
Whey Protein Hydrolysates of Sheep/Goat Origin Produced by the Action of Trypsin without pH Control: Degree of Hydrolysis, Antihypertensive Potential and Antioxidant Activities



Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrates, Hydrolysis and Sampling
2.2. Analyses
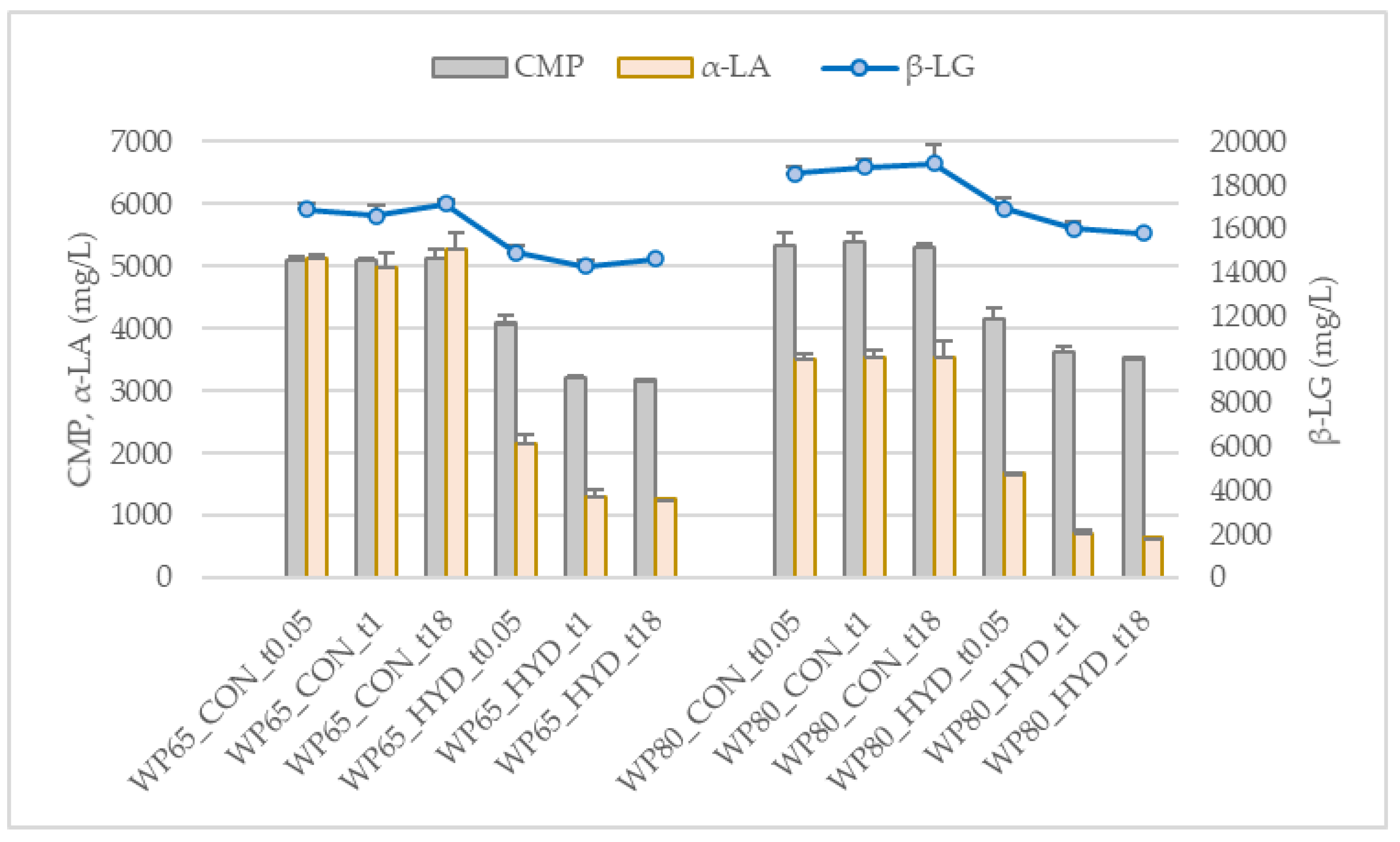
2.2.1. Protein Composition of the Substrates
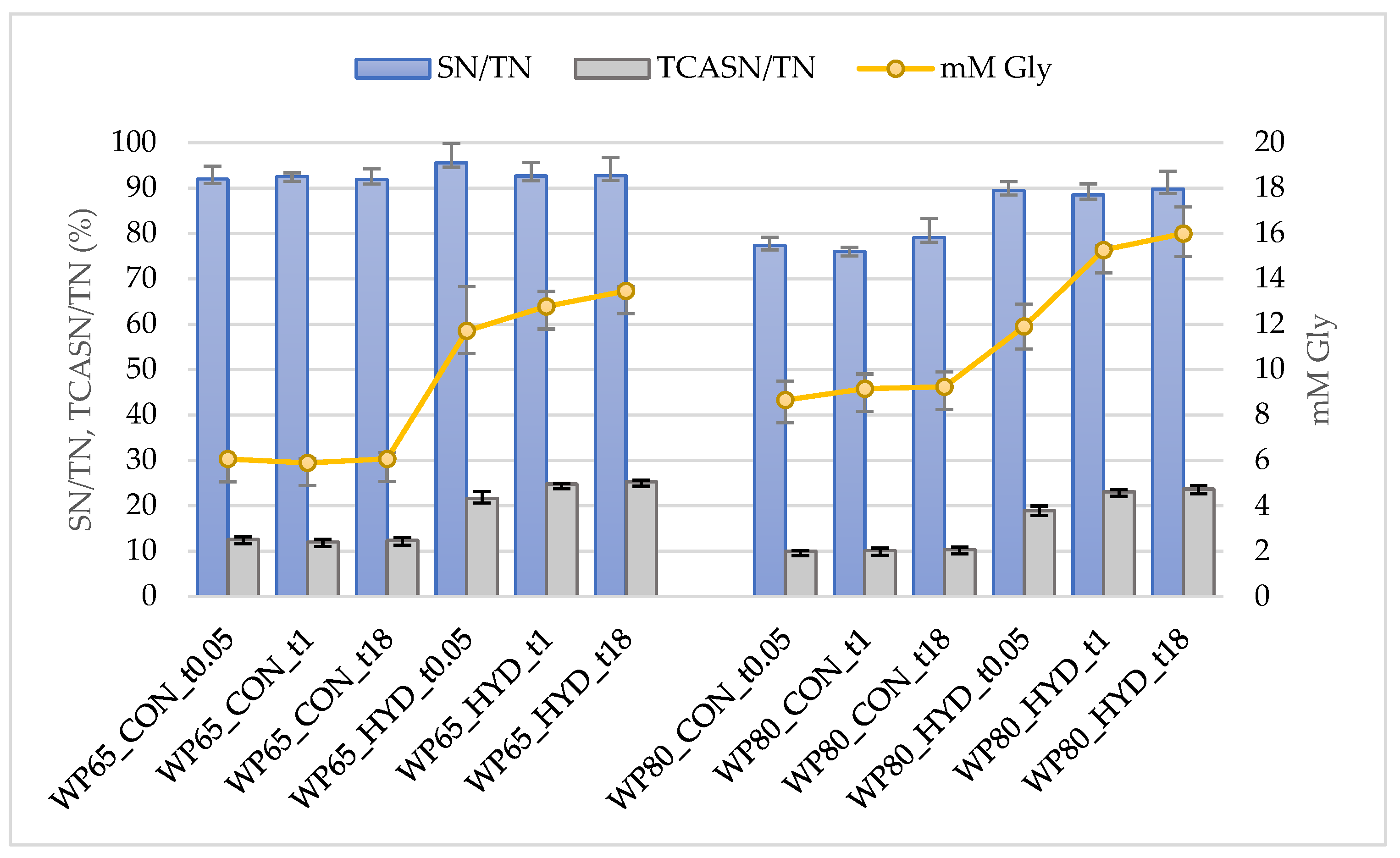
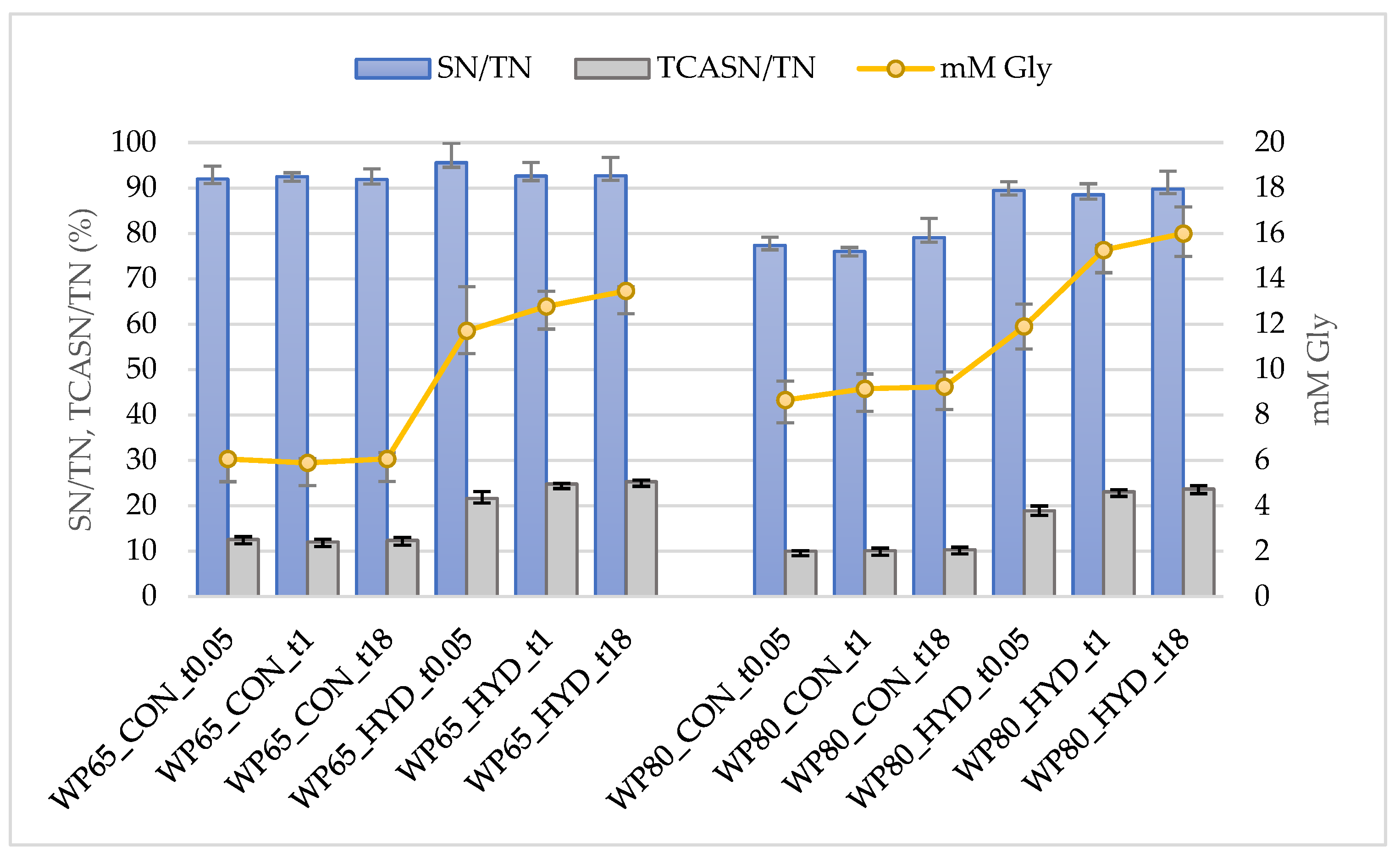
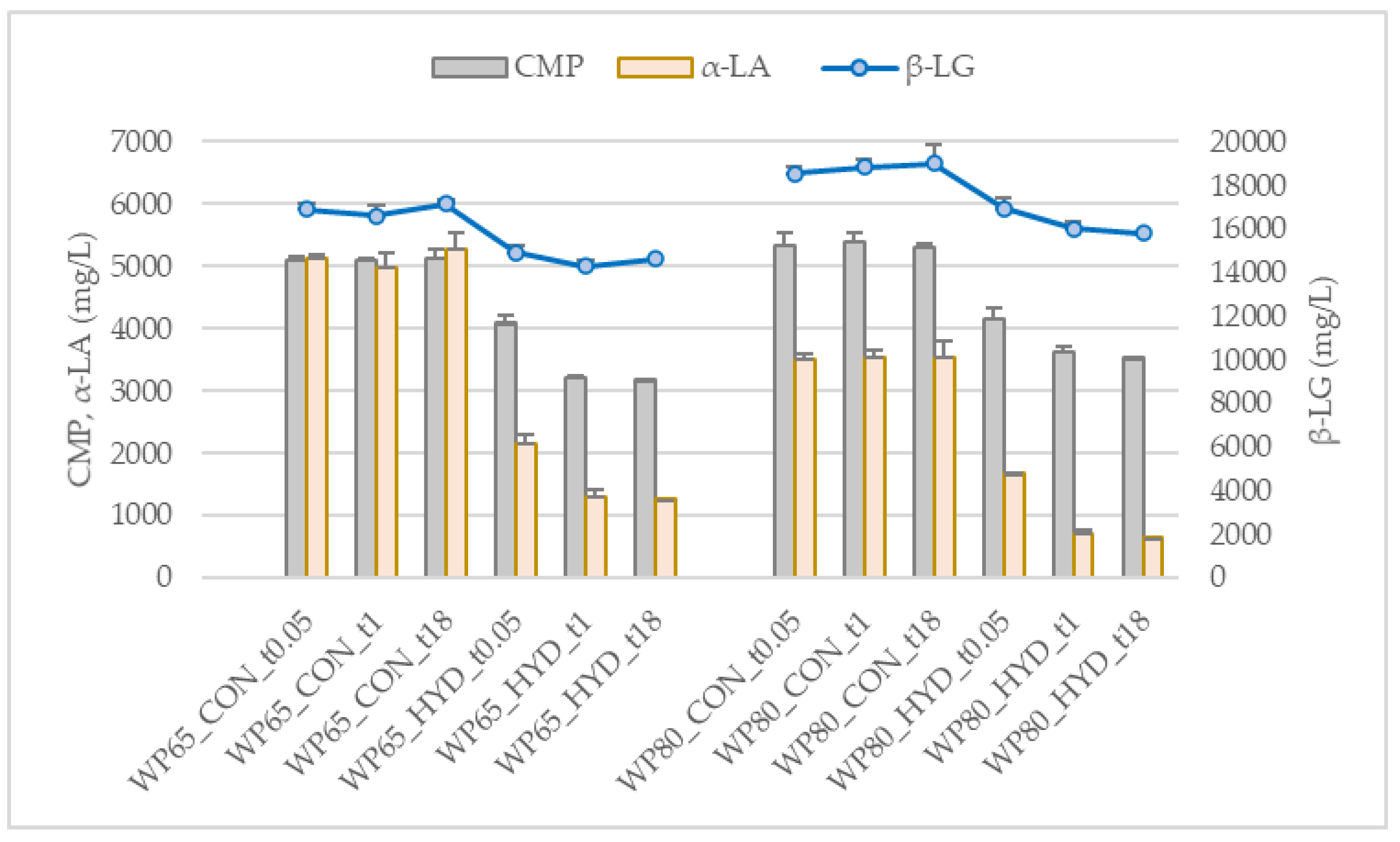
2.2.2. Degree and Extent of Hydrolysis
- i.
- The quantitative changes of nitrogen soluble at pH 4.6 (SN) and nitrogen soluble at 12% trichloroacetic acid (TCASN), as described in Section 2.2.1. The degree of hydrolysis (DH) based on SN changes was estimated according to DH_SN (%) = 100 × (SN_HYD-SN_CON)/(TN_CON-TCASN_CON), where HYD is a hydrolysate and CON is the respective control. The expression TN_CON-TCASN_CON corresponds to protein N content. The DH based on TCASN changes (DH_TCASN) was estimated accordingly.
- ii.
- Changes of free amino groups was determined by the TNBS (picrylsulfonic acid solution, P2297, Sigma Aldrich, St. Louis, MO, USA) method of Adler-Nissen [19], with some modifications. One hundred μL hydrolysate was diluted with 900 μL of 1% SDS (sodium dodecyl sulfate); 250 μL of this dilution were diluted further with 2 mL 0.21 M phosphate buffer, pH 8.0. After the addition of 2 mL 0.05%, w/w TNBS, the mixture was incubated in the dark at 50 °C for 60 min. The reaction was stopped by the addition of 4 mL 0.1 M HCl. After cooling down for 30 min, the absorbance at 340 nm was determined. Glycine was used for the construction of the standard curve. The estimation of the DH was according to Spellman et al. [20]: DH_TNBS (%) = 100 × [(AN2-AN1)/123.3]. AN1 is the amino acid nitrogen content of the control WPC solution without trypsin (CON) expressed as mg/g protein and was estimated using the reference curve. Accordingly, AN2 is the content of the respective hydrolysate (HYD). The value 123.3 mg/g is the nitrogen content of the peptide bonds of the whey proteins in the substrate.
2.2.3. Biofunctionality Assays
2.2.4. Statistical Analysis
3. Results and Discussion
3.1. Degree and Extent of Hydrolysis
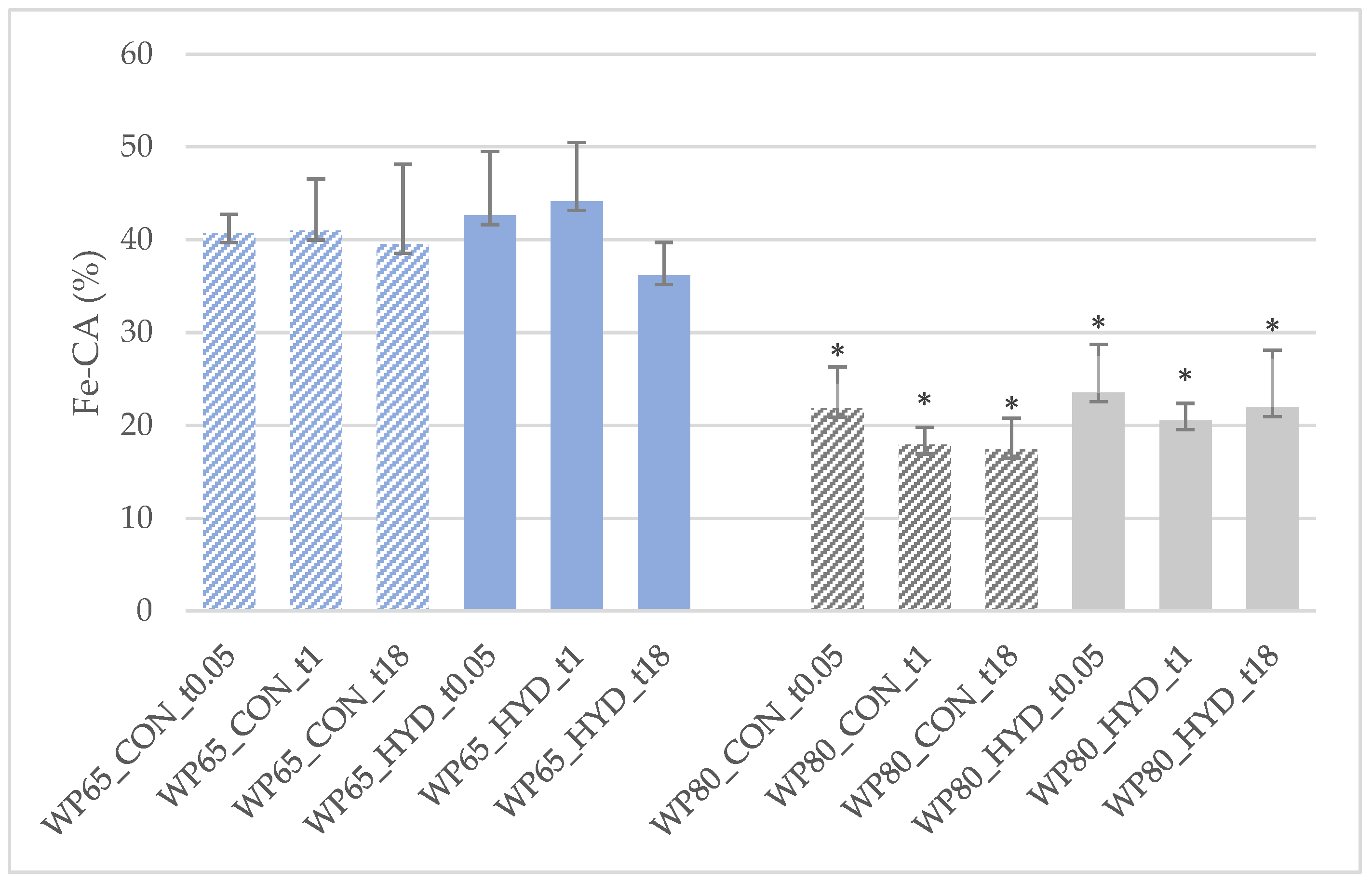
3.2. Biofunctional Potential
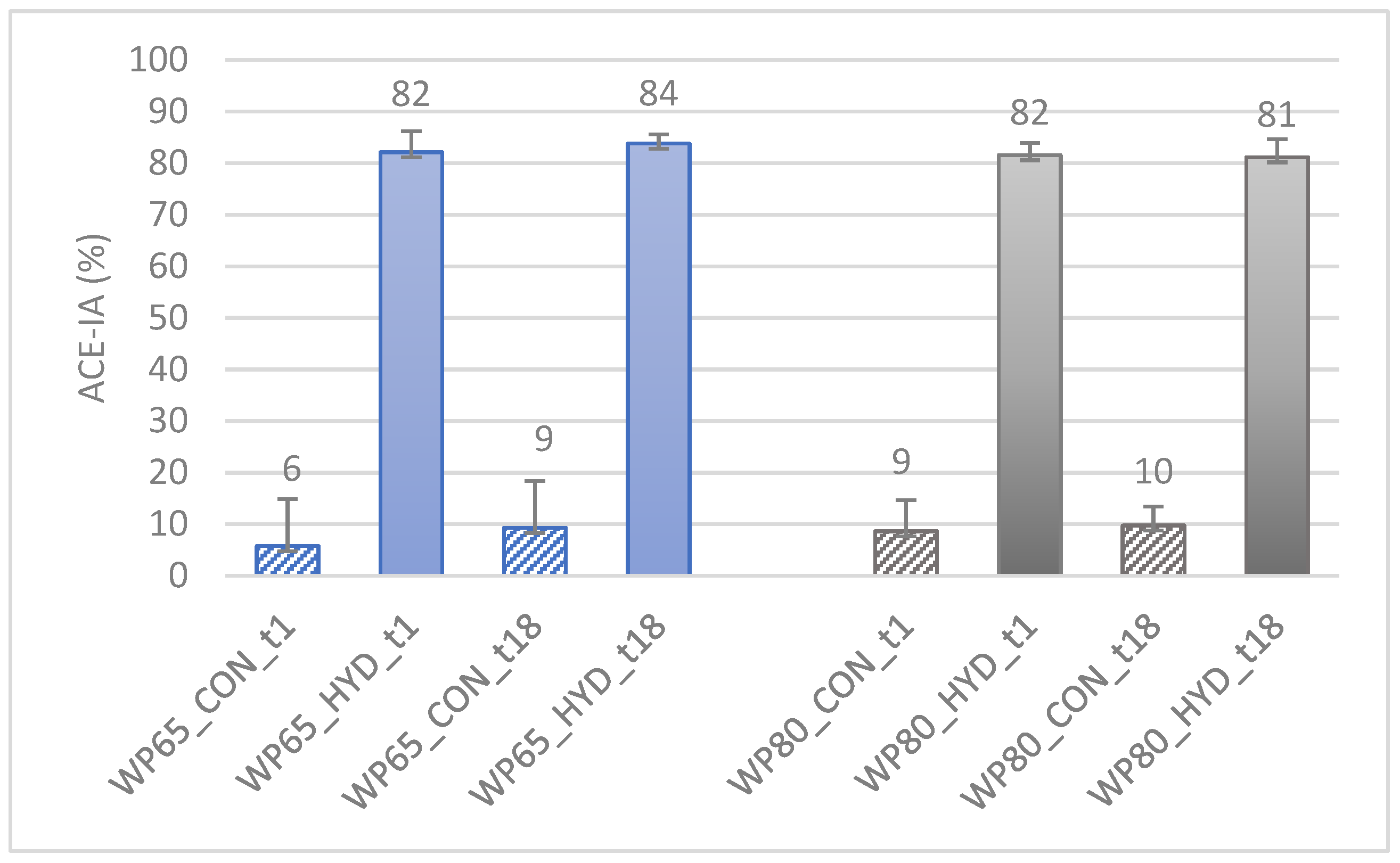
3.2.1. Peptidyl Dipeptide Hydrolase-Inhibitory Activity (ACE-IA)
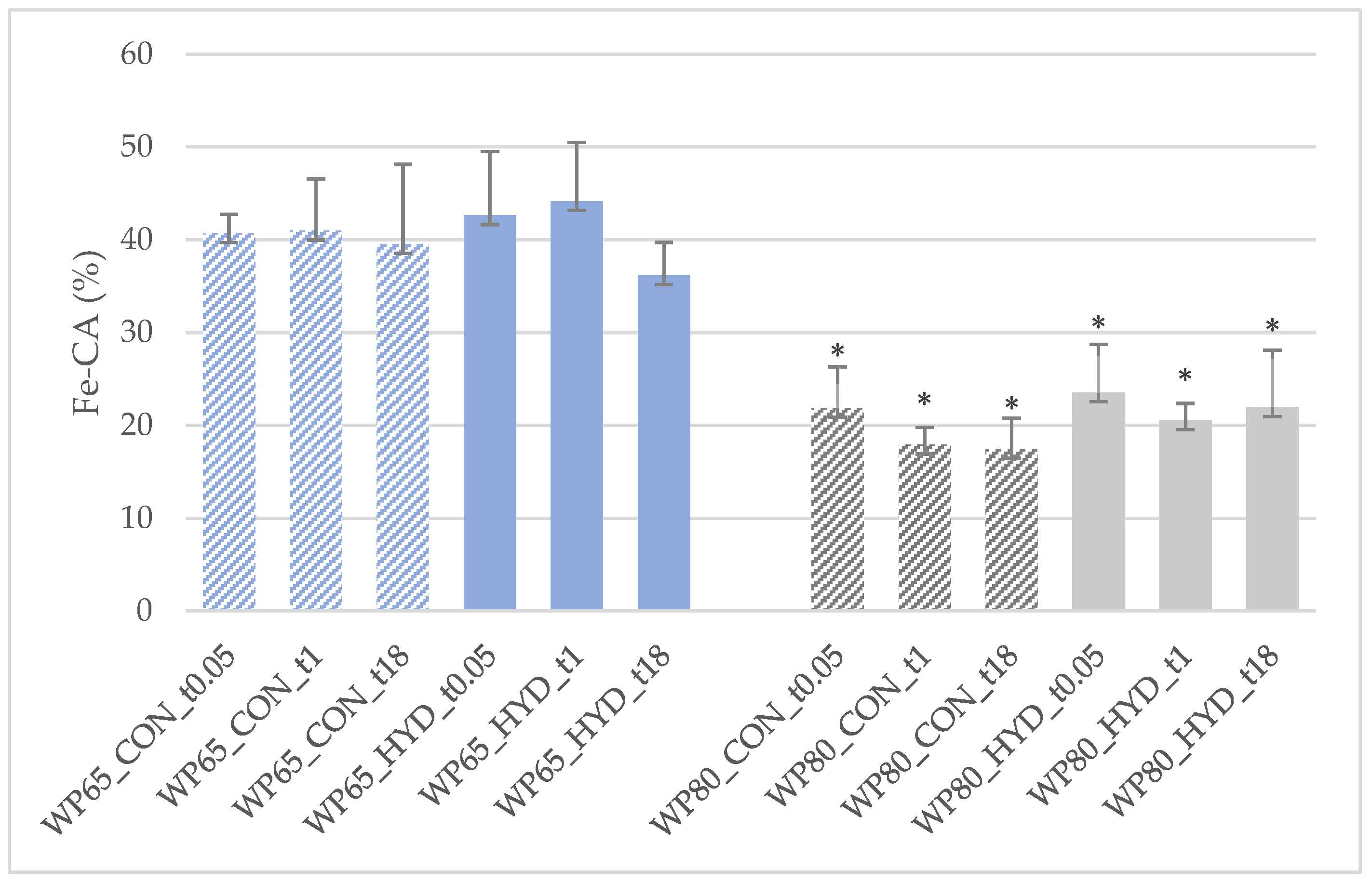
