
Improving Nutritional and Health Benefits of Biscuits by Optimizing Formulations Based on Sprouted Pseudocereal Grains

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals, Standards, and Reagents
2.2. Grains, Sprouting, and Milling Processes
2.3. Biscuit Making
- -
- Biscuit 1: SQF, SKF, and WF. This biscuit was coded as BQK.
- -
- Biscuit 2: SQF, SCF, and WF. This biscuit was coded as BQC.
- -
- Biscuit 3: SKF, SCH, and WF. This biscuit was coded as BKC.
2.4. Simulated Gastrointestinal Digestion
2.5. Chemical Characterization of Raw and Sprouted Cañihua, Kiwicha, and Quinoa Flours
2.6. Extraction and Quantification of Total Soluble Phenolic Compounds (TSPCs)
2.7. Determination of γ-Aminobutyric Acid (GABA)
2.8. Determination of Oxygen Radical Antioxidant Capacity (ORAC)
2.9. Estimation of In Vitro Glycemic Index
2.10. Simplex Centroid Mixture Design
2.11. Statistical Analysis
3. Results and Discussion
3.1. Effect of Germination on Nutritional Composition of Cañihua, Kiwicha, and Quinoa Flours
3.2. Effect of the WF Replacement by Sprouted Pseudocereal Flours on PA, GABA, TSPCs, and AA in Biscuits
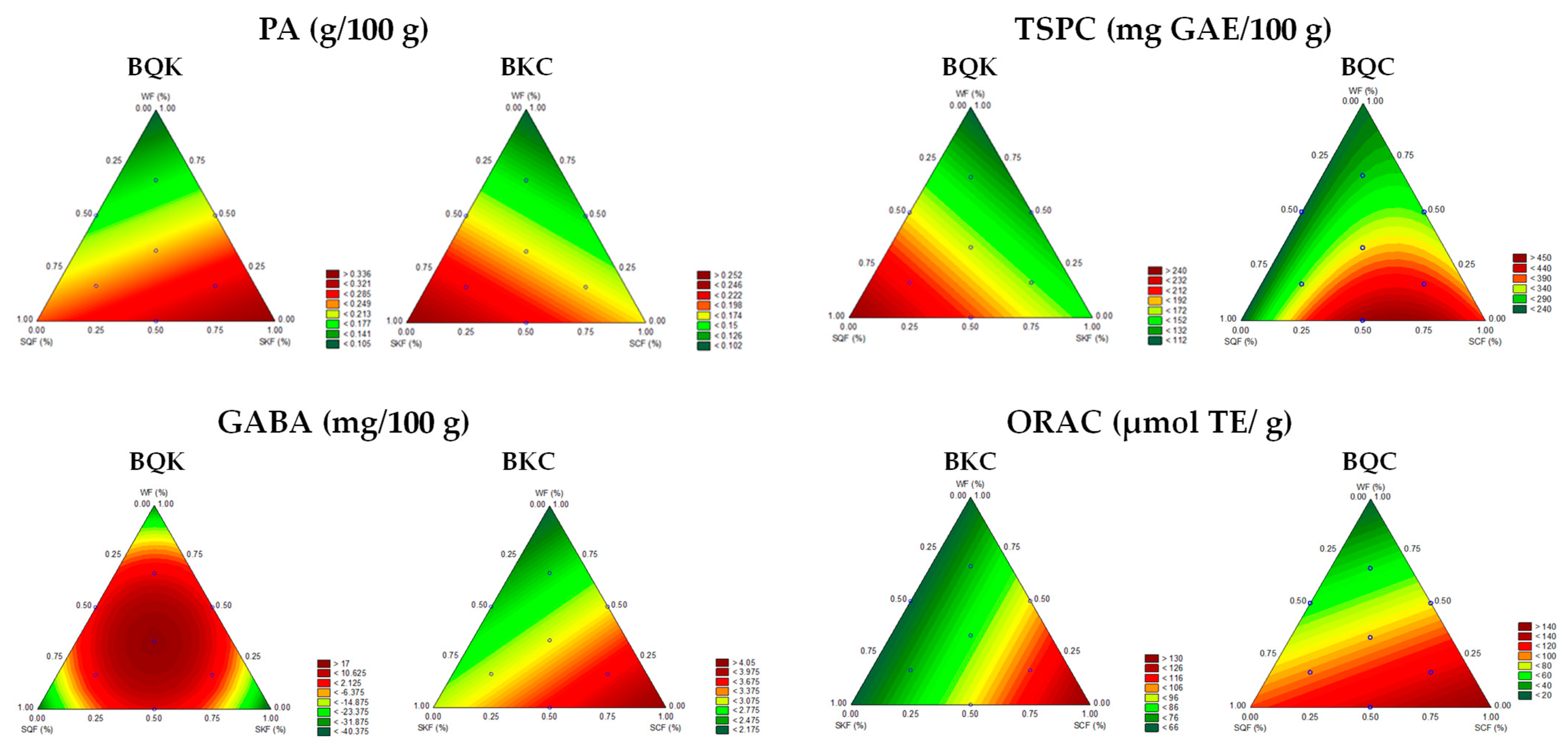
3.3. Modelization of Flour Formulation Effects on PA, GABA, TSPCs, and AA in Biscuits
3.4. In Vitro Digestion Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation
References
- De Bock, P.; Daelemans, L.; Selis, L.; Raes, K.; Vermeir, P.; Eeckhout, M.; Van Bockstaele, F. Comparison of the Chemical and Technological Characteristics of Wholemeal Flours Obtained from Amaranth (Amaranthus sp.), Quinoa (Chenopodium quinoa) and Buckwheat (Fagopyrum sp.) Seeds. Foods 2021, 10, 651. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Villaluenga, C.; Peñas, E.; Hernández-Ledesma, B. Pseudocereal grains: Nutritional value, health benefits and current applications for the development of gluten-free foods. Food Chem. Toxicol. 2020, 137, 111178. [Google Scholar] [CrossRef] [PubMed]
- Melini, V.; Melini, F. Functional Components and Anti-Nutritional Factors in Gluten-Free Grains: A Focus on Quinoa Seeds. Foods 2021, 10, 351. [Google Scholar] [CrossRef] [PubMed]
- Peñaranda, J.D.; Bueno, M.; Álvarez, F.; Pérez, P.D.; Perezábad, L. Sprouted grains in product development. Case studies of sprouted wheat for baking flours and fermented beverages. Int. J. Gastron. Food Sci. 2021, 25, 100375. [Google Scholar] [CrossRef]
- Ikram, A.; Saeed, F.; Afzaal, M.; Imran, A.; Niaz, B.; Tufail, T.; Hussain, M.; Anjum, F.M. Nutritional and end-use perspectives of sprouted grains: A comprehensive review. Food Sci. Nutr. 2021, 9, 4617–4628. [Google Scholar] [CrossRef]
- Pilco-Quesada, S.; Tian, Y.; Yang, B.; Repo-Carrasco-Valencia, R.; Suomela, J.-P. Effects of germination and kilning on the phenolic compounds and nutritional properties of quinoa (Chenopodium quinoa) and kiwicha (Amaranthus caudatus). J. Cereal Sci. 2020, 94, 102996. [Google Scholar] [CrossRef]
- Paucar-Menacho, L.M.; Martínez-Villaluenga, C.; Dueñas, M.; Frias, J.; Peñas, E. Response surface optimisation of germination conditions to improve the accumulation of bioactive compounds and the antioxidant activity in quinoa. Int. J. Food Sci. Technol. 2018, 53, 516–524. [Google Scholar] [CrossRef]
