
Recent Advances in Nattokinase-Enriched Fermented Soybean Foods: A Review

 , and
, and
Abstract
1. Introduction
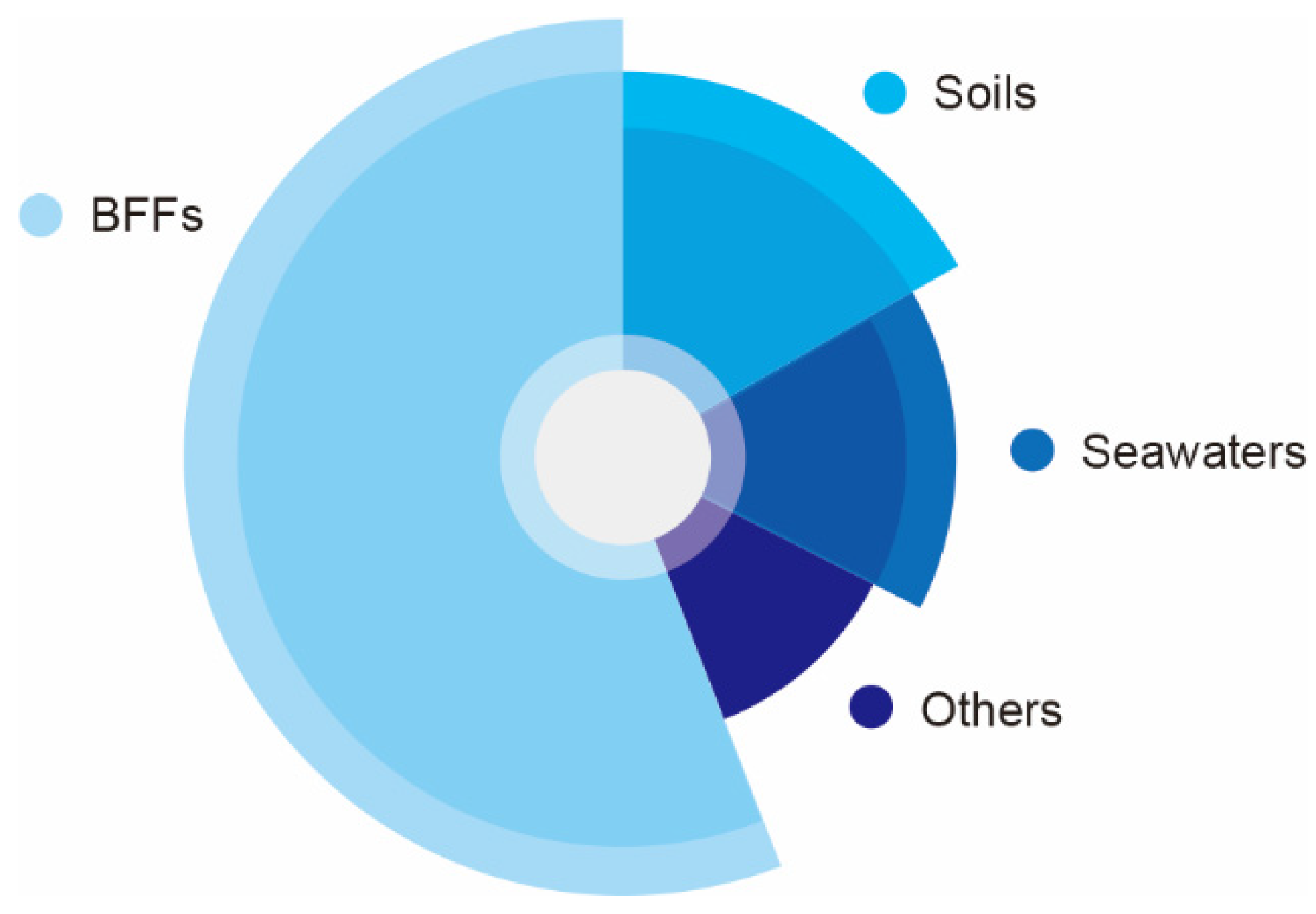
2. Fermented Soybean Foods Rich in NK
3. Production Strains of NK

{kind=link}
{kind=link}
{kind=link}
| Sources | Strains | Ref. | ||
|---|---|---|---|---|
| Fermented soybean foods | Natto | B. subtilis natto | [24] | |
| B. subtilis natto | [47] | |||
| B. subtilis REVS12 | [44] | |||
| B. subtilis natto B-12 | [48] | |||
| Da jang | B. subtilis LSSE-62 | [49] | ||
| Moromi | B. subtilis K2 | [17] | ||
| Douchi | B. subtilis DC33 | [50] | ||
| B. subtilis YF38 | [51] | |||
| B. subtilis MX-6 | [45] | |||
| B. amyloliquefaciens DC-4 | [52] | |||
| Thua nao | B. subtilis | [37] | ||
| Chungkukjang | Bacillus sp. CK 11-4 | [53] | ||
| Doenjang | B. subtilis WRL101 | [54] | ||
| Bacillus sp. DJ-4 | [55] | |||
| Gembus | B. pumilus | [56] | ||
| Soils | B. subtilis natto WTC016 | [57] | ||
| B. sublitis RJAS19 | [58] | |||
| B. subtilis IMR-NK1 | [59] | |||
| B. subtilis TKU007 | [60] | |||
| B. subtilis | [61] | |||
| Pseudomonas sp. TKU015 | [62] | |||
| Marine | Marine water | B. subtilis | [63] | |
| Marine cultures | B. subtilis ICTF-1 | [64] | ||
| Plants | Red alga Porphyra Dentata | B. subtilis N1 | [65] | |
| The root tissue of Stemona japonica (Blume) Miq | Endophytic Strain EJS-3 | [66] | ||
| Dairy products | Bovine milk | Pseudomonas aeruginosa CMSS | [67] | |
| Fermented milk | B. subtilis VITMS 2 | [68] | ||
| Rust | B. cereus VITSDVM3 | [69] | ||
| Cow dung | Bacillus spp. IND7 | [70] | ||
4. Optimization Strategies for NK
4.1. Screening and Breeding of NK Production Strains
4.2. Culture Medium and Culture Condition Optimization
| Strains | Culture Medium | Culture Methods | Results | Ref. |
|---|---|---|---|---|
| B. subtilis natto WTC016 | Luria–Bertani (LB) liquid medium: peptone (10 g/L), yeast extract (5 g/L), NaCl (10 g/L), agar (15 g/L), | Fermented at 30 °C, pH 7.0, and 60 mL of loading volume in 250 mL conical flask for 24 h | 3284 ± 58 IU/mL | [57] |
| B. subtilis VITMS2 | Sucrose (1%), soybean meal (2%), malt extract (2%), and 10 mM of CaCl2, MgSO4, Na2HPO4 and K2HPO4 | Inoculated with 4.0% inoculum, pH 7.0, 30 °C for 48 h | 171.1 ± 0.27 U/mL | [68] |
| B. subtilis NDF | Soybean milk (180 g/L) and glucose (105 g/L) | Inoculated with the 5% (V/V) of inoculant 3.7 × 108 colony forming unit/mL (CFU/mL) and fed-batch fermented at 30 °C, initial pH 7.0, 600 rpm, 1.0 vvm in 6-L fermenter with 4 L of loading volume for 25 h | 10,220 IU/mL | [77] |
| B. subtilis natto | Yeast extract (6%), soy peptone (1.2%), and glycerol (6%) | Inoculated with the 2% volume/volume (V/V) spore solution (5.2 ± 0.5 × 1010 spores/mL) and fed-batch fermented at 40 °C for 10 h in fermenter | 587 U/mL | [78] |
| B. subtilis natto | Soybean flour (16.7%) and rice husk (13.3%) with 70% water content | Solid-state fermentation incubated at 37 °C for 24 h | 2503.4 IU/g dry substrate | [80] |
| B. subtilis D21-8 | Cassava starch (20 g/L), soybean meal (10 g/L), K2HPO4 (3 g/L), KH2PO4 (1 g/L), MgSO4·7H2O (3.5 g/L), and CaCl2 (0.2 g/L) | 2% (V/V) inoculated and fermented at 34 °C and 180 rpm for 72 h in 250 mL Erlenmeyer flask with loading 50 mL medium liquid | 1754 U/mL | [81] |
| B. subtilis natto 1A752 | Ginkgo seeds | Relative humidity 80%, initial water content 73%, at 38 °C, inoculation volume 18% for 38 h | 3682 ± 43 IU/g dry substrate | [82] |
| B. subtilis natto | Chestnut | 5% (V/V) inoculum concentration at 38 °C for 56 h | 6479 IU/g dry substrate | [83] |
| B. subtilis MTCC 2616 | Tryptone (10 g/L), yeast extract (10 g/L), K2HPO4·3H2O (1 g/L), MgSO4·7H2O (0.5 g/L), and CaCl2·2H2O (0.5 g/L). | Fermented at 30 °C and 150 rpm in an orbital shaker for 51 h | 789.93 U/mL | [84] |
| B. subtilis GXA-28 | The cane molasses contained 35.2% (w/w) sucrose, 9.0% (w/w) glucose, 15.8% (w/w) fructose; the monosodium glutamate waste liquor consisted of 2.0% (w/w) glutamate, 0.3% (w/w) (NH4)2SO4 | 2% (V/V) inoculum in solid medium at pH 8.0 and 45 °C for 24 h in shallow tray | 986 U/g-substrate | [85] |
| B. subtilis natto | Glucose (6.10 g/L), soybean peptone (5.00 g/L), K2HPO4 (3.00 g/L), MgSO4.7H2O (0.25 g/L), NaCl (4.27 g/L), CaCl2 (0.05 g/L) | Inoculated 5 billion colony-forming units/mL of medium in 37 °C and pH 7.5 for 20 h | 55.82 U/mL | [87] |
| B. subtilis 13,932 | Glucose (30.868 g/L), tofu processing wastewater (93.669%), MgSO4·7H2O (1.129 g/L), CaCl2 (0.791 g/L) | Fermented at 37 °C, pH 7.0, 70 mL liquid medium, and 200 rpm in 100 L bioreactors | 7209.15 ± 195.46 IU/mL | [89] |
| B. subtilis 14,715 | 25 g of pigeon pea | 2% (V/V) inoculum (107 CFU/mL) fermented at 35 °C for 32 h | 53.03 U/g | [90] |
| B. subtilis natto | LB medium, glycerol (20 mL/L) | Fermented at 37 °C, pH 7.0, in fermenter | 7778 ± 17.28 U/mL | [91] |
| P. aeruginosa CMSS | Shrimp shell (1%), KH2PO4 (0.1%), MgSO4 (0.05%) | 1% (V/V) inoculum at 37 °C, 120 rpm, and pH 7 for 24 h in a shaker | 2581 U/ mL | [67] |
4.3. Recombinant Expression
4.4. Molecular Modification
5. Enrichment Extraction of NK
6. Maintenance of NK Activity
7. Biological Functions and Pharmaceutical Value of NK
7.1. Antithrombotic and Fibrinolytic Efficacy of NK
7.2. Anti-Atherosclerotic Effect of NK
7.3. Hyperlipidemia Reducing Effects of NK
7.4. Antihypertensive Effects of NK
7.5. Neuroprotective Effects of NK
7.6. Other Diseases
8. Toxicity Assessment of NK
9. Conclusions and Prospects