3.2.2. Antioxidant Activities
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castro, R.J.S.d.; Domingues, M.A.F.; Ohara, A.; Okuro, P.K.; Santos, J.G.d.; Brexo, R.P.; Sato, H.H. Whey protein as a key component in food systems: Physicochemical properties, production technologies and applications. Food Struct. 2017, 14, 17–29. [Google Scholar] [CrossRef]
- Zhao, C.; Chen, N.; Ashaolu, T.J. Whey proteins and peptides in health-promoting functions—A review. Intern. Dairy J. 2022, 126, 105269. [Google Scholar] [CrossRef]
- Foegeding, E.A.; Davis, J.P.; Doucet, D.; McGuffey, M.K. Advances in modifying and understanding whey protein functionality. Trends Food Sci. Technol. 2002, 13, 151–159. [Google Scholar] [CrossRef]
- Pires, A.F.; Marnotes, N.G.; Rubio, O.D.; Garcia, A.C.; Pereira, C.D. Dairy by-products: A review on the valorization of whey and second cheese whey. Foods 2021, 10, 1067. [Google Scholar] [CrossRef]
- Cheison, S.C.; Kulozik, U. Impact of the environmental conditions and substrate pre-treatment on whey protein hydrolysis: A review. Critical Rev. Food Sci. Nutr. 2017, 57, 418–453. [Google Scholar] [CrossRef]
- Guo, M.; Shen, X. Modifications of whey protein. In Whey Protein Production, Chemistry, Functionality, and Applications, 1st ed.; Guo, M., Ed.; Wiley: West Sussex, UK, 2019; pp. 205–226. [Google Scholar] [CrossRef]
- Dullius, A.; Goettert, M.I.; de Souza, C.F.V. Whey protein hydrolysates as a source of bioactive peptides for functional foods—Biotechnological facilitation of industrial scale-up. J. Funct. Foods 2018, 42, 58–74. [Google Scholar] [CrossRef]
- Qian, Z.Y.; Jollès, P.; Migliore-Samour, D.; Schoentgen, F.; Fiat, A.M. Sheep kappa-casein peptides inhibit platelet aggregation. Biochim. Biophys. Acta 1995, 1244, 411–417. [Google Scholar] [CrossRef]
- Manso, M.A.; Escudero, C.; Alijo, M.; Lopez-Fandino, R. Platelet aggregation inhibitory activity of bovine, ovine, and caprine k -casein macropeptides and their tryptic hydrolysates. J. Food Prot. 2002, 65, 1992–1996. [Google Scholar] [CrossRef]
- Chobert, J.M.; El-Zahar, K.; Sitohy, M.; Dalgalarrondo, M.; Metro, F.; Choiset, Y.; Haertlé, T. Angiotensin I-converting-enzyme (ACE)-inhibitory activity of tryptic peptides of ovine b-lactoglobulin and of milk yoghurts obtained by using different starters. Lait 2005, 85, 141–152. [Google Scholar] [CrossRef]