- Paucar-Menacho, L.M.; Peñas, E.; Dueñas, M.; Frias, J.; Martínez-Villaluenga, C. Optimizing germination conditions to enhance the accumulation of bioactive compounds and the antioxidant activity of kiwicha (Amaranthus caudatus) using response surface methodology. LWT-Food Sci. Technol. 2017, 76, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Darwish, A.M.G.; Al-Jumayi, H.A.O.; Elhendy, H.A. Effect of germination on the nutritional profile of quinoa (Cheopodium quinoa Willd.) seeds and its anti-anemic potential in Sprague–Dawley male albino rats. Cereal Chem. 2021, 98, 315–327. [Google Scholar] [CrossRef]
- Bhinder, S.; Kumari, S.; Singh, B.; Kaur, A.; Singh, N. Impact of germination on phenolic composition, antioxidant properties, antinutritional factors, mineral content and Maillard reaction products of malted quinoa flour. Food Chem. 2021, 346, 128915. [Google Scholar] [CrossRef]
- Aguilar, J.; Miano, A.C.; Obregón, J.; Soriano-Colchado, J.; Barraza-Jáuregui, G. Malting process as an alternative to obtain high nutritional quality quinoa flour. J. Cereal Sci. 2019, 90, 102858. [Google Scholar] [CrossRef]
- Abderrahim, F.; Huanatico, E.; Repo-Carrasco-Valencia, R.; Arribas, S.M.; Gonzalez, M.C.; Condezo-Hoyos, L. Effect of germination on total phenolic compounds, total antioxidant capacity, Maillard reaction products and oxidative stress markers in canihua (Chenopodium pallidicaule). J. Cereal Sci. 2012, 56, 410–417. [Google Scholar] [CrossRef]
- Hepsomali, P.; Groeger, J.A.; Nishihira, J.; Scholey, A. Effects of oral gamma-aminobutyric acid (GABA) administration on stress and sleep in humans: A systematic review. Front. Neurosci. 2020, 14, 923. [Google Scholar] [CrossRef] [PubMed]
- Ngo, D.-H.; Vo, T.S. An updated review on pharmaceutical properties of gamma-aminobutyric acid. Molecules 2019, 24, 2678. [Google Scholar] [CrossRef] [Green Version]
- Rico, D.; Penas, E.; Garcia, M.D.C.; Martinez-Villaluenga, C.; Rai, D.K.; Birsan, R.I.; Frias, J.; Martin-Diana, A.B. Sprouted Barley Flour as a Nutritious and Functional Ingredient. Foods 2020, 9, 926. [Google Scholar] [CrossRef] [Green Version]
- Aparicio-García, N.; Martínez-Villaluenga, C.; Frias, J.; Peñas, E. Sprouted oat as a potential gluten-free ingredient with enhanced nutritional and bioactive properties. Food Chem. 2021, 338, 127972. [Google Scholar] [CrossRef]
- Di Cairano, M.; Galgano, F.; Tolve, R.; Caruso, M.C.; Condelli, N. Focus on gluten free biscuits: Ingredients and issues. Trends Food Sci. Technol. 2018, 81, 203–212. [Google Scholar] [CrossRef]
- Mecha, E.; Correia, V.; Bento da Silva, A.; Ferreira, A.; Sepodes, B.; Figueira, M.E.; Vaz Patto, M.C.; Rosário Bronze, M. Improvement of wheat cookies’ nutritional quality, by partial substitution with common bean and maize flours, sustained human glycemia and enhanced satiety perception. Cereal Chem. 2021, 98, 1123–1134. [Google Scholar] [CrossRef]
- Laganà, V.; Giuffrè, A.M.; De Bruno, A.; Poiana, M. Formulation of Biscuits Fortified with a Flour Obtained from Bergamot By-Products (Citrus bergamia, Risso). Foods 2022, 11, 1137. [Google Scholar] [CrossRef]
