioxidant Activity of Lactobacillus Plantarum and Lactobacillus Casei on Cheddar Cheese and the Number of Simulated Gastrointestinal Bacteria. J. Chin. Inst. Food Sci. Technol. 2018, 18, 130–138. [Google Scholar] [CrossRef]
- Cheng, X.; Sun, Y.; Liu, N.; Huang, W.; Li, N.; Filip, A. Antioxidant Properties and Flavor Characteristics of Steamed Bread Fermented by Lactobacillus plantarum Isolated from Traditional Sourdough. Food Sci. 2015, 36, 87–92. [Google Scholar]
- Whelton, P.K.; Carey, R.M.; Aronow, W.S.; Casey, D.E., Jr.; Collins, K.J.; Himmelfarb, C.D.; DePalma, S.M.; Gidding, S.; Jamerson, K.A.; Jones, D.W.; et al. 2017 ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA Guideline for the Prevention, Detection, Evaluation, and Management of High Blood Pressure in Adults: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Hypertension 2018, 71, e13–e115. [Google Scholar] [CrossRef]
- Mills, K.T.; Stefanescu, A.; He, J. The global epidemiology of hypertension. Nat. Rev. Nephrol. 2020, 16, 223–237. [Google Scholar] [CrossRef]
- Forouzanfar, M.H.; Liu, P.; Roth, G.A. Global burden of hypertension and systolic blood pressure of at least 110 to 115 mm Hg, 1990–2015. JAMA 2017, 317, 648. [Google Scholar] [CrossRef]
- Eriksson, U.; Danilczyk, U.; Penninger, J.M. Just the Beginning: Novel Functions for Angiotensin-Converting Enzymes. Curr. Biol. 2002, 12, R745–R752. [Google Scholar] [CrossRef]
- Lewis-Mikhael, A.-M.; Davoodvandi, A.; Jafarnejad, S. Effect of Lactobacillus plantarum containing probiotics on blood pressure: A systematic review and meta-analysis. Pharmacol. Res. 2020, 153, 104663. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, Q.; Ji, Z.; Shu, G.; Chen, H. Production and fermentation characteristics of angiotensin-I-converting enzyme inhibitory peptides of goat milk fermented by a novel wild Lactobacillus plantarum 69. LWT 2018, 91, 532–540. [Google Scholar] [CrossRef]
- Omedi, J.O.; Huang, W.; Su, X.; Liu, R.; Tang, X.; Xu, Y.; Rayas-Duarte, P. Effect of five lactic acid bacteria starter type on angiotensin-I converting enzyme inhibitory activity and emulsifying properties of soy flour sourdoughs with and without wheat bran supplementation. J. Cereal Sci. 2016, 69, 57–63. [Google Scholar] [CrossRef]
- Sánchez-López, F.; Robles-Olvera, V.J.; Hidalgo-Morales, M.; Tsopmo, A. Angiotensin-I converting enzyme inhibitory activity of Amaranthus hypochondriacus seed protein hydrolysates produced with lactic bacteria and their peptidomic profiles. Food Chem. 2021, 363, 130320. [Google Scholar] [CrossRef]
- Liu, Y.-Y.; Zeng, S.-Y.; Leu, Y.-L.; Tsai, T.-Y. Antihypertensive Effect of a Combination of Uracil and Glycerol Derived from Lactobacillus plantarum Strain TWK10-Fermented Soy Milk. J. Agric. Food Chem. 2015, 63, 7333–7342. [Google Scholar] [CrossRef]
- Nahariah, N.; Legowo, A.M.; Abustam, E.; Hintono, A. Angiotensin I-Converting Enzyme Inhibitor Activity on Egg Albumen Fermentation. Asian Australasian J. Anim. Sci. 2015, 28, 855–861. [Google Scholar] [CrossRef]
- Shu, G.; Yang, H.; Chen, H.; Zhang, Q.; Tian, Y. Effect of incubation time, inoculum size, temperature, pasteurization time, goat milk powder and whey powder on ACE inhibitory activity in fermented milk by L. plantarum LP69. Acta Sci. Polonorum. Technol. Aliment. 2015, 14, 107–116. [Google Scholar] [CrossRef]
- Xia, Y.; Yu, J.; Xu, W.; Shuang, Q. Purification and characterization of angiotensin-I-converting enzyme inhibitory peptides isolated from whey proteins of milk fermented with Lactobacillus plantarum QS670. J. Dairy Sci. 2020, 103, 4919–4928. [Google Scholar] [CrossRef]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-aminobutyric acid as a bioactive compound in foods: A review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Zareian, M.; Oskoueian, E.; Forghani, B.; Ebrahimi, M. Production of a wheat-based fermented rice enriched with γ-amino butyric acid using Lactobacillus plantarum MNZ and its antihypertensive effects in spontaneously hypertensive rats. J. Funct. Foods 2015, 16, 194–203. [Google Scholar] [CrossRef]