- Hernandez-Ledesma, B.; Recio, I.; Ramos, M.; Amigo, L. Preparation of ovine and caprine β-lactoglobulin hydrolysates with ACE-inhibitory activity. Identification of active peptides from caprine β-lactoglobulin hydrolyzed with thermolysin. Int. Dairy J. 2002, 12, 805–812. [Google Scholar] [CrossRef]
- Correa, A.P.F.; Daroit, D.J.; Fontoura, R.; Meira, S.M.M.; Segalin, J.; Brandelli, A. Hydrolysates of sheep cheese whey as a source of bioactive peptides with antioxidant and angiotensin-converting enzyme inhibitory activities. Peptides 2014, 61, 48–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espejo-Carpio, F.; Pérez-Gálvez, R.; Guadix, E.; Guadix, A. Optimisation of the hydrolysis of goat milk protein for the production of ACE-inhibitory peptides. J. Dairy Res. 2013, 80, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Espejo-Carpio, F.; Pérez-Gálvez, R.; Guadix, A.; Guadix, E. Artificial Neuronal Networks (ANN) to model the hydrolysis of goat milk protein by subtilisin and trypsin. J. Dairy Res. 2018, 85, 339–346. [Google Scholar] [CrossRef]
- Du, X.; Jing, H.; Wang, L.; Huang, X.; Wang, X.; Wang, H. Characterization of structure, physicochemical properties, and hypoglycemic activity of goat milk whey protein hydrolysate processed with different proteases. LWT Food Sci. Technol. 2022, 159, 113257. [Google Scholar] [CrossRef]
- Moatsou, G.; Hatzinaki, A.; Kandarakis, I.; Anifantakis, E. Nitrogenous fractions during the manufacture of whey protein concentrates from Feta cheese whey. Food Chem. 2003, 81, 209–217. [Google Scholar] [CrossRef]
- Roumanas, D.; Moatsou, G.; Zoidou, E.; Sakkas, L.; Moschopoulou, E. Effect of enrichment of bovine milk with whey proteins on biofunctional and rheological properties of low fat yoghurt-type products. Curr. Res. Nutr. Food Sci. 2016, 4, 105–113. [Google Scholar] [CrossRef]
- Sakkas, L.; Tzevdou, M.; Zoidou, E.; Gkotzia, E.; Karvounis, A.; Samara, A.; Taoukis, P.; Moatsou, G. Yoghurt-type gels from skim sheep milk base enriched with whey protein concentrate hydrolysates and processed by heating or high hydrostatic pressure. Foods 2019, 8, 342. [Google Scholar] [CrossRef] [Green Version]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef] [PubMed]
- Spellman, D.; McEvoy, E.; O’Cuinn, G.; FitzGerald, R.J. Proteinase and exopeptidase hydrolysis of whey protein: Comparison of the TNBS, OPA and pH stat methods for quantification of degree of hydrolysis. Int. Dairy J. 2003, 13, 447–453. [Google Scholar] [CrossRef]