- Cornell, J.A. Experiments with Mixtures: Designs, Models, and the Analysis of Mixture Data; John Wiley & Sons: Hoboken, NJ, USA, 2011; Volume 403. [Google Scholar]
- Azarbad, H.; Mazaheri Tehrani, M.; Rashidi, H. Optimization of gluten-free bread formulation using sorghum, rice, and millet flour by D-optimal mixture design approach. J. Agric. Sci. Technol. 2019, 21, 101–115. [Google Scholar]
- Bravo-Núñez, Á.; Sahagún, M.; Bravo-Núñez, A.; Gómez, M. Optimisation of protein-enriched gluten-free layer cakes using a mixture design. Int. J. Food Sci. Technol. 2020, 55, 2171–2178. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef] [PubMed]
- AACC. Approved Methods of AACC: Method 08-03, 30-10, 9th ed.; The American Association of Cereal Chemists: St. Paul, MN, USA, 2010. [Google Scholar]
- McKie, V.A.; McCleary, B.V. A Novel and Rapid Colorimetric Method for Measuring Total Phosphorus and Phytic Acid in Foods and Animal Feeds. J. AOAC Int. 2019, 99, 738–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pico, J.; Pismag, R.Y.; Laudouze, M.; Martinez, M.M. Systematic evaluation of the Folin–Ciocalteu and Fast Blue BB reactions during the analysis of total phenolics in legumes, nuts and plant seeds. Food Funct. 2020, 11, 9868–9880. [Google Scholar] [CrossRef]
- Goñi, I.; Garcia-Alonso, A.; Saura-Calixto, F. A starch hydrolysis procedure to estimate glycemic index. Nutr. Res. 1997, 17, 427–437. [Google Scholar] [CrossRef]
- Sanz-Penella, J.M.; Laparra, J.M.; Haros, M. Impact of α-Amylase During Breadmaking on In Vitro Kinetics of Starch Hydrolysis and Glycaemic Index of Enriched Bread with Bran. Plant Foods Hum. Nutr. 2014, 69, 216–221. [Google Scholar] [CrossRef] [Green Version]
- Galvan, D.; Effting, L.; Cremasco, H.; Conte-Junior, C.A. Recent Applications of Mixture Designs in Beverages, Foods, and Pharmaceutical Health: A Systematic Review and Meta-Analysis. Foods 2021, 10, 1941. [Google Scholar] [CrossRef]
- Lemmens, E.; Moroni, A.V.; Pagand, J.; Heirbaut, P.; Ritala, A.; Karlen, Y.; Lê, K.-A.; Van den Broeck, H.C.; Brouns, F.J.P.H.; De Brier, N.; et al. Impact of Cereal Seed Sprouting on Its Nutritional and Technological Properties: A Critical Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 305–328. [Google Scholar] [CrossRef] [Green Version]
- Cornejo, F.; Novillo, G.; Villacrés, E.; Rosell, C.M. Evaluation of the physicochemical and nutritional changes in two amaranth species (Amaranthus quitensis and Amaranthus caudatus) after germination. Food Res. Int. 2019, 121, 933–939. [Google Scholar] [CrossRef]
- Perri, G.; Calabrese, F.M.; Rizzello, C.G.; De Angelis, M.; Gobbetti, M.; Calasso, M. Sprouting process affects the lactic acid bacteria and yeasts of cereal, pseudocereal and legume flours. LWT 2020, 126, 109314. [Google Scholar] [CrossRef]
- Padmashree, A.; Negi, N.; Handu, S.; Khan, M.A.; Semwal, A.D.; Sharma, G.K. Effect of germination on nutritional, antinutritional and rheological characteristics of chenopodium quinoa. Def. Life Sci. J. 2019, 4, 55–60. [Google Scholar] [CrossRef]