- Liu, C.F.; Tung, Y.T.; Wu, C.L.; Lee, B.-H.; Hsu, W.-H.; Pan, T.M. Antihypertensive Effects of Lactobacillus-Fermented Milk Orally Administered to Spontaneously Hypertensive Rats. J. Agric. Food Chem. 2011, 59, 4537–4543. [Google Scholar] [CrossRef]
- Torino, M.I.; Limón, R.I.; Martínez-Villaluenga, C.; Mäkinen, S.; Pihlanto, A.; Vidal-Valverde, C.; Frias, J. Antioxidant and antihypertensive properties of liquid and solid state fermented lentils. Food Chem. 2012, 136, 1030–1037. [Google Scholar] [CrossRef] [PubMed]
- Ahrén, I.L.; Xu, J.; Önning, G.; Olsson, C.; Ahrné, S.; Molin, G. Antihypertensive activity of blueberries fermented by Lactobacillus plantarum DSM 15313 and effects on the gut microbiota in healthy rats. Clin. Nutr. 2015, 34, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Saputri, F.A.; Kang, D.; Kusuma, A.S.W.; Rusdiana, T.; Hasanah, A.N.; Mutakin; Surono, I.S.; Koyama, H.; Abdulah, R. Lactobacillus plantarum IS-10506 probiotic administration increases amlodipine absorption in a rabbit model. J. Int. Med. Res. 2018, 46, 5004–5010. [Google Scholar] [CrossRef]
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Li, E.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J.; et al. Gut Dysbiosis Is Linked to Hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef]
- Robles-Vera, I.; Toral, M.; Romero, M.; Jiménez, R.; Sánchez, M.; Pérez-Vizcaíno, F.; Duarte, J. Antihypertensive Effects of Probiotics. Curr. Hypertens. Rep. 2017, 19, 26. [Google Scholar] [CrossRef]
- Wang, Z.; Shu, G.; Chen, L.; Dai, C.; Li, Y.; Niu, J.; Wan, H. Directed-Vat-Set starter producing ACE-inhibitory peptides: Optimization and evaluation of stability. J. Food Process. Preserv. 2022, 46, e16475. [Google Scholar] [CrossRef]
- Jitpakdee, J.; Kantachote, D.; Kanzaki, H.; Nitoda, T. Potential of lactic acid bacteria to produce functional fermented whey beverage with putative health promoting attributes. LWT 2022, 160, 113269. [Google Scholar] [CrossRef]
- Hernández-Sánchez, H.; Nacional, I.P.; Fajardo-Espinoza, F.; Gutiérrez-López, G.; Ávila-Reyes, S.; Cano-Sarmiento, C.; Figueroa-Hernández, C. ACE-Inhibitory and metal-binding activity produced during milk fermentation by three probiotic potential LAB strains isolated from Chiapas double cream cheese. Rev. Mex. De Ing. Quim. 2020, 20, 97–112. [Google Scholar] [CrossRef]
- Min, Z.; Yunyun, J.; Miao, C.; Zhennai1, Y. Characterization and ACE Inhibitory Activity of Fermented Milk with Probiotic Lactobacillus plantarum K25 as Analyzed by GC-MS-Based Metabolomics Approach. J. Microbiol. Biotechnol. 2020, 30, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Filho, M.L.M.; Busanello, M.; Garcia, S. Probiotic creamy soy sauce with Lactobacillus plantarum BG 112. Br. Food J. 2019, 121, 2746–2758. [Google Scholar] [CrossRef]
- Singh, B.P.; Vij, S. Growth and bioactive peptides production potential of Lactobacillus plantarum strain C2 in soy milk: A LC-MS/MS based revelation for peptides biofunctionality. LWT 2017, 86, 293–301. [Google Scholar] [CrossRef]
- Jain, S.; Anal, A.K. Production and characterization of functional properties of protein hydrolysates from egg shell membranes by lactic acid bacteria fermentation. J. Food Sci. Technol. 2017, 54, 1062–1072. [Google Scholar] [CrossRef]
- Rui, X.; Wen, D.; Li, W.; Chen, X.; Jiang, M.; Dong, M. Enrichment of ACE inhibitory peptides in navy bean (Phaseolus vulgaris) using lactic acid bacteria. Food Funct. 2014, 6, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Fujita, A.; Sarkar, D.; Genovese, M.I.; Shetty, K. Improving anti-hyperglycemic and anti-hypertensive properties of camu-camu (Myriciaria dubia Mc. Vaugh) using lactic acid bacterial fermentation. Process Biochem. 2017, 59, 133–140. [Google Scholar] [CrossRef]
- Kim, E.-D.; Lee, H.-S.; Kim, K.-T.; Paik, H.-D. Antioxidant and Angiotensin-Converting Enzyme (ACE) Inhibitory Activities of Yogurt Supplemented with Lactiplantibacillus plantarum NK181 and Lactobacillus delbrueckii KU200171 and Sensory Evaluation. Foods 2021, 10, 2324. [Google Scholar] [CrossRef]