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Richard, A.; Margaritis, A. World Health Statistics 2022: Monitoring Health for the SDGs, Sustainable Development Goals; World Health Organization: Geneva, The Netherlands, Licence: CC BY-NC-SA 3.0 IGO; 2022; Available online: https://www.who.int/data/gho/publications/world-health-statistics (accessed on 19 May 2022).
- Geraghty, L.; Figtree, G.A.; Schutte, A.E.; Patel, S.; Arnott, C. Cardiovascular disease in women: From pathophysiology to novel and emerging risk factors. Heart Lung Circ. 2020, 30, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Falkner, B.; Gidding, S. Life-course implications of pediatric risk factors for cardiovascular disease. Can. J. Cardiol. 2021, 37, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Nozue, M.; Shimazu, T.; Charvat, H.; Mori, N.; Tsugane, S. Fermented soy products intake and risk of cardiovascular disease and total cancer incidence: The Japan Public Health Center–based Prospective study. Eur. J. Clin. Nutr. 2021, 75, 954–968. [Google Scholar] [CrossRef] [PubMed]
- Kokubo, Y.; Iso, H.; Ishihara, J.; Okada, K.; Inoue, M.; Tsugane, S. Association of dietary intake of soy, beans, and isoflavones. Circulation 2007, 116, 2553–2562. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Zhang, X.; Li, C.; Jiao, S.; Dong, W. Association between consumption of soy and risk of cardiovascular disease: A meta-analysis of observational studies. Eur. J. Prev. Cardiol. 2017, 24, 735. [Google Scholar] [CrossRef]
- Yong, P.; Yang, X.; Zhang, Y. Microbial fibrinolytic enzymes: An overview of source, production, properties, and thrombolytic activity in vivo. Appl. Microbiol. Biotechnol. 2005, 69, 126–132. [Google Scholar] [CrossRef]
- Dabbagh, F.; Negahdaripour, M.; Berenjian, A.; Behfar, A.; Mohammadi, F.; Zamani, M.; Irajie, C.; Ghasemi, Y. Nattokinase: Production and application. Appl. Microbiol. Biotechnol. 2014, 98, 9199–9206. [Google Scholar] [CrossRef]
- Weng, Y.; Jian, Y.; Sawyer, S.; Wang, K. Nattokinase: An oral antithrombotic agent for the prevention of cardiovascular disease. Int. J. Mol. Sci. 2017, 18, 523. [Google Scholar] [CrossRef]
- Chen, H.; Mcgowan, E.M.; Ren, N.; Lal, S.; Lin, Y. Nattokinase: A promising alternative in prevention and treatment of cardiovascular diseases. Biomark. Insights 2018, 13, 117727191878513. [Google Scholar] [CrossRef]
- Yatagai, C.; Maruyama, M.; Kawahara, T.; Sumi, H. Nattokinase-promoted tissue plasminogen activator release from human cells. Pathophysiol. Haemost. Thromb. 2008, 36, 227–232. [Google Scholar] [CrossRef]
- Chien-Hsun, H.; Ming-Ching, S.; Jen-Shiou, L.; Yao-Ke, W.; Kai-Lin, H.; Thau-Ming, C.; Nae-Cherng, Y. Nattokinase decreases plasma levels of fibrinogen, factor VII, and factor VIII in human subjects. sciencedirect. Nutr. Res. 2009, 29, 190–196. [Google Scholar] [CrossRef]
- You, W.K.; Sohn, Y.D.; Kim, K.Y.; Park, D.H.; Jang, Y.; Chung, K.H. Purification and molecular cloning of a novel serine protease from the centipede, scolopendra subspinipes mutilans. Insect Biochem. Mol. Biol. 2004, 34, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Yamanaka, N.; Ohnishi, K.; Fukayama, M.; Yoshino, M. Inhibition of angiotensin I converting enzyme by Subtilisin NAT (nattokinase) in natto, a Japanese traditional fermented food. Food Funct. 2012, 3, 674–678. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.-H.; Green-Johnson, J.M.; Buckley, N.D.; Lin, Q.-Y. Bioactivity of soy-based fermented foods: A review. Biotechnol. Adv. 2019, 37, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Keitarou, K.; Satoshi, Y. Trends in the application of Bacillus in fermented foods. Curr. Opin. Biotechnol. 2018, 56, 36–42. [Google Scholar] [CrossRef]
- Syahbanu, F.; Giriwono, P.E.; Tjandrawinata, R.R.; Suhartono, M.T. Molecular analysis of a fibrin-degrading enzyme from Bacillus subtilis K2 isolated from the indonesian soybean–based fermented food moromi. Mol. Biol. Rep. 2020, 47, 8553–8563. [Google Scholar] [CrossRef]
- Jeong, D.Y.; Ryu, M.S.; Yang, H.J.; Park, S. γ-PGA-rich chungkookjang, short-term fermented soybeans: Prevents memory impairment by modulating brain insulin sensitivity, neuro-inflammation, and the gut–microbiome–brain axis. Foods 2021, 10, 221. [Google Scholar] [CrossRef]
- Ohta, T. Natto: In Legume-Based Fermented Foods; Reddy, N.R., Pierson, M.D., Salunkhe, D.K., Eds.; CRC Press: Boca Raton, FL, USA, 1986; pp. 85–93. [Google Scholar]
- Mine, Y.; Wong, A.H.K.; Jiang, B. Fibrinolytic enzymes in Asian traditional fermented foods. Food Res. Int. 2005, 38, 243–250. [Google Scholar] [CrossRef]
- Miyake, S.; Wantanabe, K.; Yoshikawa, M.; Nonoguchi, Y. Studies on Bacillus natto protease: The constituting amino acids of the crystalline protease of Bac. Natto. Sawamura. Sci. Rep. Hyogo Univ. Agric. 1956, 28, 527–529. [Google Scholar] [CrossRef]
- Oshima, K. The properties of protease A, the proteolytic enzyme of natto bacteria. J. Soc. Agric. For. Sapporo 1925, 71, 387–403. [Google Scholar]
- Miyake, S.; Shimizu, J. The studies on Bacillus natto protease (1): On crystallization of protease. Sci. Rep. Hyogo Univ. Agric. 1953, 32, 260–262. [Google Scholar] [CrossRef]
- Sumi, H.; Hamada, H.; Tsushima, H.; Mihara, H.; Muraki, H. A novel fibrinolytic enzyme (nattokinase) in the vegetable cheese natto; a typical and popular soybean food in the Japanese diet. Experientia 1987, 43, 1110. [Google Scholar] [CrossRef] [PubMed]
- Ito, K. Review of the health benefits of habitual consumption of miso soup: Focus on the effects on sympathetic nerve activity, blood pressure, and heart rate. Environ. Health Prev. Med. 2020, 25, 45. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Tatsumi, E.; Ding, C.H.; Li, L.T. Angiotensin I–converting enzyme inhibitory peptides in douchi, a chinese traditional fermented soybean product. Food Chem. 2006, 98, 551–557. [Google Scholar] [CrossRef]