- Finch-Savage, B. Seeds: Physiology of development, germination and dormancy (3rd edition)—Book review. Seed Sci. Res. 2013, 23, 289. [Google Scholar] [CrossRef]
- Paucar-Menacho, L.M.; Dueñas, M.; Peñas, E.; Frias, J.; Martínez-Villaluenga, C. Effect of Dry Heat Puffing on Nutritional Composition, Fatty Acid, Amino Acid and Phenolic Profiles of Pseudocereals Grains. Pol. J. Food Nutr. Sci. 2018, 68, 289–297. [Google Scholar] [CrossRef]
- Demir, B.; Bilgiçli, N. Changes in chemical and anti-nutritional properties of pasta enriched with raw and germinated quinoa (Chenopodium quinoa Willd.) flours. J. Food Sci. Technol. 2020, 57, 3884–3892. [Google Scholar] [CrossRef]
- Velarde-Salcedo, A.J.; Bojórquez-Velázquez, E.; de la Rosa, A.P.B. Amaranth. In Whole Grains and Their Bioactives; John Wiley & Sons: Hoboken, NJ, USA, 2019; Chapter 8; pp. 209–250. [Google Scholar] [CrossRef]
- Diez-Gutiérrez, L.; San Vicente, L.; Barrón, L.J.R.; Villarán, M.d.C.; Chávarri, M. Gamma-aminobutyric acid and probiotics: Multiple health benefits and their future in the global functional food and nutraceuticals market. J. Funct. Foods 2020, 64, 103669. [Google Scholar] [CrossRef]
- Ding, J.; Hou, G.G.; Nemzer, B.V.; Xiong, S.; Dubat, A.; Feng, H. Effects of controlled germination on selected physicochemical and functional properties of whole-wheat flour and enhanced γ-aminobutyric acid accumulation by ultrasonication. Food Chem. 2018, 243, 214–221. [Google Scholar] [CrossRef]
- Chungcharoen, T.; Prachayawarakorn, S.; Tungtrakul, P.; Soponronnarit, S. Effects of germination time and drying temperature on drying characteristics and quality of germinated paddy. Food Bioprod. Proc. 2015, 94, 707–716. [Google Scholar] [CrossRef]
- Peñarrieta, J.M.; Alvarado, J.A.; Åkesson, B.; Bergenståhl, B. Total antioxidant capacity and content of flavonoids and other phenolic compounds in canihua (Chenopodium pallidicaule): An Andean pseudocereal. Mol. Nutr. Food Res. 2008, 52, 708–717. [Google Scholar] [CrossRef]
- Li, H.; Deng, Z.; Liu, R.; Zhu, H.; Draves, J.; Marcone, M.; Sun, Y.; Tsao, R. Characterization of phenolics, betacyanins and antioxidant activities of the seed, leaf, sprout, flower and stalk extracts of three Amaranthus species. J. Food Comp. Anal. 2015, 37, 75–81. [Google Scholar] [CrossRef]
- Abderrahim, F.; Huanatico, E.; Segura, R.; Arribas, S.; Gonzalez, M.C.; Condezo-Hoyos, L. Physical features, phenolic compounds, betalains and total antioxidant capacity of coloured quinoa seeds (Chenopodium quinoa Willd.) from Peruvian Altiplano. Food Chem. 2015, 183, 83–90. [Google Scholar] [CrossRef]
- Tomé-Sánchez, I.; Martín-Diana, A.B.; Peñas, E.; Bautista-Expósito, S.; Frias, J.; Rico, D.; González-Maillo, L.; Martinez-Villaluenga, C. Soluble Phenolic Composition Tailored by Germination Conditions Accompany Antioxidant and Anti-inflammatory Properties of Wheat. Antioxidants 2020, 9, 426. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, A.V.F.; Tetens, I.; Meyer, A.S. Potential of Phytase-Mediated Iron Release from Cereal-Based Foods: A Quantitative View. Nutrients 2013, 5, 3074–3098. [Google Scholar] [CrossRef] [Green Version]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-aminobutyric acid as a bioactive compound in foods: A review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Garzón, A.G.; Erben, M.; Osella, C.A.; Drago, S.R. Effects of baking on γ-aminobutyric acid and free phenolic acids from gluten-free cookies made with native and malted whole sorghum flours. J. Food Proc. Preserv. 2020, 44, e14571. [Google Scholar] [CrossRef]
