
The Antifreeze and Cryoprotective Activities of a Novel Antifreeze Peptide from Ctenopharyngodon idella Scales

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Pretreatment of Ctenopharyngodon idella Scales
2.3. Screening Proteases
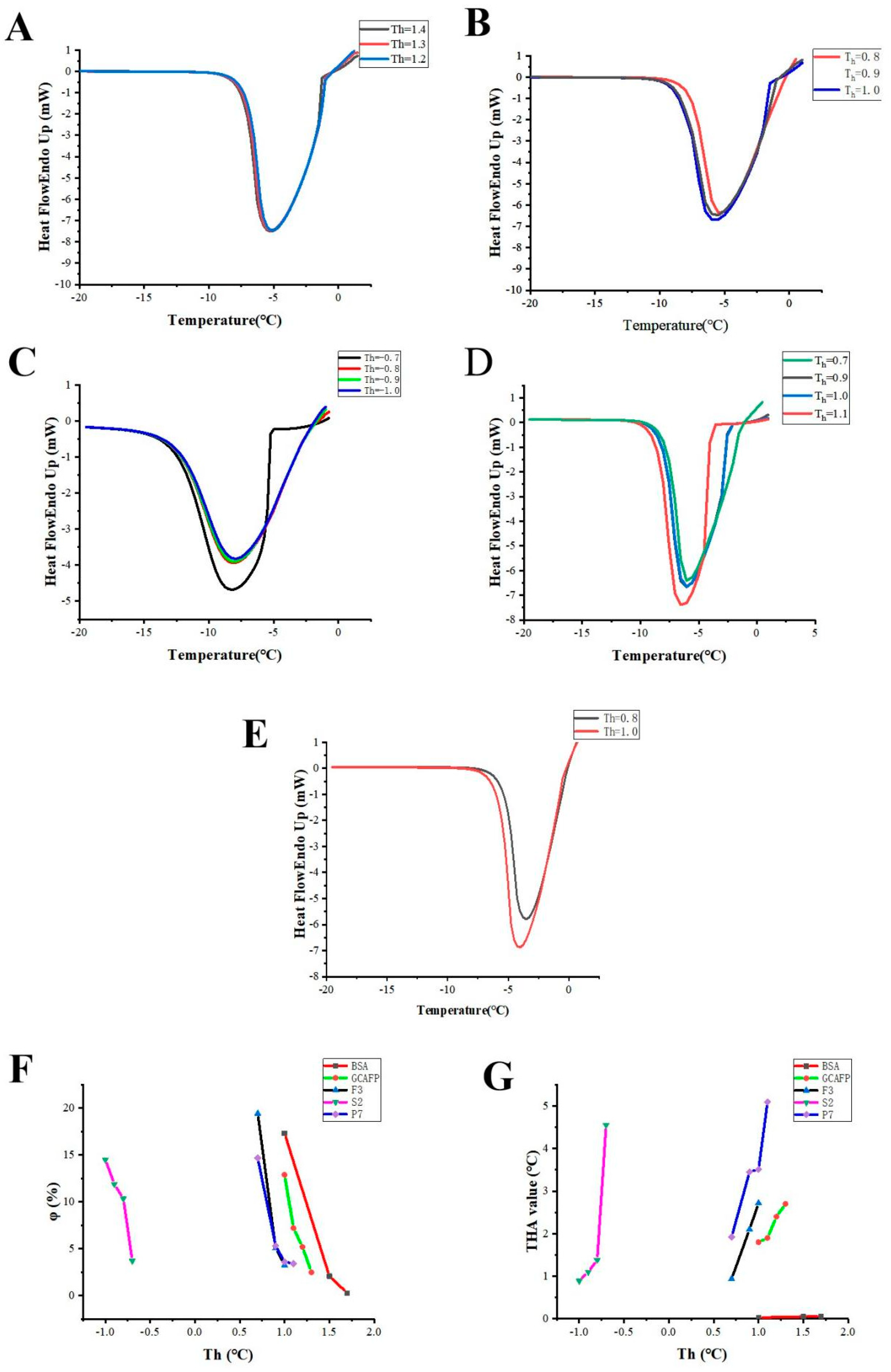
2.4. Optimizing AFP Preparation Conditions
2.5. Probing the Protection from Hypothermia
2.6. THA Determination
2.7. Isolating the Ice-Binding Collagen Peptides
2.7.1. Gel Filtration Chromatography
2.7.2. Isolation and Purification by Ion-Exchange Chromatography
2.7.3. Reversed-Phase HPLC
2.8. Peptide Sequence Analysis by Q Exactive LC-MS/MS
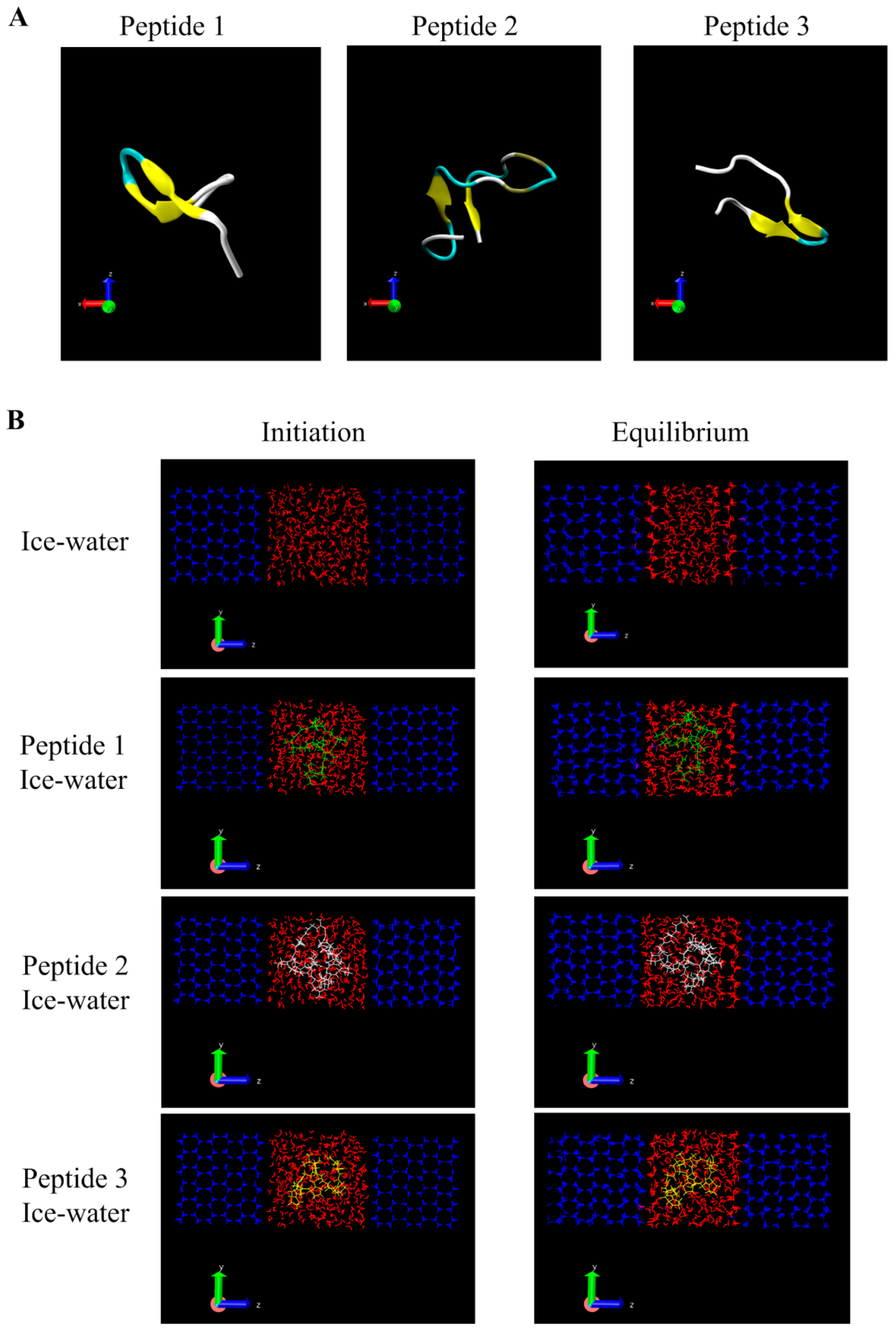
2.9. Homology Modeling of AFPs
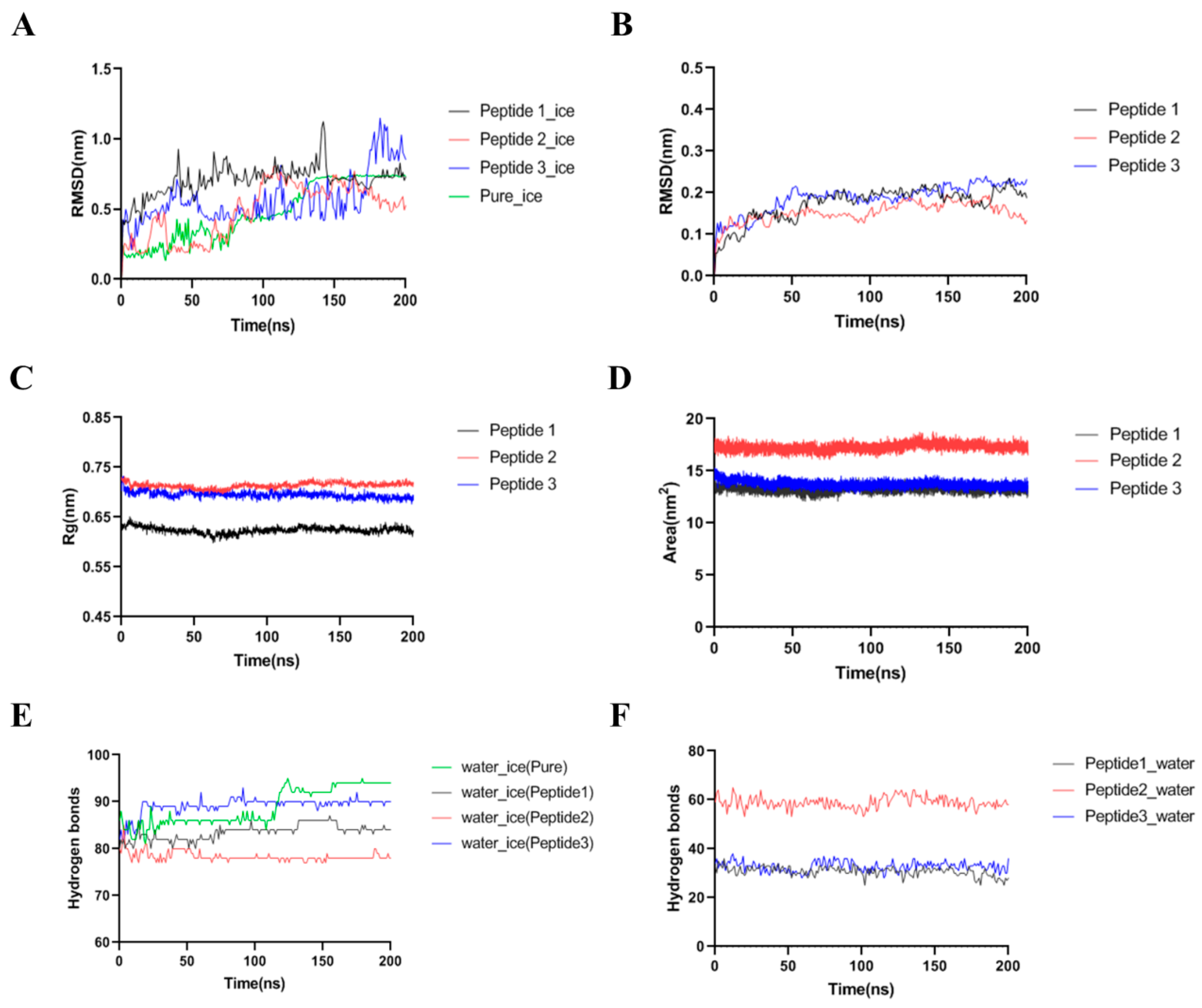
2.10. Molecular Dynamic Simulation of Ice-Water-Peptide System
2.11. Data Processing and Statistical Analysis
3. Results and Discussion
3.1. Optimizing the AFPs Preparation Conditions
3.2. Isolating Promising AFPs
3.3. Primary Structure of the Purified GCFSC-AFPs
3.4. Molecular Modeling of GCFSC-AFPs-Ice Interaction
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Feeney, R.E.; Yeh, Y. Antifreeze proteins: Current status and possible food uses. Trends Food Sci. Technol. 1998, 9, 102–106. [Google Scholar] [CrossRef]
- Chen, X.; Wu, J.; Cai, X.; Wang, S. Production, structure–function relationships, mechanisms, and applications of antifreeze peptides. Compr. Rev. Food Sci. Food Saf. 2021, 20, 542–562. [Google Scholar] [CrossRef] [PubMed]
- Graham, L.A.; Davies, P.L. Glycine-rich antifreeze proteins from snow fleas. Science 2005, 310, 461. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chen, M.; Wu, J.; Wang, S. Hypothermia protection effect of antifreeze peptides from pigskin collagen on freeze-dried Streptococcus thermophiles and its possible action mechanism. LWT-Food Sci. Technol. 2015, 63, 878–885. [Google Scholar] [CrossRef]