- Guan, R.F.; Liu, Z.F.; Zhang, J.J.; Wei, Y.X.; Said, W.; Liu, D.H.; Ye, X.Q. Investigation of biogenic amines in sufu (furu): A Chinese traditional fermented soybean food product. Food Control 2013, 31, 345–352. [Google Scholar] [CrossRef]
- Kwon, D.Y.; Chung, K.R.; Yang, H.J.; Jang, D.J. Gochujang (Korean red pepper paste): A Korean ethnic sauce, its role and history. J. Ethn. Foods 2015, 2, 29–35. [Google Scholar] [CrossRef]
- Shin, D.; Jeong, D. Korean traditional fermented soybean products: Jang. J. Ethn. Foods 2015, 2, 2–7. [Google Scholar] [CrossRef]
- Kim, D.C.; Quang, T.H.; Yoon, C.S.; Ngan, N.; Lim, S.I.; Lee, S.Y.; Kim, Y.C.; Oh, H. Anti-neuroinflammatory activities of indole alkaloids from kanjang (Korean fermented soy source) in lipopolysaccharide-induced BV2 microglial cells. Food Chem. 2016, 213, 69–75. [Google Scholar] [CrossRef]
- Tamang, J.P. Native microorganisms in the fermentation of kinema. Indian J. Microbiol. 2003, 43, 127–130. [Google Scholar]
- Chettri, R.; Tamang, J.P. Bacillus species isolated from tungrymbai and bekang, naturally fermented soybean foods of india. Int. J. Food Microbiol. 2015, 197, 72–76. [Google Scholar] [CrossRef]
- Gupta, A.; Tiwari, S.K. Probiotic potential of Lactobacillus plantarum ID1 isolated from batter of dosa, a south indian fermented food. Probiotics Antimicrob. Proteins 2014, 6, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Syahbanu, F.; Giriwono, P.E.; Tjandrawinata, R.R.; Suhartono, M.T. Molecular docking of Subtilisin K2, a fibrin-degrading enzyme from indonesian moromi, with its substrates. Ciência E Tecnol. Aliment. 2021, 42, e61820. [Google Scholar] [CrossRef]
- Hachmeister, K.A.; Fung, D.Y.C. Tempeh: A mold-modified indigenous fermented food made from soybeans and/or cereal grains. Crit. Rev. Microbiol. 1993, 19, 137–188. [Google Scholar] [CrossRef] [PubMed]
- Press, C. Fermented Foods and Beverages of the World; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar] [CrossRef]
- Inatsu, Y.; Nakamura, N.; Yuriko, Y.; Fushimi, T.; Kawamoto, S. Characterization of Bacillus subtilis strains in thua nao, a traditional fermented soybean food in northern Thailand. Lett. Appl. Microbiol. 2010, 43, 237–242. [Google Scholar] [CrossRef]
- Antai, S.P.; Ibrahim, M.H. Micro-organisms associated with African locust bean (parkia filicoidea welw) fermentation for ‘dawadawa’ production. J. Appl. Bacteriol. 2010, 61, 145–148. [Google Scholar] [CrossRef]
- Selvarajan, E.; Bhatnagar, N. Nattokinase: An updated critical review on challenges and perspectives. Cardiovasc. Hematol. Agents Med. Chem. 2017, 16, CD012175. [Google Scholar] [CrossRef]
- Tamang, J.P.; Koichi, W.; Holzapfel, W.H. Review: Diversity of microorganisms in global fermented foods and beverages. Front. Microbiol. 2016, 7, 377. [Google Scholar] [CrossRef]
- Owusu-Kwarteng, J.; Parkouda, C.; Adewumi, G.A.; Ouoba, L.I.I.; Jespersen, L. Technologically relevant Bacillus species and microbial safety of west african traditional alkaline fermented seed condiments. Crit. Rev. Food Sci. Nutr. 2020, 62, 871–888. [Google Scholar] [CrossRef]
- Zhang, Q.; Wu, Y.; Gong, M.; Zhang, H.; Liu, L. Production of proteins and commodity chemicals using engineered Bacillus subtilis platform strain. Essays Biochem. 2021, 65, 173–185. [Google Scholar] [CrossRef]
- Xiang, M.; Kang, Q.; Zhang, D. Advances on systems metabolic engineering of Bacillus subtilis as a chassis cell. Synth. Syst. Biotechnol. 2020, 5, 245–251. [Google Scholar] [CrossRef]
- Vignesh, H.; Eajas, B.M.; Ramesh, B.; Saravanan, N. Production, optimization and characterization of nattokinase from Bacillus subtilis REVS 12 isolated from natto. Int. J. Sci. Eng. Res. 2014, 5, 423. [Google Scholar]
- Man, L.L.; Xiang, D.J.; Zhang, C.L. Strain screening from traditional fermented soybean foods and induction of nattokinase production in Bacillus subtilis MX–6. Probiotics Antimicro. Proteins 2019, 11, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Wang, H.; Chen, Z.; Lv, Z.; Xie, Y.; Lu, F. Profiling of dynamic changes in the microbial community during the soy sauce fermentation process. Appl. Microbiol. Biotechnol. 2013, 97, 9111–9119. [Google Scholar] [CrossRef] [PubMed]
- Urano, T.; Hayato, I.; Kazuo, U.; Yasuhiro, S.; Masaki, O.; Sumio, A.; Yoshinori, T.; Isao, S.; Akikazu, T. The profibrinolytic enzyme Subtilisin NAT purified from Bacillus subtilis cleaves and inactivates plasminogen activator inhibitor type 1. J. Biol. Chem. 2021, 276, 24690–24696. [Google Scholar] [CrossRef]
- Wang, C.; Du, M.; Zheng, D.; Kong, F.; Zu, G.; Feng, Y. Purification and characterization of nattokinase from Bacillus subtilis natto B-12. J. Agric. Food Chem. 2009, 57, 9722–9729. [Google Scholar] [CrossRef]
- Wei, X.T.; Luo, M.F.; Xu, L.; Zhang, Y.W.; Lin, X.; Kong, P.; Liu, H.Z. Production of fibrinolytic enzyme from Bacillus amyloliquefaciens by fermentation of chickpeas, with the evaluation of the anticoagulant and antioxidant properties of chickpeas. J. Agric. Food Chem. 2011, 59, 3957–3963. [Google Scholar] [CrossRef]
- Cheng, T.; Bao, P.; Bo, L.; Nout, R.; Ping, L.; Hong, J.; Chen, L. Purification and characterization of a fibrinolytic enzyme of Bacillus subtilis DC33, isolated from Chinese traditional douchi. J. Ind. Microbiol. Biotechnol. 2006, 33, 750–758. [Google Scholar] [CrossRef]
- Liang, X.; Jia, S.; Sun, Y.; Chen, M.; Chen, X.; Zhong, J.; Huan, L. Secretory Expression of Nattokinase from Bacillus subtilis YF38 in Escherichia coli. Mol. Biotechnol. 2007, 37, 187–194. [Google Scholar] [CrossRef]