- Oketch-Rabah, H.A.; Madden, E.F.; Roe, A.L.; Betz, J.M. United States Pharmacopeia (USP) Safety Review of Gamma-Aminobutyric Acid (GABA). Nutrients 2021, 13, 2742. [Google Scholar] [CrossRef] [PubMed]
- Cannas, M.; Pulina, S.; Conte, P.; Del Caro, A.; Urgeghe, P.P.; Piga, A.; Fadda, C. Effect of Substitution of Rice Flour with Quinoa Flour on the Chemical-Physical, Nutritional, Volatile and Sensory Parameters of Gluten-Free Ladyfinger Biscuits. Foods 2020, 9, 808. [Google Scholar] [CrossRef]
- Demir, M.K.; Kilinc, M. Utilization of quinoa flour in cookie production. Int. Food Res. J. 2017, 24, 2394–2401. [Google Scholar]
- Jan, R.; Saxena, D.C.; Singh, S. Physico-chemical, textural, sensory and antioxidant characteristics of gluten—Free cookies made from raw and germinated Chenopodium (Chenopodium album) flour. LWT-Food Sci. Technol. 2016, 71, 281–287. [Google Scholar] [CrossRef]
- Costa, C.; Tsatsakis, A.; Mamoulakis, C.; Teodoro, M.; Briguglio, G.; Caruso, E.; Tsoukalas, D.; Margina, D.; Dardiotis, E.; Kouretas, D.; et al. Current evidence on the effect of dietary polyphenols intake on chronic diseases. Food Chem. Toxicol. 2017, 110, 286–299. [Google Scholar] [CrossRef]
- Kapolou, A.; Karantonis, H.C.; Rigopoulos, N.; Koutelidakis, A.E. Association of Mean Daily Polyphenols Intake with Mediterranean Diet Adherence and Anthropometric Indices in Healthy Greek Adults: A Retrospective Study. Appl. Sci. 2021, 11, 4664. [Google Scholar] [CrossRef]
- Chauhan, A.; Saxena, D.C.; Singh, S. Total dietary fibre and antioxidant activity of gluten free cookies made from raw and germinated amaranth (Amaranthus spp.) flour. LWT-Food Sci. Technol. 2015, 63, 939–945. [Google Scholar] [CrossRef]
- Brouns, F. Phytic Acid and Whole Grains for Health Controversy. Nutrients 2022, 14, 25. [Google Scholar] [CrossRef] [PubMed]
- Hussin, F.S.; Chay, S.Y.; Hussin, A.S.M.; Wan Ibadullah, W.Z.; Muhialdin, B.J.; Abd Ghani, M.S.; Saari, N. GABA enhancement by simple carbohydrates in yoghurt fermented using novel, self-cloned Lactobacillus plantarum Taj-Apis362 and metabolomics profiling. Sci. Rep. 2021, 11, 9417. [Google Scholar] [CrossRef] [PubMed]
- Dala-Paula, B.M.; Deus, V.L.; Tavano, O.L.; Gloria, M.B.A. In vitro bioaccessibility of amino acids and bioactive amines in 70% cocoa dark chocolate: What you eat and what you get. Food Chem. 2021, 343, 128397. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, A.; Ferraretto, A.; De Noni, I.; Bottani, M.; Cattaneo, S.; Galli, S.; Brandolini, A. Bioactive compounds and antioxidant properties of pseudocereals-enriched water biscuits and their In vitro digestates. Food Chem. 2018, 240, 799–807. [Google Scholar] [CrossRef]
- Wang, X.; Lao, X.; Bao, Y.; Guan, X.; Li, C. Effect of whole quinoa flour substitution on the texture and In vitro starch digestibility of wheat bread. Food Hydrocol. 2021, 119, 106840. [Google Scholar] [CrossRef]