- Moschopoulou, E.; Sakkas, L.; Zoidou, E.; Theodorou, G.; Sgouridou, E.; Kalathaki, C.; Liarakou, A.; Chatzigeorgiou, A.; Politis, I.; Moatsou, G. Effect of milk kind and storage on the biochemical, textural and biofunctional characteristics of set-type yoghurt. Intern. Dairy J. 2018, 77, 47–55. [Google Scholar] [CrossRef]
- Le Maux, S.; Nongonierma, A.B.; Barre, C.; FitzGerald, R.J. Enzymatic generation of whey protein hydrolysates under pH-controlled and non pH-controlled conditions: Impact on physicochemical and bioactive properties. Food Chem. 2016, 199, 246–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Carvalho, N.C.; Pessato, T.B.; Fernandes, L.G.R.; De Lima Zollner, R.; Netto, F.M. Physicochemical characteristics and antigenicity of whey protein hydrolysates obtained with and without pH control. Int. Dairy J. 2017, 71, 24–34. [Google Scholar] [CrossRef]
- Kleekayai, T.; Le Gouic, A.V.; Deracinois, B.; Cudennec, B.; FitzGerald, R.J. In vitro characterisation of the antioxidative properties of whey protein hydrolysates generated under pH- and non pH-controlled conditions. Foods 2020, 9, 582. [Google Scholar] [CrossRef]
- Martin-Orue, C.; Henry, G.; Bouhallab, S. Tryptic hydrolysis of κ-caseinomacropeptide: Control of the enzymic reaction in a continuous membrane reactor. Enzyme Microbial Technol. 1999, 24, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Furet, J.P.; Mercier, J.C.; Gaye, P.; Huedelahaie, D.; Vilotte, J.L. Nucleotide sequence of ovine κ-casein cDNA. Nucleic Acids Res. 1990, 18, 5286. [Google Scholar] [CrossRef] [Green Version]
- Coll, A.; Folch, J.M.; Sànchez, A. Nucleotide sequence of the goat kappa-casein cDNA. J. Anim Sci. 1993, 71, 2833. [Google Scholar] [CrossRef] [Green Version]
- Cheison, S.C.; Leeb, E.; Toro-Siera, J.; Kulozik, U. Influence of hydrolysis temperature and pH on the selective hydrolysis of whey proteins by trypsin and potential recovery of native alpha-lactalbumin. Intern. Dairy J. 2011, 21, 166–171. [Google Scholar] [CrossRef]
- Hinnenkamp, C.; Ismail, B.P.A. Proteomics approach to characterizing limited hydrolysis of whey protein concentrate. Food Chem. 2021, 350, 129235. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.M.P.L.V.O.; Pinho, O.; Mota, M.V.; Tavares, P.; Pereira, A.; Gonçalves, M.P.; Torres, D.; Rocha, C.; Teixeira, J.A. Preparation of ingredients containing an ACE-inhibitory peptide by tryptic hydrolysis of whey protein concentrates. Intern. Dairy J. 2007, 17, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Fernández, A.; Kelly, P. pH-stat vs. free-fall pH techniques in the enzymatic hydrolysis of whey proteins. Food Chem. 2016, 199, 409–415. [Google Scholar] [CrossRef]