- Zhang, Z. Purification and characterization of a fibrinolytic enzyme produced by Bacillus amyloliquefaciens DC-4 screened from douchi, a traditional chinese soybean food. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2003, 134, 45–52. [Google Scholar] [CrossRef]
- Kim, W. Purification and characterization of a fibrinolytic enzyme produced from Bacillus sp. strain CK 11-4 screened from chungkook–jang. Appl. Environ. Microbiol. 1996, 62, 2482. [Google Scholar] [CrossRef]
- Chang, S.P.; Dong, H.K.; Lee, W.Y.; Kang, D.O.; Song, J.J.; Choi, N.S. Identification of fibrinogen-induced nattokinase WRL101 from Bacillus subtilis WRL101 isolated from doenjang. Afr. J. Microbiol. Res. 2013, 7, 1983–1992. [Google Scholar] [CrossRef]
- Kim, S.H.; Choi, N.S. Purification and characterization of subtilisin DJ-4 secreted by Bacillus sp. strain DJ-4 screened from doen-jang. J. Agric. Chem. Soc. Jpn. 2020, 64, 1722–1725. [Google Scholar] [CrossRef]
- Afifah, D.N.; Sulchan, M.; Syah, D.; Yanti Suhartono, M.T. Purification and characterization of a fibrinolytic enzyme from Bacillus pumilus 2.g isolated from gembus, an indonesian fermented food. Prev. Nutr. Food Sci. 2014, 19, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Ju, S.; Cao, Z.; Wong, C.; Liu, Y.; Li, J. Isolation and optimal fermentation condition of the Bacillus subtilis subsp. natto strain WTC016 for nattokinase production. Fermentation 2019, 5, 92. [Google Scholar] [CrossRef]
- Kumar, D.; Rakshitha, R.; Vidhya, M.A.; Jennifer, P.S.; Kalaichelv, P.T. Production, optimization and characterization of fibrinolytic enzyme by Bacillus subtilis RJAS19. Pak. J. Biol. Sci. PJBS 2014, 17, 529–534. [Google Scholar] [CrossRef]
- Chang, C.T.; Fan, M.H.; Kuo, F.C.; Sung, H.Y. Potent fibrinolytic enzyme from a mutant of Bacillus subtilisimr NK1. J. Agric. Food Chem. 2020, 48, 3210–3216. [Google Scholar] [CrossRef]
- Wang, S.L.; Wu, Y.Y.; Liang, T.W. Purification and biochemical characterization of a nattokinase by conversion of shrimp shell with Bacillus subtilis TKU007. New Biotechnol. 2011, 28, 196–202. [Google Scholar] [CrossRef]
- Uppuluri, S. Screening, production, purification, characterization and evaluation of nattokinase from Bacillus subtilis isolated from blood soil Andhra Pradesh. Am. Res. Thoughts 2016, 2, 3691–3698. [Google Scholar]
- Wang, S.L.; Chen, H.J.; Liang, T.W.; Lin, Y.D. A novel nattokinase produced by pseudomonas sp. TKU015 using shrimp shells as substrate. Process Biochem. 2009, 44, 70–76. [Google Scholar] [CrossRef]
- Borah, D.; Shahin, L.; Sangra, A. Production, purification and characterization of nattokinase from Bacillus subtilis, isolated from sewage water. Int. J. Pharm. Arch. 2012, 1, 21–24. [Google Scholar]
- Mahajan, P.M.; Nayak, S.; Lele, S.S. Fibrinolytic enzyme from newly isolated marine bacterium Bacillus subtilis ICTF-1: Media optimization, purification and characterization. J. Biosci. Bioeng. 2012, 113, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.T.V.; Wu, G.J.; Hsieh, M.C.; Chang, S.H.; Tsai, G.J. Purification and characterization of nattokinase from cultural filtrate of red alga porphyra dentata fermented by Bacillus subtilis n1. J. Mar. Sci. Technol. 2015, 23, 240–248. [Google Scholar] [CrossRef]
- Lu, F.; Sun, L.; Lu, Z.; Bie, X.; Fang, Y.; Shu, L. Isolation and identification of an endophytic strain EJS-3 producing novel fibrinolytic enzymes. Curr. Microbiol. 2007, 54, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, S.D.; Vaithilingam, M.; Shanker, R.; Kumar, S.; Thiyur, S.; Babu, V.; Selvakumar, J.N.; Prakash, S. Exploring the in vitro thrombolytic activity of nattokinase from a new strain Pseudomonas aeruginosa CMSS. Jundishapur J. Microbiol. 2015, 8, e23567. [Google Scholar] [CrossRef] [PubMed]
- Keziah, S.M.; Devi, C.S. Fibrinolytic and ACE Inhibitory Activity of Nattokinase Extracted from Bacillus subtilis VITMS 2: A Strain Isolated from Fermented Milk of Vigna unguiculata. Protein J. 2021, 40, 876–890. [Google Scholar] [CrossRef]
- Vaithilingam, M.; Chandrasekaran, S.D.; Gupta, S.; Paul, D.; Sahu, P.; Selvaraj, J.N.; Babu, V. Extraction of Nattokinase Enzyme from Bacillus cereus Isolated from Rust. Natl. Acad. Sci. Lett. 2016, 39, 263–267. [Google Scholar] [CrossRef]
- Ponnuswamy, V.; Arumugaperumal, A.; Prakash, V.; Valan, A.M.; Abdullah, A. Cow dung is a novel feedstock for fibrinolytic enzyme production from newly isolated Bacillus sp. IND7 and its application in in vitro clot lysis. Front. Microbiol. 2016, 7, 361. [Google Scholar] [CrossRef]
- Huang, Y.; Ding, S.; Liu, M.; Gao, C.; Yang, J.; Zhang, X.; Ding, B. Ultra–small and anionic starch nanospheres: Formation and vitro thrombolytic behavior study. Carbohydr. Polym. 2013, 96, 426–434. [Google Scholar] [CrossRef]
- Cai, D.; Zhu, C.; Chen, S. Microbial production of nattokinase: Current progress, challenge and prospect. World J. Microbiol. Biotechnol. 2017, 33, 84. [Google Scholar] [CrossRef]
- Li, S.Y. Screening of strains with the high activity and thermostability nattokinase by 60co γ–ray irradiation. J. Nucl. Agric. Sci. 2013, 6, 782–785. [Google Scholar]
- Liu, S.; Jiang, M.J.; Chen, X.H.; Chen, G.; Dong, M.S. Effect of the 36th amino acid site mutagenesis on the activity and thermostability of nattokinase. J. Nanjing Agric. Univ. 2008, 31, 130–136. [Google Scholar] [CrossRef]
- Guan, Z.; Li, Z.; Sun, H. Research on the breeding of Bacillus subtilis natto highly producing nattokinase by the compound mutation of hydroxylamine hydrochloride and UV radiation. Food Ferment. Ind. 2009, 35, 71–74. [Google Scholar] [CrossRef]