- Herrero-Fernandez, B.; Gomez-Bris, R.; Somovilla-Crespo, B.; Gonzalez-Granado, J.M. Immunobiology of Atherosclerosis: A Complex Net of Interactions. Int. J. Mol. Sci. 2019, 20, 5293. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Z.; Jin, G.; Yang, X.; Zhou, H. Regulating dyslipidemia effect of polysaccharides from Pleurotus ostreatus on fat-emulsion-induced hyperlipidemia rats. Int. J. Biol. Macromol. 2017, 101, 107–116. [Google Scholar] [CrossRef]
- Hasenfuß, G. Secondary prevention of cardiovascular diseases: Current state of the art. Kardiol. Pol. 2018, 76, 1671–1679. [Google Scholar] [CrossRef]
- Qu, T.; Yang, L.; Wang, Y.; Jiang, B.; Shen, M.; Ren, D. Reduction of serum cholesterol and its mechanism by Lactobacillus plantarum H6 screened from local fermented food products. Food Funct. 2020, 11, 1397–1409. [Google Scholar] [CrossRef] [PubMed]
- Colautti, A.; Arnoldi, M.; Comi, G.; Iacumin, L. Antibiotic resistance and virulence factors in lactobacilli: Something to carefully consider. Food Microbiol. 2022, 103, 103934. [Google Scholar] [CrossRef] [PubMed]
- Ding, P.; Ding, K.; Yu, Z.; Li, W.; Li, Y.; Liu, Y.; He, W.; Zhao, Z.; Wang, Y.; Cheng, X.; et al. Screening of Lactobacillus with Cholesterol-Degrading Property and Its Effect on Serum Cholesterol in Mice. Food Sci. 2016, 37, 192–197. [Google Scholar]
- Ding, W.; Shi, C.; Chen, M.; Zhou, J.; Long, R.; Guo, X. Screening for lactic acid bacteria in traditional fermented Tibetan yak milk and evaluating their probiotic and cholesterol-lowering potentials in rats fed a high-cholesterol diet. J. Funct. Foods 2017, 32, 324–332. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Q.; Sun, J.; Jiang, B.; Yu, J.; Qi, X. Screening and cholesterol removal ability of lactic acid bacteria DCel3 from Phasianus colchicus. Food Ferment. Ind. 2015, 41, 47–51. [Google Scholar]
- Tang, Y.; Yu, S.; Guo, L.; Wang, N.; Huo, G. Screening and study on probiotic characteristics of a cholesterol-lowering lactobacillus. Sci. Technol. Food Ind. 2016, 37, 142–144, 152. [Google Scholar]
- Wang, J.; Gu, Y.; Ma, W.; Yang, Z. Screening of Potential Probiotic Lactic Acid Bacteria from Inner Mongolian Dairy Tofu. Food Sci. 2014, 35, 171–177. [Google Scholar]
- Xin, J.; Ni, X.; Zeng, D.; Wang, H.; Li, T.; Shen, X. Screening of the hypocholesterolemic effect and body fat control ability of Lactobacillus plantarum F. J. China Agric. Univ. 2014, 19, 137–142. [Google Scholar]
- Zhang, K.; Tian, J. Screening of Cholesterol-Reducing Lactobacillus and Adverse Effects in Mice. J. Food Sci. Biotechnol. 2014, 33, 1222–1227. [Google Scholar]
- Zhao, F.; Li, Y.; Li, B. Screening of probiotic Lactobacillus in simulated gastrointestinal environment. Microbiol. China 2016, 43, 1396–1403. [Google Scholar]
- Ooi, L.-G.; Liong, M.-T. Cholesterol-Lowering Effects of Probiotics and Prebiotics: A Review of in Vivo and in Vitro Findings. Int. J. Mol. Sci. 2010, 11, 2499–2522. [Google Scholar] [CrossRef]
- Lye, H.-S.; Rusul, G.; Liong, M.-T. Removal of cholesterol by lactobacilli via incorporation and conversion to coprostanol. J. Dairy Sci. 2010, 93, 1383–1392. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.-L.; Yu, B.; Li, Z.; Jiang, W.-X.; Jiang, J.-D.; Kong, W.-J. Gastrodin Ameliorates Oxidative Stress and Proinflammatory Response in Nonalcoholic Fatty Liver Disease through the AMPK/Nrf2 Pathway. Phytotherapy Res. 2015, 30, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Lew, L.-C.; Choi, S.-B.; Khoo, B.-Y.; Sreenivasan, S.; Ong, K.-L.; Liong, M.-T. Lactobacillus plantarum DR7 Reduces Cholesterol via Phosphorylation of AMPK That Down-regulated the mRNA Expression of HMG-CoA Reductase. Korean J. Food Sci. Anim. Resour. 2018, 38, 350–361. [Google Scholar] [CrossRef] [PubMed]
- Leung, C.; Rivera, L.; Furness, J.B.; Angus, C.L.P.W. The role of the gut microbiota in NAFLD. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 412–425. [Google Scholar] [CrossRef]