- Sáez, L.; Murphy, E.; FitzGerald, R.J.; Kelly, P. Exploring the use of a modified high-temperature, short-time continuous heat exchanger with extended holding time (HTST-EHT) for thermal inactivation of trypsin following selective enzymatic hydrolysis of the β-lactoglobulin fraction in whey protein isolate. Foods 2019, 8, 367. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Ledesma, B.; Ramos, M.; Gómez-Ruiz, J.A. Bioactive components of ovine and caprine cheese whey. Small Rumin. Res. 2011, 101, 196–204. [Google Scholar] [CrossRef] [Green Version]
- Gruppi, A.; Dermiki, M.; Spigno, G.; FitzGerald, R.J. Impact of enzymatic hydrolysis and heat inactivation on the physicochemical properties of milk protein hydrolysates. Foods 2022, 11, 516. [Google Scholar] [CrossRef] [PubMed]
- Walstra, P.; Wouters, J.T.M.; Geurts, T.J. Serum proteins. In Dairy Science and Technology, 2nd ed.; CRC Press Taylor and Francis Group: Boca Raton, FL, USA, 2006; pp. 76–77. [Google Scholar] [CrossRef]
- Guo, M.; Wang, C. Chemistry of whey proteins. In Whey Protein Production, Chemistry, Functionality, and Applications, 1st ed.; Guo, M., Ed.; Wiley: West Sussex, UK, 2019; pp. 39–66. [Google Scholar] [CrossRef]
- Cheison, S.C.; Lai, M.-Y.; Leeb, E.; Kulozik, U. Hydrolysis of b-lactoglobulin by trypsin under acidic pH and analysis of the hydrolysates with MALDI–TOF– MS/MS. Food Chem. 2011, 125, 1241–1248. [Google Scholar] [CrossRef]
- Cheison, S.C.; Schmitt, M.; Leeb, E.; Letzel, T.; Kulozik, U. Influence of the temperature and the degree of hydrolysis on the peptide composition of trypsin hydrolysates of b-lactoglobulin: Analysis by LC-ESI-TOF/MS. Food Chem. 2010, 121, 457–467. [Google Scholar] [CrossRef]
- Deng, Y.; Butre, C.I.; Wierenga, P.A. Influence of substrate concentration on the extent of protein enzymatic hydrolysis. Intern. Dairy J. 2018, 86, 39–48. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, R.; Zhang, J.; Zhou, P. Heat-induced denaturation and bioactivity changes of whey proteins. Intern. Dairy J. 2021, 123, 105175. [Google Scholar] [CrossRef]
- Morgan, F.; Léonil, J.; Mollé, D.; Bouhallab, S. Nonenzymatic lactosylation of bovine β-lactoglobulin under mild heat treatment leads to structural heterogeneity of the glycoforms. Biochem. Biophys. Res. Commun. 1997, 236, 413–417. [Google Scholar] [CrossRef]