- Wolter, A.; Hager, A.-S.; Zannini, E.; Arendt, E.K. In vitro starch digestibility and predicted glycaemic indexes of buckwheat, oat, quinoa, sorghum, teff and commercial gluten-free bread. J. Cereal Sci. 2013, 58, 431–436. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Parameters | WF | CF | SCF | KF | SKF | QF | SQF |
|---|---|---|---|---|---|---|---|
| Starch (g/100 g dw) | 75.07 ± 0.97 f | 46.66 ± 0.45 d | 41.21 ± 1.47 c | 48.33 ± 0.93 d | 32.45 ± 0.85 b | 40.98 ± 0.88 c | 16.97± 0.85 a |
| Protein (g/100 g dw) | 12.26 ± 0.06 a | 20.61 ± 0.26 g | 19.11 ± 0.27 f | 16.05 ± 0.14 d | 15.38 ± 0.11 c | 16.87 ± 0.13 e | 13.52 ± 0.26 b |
| Fat (g/100 g dw) | 0.61 ± 0.04 a | 6.18 ± 0.04 d | 6.23 ± 0.25 d | 5.10 ± 0.08 b | 5.86 ± 0.38 cd | 5.21 ± 0.07 bc | 6.55 ± 0.11 e |
| Ash (g/100 g dw) | 0.55 ± 0.05 a | 2.76 ± 0.13 c | 2.68 ± 0.05 c | 2.66 ± 0.19 bc | 2.85 ± 0.08 c | 2.48 ± 0.17 bc | 2.29 ± 0.08 b |
| PA (g/100 g dw) | 0.09 ± 0.01 a | 1.17 ± 0.02 b | 0.88 ± 0.01 b | 1.23 ± 0.02 c | 1.24 ± 0.02 c | 0.90 ± 0.02 b | 0.93 ± 0.02 b |
| GABA (mg/100 g dw) | 12.96 ± 0.35 a | 24.34 ± 4.83 b | 217.98 ± 1.48 d | 37.38 ± 1.58 b | 100.00 ± 22.45 c | 32.98 ± 4.42 b | 202.54 ± 32.05 d |
| TSPC (mg GAE/100 g) | 44.27 ± 2.43 a | 314.39 ± 22.38 d | 386.12 ± 27.83 e | 149.27 ± 1.80 b | 244.72 ± 2.09 c | 525.5 ± 38.14 f | 612.81 ± 13.25 g |
| ORAC (μmol TE/g) | 20.64 ± 2.71 b | 48.74 ± 5.98 b | 114.92 ± 14.17 c | 10.23 ± 2.16 a | 35.44 ± 4.55 d | 46.62 ± 3.53 b | 45.30 ± 3.96 d |
| Biscuit Type | Recipe No. | Proportion of Flours a | PA (g/100 g) | GABA (mg/100 g) | TSPC (mg GAE/100 g) | ORAC (μmol TE/g) | ||
|---|---|---|---|---|---|---|---|---|
| Sprouted Pseudocereal Flour 1 (X1) | Sprouted Pseudocereal Flour 2 (X2) | Wheat Flour (X3) | ||||||
| BQK | 1 | 15 | 15 | 70 | 0.22 ± 0.00 c | 4.01 ± 0.06 de | 148.38 ± 3.80 bcd | 57.03 ± 1.39 bc |
| 2 | 20 | 20 | 60 | 0.27 ± 0.01 de | 4.41 ± 0.11 e | 192.96 ± 1.69 fgh | 72.43 ± 6.25 def | |
| 3 | 5 | 20 | 75 | 0.23 ± 0.01 c | 2.44 ± 0.16 b | 133.00 ± 0.68 bc | 52.50 ± 6.27 b | |
| 4 | 20 | 20 | 60 | 0.32 ± 0.01 fg | 3.62 ± 0.26 cd | 222.30 ± 4.16 hi | 91.76 ± 1.94 g | |
| 5 | 5 | 20 | 75 | 0.22 ± 0.01 c | 2.54 ± 0.05 b | 125.71 ± 5.56 b | 59.83 ± 2.33 bc | |
| 6 | 5 | 20 | 75 | 0.22 ± 0.00 c | 2.50 ± 0.04 b | 128.86 ± 3.74 b | 52.39 ± 0.50 b | |
| 7 | 20 | 20 | 60 | 0.33 ± 0.01 g | 3.37 ± 0.04 c | 246.49 ± 15.01 i | 75.81 ± 2.44 f | |
| 8 | 10 | 25 | 65 | 0.29 ± 0.00 ef | 3.73 ± 0.00 cd | 160.46 ± 4.87 cde | 52.71 ± 1.50 b | |
| 9 | 20 | 5 | 75 | 0.16 ± 0.01 b | 3.85 ± 0.18 cd | 165.11 ± 4.65 def | 74.57 ± 5.01 ef | |
| 10 | 20 | 5 | 75 | 0.17 ± 0.00 b | 3.41 ± 0.07 c | 188.39 ± 16.35 efg | 63.03 ± 5.02 bcde | |
| 11 | 10 | 10 | 80 | 0.18 ± 0.02 b | 2.63 ± 0.02 b | 151.36 ± 9.06 bcd | 53.98 ± 2.48 bc | |
| 12 | 25 | 10 | 65 | 0.24 ± 0.01 cd | 4.49 ± 0.08 e | 212.15 ± 2.73 gh | 66.14 ± 5.52 cdef | |
| 13 | 20 | 5 | 75 | 0.22 ± 0.01 c | 4.12 ± 0.18 de | 222.31 ± 1.08 hi | 60.01 ± 4.08 bcd | |
| 14 | 25 | 10 | 65 | 0.15 ± 0.01 b | 3.38 ± 0.01 c | 175.10 ± 3.36 def | 51.30 ± 4.32 b | |