- Lin, J.; Hong, H.; Zhang, L.; Zhang, C.; Luo, Y. Antioxidant and cryoprotective effects of hydrolysate from gill protein of bighead carp (Hypophthalmichthys nobilis) in preventing denaturation of frozen surimi. Food Chem. 2019, 298, 124868. [Google Scholar] [CrossRef]
- Damodaran, S. Inhibition of ice crystal growth in ice cream mix by gelatin hydrolysate. J. Agric. Food Chem. 2007, 55, 10918–10923. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, J.; Chen, L.; Zhou, Y.; Wu, J. Preparation, isolation and hypothermia protection activity of antifreeze peptides from shark skin collagen. LWT-Food Sci. Technol. 2014, 55, 210–217. [Google Scholar] [CrossRef]
- Chen, X.; Wu, J.; Li, L.; Wang, S. Cryoprotective activity and action mechanism of antifreeze peptides obtained from tilapia scales on Streptococcus thermophilus during cold stress. J. Agric. Food Chem. 2019, 67, 1918–1926. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, H.; Wang, L.; Qian, H.; Qi, X. The preparation of antifreeze peptides from salmon skin gelatin and their effects on the thermal properties of frozen dough. Food Ferment. Ind. 2017, 43, 87–92. [Google Scholar]
- Fu, W.; Wang, P.; Chen, Y.; Lin, J.; Zheng, B.; Zeng, H.; Zhang, Y. Preparation, primary structure and antifreeze activity of antifreeze peptides from Scomberomorus niphonius skin. LWT 2019, 101, 670–677. [Google Scholar] [CrossRef]