- Corzo-Martínez, M.; Soria, A.C.; Belloque, J.; Villamiel, M.; Moreno, F.J. Effect of glycation on the gastrointestinal digestibility and immunoreactivity of bovine β-lactoglobulin. Intern. Dairy J. 2010, 20, 742–752. [Google Scholar] [CrossRef] [Green Version]
- Leeb, E.; Götz, A.; Letzel, T.; Cheison, S.C.; Kulozik, U. Influence of denaturation and aggregation of β-lactoglobulin on its tryptic hydrolysis and the release of functional peptides. Food Chem. 2015, 187, 545–554. [Google Scholar] [CrossRef]
- Petit, J.; Moreau, A.; Ronse, G.; Debreyne, P.; Bouvier, L.; Blanpain-Avet, P.; Jeantet, R.; Delaplace, G. Role of whey components in the kinetics and thermodynamics of β-lactoglobulin unfolding and aggregation. Food Bioprocess Technol. 2016, 9, 1367–1379. [Google Scholar] [CrossRef]
- Madureira, A.R.; Tavares, T.; Gomes, A.M.P.; Pintado, M.E.; Malcata, F.X. Invited review: Physiological properties of bioactive peptides obtained from whey proteins. J. Dairy Sci. 2010, 93, 437–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, G.; Huang, J.; Bao, C.; Meng, J.; Chen, H.; Cao, J. Effect of different proteases on the degree of hydrolysis and angiotensin I-converting enzyme-inhibitory activity in goat and cow milk. Biomolecules 2018, 8, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullally, M.M.; Meisel, H.; FitzGerald, R.J. Angiotensin-I-converting enzyme inhibitory activities of gastric and pancreatic proteinase digests of whey proteins. Int. Dairy J. 1997, 7, 299–303. [Google Scholar] [CrossRef]
- Pihlanto-Leppala, A.; Koskinen, P.; Piilola, K.; Tupasela, T.; Korhonen, H. Angiotensin-I converting enzyme inhibitory properties of whey protein digests: Concentration and characterization of active peptides. J. Dairy Res. 2000, 67, 53–64. [Google Scholar] [CrossRef]
- Manso, M.A.; López-Fandiño, R. Angiotensin I converting enzyme—Inhibitory activity of bovine, ovine, and caprine κ-casein macropeptides and their tryptic hydrolysates. J. Food Prot. 2003, 66, 1686–1692. [Google Scholar] [CrossRef]
- Lindmark-Mansson, H.; Akesson, B. Antioxidative factors in milk. British J. Nutr. 2000, 84, S103–S110. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.T.; Nadeem, M.; Imran, M.; Ullah, R.; Ajmal, M.; Jaspal, M.H. Antioxidant properties of milk and dairy products: A comprehensive review of the current knowledge. Lipids Health Dis. 2019, 18, 41. [Google Scholar] [CrossRef] [Green Version]