- Tang, T.; Song, J.J.; Li, J.; Wang, H.W.; Zhang, Y.; Suo, H.Y. A synbiotic consisting of Lactobacillus plantarum S58 and hull-less barley β-glucan ameliorates lipid accumulation in mice fed with a high-fat diet by activating AMPK signaling and modulating the gut microbiota. Carbohydr. Polym. 2021, 243, 116398. [Google Scholar] [CrossRef]
- Teng, Y.; Wang, Y.; Tian, Y.; Chen, Y.-Y.; Guan, W.-Y.; Piao, C.-H. Lactobacillus plantarum LP104 ameliorates hyperlipidemia induced by AMPK pathways in C57BL/6N mice fed high-fat diet. J. Funct. Foods 2019, 64, 103665. [Google Scholar] [CrossRef]
- Xu, Y.; Wu, Y.; Wang, S.; Lin, X. Liver Protection Mechanism of Lactobacillus plantarum in Nonalcoholic Fatty Liver Mice. Genom. Appl. Biol. 2020, 39, 1845–1851. [Google Scholar]
- Zhao, Z.; Wang, C.; Zhang, L.; Zhao, Y.; Duan, C.; Zhang, X.; Gao, L.; Li, S. Lactobacillus plantarum NA136 improves the non-alcoholic fatty liver disease by modulating the AMPK/Nrf2 pathway. Appl. Microbiol. Biotechnol. 2019, 103, 5843–5850. [Google Scholar] [CrossRef]
- Rosso, C.; Younes, R.; Eslam, M.; Chen, F.; Cucco, M.; Gaggini, M.; Coulter, S.; Carli, F.; Barbieri, C.; Della Latta, V.; et al. Bile acid composition modulates insulin resistance in non-diabetic patients with NAFLD. Dig. Liver Dis. 2018, 50, 17. [Google Scholar] [CrossRef]
- Kumar, R.; Rajkumar, H.; Kumar, M.; Varikuti, S.R.; Athimamula, R.; Shujauddin, M.; Ramagoni, R.; Kondapalli, N. Molecular cloning, characterization and heterologous expression of bile salt hydrolase (Bsh) from Lactobacillus fermentum NCDO394. Mol. Biol. Rep. 2013, 40, 5057–5066. [Google Scholar] [CrossRef] [PubMed]
- Sridevi, N.; Prabhune, A.A. Brevibacillus sp: A Novel Thermophilic Source for the Production of Bile Salt Hydrolase. Appl. Biochem. Biotechnol. 2008, 157, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, X.; Wang, J.; Wu, F.; Sui, Y.; Yang, L.; Wang, Z. Lactobacillus plantarum strains as potential probiotic cultures with cholesterol-lowering activity. J. Dairy Sci. 2013, 96, 2746–2753. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Huang, W.; Xia, Y.; Xiong, Z.; Ai, L. Cholesterol-lowering potentials of Lactobacillus strain overexpression of bile salt hydrolase on high cholesterol diet-induced hypercholesterolemic mice. Food Funct. 2019, 10, 1684–1695. [Google Scholar] [CrossRef]
- Guo, L.; Wang, L.; Liu, F.; Li, B.; Tang, Y.; Yu, S.; Zhang, D.; Huo, G. Effect of bile salt hydrolase-active Lactobacillus plantarum KLDS 1.0344 on cholesterol metabolism in rats fed a high-cholesterol diet. J. Funct. Foods 2019, 61, 103497. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, K.; Zhao, Y.; Du, R.; Tian, J.; Jin, Y. Research Progress of Regulatory Key Factors Involved in Cholesterol Metabolism by Lactic Acid Bacteria. J. Chin. Inst. Food Sci. Technol. 2021, 21, 341–350. [Google Scholar]
- Aarti, C.; Khusro, A. Functional and technological properties of exopolysaccharide producing autochthonous Lactobacillus plantarum strain AAS3 from dry fish based fermented food. LWT 2019, 114, 108387. [Google Scholar] [CrossRef]
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.A.; Hirschfield, G.M.; Hold, G.; Quraishi, M.N.; Kinross, J.; Smidt, H.; Tuohy, K.M.; et al. The gut microbiota and host health: A new clinical frontier. Gut 2016, 65, 330–339. [Google Scholar] [CrossRef]
- Shao, Y.; Huo, D.; Peng, Q.; Pan, Y.; Jiang, S.; Liu, B.; Zhang, J. Lactobacillus plantarum HNU082-derived improvements in the intestinal microbiome prevent the development of hyperlipidaemia. Food Funct. 2017, 8, 4508–4516. [Google Scholar] [CrossRef]
- Yang, B.; Chen, H.; Gu, Z.; Tian, F.; Ross, R.P.; Stanton, C.; Chen, Y.Q.; Chen, W.; Zhang, H. Synthesis of conjugated linoleic acid by the linoleate isomerase complex in food-derived lactobacilli. J. Appl. Microbiol. 2014, 117, 430–439. [Google Scholar] [CrossRef]