- Cao, H.; Zhao, Y.; Zhu, Y.B.; Xu, F.; Yu, J.S.; Yuan, M. Antifreeze and cryoprotective activities of ice-binding collagen peptides from pig skin. Food Chem. 2016, 194, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.K.; Liu, D.C. Comparison of physical–chemical properties of type I collagen from different species. Food Chem. 2006, 99, 244–251. [Google Scholar] [CrossRef]
- Jia, C.; Huang, W.; Wu, C.; Lv, X.; Rayas-Duarte, P.; Zhang, L. Characterization and yeast cryoprotective performance for thermostable ice-structuring proteins from Chinese Privet (Ligustrum vulgare) leaves. Food Res. Int. 2012, 49, 280–284. [Google Scholar] [CrossRef]
- Ding, X.; Zhang, H.; Wang, L.; Qian, H.; Qi, X.; Xiao, J. Effect of barley antifreeze protein on thermal properties and water state of dough during freezing and freeze-thaw cycles. Food Hydrocoll. 2015, 47, 32–40. [Google Scholar] [CrossRef]
- Wu, J.; Rong, Y.; Wang, Z.; Zhou, Y.; Wang, S.; Zhao, B. Isolation and characterisation of sericin antifreeze peptides and molecular dynamics modelling of their ice-binding interaction. Food Chem. 2015, 174, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Haddad, Y.; Adam, V.; Heger, Z. Ten quick tips for homology modeling of high-resolution protein 3D structures. PLoS Comput. Biol. 2020, 16, e1007449. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Lamiable, A.; Thévenet, P.; Rey, J.; Vavrusa, M.; Derreumaux, P.; Tufféry, P. PEP-FOLD3: Faster de novo structure prediction for linear peptides in solution and in complex. Nucleic Acids Res. 2016, 44, W449–W454. [Google Scholar] [CrossRef] [Green Version]
- Lovell, S.C.; Davis, I.W.; Arendall, W.B., III; De Bakker, P.I.; Word, J.M.; Prisant, M.G.; Richardson, J.S.; Richardson, D.C. Structure validation by Cα geometry: ϕ, ψ and Cβ deviation. Proteins Struct. Funct. Bioinform. 2003, 50, 437–450. [Google Scholar] [CrossRef]