- Conway, V.; Gauthier, S.F.; Pouliot, Y. Antioxidant activities of buttermilk proteins, whey proteins, and their enzymatic hydrolysates. J. Agric. Food Chem. 2013, 61, 364–372. [Google Scholar] [CrossRef]
- Cichosz, G.; Czeczot, H.; Ambroziak, A.; Bielecka, M.M. Natural antioxidants in milk and dairy products. Int. J. Dairy Technol. 2017, 70, 165–178. [Google Scholar] [CrossRef]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Krunic, T.Z.; Rakin, M.B. Enriching alginate matrix used for probiotic encapsulation with whey protein concentrate or its trypsin-derived hydrolysate: Impact on antioxidant capacity and stability of fermented whey-based beverages. Food Chem. 2022, 370, 130931. [Google Scholar] [CrossRef] [PubMed]
- Vavrusova, M.; Pindstrup, H.; Johansen, L.B.; Andersen, M.L.; Andersen, H.J.; Skibsted, L.H. Characterisation of a whey protein hydrolysate as antioxidant. Intern. Dairy J. 2015, 47, 86–93. [Google Scholar] [CrossRef]
- Giblin, L.; Yalçın, A.S.; Biçim, G.; Krämer, A.C.; Chen, Z.; Callanan, M.J.; Arranz, E.; Davies, M.J. Whey proteins: Targets of oxidation, or mediators of redox protection. Free Radic. Res. 2019, 53, 1136–1152. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Code 2 | Trypsin (0.25% on Protein) | Time (h) 3 | Heating (68 °C for 10 min) 4 | Storage (h) 5 |
|---|---|---|---|---|
| WP(65 or 80)_CON_t0.05 | - | 0.05 | + | - |
| WP(65 or 80)_CON_t1 | - | 1 | + | - |
| WP(65 or 80)_CON_t18 | - | 1 | + | 17 |
| WP(65 or 80)_HYD_t0.05 | + | 0.05 | + | - |
| WP(65 or 80)_HYD_t1 | + | 1 | + | - |
| WP(65 or 80)_HYD_t18 | + | 1 | + | 17 |
| WPC | TP g/100 g | SN/TN % | TCASN/TN % | CMP g/100 g | α-LA 1 g/100 g | β-LG 1 g/100 g |
|---|---|---|---|---|---|---|
| WP65 | 61.25 ± 1.98 | 91.2 ± 2.62 | 12.9 ± 1.73 | 10.1 ± 0.32 | 10.4 ± 0.45 | 33.0 ± 3.35 |
| WP80 | 73.27 ± 2.18 | 76.5 ± 2.74 | 10.6 ± 1.90 | 10.3 ± 0.58 | 6.7 ± 0.33 | 36.64 ± 1.04 |
| Time (h) | WP65 | WP80 | ||
|---|---|---|---|---|
| Control | +Trypsin | Control | +Trypsin | |
| 0.05 1 | 6.28 ± 0.03 | 6.28 ± 0.03 | 6.11 ± 0.10 | 6.11 ± 0.09 |
| 0.5 1 | 5.92 ± 0.08 | 5.85 ± 0.07 | 5.75 ± 0.07 | 5.65 ± 0.07 |
| 1 1,2 | 6.07 ± 0.19 | 5.95 ± 0.19 | 5.88 ± 0.18 | 5.84 ± 0.24 |
| 18 3 | 6.35 ± 0.07 | 6.29 ± 0.12 | 6.19 ± 0.10 | 6.17 ± 0.12 |
| Hydrolysis Stage | DH_TCASN | DH_TNBS | EH_β-lg | EH_α-LA | EH_CMP |
|---|---|---|---|---|---|