| BQC | 1 | 15 | 15 | 70 | 0.19 ± 0.00 de | 3.50 ± 0.01 bcd | 409.48 ± 38.06 ef | 95.46 ± 2.45 c |
| 2 | 20 | 20 | 60 | 0.25 ± 0.01 f | 4.66 ± 0.06 de | 486.86 ± 15.14 f | 133.28 ± 2.65 d | |
| 3 | 5 | 20 | 75 | 0.18 ± 0.00 cde | 4.01 ± 0.02 cde | 334.40 ± 2.85 de | 91.60 ± 6.44 c | |
| 4 | 20 | 20 | 60 | 0.23 ± 0.00 f | 4.58 ± 0.28 de | 441.61 ± 14.63 f | 138.39 ± 2.03 d | |
| 5 | 5 | 20 | 75 | 0.15 ± 0.00 b | 4.06 ± 0.08 cde | 320.15 ± 26.96 cd | 84.49 ± 0.25 c | |
| 6 | 5 | 20 | 75 | 0.17 ± 0.00 bcd | 3.63 ± 0.03 bcde | 280.14 ± 2.58 bcd | 89.03 ± 4.07 c | |
| 7 | 20 | 20 | 60 | 0.25 ± 0.00 f | 4.94 ± 0.22 e | 428.78 ± 6.39 f | 136.25 ± 2.19 d | |
| 8 | 10 | 25 | 65 | 0.23 ± 0.00 f | 3.60 ± 0.41 bcde | 411.83 ± 21.66 ef | 98.35 ± 8.20 c | |
| 9 | 20 | 5 | 75 | 0.18 ± 0.00 cde | 3.18 ± 0.02 bc | 204.43 ± 5.60 b | 67.38 ± 2.92 b | |
| 10 | 20 | 5 | 75 | 0.19 ± 0.00 cde | 3.12 ± 0.03 bc | 236.8 ± 0.310 b | 61.64 ± 5.84 b | |
| 11 | 10 | 10 | 80 | 0.17 ± 0.00 bc | 2.61 ± 0.02 b | 275.67 ± 3.05 bcd | 56.06 ± 5.06 b | |
| 12 | 25 | 10 | 65 | 0.24 ± 0.00 f | 4.58 ± 0.00 de | 339.32 ± 24.67 de | 90.22 ± 9.48 c | |
| 13 | 20 | 5 | 75 | 0.20 ± 0.01 e | 3.98 ± 1.03 cde | 247.89 ± 19.14 bc | 56.68 ± 5.22 b | |
| 14 | 25 | 10 | 65 | 0.23 ± 0.00 f | 6.47 ± 0.02 f | 325.52 ± 24.66 cd | 92.66 ± 2.98 c | |
| BKC | 1 | 15 | 15 | 70 | 0.15 ± 0.01 bc | 3.32 ± 0.05 ef | 228.92 ± 16.62 def | 93.69 ± 1.03 ef |
| 2 | 20 | 20 | 60 | 0.22 ± 0.01 e | 3.36 ± 0.08 ef | 328.63 ± 0.00 g | 98.96 ± 14.25 fg | |
| 3 | 5 | 20 | 75 | 0.15 ± 0.00 bc | 3.32 ± 0.09 ef | 260.76 ± 7.13 ef | 114.45 ± 2.37 g | |
| 4 | 20 | 20 | 60 | 0.21 ± 0.01 e | 3.64 ± 0.00 f | 279.73 ± 25.94 fg | 106.35 ± 3.91 fg | |
| 5 | 5 | 20 | 75 | 0.14 ± 0.00 b | 3.06 ± 0.03 cde | 230.75 ± 19.53 def | 106.14 ± 1.32 fg | |
| 6 | 5 | 20 | 75 | 0.14 ± 0.00 b | 3.23 ± 0.07 e | 287.82 ± 20.39 fg | 89.74 ± 2.97 def | |
| 7 | 20 | 20 | 60 | 0.25 ± 0.00 f | 4.11 ± 0.06 g | 236.09 ± 22.07 def | 99.16 ± 1.75 fg | |
| 8 | 10 | 25 | 65 | 0.17 ± 0.00 d | 3.32 ± 0.04 ef | 185.48 ± 13.65 bcd | 100.34 ± 3.40 fg | |
| 9 | 20 | 5 | 75 | 0.22 ± 0.01 e | 2.58 ± 0.05 b | 285.27 ± 8.29 fg | 68.19 ± 2.21 bc | |
| 10 | 20 | 5 | 75 | 0.16 ± 0.00 cd | 2.80 ± 0.06 bcd | 127.11 ± 4.57 b | 80.44 ± 4.35 cde | |
| 11 | 10 | 10 | 80 | 0.14 ± 0.00 b | 2.52 ± 0.10 b | 167.04 ± 13.22 bc | 62.95 ± 0.94 b | |
| 12 | 25 | 10 | 65 | 0.14 ± 0.00 b | 2.71 ± 0.22 bc | 240.33 ± 3.93 def | 67.51 ± 0.33 bc | |
| 13 | 20 | 5 | 75 | 0.18 ± 0.01 d | 2.46 ± 0.06 b | 147.29 ± 5.00 b | 59.81 ± 1.33 b | |
| 14 | 25 | 10 | 65 | 0.26 ± 0.01 f | 3.13 ± 0.13 de | 218.18 ± 8.62 cde | 76.48 ± 11.73 bcd | |
| Control | 15 | 0 | 0 | 100 | 0.04 ± 0.01 a | 0.70 ± 0.04 a | 52.59 ± 3.74 a | 25.50 ± 2.80 a |
| Biscuit Type | Dependent Variables | Mathematical Models | R2 (Pred) | R2 (Adj) |
|---|---|---|---|---|
| BQK | PA | ŷ = 0.25X1 + 0.44X2 + 0.18X3 | 0.88 | 0.80 |
| GABA | ŷ = −39.67X1 − 41.12X2 − 31.30X3 + 179.69X1X2 + 154.65X1X3 + 161.99X2X3 | 0.76 | 0.71 | |
| TSPC | ŷ = 130.89X1 + 291.14X2 + 368.00X3 | 0.78 | 0.74 | |
| BQC | TSPC | ŷ = −1.33X1 + 4.00X2 + 2.34X3 + 0.67X1X2 | 0.86 | 0.89 |
| ORAC | ŷ = 2.68X1 + 4.24X2 − 0.17X3 | 0.84 | 0.87 | |