- Wei, X.; Luo, M.; Xie, Y.; Yang, L.; Li, H.; Xu, L.; Liu, H. Strain Screening, Fermentation, Separation, and Encapsulation for Production of Nattokinase Functional Food. Appl. Biochem. Biotechnol. 2012, 168, 1753–1764. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Shen, J.; Li, Y.; Wang, Z.; Zhao, Y.; Chen, Y.; Zhao, J. High and Economical Nattokinase Production with Acetoin as a Useful Byproduct from Soybean Milk and Glucose. Probiotics Antimicro. Proteins 2021. [Google Scholar] [CrossRef]
- Aydin, B.; Raja, M.; John, K.; Fariba, D.; Younes, G. Nattokinase production: Medium components and feeding strategy studies. Chem. Ind. Chem. Eng. Q. 2014, 20, 541–547. [Google Scholar] [CrossRef]
- Meng, F.Y.; Xue, F.; Shi, H. Effects of natto kinase on blood lipid and blood rheology in atherosclerosis model rats. Chin. J. Lab. Diagn. 2013, 17, 1567–1569. [Google Scholar]
- Nie, G.; Zhu, Z.; Fang, L.; Nie, Z.; Ye, Y.; Yue, W. Co-production of nattokinase and poly (γ-glutamic acid) under solid-state fermentation using soybean and rice husk. Braz. Arch. Biol. Technol. 2015, 58, 718–724. [Google Scholar] [CrossRef]
- Pan, S.; Chen, G.; Zeng, J.; Cao, X.; Zheng, X.; Zeng, W.; Liang, Z. Fibrinolytic enzyme production from low-cost substrates by marine Bacillus subtilis: Process optimization and kinetic modeling. Biochem. Eng. J. 2018, 141, 268–277. [Google Scholar] [CrossRef]
- Guo, N.; Jiang, Y.W.; Song, X.R.; Li, Y.Y.; Liu, Z.M.; Fu, Y.J. Effect of Bacillus natto solid-state fermentation on the functional constituents and properties of ginkgo seeds. J. Food Biochem. 2019, 43, e12820. [Google Scholar] [CrossRef]
- Dong, M.Z.; An, J.Y.; Wang, L.T.; Fan, X.H.; Fu, Y.J. Development of fermented chestnut with Bacillus natto: Functional and sensory properties. Food Res. Int. 2020, 130, 108941. [Google Scholar] [CrossRef]
- Sahoo, A.; Mahanty, B.; Daverey, A.; Dutta, K. Nattokinase production from Bacillus subtilis using cheese whey: Effect of nitrogen supplementation and dynamic modelling. J. Water Process Eng. 2020, 38, 101533. [Google Scholar] [CrossRef]
- Zeng, W.; Li, W.; Shu, L.; Yi, J.; Chen, G.; Liang, Z. Non-sterilized fermentative co–production of poly (γ–glutamic acid) and fibrinolytic enzyme by a thermophilic Bacillus subtilis GXA-28. Bioresour. Technol. 2013, 142, 697–700. [Google Scholar] [CrossRef]
- Zhou, H.M.; Zhang, H.X.; Xie, Y.H.; Zhou, T.T.; Liu, H.; Luo, Y.B. Optimization of liquid fermentation conditions and encapsulation for nattokinase production. Adv. Mater. Res. 2013, 781–784, 1403–1409. [Google Scholar] [CrossRef]
- Nguyen, A.T.; Thuan, D.T.H.; Tran, T.M.T.; Nguyen, T.H. Determination the optimum fermentation in obtaining nattokinase by Bacillus subtilis natto. ISSR J. 2015, 13, 663–668. [Google Scholar]
- Zhang, J.; Cui, Q.; Qian, B.; Sun, X. High Cell Density Cultivation of A Recombinant Bacillus Subtilis for Nattokinase Production. Authorea 2020. [Google Scholar] [CrossRef]
- Li, T.; Zhan, C.; Guo, G.; Liu, Z.; Ouyang, P. Tofu processing wastewater as a low-cost substrate for high activity nattokinase production using Bacillus subtilis. BMC Biotechnol. 2021, 21, 57. [Google Scholar] [CrossRef]
- Lee, B.H.; Lai, Y.S.; Wu, S.C. Antioxidation, angiotensin converting enzyme inhibition activity, nattokinase, and antihypertension of Bacillus subtilis (natto)-fermented pigeon pea. J. Food Drug Anal. 2015, 23, 750–757. [Google Scholar] [CrossRef]
- Zhang, X.; Lyu, X.; Tong, Y.; Wang, J.; Ye, J.; Yang, R. Chitosan/casein based microparticles with a bilayer shell–core structure for oral delivery of nattokinase. Food Funct. 2020, 11, 10799–10816. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Xiong, S.; Zhang, J.; Yang, C.Y. Expression and purification of recombinant nattokinase in Spodoptera frugiperda cells. Biotechnol. Lett. 2007, 29, 1459–1464. [Google Scholar] [CrossRef]
- Wang, K.; Tull, L. Expression of blood clot-dissolving proteins in transgenic plant (LB852). FASEB J. 2007, 28, LB852. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Quyen, T.D.; Le, H.T. Cloning and enhancing production of a detergent- and organic-solvent-resistant nattokinase from Bacillus subtilis VTCC-DVN-12-01 by using an eight-protease-gene-deficient Bacillus subtilis WB800. Microb. Cell Fact. 2013, 12, 79. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, Z.M.; Zhao, R.L.; Jin, T.T.; Zhang, X.M.; Chen, X.D. Deleting multiple lytic genes enhances biomass yield and production of recombinant proteins by Bacillus subtilis. Microb. Cell Fact. 2014, 13, 129. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhou, Y.; Chen, J.; Cai, D.; Wang, D.; Qi, G.; Chen, S. Efficient expression of nattokinase in Bacillus licheniformis: Host strain construction and signal peptide optimization. J. Ind. Microbiol. Biotechnol. 2015, 42, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Jing, J.F.; Chen, B.; Li, Y.; He, Z.; Huo, J. Cloning of nattokinase gene and expression in pichia pastoris. J. Biol. 2011, 28(05), 55–57. [Google Scholar] [CrossRef]
- Cai, D.; Zhang, B.; Rao, Y.; Li, L.; Zhu, J.; Li, J.; Ma, X.; Chen, S. Improving the utilization rate of soybean meal for efficient production of bacitracin and heterologous proteins in the aprA-deficient strain of Bacillus licheniformis. Appl. Microbiol. Biotechnol. 2019, 103, 4789–4799. [Google Scholar] [CrossRef]
- Ni, H.; Guo, P.C.; Jiang, W.L.; Fan, X.M.; Luo, X.Y.; Li, H.H. Expression of nattokinase in Escherichia coli and renaturation of its inclusion body. J. Biotechnol. 2016, 231, 65–71. [Google Scholar] [CrossRef]