- Kishino, S.; Takeuchi, M.; Park, S.-B.; Hirata, A.; Kitamura, N.; Kunisawa, J.; Kiyono, H.; Iwamoto, R.; Isobe, Y.; Arita, M.; et al. Polyunsaturated fatty acid saturation by gut lactic acid bacteria affecting host lipid composition. Proc. Natl. Acad. Sci. USA 2013, 110, 17808–17813. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.-J.; Yan, L.-H.; Li, Q.; Zhang, C.-J.; Si, C.-L.; Li, Z.-Y.; Song, Y.-J.; Zhou, H.; Zhang, T.-C.; Luo, X.-G. Bioconversion of conjugated linoleic acid by Lactobacillus plantarum CGMCC8198 supplemented with Acer truncatum bunge seeds oil. Food Sci. Biotechnol. 2017, 26, 1595–1611. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhao, W.; Zhang, F.; Yang, X.; Zhu, X.; Wang, L.; Gao, P.; Shu, G.; Jiang, Q. Effects of different conjugated linoleic acid isomers on fat deposition, energy metabolism and intestinal microorganisms in mice. J. S. China Agric. Univ. 2022, 43, 1–8. [Google Scholar]
- Hütt, P.; Songisepp, E.; Rätsep, M.; Mahlapuu, R.; Kilk, K.; Mikelsaar, M. Impact of probiotic Lactobacillus plantarum TENSIA in different dairy products on anthropometric and blood biochemical indices of healthy adults. Benef. Microbes 2015, 6, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Kwon, M.H.; Kim, H.K.; Lee, H.S.; Cho, J.S.; Lee, Y.I. Anti-Obesity Effect of Lactobacillus plantarum LB818 Is Associated with Regulation of Gut Microbiota in High-Fat Diet-Fed Obese Mice. J. Med. Food 2020, 23, 750–759. [Google Scholar] [CrossRef]
- Andersson, U.; Bränning, C.; Ahrné, S.; Molin, G.; Alenfall, J.; Önning, G.; Nyman, M.; Holm, C. Probiotics lower plasma glucose in the high-fat fed C57BL/6J mouse. Benef. Microbes 2010, 1, 189–196. [Google Scholar] [CrossRef]
- da Costa, W.K.A.; Brandão, L.R.; Martino, M.E.; Garcia, E.F.; Alves, A.F.; de Souza, E.L.; Aquino, J.D.S.; Saarela, M.; Leulier, F.; Vidal, H.; et al. Qualification of tropical fruit-derived Lactobacillus plantarum strains as potential probiotics acting on blood glucose and total cholesterol levels in Wistar rats. Food Res. Int. 2018, 124, 109–117. [Google Scholar] [CrossRef]
- Lin, S.; Yang, X.; Long, Y.; Zhong, H.; Wang, P.; Yuan, P.; Zhang, X.; Che, L.; Feng, B.; Li, J.; et al. Dietary supplementation with Lactobacillus plantarum modified gut microbiota, bile acid profile and glucose homoeostasis in weaning piglets. Br. J. Nutr. 2020, 124, 797–808. [Google Scholar] [CrossRef]
- Lee, E.; Jung, S.-R.; Lee, S.-Y.; Lee, N.-K.; Paik, H.-D.; Lim, S.-I. Lactobacillus plantarum Strain Ln4 Attenuates Diet-Induced Obesity, Insulin Resistance, and Changes in Hepatic mRNA Levels Associated with Glucose and Lipid Metabolism. Nutrients 2018, 10, 643. [Google Scholar] [CrossRef]
- Saltiel, A.R.; Kahn, C.R. Insulin signaling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef]
- Pessin, J.E.; Thurmond, D.C.; Elmendorf, J.S.; Coker, K.J.; Okada, S. Molecular Basis of Insulin-stimulated GLUT4 Vesicle Trafficking. Location! Location! Location! J. Biol. Chem. 1999, 274, 2593–2596. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-S.; Lee, D.; Park, G.-S.; Ko, S.-H.; Park, J.; Lee, Y.-K.; Kang, J. Lactobacillus plantarum HAC01 ameliorates type 2 diabetes in high-fat diet and streptozotocin-induced diabetic mice in association with modulating the gut microbiota. Food Funct. 2021, 12, 6363–6373. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, O.A.; Sumlu, E.; Koca, H.B.; Pektas, M.B.; Kocabas, A.; Sadi, G.; Akar, F. Effects of Lactobacillus Plantarum and Lactobacillus Helveticus on Renal Insulin Signaling, Inflammatory Markers, and Glucose Transporters in High-Fructose-Fed Rats. Medicina 2019, 55, 207. [Google Scholar] [CrossRef]
- He, Q.; Cao, C.; Hui, W.; Yu, J.; Zhang, H.; Zhang, W. Genomic resequencing combined with quantitative proteomic analyses elucidate the survival mechanisms of Lactobacillus plantarum P-8 in a long-term glucose-limited experiment. J. Proteom. 2018, 176, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Sakai, T.; Taki, T.; Nakamoto, A.; Shuto, E.; Tsutsumi, R.; Toshimitsu, T.; Makino, S.; Ikegami, S. Lactobacillus plantarum OLL2712 Regulates Glucose Metabolism in C57BL/6 Mice Fed a High-Fat Diet. J. Nutr. Sci. Vitaminol. 2013, 59, 144–147. [Google Scholar] [CrossRef]
- Gao, H.; Wen, J.-J.; Hu, J.-L.; Nie, Q.-X.; Chen, H.-H.; Nie, S.-P.; Xiong, T.; Xie, M.-Y. Momordica charantia juice with Lactobacillus plantarum fermentation: Chemical composition, antioxidant properties and aroma profile. Food Biosci. 2019, 29, 62–72. [Google Scholar] [CrossRef]