- Abraham, M.; van der Spoel, D.; Lindahl, E.; Hess, B.; Team, G. GROMACS User Manual, Version 2016.3; GROMACS Development Team: Groningen, The Netherlands, 2017.
- Best, R.B.; Zhu, X.; Shim, J.; Lopes, P.E.; Mittal, J.; Feig, M.; MacKerell, A.D., Jr. Optimization of the additive CHARMM all-atom protein force field targeting improved sampling of the backbone ϕ, ψ and side-chain χ1 and χ2 dihedral angles. J. Chem. Theory Comput. 2012, 8, 3257–3273. [Google Scholar] [CrossRef] [Green Version]
- Kaminski, G.A.; Friesner, R.A.; Tirado-Rives, J.; Jorgensen, W.L. Evaluation and reparametrization of the OPLS-AA force field for proteins via comparison with accurate quantum chemical calculations on peptides. J. Phys. Chem. B 2001, 105, 6474–6487. [Google Scholar] [CrossRef]
- Cerutti, D.S.; Duke, R.E.; Darden, T.A.; Lybrand, T.P. Staggered mesh Ewald: An extension of the smooth particle-mesh Ewald method adding great versatility. J. Chem. Theory Comput. 2009, 5, 2322–2338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Kun, H.; Mastai, Y. Activity of short segments of type I antifreeze protein. Pept. Sci. Orig. Res. Biomol. 2007, 88, 807–814. [Google Scholar] [CrossRef]
- Nobekawa, T.; Hagiwara, Y. Interaction among the twelve-residue segment of antifreeze protein type I, or its mutants, water and a hexagonal ice crystal. Mol. Simul. 2008, 34, 591–610. [Google Scholar] [CrossRef]
- Damodaran, S.; Wang, S. Ice crystal growth inhibition by peptides from fish gelatin hydrolysate. Food Hydrocoll. 2017, 70, 46–56. [Google Scholar] [CrossRef]