- Li, C.; Du, Z.; Qi, S.; Zhang, X.; Tian, H. Food-grade expression of nattokinase in Lactobacillus delbrueckii subsp. bulgaricus and its thrombolytic activity in vitro. Biotechnol. Lett. 2020, 42, 2179–2187. [Google Scholar] [CrossRef]
- Yan, G.; Shu, M.; Shen, W.; Ma, L.; Zhai, C.; Wang, Y.; H, Z. Heterologous expression of nattokinase from B. subtilis natto using Pichia pastoris GS115 and assessment of its thrombolytic activity. BMC Biotechnol. 2021, 21, 49. [Google Scholar] [CrossRef]
- Han, L.; Zhang, L.; Liu, J.; Li, H.; Wang, Y.; Hasi, A. Transient expression of optimized and synthesized nattokinase gene in melon (Cucumis melo L.) fruit by agroinfiltration. Plant Biotechnol. 2015, 32, 175–180. [Google Scholar] [CrossRef]
- Han, L.; Ju, L.F.; Niu, Y.D.; Wang, Y.C.; Agula, H. Optimization, synthesis and transient expression of nattokinase gene in tobacco(Nicotiana tabacum L.) leaves. China Biotechnol. 2015, 2, e25271. [Google Scholar] [CrossRef]
- Kyoko, H.T.; Hiroshi, E. Molecular breeding to create optimized crops: From genetic manipulation to potential applications in plant factories. Front. Plant Sci. 2016, 7, 539. [Google Scholar] [CrossRef]
- Shahid, N.; Daniell, H. Plant-based oral vaccines against zoonotic and non-zoonotic diseases. Plant Biotechnol. J. 2016, 14, 2079–2099. [Google Scholar] [CrossRef] [PubMed]
- Buyel, J.F.; Twyman, R.M.; Fischer, R. Extraction and downstream processing of plant-derived recombinant proteins. Biotechnol. Adv. 2015, 33, 902–913. [Google Scholar] [CrossRef]
- Yao, J.; Weng, Y.; Alexia, D.; Wang, K. Plants as factories for human pharmaceuticals: Applications and challenges. Int. J. Mol. Sci. 2015, 16, 26122. [Google Scholar] [CrossRef]
- Guan, C.; Cui, W.; Cheng, J.; Liu, Z.; Zhou, Z. Development of an efficient autoinducible expression system by promoter engineering in Bacillus subtilis. Microb. Cell Fact. 2016, 15, 66. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, L.; Jin, Z.; Huan, L. Secretory expression of a heterologous nattokinase in Lactococcus lactis. Appl. Microbiol. Biotechnol. 2007, 75, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Zhongmei, L.; Wenhui, Z.; Chunlei, G.; Wenjing, C.; Li, Z.; Zhemin, Z. High-level extracellular production of recombinant nattokinase in Bacillus subtilis WB800 by multiple tandem promoters. BMC Microbiol. 2019, 19, 89. [Google Scholar] [CrossRef]
- Wu, S.M.; Feng, C.; Zhong, J.; Huan, L.D. Roles of S3 site residues of nattokinase on its activity and substrate specificity. J. Biochem. 2007, 142, 357–364. [Google Scholar] [CrossRef]
- Weng, M.Z.; Zheng, Z.L.; Wei, B.; Cai, Y.J.; Yin, Y.; Zou, G.L. Enhancement of oxidative stability of the subtilisin nattokinase by site-directed mutagenesis expressed in Escherichia coli. BBA Proteins Proteom. 2009, 1794, 1566–1572. [Google Scholar] [CrossRef]
- Weng, M.Z.; Deng, X.W.; Bao, W.; Zhu, L.; Wu, J.; Cai, Y.; Jia, Y.; Zheng, Z.; Zou, G. Improving the activity of the subtilisin nattokinase by site–directed mutagenesis and molecular dynamics simulation. Biochem. Biophys. Res. Commun. 2015, 465, 580–586. [Google Scholar] [CrossRef]
- Liu, Z.; Zhao, H.; Han, L.; Cui, W.; Zhou, L.; Zhou, Z. Improvement of the acid resistance, catalytic efficiency, and thermostability of nattokinase by multisite-directed mutagenesis. Biotechnol. Bioeng. 2019, 116, 1833–1843. [Google Scholar] [CrossRef] [PubMed]
- Vianney, Y.M.; Tjoa, S.E.E.; Aditama, R.; Putra, S.E.D. Designing a less immunogenic nattokinase from Bacillus subtilis subsp. natto: A computational mutagenesis. J. Mol. Model. 2019, 25, 337. [Google Scholar] [CrossRef] [PubMed]
- Purwaeni, E.; Riani, C.; Retnoningrum, D.S. Molecular Characterization of Bacterial Fibrinolytic Proteins from Indonesian Traditional Fermented Foods. Protein J. 2020, 39, 258–267. [Google Scholar] [CrossRef]
- Rasagnya, P.S.; Vangalapati, A.M. Studies on Optimization of Process Parameters for Nattokinase Production by Bacillus subtilis NCIM 2724 and Purification by Liquid-Liquid Extraction. Int. J. Innov. Res. Sci. Eng. Technol. 2013, 2, 4516–4521. [Google Scholar]
- Garg, R.; Thorat, B.N. Nattokinase purification by three phase partitioning and impact of t-butanol on freeze drying. Sep. Purif. Technol. 2014, 131, 19–26. [Google Scholar] [CrossRef]
- Liu, J.G.; Xing, J.M.; Chang, T.S.; Liu, H.Z. Purification of nattokinase by reverse micelles extraction from fermentation broth: Effect of temperature and phase volume ratio. Bioprocess Biosyst. Eng. 2006, 28, 267. [Google Scholar] [CrossRef]
- Yang, C.; Xing, J.; Guan, Y.; Liu, H. Superparamagnetic poly (methyl methacrylate) beads for nattokinase purification from fermentation broth. Appl. Microbiol. Biotechnol. 2006, 72, 616–622. [Google Scholar] [CrossRef]
- Ero, M.P.; Ng, C.M.; Mihailovski, T.; Harvey, N.R.; Lewis, B.H. A pilot study on the serum pharmacokinetics of nattokinase in humans following a single, oral, daily dose. Altern. Ther. Health Med. 2013, 19, 16–19. [Google Scholar]
- Arsalan, A.; Younus, H. Enzymes and nanoparticles: Modulation of enzymatic activity via nanoparticles. Int. J. Biol. Macromol. 2018, 118 Pt B, 1833–1847. [Google Scholar] [CrossRef]
- Deepak, V.; Pandian, S.; Kalishwaralal, K.; Gurunathan, S. Purification, immobilization, and characterization of nattokinase on PHB nanoparticles. Bioresour. Technol. 2009, 100, 6644–6646. [Google Scholar] [CrossRef]
- Chen, C.; Duan, H.; Gao, C.; Liu, M.; Wu, X.; Wei, Y.; Zhang, X.; Liu, Z. Non-covalent modification of thrombolytic agent nattokinase: Simultaneous improvement of fibrinolysis activity and enzymatic stability. RSC Adv. 2014, 4, 27422–27429. [Google Scholar] [CrossRef]