- Chen, R.; Chen, W.; Chen, H.; Zhang, G.; Chen, W. Comparative Evaluation of the Antioxidant Capacities, Organic Acids, and Volatiles of Papaya Juices Fermented by Lactobacillus acidophilus and Lactobacillus plantarum. J. Food Qual. 2018, 2018, 1–12. [Google Scholar] [CrossRef]
- Zhong, H.; Abdullah; Zhao, M.; Tang, J.; Deng, L.; Feng, F. Probiotics-fermented blueberry juices as potential antidiabetic product: Antioxidant, antimicrobial and antidiabetic potentials. J. Sci. Food Agric. 2021, 101, 4420–4427. [Google Scholar] [CrossRef]
- Shikano, A.; Kuda, T.; Shibayama, J.; Toyama, A.; Ishida, Y.; Takahashi, H.; Kimura, B. Effects of Lactobacillus plantarum Uruma-SU4 fermented green loofah on plasma lipid levels and gut microbiome of high-fat diet fed mice. Food Res. Int. 2019, 121, 817–824. [Google Scholar] [CrossRef]
- Hu, J.-P.; Zheng, T.-T.; Zeng, B.-F.; Wu, M.-L.; Shi, R.; Zhang, Y.; Chen, L.-J.; Cheng, W.-J.; Liang, P. Effects of Lactobacillus plantarum FZU3013-Fermented Laminaria japonica on Lipid Metabolism and Gut Microbiota in Hyperlipidaemic Rats. Front. Nutr. 2021, 8, 786571. [Google Scholar] [CrossRef]
- Tsui, C.-Y.; Yang, C.-Y. Evaluation of Semi-Solid-State Fermentation of Elaeocarpus serratus L. Leaves and Black Soymilk by Lactobacillus plantarum on Bioactive Compounds and Antioxidant Capacity. Foods 2021, 10, 704. [Google Scholar] [CrossRef] [PubMed]
- Mirlohi, M.; Babashahi, M.; Ghiasvand, R.; Azadbakht, L.; Mosharaf, L.; Torki-Baghbadorani, S. Effects of probiotic soy milk fermented by lactobacillus plantarum A7 (KC 355240) added with Cuminum Cyminum essential oil on fasting blood glucose levels, serum lipid profile and body weight in diabetic Wistar rats. Int. J. Prev. Med. 2020, 11, 8. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Wu, R.; Zhu, X.; Li, Y.; Li, M.; An, F.; Wu, J. Ameliorative effect of Lactobacillus plantarum WW-fermented soy extract on rat fatty liver via the PPAR signaling pathway. J. Funct. Foods 2019, 60, 103439. [Google Scholar] [CrossRef]
- de la Fuente, B.; Luz, C.; Puchol, C.; Meca, G.; Barba, F.J. Evaluation of fermentation assisted by Lactobacillus brevis POM, and Lactobacillus plantarum (TR-7, TR-71, TR-14) on antioxidant compounds and organic acids of an orange juice-milk based beverage. Food Chem. 2020, 343, 128414. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, M.; Gani, A.; Masoodi, F. Himalayan cheese (Kalari/Kradi) fermented with different probiotic strains: In vitro investigation of nutraceutical properties. LWT 2019, 104, 53–60. [Google Scholar] [CrossRef]
- Moreno-Montoro, M.; Olalla-Herrera, M.; Rufián-Henares, J.; Martínez, R.G.; Miralles, B.; Bergillos, T.; Navarro-Alarcón, M.; Jauregi, P. Antioxidant, ACE-inhibitory and antimicrobial activity of fermented goat milk: Activity and physicochemical property relationship of the peptide components. Food Funct. 2017, 8, 2783–2791. [Google Scholar] [CrossRef]
- Li, Y.; Wu, J.; Cao, C.; Zhu, X.; Sun, X.; Wu, R. Effects of skim milk fermented with Lactobacillus plantarum WW on the constitutions of rats fed a high-fat diet. J. Dairy Sci. 2020, 103, 5019–5029. [Google Scholar] [CrossRef]
- Albano, C.; Morandi, S.; Silvetti, T.; Casiraghi, M.; Manini, F.; Brasca, M. Lactic acid bacteria with cholesterol-lowering properties for dairy applications: In vitro and in situ activity. J. Dairy Sci. 2018, 101, 10807–10818. [Google Scholar] [CrossRef]
- Wang, M.; Lei, M.; Samina, N.; Chen, L.; Liu, C.; Yin, T.; Yan, X.; Wu, C.; He, H.; Yi, C. Impact of Lactobacillus plantarum 423 fermentation on the antioxidant activity and flavor properties of rice bran and wheat bran. Food Chem. 2020, 330, 127156. [Google Scholar] [CrossRef]
- Wu, H.; Rui, X.; Li, W.; Xiao, Y.; Zhou, J.; Dong, M. Whole-grain oats (Avena sativa L.) as a carrier of lactic acid bacteria and a supplement rich in angiotensin I-converting enzyme inhibitory peptides through solid-state fermentation. Food Funct. 2018, 9, 2270–2281. [Google Scholar] [CrossRef]