- Jinhong, H.J.W.S.W.; Pingfan, R. Preparation of Food-based Antifreeze Peptides and Research on the Ice Crystal Inhibition. J. Chin. Inst. Food Sci. Technol. 2013, 13, 11–18. [Google Scholar]
- Fei, Y.; Cao, P.; Gao, S.; Wang, B.; Wei, L.; Zhao, J.; Chen, G.; Wang, B. Purification and structure analysis of antifreeze proteins from Ammopiptanthus mongolicus. Prep. Biochem. Biotechnol. 2008, 38, 172–183. [Google Scholar] [CrossRef]
- Liu, Q.; Liu, Y.; Yu, J.; Wang, F.; Li, X. Identification and characterization of cryoprotective peptides extracted from silver carp (Hypophthalmichthys molitrix) hydrolysates. Int. J. Food Prop. 2019, 22, 1011–1023. [Google Scholar] [CrossRef] [Green Version]
- Carignano, M.; Shepson, P.; Szleifer, I. Molecular dynamics simulations of ice growth from supercooled water. Mol. Phys. 2005, 103, 2957–2967. [Google Scholar] [CrossRef]
- Kim, J.S.; Damodaran, S.; Yethiraj, A. Retardation of ice crystallization by short peptides. J. Phys. Chem. A 2009, 113, 4403–4407. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Yu, J.; Chen, X.; Zhang, Q.; Cai, X.; Ding, Y.; Zhou, X.; Wang, S. Dual cryoprotective strategies for ice-binding and stabilizing of frozen seafood: A review. Trends Food Sci. Technol. 2021, 111, 223–232. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Papain Concentration (U/g) | Temperature (°C) | pH | Substrate Concentration | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3000 | 3500 | 4000 | 4500 | 45 | 50 | 55 | 60 | 3 | 4 | 5 | 6 | 1:10 | 1:15 | 1:20 | BSA | |
| Th (°C) | 1.1 | 1.4 | 1.3 | 0.8 | 0.8 | 1.2 | 0.8 | 1.3 | 0.7 | 0.9 | 1.4 | 1.1 | 1.1 | 0.9 | 0.8 | 1.5 |
| To (°C) | −0.8 | −1.26 | −1.4 | 0 | 0.2 | 0.1 | 0.2 | 0.1 | −0.4 | −0.4 | −1.28 | −0.9 | −0.9 | −0.8 | 0 | 1.45 |