| WP65_HYD_t0.05 | 11 ± 2.4 a | 10.6 ± 1.8 a,* | 11.6 ± 2.3 | 57.6 ± 4.6 a | 19.9 ± 2.9 a |
| WP65_HYD_t1 | 17.1 ± 0.3 b | 13.7 ± 1.5 b,* | 14 ± 1.3 | 73.9 ± 2.6 b | 37 ± 2.6 b |
| WP65_HYD_t18 | 17± 1.2 b | 14.8 ± 0.3 b,* | 14.6 ± 3.7 | 75.4 ± 3.3 b | 37.8 ± 0.9 b |
| WP80_HYD_t0.05 | 10.9 ± 1.2 a | 5.9 ± 0.9 a | 8.6 ± 2.9 a | 51.9 ± 6.7 a | 21.9 ± 3.7 a |
| WP80_HYD_t1 | 16.7 ± 1 b | 10.3 ± 1.1 b | 14.9 ± 1.7 b | 79.9 ± 1 b,* | 32.7 ± 4.8 b |
| WP80_HYD_t18 | 17.4 ± 0.5 b | 11.3 ± 2 b | 16.9 ± 1.4 b | 82 ± 1.3 b,* | 33.6 ± 0.3 b |
| DH_TCASN | DH_TNBS | ED_β-LG | ED_α-LA | ED_CMP | |
|---|---|---|---|---|---|
| DH_TCASN | 0.66 | 0.72 | 0.91 | 0.89 | |
| DH_TNBS | 0.66 | 0.58 | 0.61 | 0.73 | |
| ED_β-LG | 0.72 | 0.58 | 0.83 | 0.64 | |
| ED_α-LA | 0.91 | 0.61 | 0.83 | 0.81 | |
| ED_CMP | 0.89 | 0.73 | 0.64 | 0.81 |
| WP65_CON _t0.05 | WP65_CON _t1 | WP65_CON _t18 | WP65_HYD _t0.05 | WP65_HYD _t1 | WP65_HYD _t18 | Trolox | |
| mean | 50.77 | 48.92 | 46.01 | 49.67 | 47.21 | 45.21 | 94.92 |
| sd | 7.05 | 4.40 | 3.48 | 5.26 | 5.44 | 4.64 | 2.25 |
| WP80_CON _t0.05 | WP80_CON _t1 | WP80_CON _t18 | WP80_HYD _t0.05 | WP80_HYD _t1 | WP80_HYD _t18 | Trolox | |
| mean | 49.13 | 51.12 | 54.05 | 49.68 | 51.55 | 51.39 | 96.51 |
| sd | 5.63 | 8.77 | 2.36 | 7.22 | 6.75 | 3.66 | 0.57 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakkas, L.; Lekaki, E.; Moatsou, G. Whey Protein Hydrolysates of Sheep/Goat Origin Produced by the Action of Trypsin without pH Control: Degree of Hydrolysis, Antihypertensive Potential and Antioxidant Activities. Foods 2022, 11, 2103. https://doi.org/10.3390/foods11142103
Sakkas L, Lekaki E, Moatsou G. Whey Protein Hydrolysates of Sheep/Goat Origin Produced by the Action of Trypsin without pH Control: Degree of Hydrolysis, Antihypertensive Potential and Antioxidant Activities. Foods. 2022; 11(14):2103. https://doi.org/10.3390/foods11142103
Chicago/Turabian StyleSakkas, Lambros, Eleni Lekaki, and Golfo Moatsou. 2022. "Whey Protein Hydrolysates of Sheep/Goat Origin Produced by the Action of Trypsin without pH Control: Degree of Hydrolysis, Antihypertensive Potential and Antioxidant Activities" Foods 11, no. 14: 2103. https://doi.org/10.3390/foods11142103
APA StyleSakkas, L., Lekaki, E., & Moatsou, G. (2022). Whey Protein Hydrolysates of Sheep/Goat Origin Produced by the Action of Trypsin without pH Control: Degree of Hydrolysis, Antihypertensive Potential and Antioxidant Activities. Foods, 11(14), 2103. https://doi.org/10.3390/foods11142103