- Dong, X.Y.; Kong, F.P.; Yuan, G.Y.; Wei, F.; Jiang, M.L.; Li, G.M.; Wang, Z.; Zhao, Y.D.; Chen, H. Optimisation of preparation conditions and properties of phytosterol liposome-encapsulating nattokinase. Nat. Prod. Res. 2012, 26, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Darwesh, O.M.; Ali, S.S.; Matter, I.A.; Elsamahy, T.; Mahmoud, Y.A. Enzymes immobilization onto magnetic nanoparticles to improve industrial and environmental applications. Methods Enzymol. 2019, 630, 481–502. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Li, N.; Zhong, X.; Jiang, Y. Metal–organic frameworks: A potential platform for enzyme immobilization and related applications. Front. Bioeng. Biotechnol. 2020, 8, 695. [Google Scholar] [CrossRef]
- Wen, N.; Lü, S.; Xu, X.; Ning, P.; Wang, Z.; Zhang, Z.; Gao, C.; Liu, Y.; Liu, M. A polysaccharide-based micelle-hydrogel synergistic therapy system for diabetes and vascular diabetes complications treatment. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 100, 94–103. [Google Scholar] [CrossRef]
- Huang, M.; Zhang, S.; Lü, S.; Qi, T.; Yan, J. Synthesis of mesoporous silica/polyglutamic acid peptide dendrimer with dual targeting and its application in dissolving thrombus. J. Biomed. Mater. Res. Part A 2019, 107, 1824–1831. [Google Scholar] [CrossRef]
- Ren, L.; Wang, X.; Wu, H.; Shang, B.; Wang, J. Conjugation of nattokinase and lumbrukinase with magnetic nanoparticles for the assay of their thrombolytic activities. J. Mol. Catal. B Enzym. 2010, 62, 190–196. [Google Scholar] [CrossRef]
- Law, D.; Zhang, Z. Stabilization and target delivery of nattokinase using compression coating. Drug Dev. Ind. Pharm. 2007, 33, 495. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, S.; Wang, X.; Jie, L.; Yin, Z. Preparation and evaluation of nattokinase-loaded self-double-emulsifying drug delivery system. Asian J. Pharm. Sci. 2015, 10, 386–395. [Google Scholar] [CrossRef]
- Prakash, B.; Amita, V.; Fahad, A.A.; Firoz, A.; Vikas, K.; Bibhu, P. Development of surface-engineered plga nanoparticulate-delivery system of Tet-1-conjugated nattokinase enzyme for inhibition of Aβ40 plaques in Alzheimer’s disease. Int. J. Nanomed. 2017, 12, 8749–8768. [Google Scholar] [CrossRef]
- Fadl, N.; Ahmed, H.; Booles, H.; Sayed, A. Serrapeptase and nattokinase intervention for relieving Alzheimer’s disease pathophysiology in rat model. Hum. Exp. Toxicol. 2013, 32, 721–735. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.-Y.; Liu, L.-H.; Chen, C.-Y.; Lin, I.-F.; Ali, D.; Lee, A.Y.-L.; Wang, H.-M.D. Bifunctional mechanisms of autophagy and apoptosis regulations in melanoma from Bacillus subtilis natto fermentation extract. Food Chem. Toxicol. 2021, 150, 112020. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, A.; Belur, P.D.; Subramanya, S.B. Methods available to assess therapeutic potential of fibrinolytic enzymes of microbial origin: A review. J. Anal. Sci. Technol. 2018, 9, 10. [Google Scholar] [CrossRef]
- Guo, H.; Ban, Y.H.; Cha, Y.; An, E.S.; Choi, J.; Seo, D.W.; Park, D.; Choi, E.K.; Kim, Y.B. Comparative anti–thrombotic activity and haemorrhagic adverse effect of nattokinase and tissue–type plasminogen activator. Food Sci. Biotechnol. 2019, 28, 1535–1542. [Google Scholar] [CrossRef] [PubMed]
- Kurosawa, Y.; Nirengi, S.; Homma, T.; Esaki, K.; Ohta, M.; Clark, J.F.; Hamaoka, T. A single-dose of oral nattokinase potentiates thrombolysis and anti-coagulation profiles. Sci. Rep. 2015, 5, 11601. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.H.; Chen, K.T.; Lee, T.H.; Wang, C.H.; Kuo, Y.W.; Chiu, Y.H.; Hsieh, C.L.; Wu, C.J.; Chang, Y.L. Effects of natto extract on endothelial injury in a rat model. Acta Med. Okayama 2010, 64, 399–406. [Google Scholar] [CrossRef]
- Ren, N.N.; Chen, H.J.; Li, Y.; Mcgowan, E.; Lin, Y.G. A clinical study on the effect of nattokinase on carotid artery atherosclerosis and hyperlipidaemia. Zhonghua Yi Xue Za Zhi 2017, 97, 2038–2042. [Google Scholar] [CrossRef]
- Wu, D.J.; Lin, C.S.; Lee, M.Y. Lipid-lowering effect of nattokinase in patients with primary hypercholesterolemia. Acta Cardiol. Sin. 2009, 25, 26–30. [Google Scholar]
- Yang, N.C.; Chou, C.W.; Chen, C.Y.; Hwang, K.L.; Yang, Y.C. Combined nattokinase with red yeast rice but not nattokinase alone has potent effects on blood lipids in human subjects with hyperlipidemia. Asia Pac. J. Clin. Nutr. 2009, 18, 310–317. [Google Scholar] [CrossRef]
- Kang, S.J.; Lim, Y.; Kim, A.J. Korean red ginseng combined with nattokinase ameliorates dyslipidemia and the area of aortic plaques in high cholesterol-diet fed rabbits. Food Sci. Biotechnol. 2014, 23, 283–287. [Google Scholar] [CrossRef]
- Kim, J.; Gum, S.; Paik, J.; Lim, H.; Kim, K.C.; Ogasawara, K.; Inoue, K.; Park, S.; Jang, Y.; Lee, J. Effects of Nattokinase on Blood Pressure: A Randomized, Controlled Trial. Hypertens. Res. 2008, 31, 1583–1588. [Google Scholar] [CrossRef] [PubMed]
- Gitte, J.; Miki, L.; Ero, M.P.; Kathleen, B. Consumption of nattokinase is associated with reduced blood pressure and von willebrand factor, a cardiovascular risk marker: Results from a randomized, double–blind, placebo–controlled, multicenter north american clinical trial. Integr. Blood Press. Control 2016, 9, 95–104. [Google Scholar] [CrossRef]
- Ji, H.; Yu, L.; Liu, K.; Yu, Z.; Zhang, Q.; Zou, F.; Liu, B. Mechanisms of nattokinase in protection of cerebral ischemia. Eur. J. Pharmacol. 2014, 745C, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.J.; Kim, M.H.; Kim, J.; An, G.; Kim, D.; Kwon, Y.R.; Lee, A.M.; Kim, Y.B.; Kim, H.H. Abstract W P 262: Neuroprotective effect of nattokinase mediated by inhibition of platelet aggregation and thrombosis in photothrombotic stroke. Stroke 2015, 46, AWP262. [Google Scholar] [CrossRef]