| ΔHr (J/g) | −361.1 | −401.1 | −386.1 | −329.8 | −404.4 | −371 | −402.1 | −363.7 | −249.4 | −369.5 | −398.9 | −409.2 | −360.2 | −320.9 | −347.7 | −378.6 |
| φ (%) | 0.77 | 1.67 | 2.48 | 1.61 | 1.10 | 0.78 | 0.60 | 0.08 | 7.32 | 2.33 | 1.48 | 2.57 | 3.30 | 2.30 | 4.77 | 0.02 |
| THA (°C) | 1.9b | 2.66a | 2.7a | 0.8c | 0.60b | 1.10a | 0.6b | 1.2a | 1.1c | 1.3c | 2.68a | 2.0b | 1.9a | 1.7b | 0.8c | 0.05 |
| Purification Step | THA (°C) | Purification Fold (Based on Activity) |
|---|---|---|
| Hydrolysate GCAFP | 2.68bc | 1.0b |
| Sephadex G-50 (Fraction F2) | 2.7b | 1.03b |
| SP-Sephadex C-25 (Fraction S2) | 4.55ab | 1.70a |
| C18 HPLC (Fraction P7) GCFSC-AFPs | 5.09a | 1.90a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dang, M.; Wang, R.; Jia, Y.; Du, J.; Wang, P.; Xu, Y.; Li, C. The Antifreeze and Cryoprotective Activities of a Novel Antifreeze Peptide from Ctenopharyngodon idella Scales. Foods 2022, 11, 1830. https://doi.org/10.3390/foods11131830
Dang M, Wang R, Jia Y, Du J, Wang P, Xu Y, Li C. The Antifreeze and Cryoprotective Activities of a Novel Antifreeze Peptide from Ctenopharyngodon idella Scales. Foods. 2022; 11(13):1830. https://doi.org/10.3390/foods11131830
Chicago/Turabian StyleDang, Meizhu, Ruifeng Wang, Yangyang Jia, Jing Du, Ping Wang, Yawei Xu, and Chunmei Li. 2022. "The Antifreeze and Cryoprotective Activities of a Novel Antifreeze Peptide from Ctenopharyngodon idella Scales" Foods 11, no. 13: 1830. https://doi.org/10.3390/foods11131830
APA StyleDang, M., Wang, R., Jia, Y., Du, J., Wang, P., Xu, Y., & Li, C. (2022). The Antifreeze and Cryoprotective Activities of a Novel Antifreeze Peptide from Ctenopharyngodon idella Scales. Foods, 11(13), 1830. https://doi.org/10.3390/foods11131830