- Luan, X.; Feng, M.; Sun, J. Effect of Lactobacillus plantarum on antioxidant activity in fermented sausage. Food Res. Int. 2021, 144, 110351. [Google Scholar] [CrossRef]
- Hashemi, S.; Raeisi, S.; Zanardi, E.; Paciulli, M.; Chiavaro, E. Antimicrobial and Antioxidant Activities of Fermented Meat Patty with Lactobacillus Strains. Ital. J. Food Sci. 2018, 30, 268–279. [Google Scholar] [CrossRef]
- Ayyash, M.; Liu, S.-Q.; Al Mheiri, A.; Aldhaheri, M.; Raeisi, B.; Al-Nabulsi, A.; Osaili, T.; Olaimat, A. In vitro investigation of health-promoting benefits of fermented camel sausage by novel probiotic Lactobacillus plantarum: A comparative study with beef sausages. LWT 2018, 99, 346–354. [Google Scholar] [CrossRef]
- Kong, Y.-W.; Feng, M.-Q.; Sun, J. Effects of Lactobacillus plantarum CD101 and Staphylococcus simulans NJ201 on proteolytic changes and bioactivities (antioxidant and antihypertensive activities) in fermented pork sausage. LWT 2020, 133, 109985. [Google Scholar] [CrossRef]
- Wu, Y.; Li, S.; Tao, Y.; Li, D.; Han, Y.; Show, P.L.; Wen, G.; Zhou, J. Fermentation of blueberry and blackberry juices using Lactobacillus plantarum, Streptococcus thermophilus and Bifidobacterium bifidum: Growth of probiotics, metabolism of phenolics, antioxidant capacity in vitro and sensory evaluation. Food Chem. 2021, 348, 129083. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Duan, W.; Zou, J.; Zhou, H.; Liu, C.; Yang, H. Flavor and antioxidant activity improvement of carrot juice by fermentation with Lactobacillus plantarum WZ-01. J. Food Meas. Charact. 2019, 13, 3366–3375. [Google Scholar] [CrossRef]
- Bin Jeon, Y.; Lee, J.; Chang, H.C. Characterization of juice fermented with Lactobacillus plantarum EM and its cholesterol-lowering effects on rats fed a high-fat and high-cholesterol diet. Food Sci. Nutr. 2019, 7, 3622–3634. [Google Scholar] [CrossRef]
- Huang, L.; Feng, M.; Sun, J. Angiotensin-converting enzyme (ACE) inhibitory peptides from fermented sausages inoculated with Lactobacillus plantarum CD101 and Staphylococcus simulans NJ201. Int. J. Food Sci. Technol. 2022, 57, 4985–4997. [Google Scholar] [CrossRef]
- de Oliveira, S.D.; Araújo, C.M.; Borges, G.D.S.C.; Lima, M.D.S.; Viera, V.B.; Garcia, E.F.; de Souza, E.L.; de Oliveira, M.E.G. Improvement in physicochemical characteristics, bioactive compounds and antioxidant activity of acerola (Malpighia emarginata D.C.) and guava (Psidium guajava L.) fruit by-products fermented with potentially probiotic lactobacilli. LWT 2020, 134, 110200. [Google Scholar] [CrossRef]
- Zhang, Z.-P.; Ma, J.; He, Y.-Y.; Lu, J.; Ren, D.-F. Antioxidant and hypoglycemic effects of Diospyros lotus fruit fermented with Microbacterium flavum and Lactobacillus plantarum. J. Biosci. Bioeng. 2018, 125, 682–687. [Google Scholar] [CrossRef]
- Ayyash, M.; Olaimat, A.; Al-Nabulsi, A.; Liu, S.-Q. Bioactive Properties of Novel Probiotic Lactococcus lactis Fermented Camel Sausages: Cytotoxicity, Angiotensin Converting Enzyme Inhibition, Antioxidant Capacity, and Antidiabetic Activity. Korean J. Food Sci. Anim. Resour. 2020, 40, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, S.M.B.; Gholamhosseinpour, A. Fermentation of table cream by Lactobacillus plantarum strains: Effect on fungal growth, aflatoxin M1 and ochratoxin A. Int. J. Food Sci. Technol. 2018, 54, 347–353. [Google Scholar] [CrossRef]
- Dan, T.; Chen, H.; Li, T.; Tian, J.; Ren, W.; Zhang, H.; Sun, T. Influence of Lactobacillus plantarum P-8 on Fermented Milk Flavor and Storage Stability. Front. Microbiol. 2019, 9, 3133. [Google Scholar] [CrossRef] [PubMed]
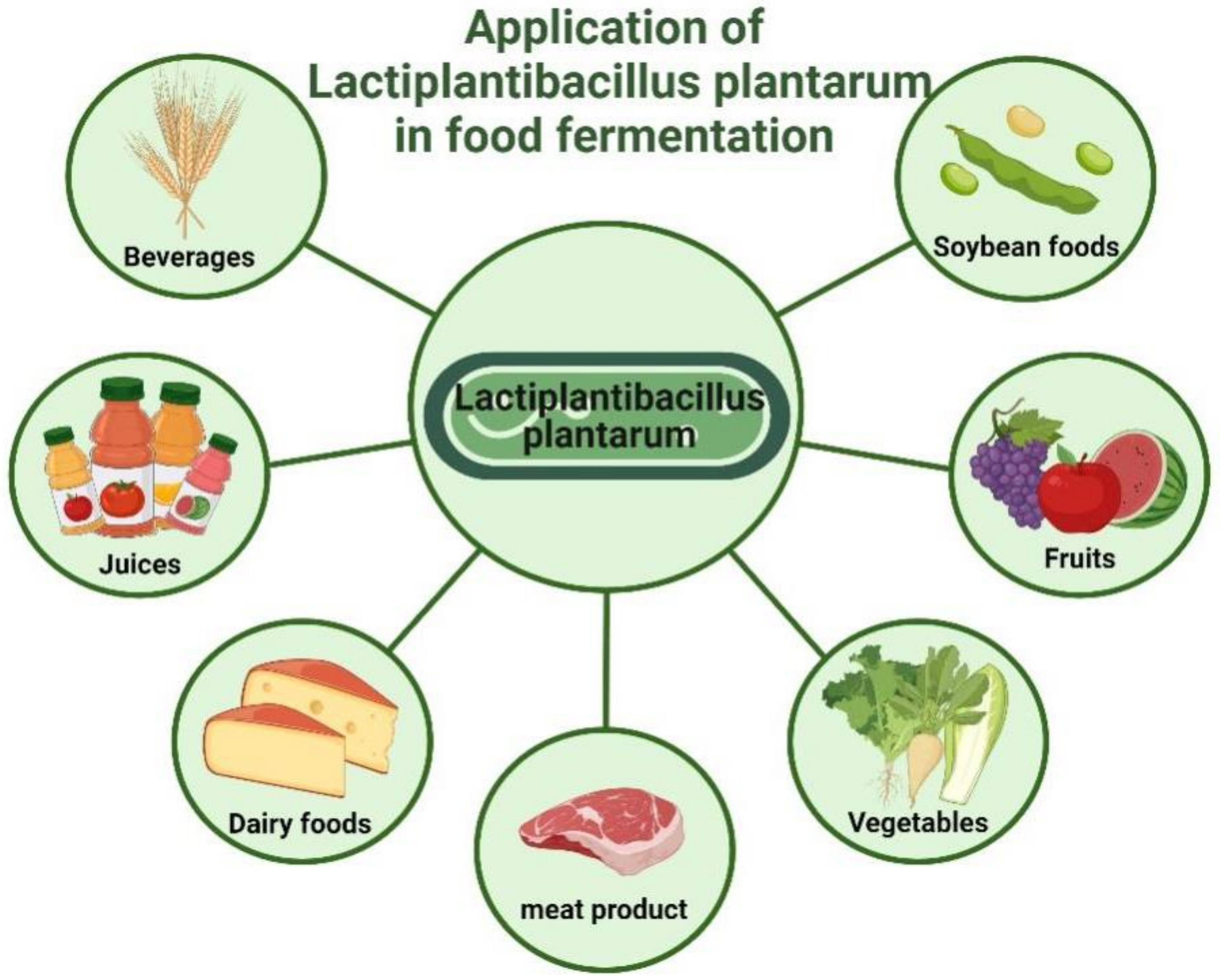
| Types of Food | Fermented Food | Lb. plantarum | Function | Mechanism | References |
|---|---|---|---|---|---|
| Fruit and vegetable juice | charantia juice | Lb. plantarum NCU116 | antioxidant | increase the content of phenolic compounds and promote the biotransformation to provide stronger antioxidant properties | [124] |
| papaya juice | Lb. plantarum GIM1.140 | antioxidant | increase the content of total flavonoids and improve inhibition of DPPH free radicals | [125] | |