- Bhatt, P.C.; Pathak, S.; Kumar, V.; Panda, B. Attenuation of neurobehavioral and neurochemical abnormalities in animal model of cognitive deficits of Alzheimer’s disease by fermented soybean nanonutraceutical. Inflammopharmacology 2018, 26, 105–118. [Google Scholar] [CrossRef]
- Huang, Z.; Ng, T.K.; Chen, W.; Sun, X.; Chen, S. Nattokinase attenuates retinal neovascularization via modulation of Nrf2/HO-1 and glial activation. Investig. Ophthalmol. Vis. Sci. 2021, 62, 25. [Google Scholar] [CrossRef]
- Takano, A.; Hirata, A.; Ogasawara, K.; Sagara, N.; Inomata, Y.; Kawaji, T.; Tanihara, H. Posterior vitreous detachment induced by nattokinase (Subtilisin NAT): A novel enzyme for pharmacologic vitreolysis. Investig. Opthalmology Vis. Sci. 2006, 47, 2075–2079. [Google Scholar] [CrossRef]
- Takabayashi, T.; Imoto, Y.; Sakashita, M.; Kato, Y.; Tokunaga, T.; Yoshida, K.; Narita, N.; Ishizuka, T.; Fujieda, S. Nattokinase, profibrinolytic enzyme, effectively shrinks the nasal polyp tissue and decreases viscosity of mucus. Allergol. Int. 2017, 66, 594–602. [Google Scholar] [CrossRef]
- Yan, Y.M.; Wang, Y.J.; Qian, J.L.; Wu, S.; Ji, Y.; Liu, Y.; Zeng, J.; Gong, A. Nattokinase crude extract inhibits Hepatocellular Carcinoma growth in mice. J. Microbiol. Biotechnol. 2003, 29, 1281–1287. [Google Scholar] [CrossRef]
- Lampe, B.J.; English, J.C. Toxicological assessment of nattokinase derived from bacillus subtilis var. natto. Food Chem. Toxicol. 2016, 88, 87–99. [Google Scholar] [CrossRef]
- Hsieh, C.W.; Lu, W.C.; Hsieh, W.C.; Huang, Y.P.; Lai, C.H.; Ko, W.C. Improvement of the stability of nattokinase using γ-polyglutamic acid as a coating material for microencapsulation. LWT-Food Sci. Technol. 2009, 42, 144–149. [Google Scholar] [CrossRef]



| Geographical Location | Country | Traditional BFFs | Ref. |
|---|---|---|---|
| Asia | Japan | Natto | [24] |
| Miso | [25] | ||
| China | Douchi | [26] | |
| Sufu/Furu | [27] | ||
| Korea | Chungkukjang/Chongkukjang | [18] | |
| Gochujang | [28] | ||
| Doenjang | [29] | ||
| Kanjang | [30] | ||
| India | Kinema | [31] | |
| Tungrymbai | [32] | ||
| Bekang | |||
| Dosa batter | [33] | ||
| Indonesian | Moromi | [34] | |
| Tempeh | [35] | ||
| Cambodia | Sieng | [36] | |
| Laos | |||
| Thailand | Thua Nao | [37] | |
| Nepal | Kinema | [31] | |
| Bhutan | |||
| Myanmar | Pepok | [36] | |
| Africa | Ghana and Nigeria | Dawadawa | [38] |
| Expression Systems | Hosts | Engineering Methods | Results | Ref. |
|---|---|---|---|---|
| Microorganisms | Bacillus spp. | Eight-proteases-gene-deficient B. subtilis WB800 | NK was expressed at a high level of 600 mg protein per liter culture medium | [94] |
| Multiple lytic genes deficient mutant B. subtilis LM2531 | The NK activity increased about 2.6-fold compared to B. subtilis 168 | [95] | ||
| Eight-proteases-gene-deficient B. licheniformis BL10 | The fermentation activity and product activity per unit of biomass of NK increased by 39% and 156% compared to B. licheniformis WX-02 | [96] | ||
| E. coli | Cytoplasmic expression NK in E. coli by cloning the aprN gene | The NK yield (49.3 mg/L) lower than that of B. subtilis YF38 | [51] | |
| E. coli DH5α acted as the host for plasmid construction from | 50.53% increase in NK activity compared to B. licheniformis DW2 | [98] | ||
| The aprN gene were cloned in E. coli pQE30 | 79.3 IU/mg fibrinolytic activity of NK compared to the 52.0 IU/mg from B. subtilis natto | [99] | ||
| LAB | The recombinant plasmid pMGthyA-ppNK carrying the aprN gene in L. bulgaricus | Recombinant strains expressed NK intracellularly, NK activity gradually decreased to 25% after 2 h in artificial gastric juice. The highest relative activity was approximately 94% after 3 h in artificial intestinal juice | [100] | |
| Eucaryote | Heterologous expression cloned the aprN gene on vector pPICZaA-NK and transformed into Pichia pastoris x-33 | The final fibrinolytic activity of NK is 195 U/mL | [97] | |
| Heterologous expression cloned the aprN gene in Pichia pastoris | Expressed high levels of NK approximately 9.5 g/L in high-density fermentation | [101] | ||
| Animals | Insects | Rv-egfp-NK (a recombinant baculovirus) containing the aprN gene expressed in Spodoptera frugiperda (SF-9) cells | The fibrinolytic activity of recombinant NK was 60 U/mL | [92] |
| Plants | Fruits | A synthetic gene (s NK) encoding NK was transiently expressed in Cucumis melo L. | The maximum fibrinolytic activity of 79.30 U/mL | [102] |
| Tobacco | Synthetic genes (sNK and sNKi) encoding NK was transiently expressed in tobacco leaves | The maximum fibrinolytic activity of 16.73 U/mL | [103] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Hou, L.; Hu, M.; Gao, Y.; Tian, Z.; Fan, B.; Li, S.; Wang, F. Recent Advances in Nattokinase-Enriched Fermented Soybean Foods: A Review. Foods 2022, 11, 1867. https://doi.org/10.3390/foods11131867
Li D, Hou L, Hu M, Gao Y, Tian Z, Fan B, Li S, Wang F. Recent Advances in Nattokinase-Enriched Fermented Soybean Foods: A Review. Foods. 2022; 11(13):1867. https://doi.org/10.3390/foods11131867
Chicago/Turabian StyleLi, Danfeng, Lizhen Hou, Miao Hu, Yaxin Gao, Zhiliang Tian, Bei Fan, Shuying Li, and Fengzhong Wang. 2022. "Recent Advances in Nattokinase-Enriched Fermented Soybean Foods: A Review" Foods 11, no. 13: 1867. https://doi.org/10.3390/foods11131867
APA StyleLi, D., Hou, L., Hu, M., Gao, Y., Tian, Z., Fan, B., Li, S., & Wang, F. (2022). Recent Advances in Nattokinase-Enriched Fermented Soybean Foods: A Review. Foods, 11(13), 1867. https://doi.org/10.3390/foods11131867