| BKC | PA | ŷ = 0.25X1 + 0.18X2 + 0.10X3 | 0.80 | 0.81 |
| GABA | ŷ = 3.02X1 + 4.18X2 + 2.14X3 | 0.74 | 0.70 | |
| ORAC | ŷ = 65.46X1 + 133.01X2 + 66.30X3 | 0.74 | 0.69 |
| Biscuit Type | Optimum Desirability Value (D) | Flour Formulation at Optimum D | Response Variables | Predicted Values | −95% CI | +95% CI |
|---|---|---|---|---|---|---|
| BQK | 0.487 | 21% SQF, 16% SKF, 63% WF | PA (g/100 g) | 0.24 | 0.16 | 0.33 |
| GABA (mg/100 g) | 13.57 | 2.44 | 17.78 | |||
| TSPC (mg GAE/100 g) | 177.95 | 125.72 | 246.49 | |||
| BQC | 0.959 | 15% SQF, 25% SCF, 60% WF | TSPC (mg GAE/100 g) | 472.44 | 437.96 | 506.92 |
| ORAC (μmol TE/g) | 135.87 | 124.70 | 147.04 | |||
| BKC | 0.704 | 5% SKF, 23% SCF, 72% WF | PA (g/100 g) | 0.15 | 0.11 | 0.19 |
| GABA (mg/100 g) | 3.37 | 3.09 | 3.65 | |||
| ORAC (μmol TE/g) | 106.53 | 95.38 | 117.67 |
| Digestion Phase/Time | Parameters | Control Biscuit | BQC |
|---|---|---|---|
| None/0 min | PA (g/100 g) | 0.21 ± 0.01 a | 0.25 ± 0.00 a,* |
| GABA (mg/100 g) | 0.7 ± 0.04 a | 6.23 ± 0.06 b,* | |
| TSPC (mg GAE/100 g) | 52.59 ± 3.74 a | 428.78 ± 6.39 a,* | |
| ORAC (μmol TE/g) | 25.51 ± 2.80 a | 136.25 ± 2.19 b,* | |
| Gastric/120 min | PA (g/100 g) | 0.32 ± 0.01 a | 0.25 ± 0.07 a,* |
| GABA (mg/100 g) | 0.67 ± 0.06 a | 6.34 ± 0.48 a,* | |
| TSPC (mg GAE/100 g) | 128.29 ± 0.99 b | 429.28 ± 6.77 a,* | |
| ORAC (μmol TE/g) | 41.69 ± 0.62 b | 119.47 ± 12.60 ab,* | |
| Intestinal/120 min | PA (g/100 g) | 0.26 ± 0.39 a | 0.39 ± 0.04 b,* |
| GABA (mg/100 g) | 1.75 ± 0.06 b | 8.97 ± 0.11 b,* | |
| TSPC (mg GAE/100 g) | 401.73 ± 8.19 c | 638.49 ± 2.65 b,* | |
| ORAC (μmol TE/g) | 103.59 ± 8.02 c | 195.26 ± 8.48 c,* | |
| In vitro starch digestibility | HI | 87.3 ± 0.27 | 75.99 ± 0.96 * |
| AUC | 27,633.1 ± 85.8 | 24,046.0 ± 304.5 * | |
| GI | 87.65 ± 0.15 | 81.43 ± 0.53 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paucar-Menacho, L.M.; Simpalo-López, W.D.; Castillo-Martínez, W.E.; Esquivel-Paredes, L.J.; Martínez-Villaluenga, C. Improving Nutritional and Health Benefits of Biscuits by Optimizing Formulations Based on Sprouted Pseudocereal Grains. Foods 2022, 11, 1533. https://doi.org/10.3390/foods11111533
Paucar-Menacho LM, Simpalo-López WD, Castillo-Martínez WE, Esquivel-Paredes LJ, Martínez-Villaluenga C. Improving Nutritional and Health Benefits of Biscuits by Optimizing Formulations Based on Sprouted Pseudocereal Grains. Foods. 2022; 11(11):1533. https://doi.org/10.3390/foods11111533
Chicago/Turabian StylePaucar-Menacho, Luz María, Wilson Daniel Simpalo-López, Williams Esteward Castillo-Martínez, Lourdes Jossefyne Esquivel-Paredes, and Cristina Martínez-Villaluenga. 2022. "Improving Nutritional and Health Benefits of Biscuits by Optimizing Formulations Based on Sprouted Pseudocereal Grains" Foods 11, no. 11: 1533. https://doi.org/10.3390/foods11111533
APA StylePaucar-Menacho, L. M., Simpalo-López, W. D., Castillo-Martínez, W. E., Esquivel-Paredes, L. J., & Martínez-Villaluenga, C. (2022). Improving Nutritional and Health Benefits of Biscuits by Optimizing Formulations Based on Sprouted Pseudocereal Grains. Foods, 11(11), 1533. https://doi.org/10.3390/foods11111533