| blueberry juice | A variety of mixed strains including Lb. plantarum | anti-diabetes | maintain glucose homeostasis and promote glucose consumption | [126] | |
| green loofah | Lb. plantarum SU4 | cholesterol lowering | high bile acid lowering capacity in vitro and in vivo to promote cholesterol consumption | [127] | |
| Aquatic product | laminaria japonica | Lb. plantarum FZU3013 | cholesterol lowering | reduce expression levels of genes involved in lipid metabolism and bile acid homeostasis to promote cholesterol consumption | [128] |
| Soybean products | black Soymilk | Lb. plantarum BCRC 10357 | antioxidant | increase the ferric reducing antioxidant capacity | [129] |
| soy milk added with cuminum cyminum essential oil | Lb. plantarum A7 (KC 355240) | anti-diabetes and cholesterol lowering | significantly reduces postprandial serum glucose concentrations and TG levels. | [130] | |
| soy extract | Lb. plantarum WW | cholesterol lowering | regulate the expression levels of genes involved in lipid metabolism and oxidation-reduction processes to promote cholesterol catabolism | [131] | |
| Dairy products | orange juice-milk based beverage | Lb. plantarum (TR-7, TR-71, TR-14) | antioxidant | increase the content of carotenoids and the total antioxidant activity | [132] |
| kalari cheese | Lb. plantarum NCDC 012 | anti-diabetes | produce a variety of bioactive peptides to enhance the inhibitory activity of α-amylase and α-glucosidase, and inhibit carbohydrate decomposition to lower glucose | [133] | |
| goat milk | Lb. plantarum C4 | blood pressure lowering | enhance the ACE inhibitory activity by fermentation | [134] | |
| skim milk | Lb. plantarum WW | cholesterol lowering | regulate the intestinal flora and lower cholesterol levels | [135] | |
| cheese | Lb. plantarum VC213 | cholesterol lowering | significantly lower cholesterol content than before fermentation | [136] | |
| Cereal grains | rice bran and wheat bran | Lb. plantarum NCU116 | antioxidant | enhance the hydroxyl radical-scavenging activity and the oxygen radical-quenching activity | [137] |
| whole-grain oats | Lb. plantarum B1-6 | blood pressure lowering | present higher ACE inhibitory activities | [138] | |
| Meat products | Chinese fermented sausages | Lb. plantarum CD101 | antioxidant | reduce pH, and promote the formation of antioxidant peptides | [139] |
| fermented meat patty | Lb. plantarum PTCC 1745 | antioxidant | radical scavenging activity significantly higher than before fermentation | [140] | |
| fermented camel sausages | Lb. plantarum KX881772 | anti-diabetes | higher α-amylase and higher α-glucosidase inhibitions to control diabetes by reducing carbohydrate hydrolysis | [141] | |
| fermented sausage | Lb. plantarum CD101 | blood pressure lowering | significantly increase the ACE inhibitory activity | [142] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Wu, J.; Tian, Z.; Si, Y.; Chen, H.; Gan, J. The Mechanisms of the Potential Probiotic Lactiplantibacillus plantarum against Cardiovascular Disease and the Recent Developments in its Fermented Foods. Foods 2022, 11, 2549. https://doi.org/10.3390/foods11172549
Wang Z, Wu J, Tian Z, Si Y, Chen H, Gan J. The Mechanisms of the Potential Probiotic Lactiplantibacillus plantarum against Cardiovascular Disease and the Recent Developments in its Fermented Foods. Foods. 2022; 11(17):2549. https://doi.org/10.3390/foods11172549
Chicago/Turabian StyleWang, Zhe, Juanjuan Wu, Zichen Tian, Yue Si, Hao Chen, and Jing Gan. 2022. "The Mechanisms of the Potential Probiotic Lactiplantibacillus plantarum against Cardiovascular Disease and the Recent Developments in its Fermented Foods" Foods 11, no. 17: 2549. https://doi.org/10.3390/foods11172549
APA StyleWang, Z., Wu, J., Tian, Z., Si, Y., Chen, H., & Gan, J. (2022). The Mechanisms of the Potential Probiotic Lactiplantibacillus plantarum against Cardiovascular Disease and the Recent Developments in its Fermented Foods. Foods, 11(17), 2549. https://doi.org/10.3390/foods11172549