
Peeling Affects the Nutritional Properties of Carrot Genotypes




Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials, Collecting Site and Sampling
2.2. Standards and Reagents
2.3. Inorganic Cations and Anions
2.4. Simple Sugars and Sweetness Index
2.5. Carotenoids
2.6. Organic Acids
2.7. Phenolic Compounds
2.8. Statistical Analysis
3. Results and Discussion
3.1. Bio-Morphological Traits
3.2. Inorganic Ions
3.3. Simple Sugars
3.4. Carotenoids
3.5. Organic Acids (OA)
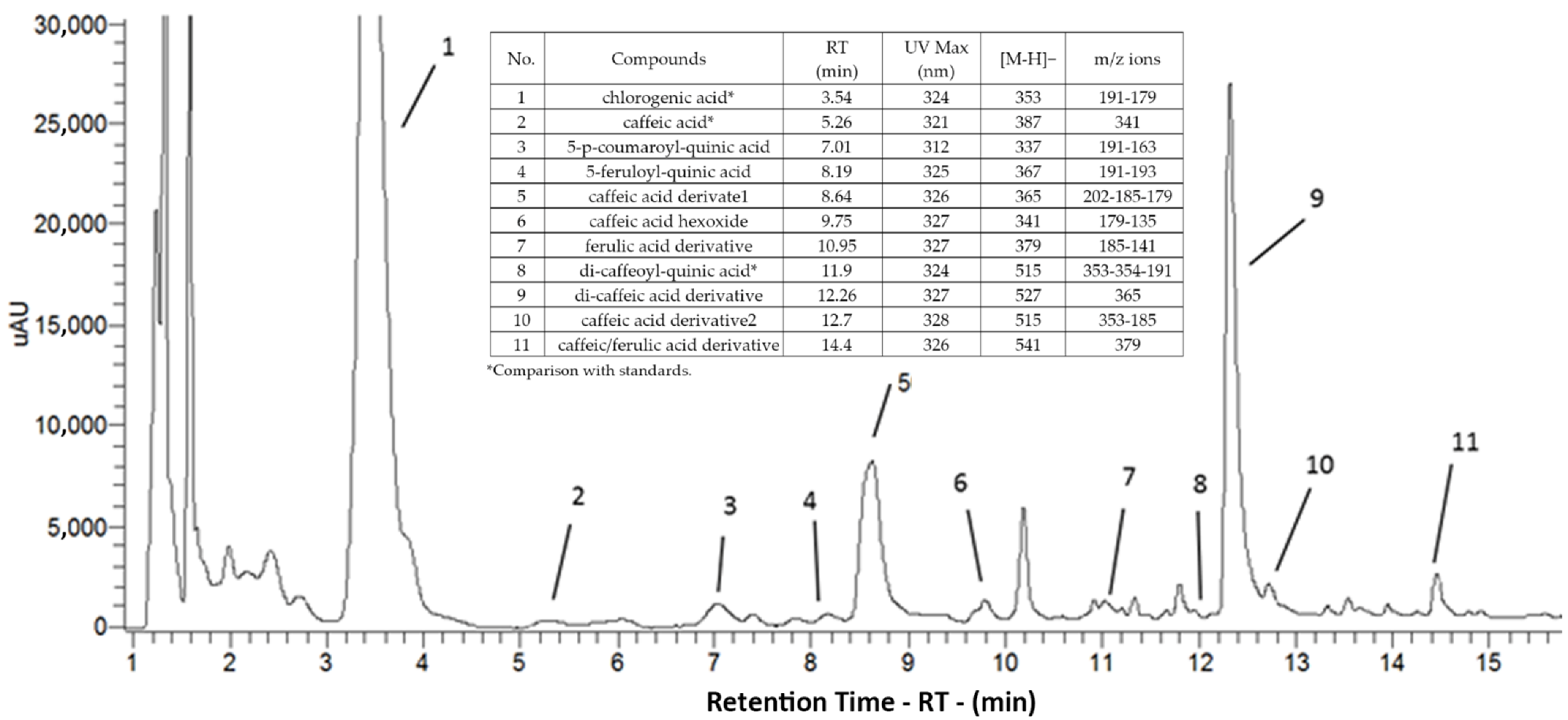
3.6. Phenols
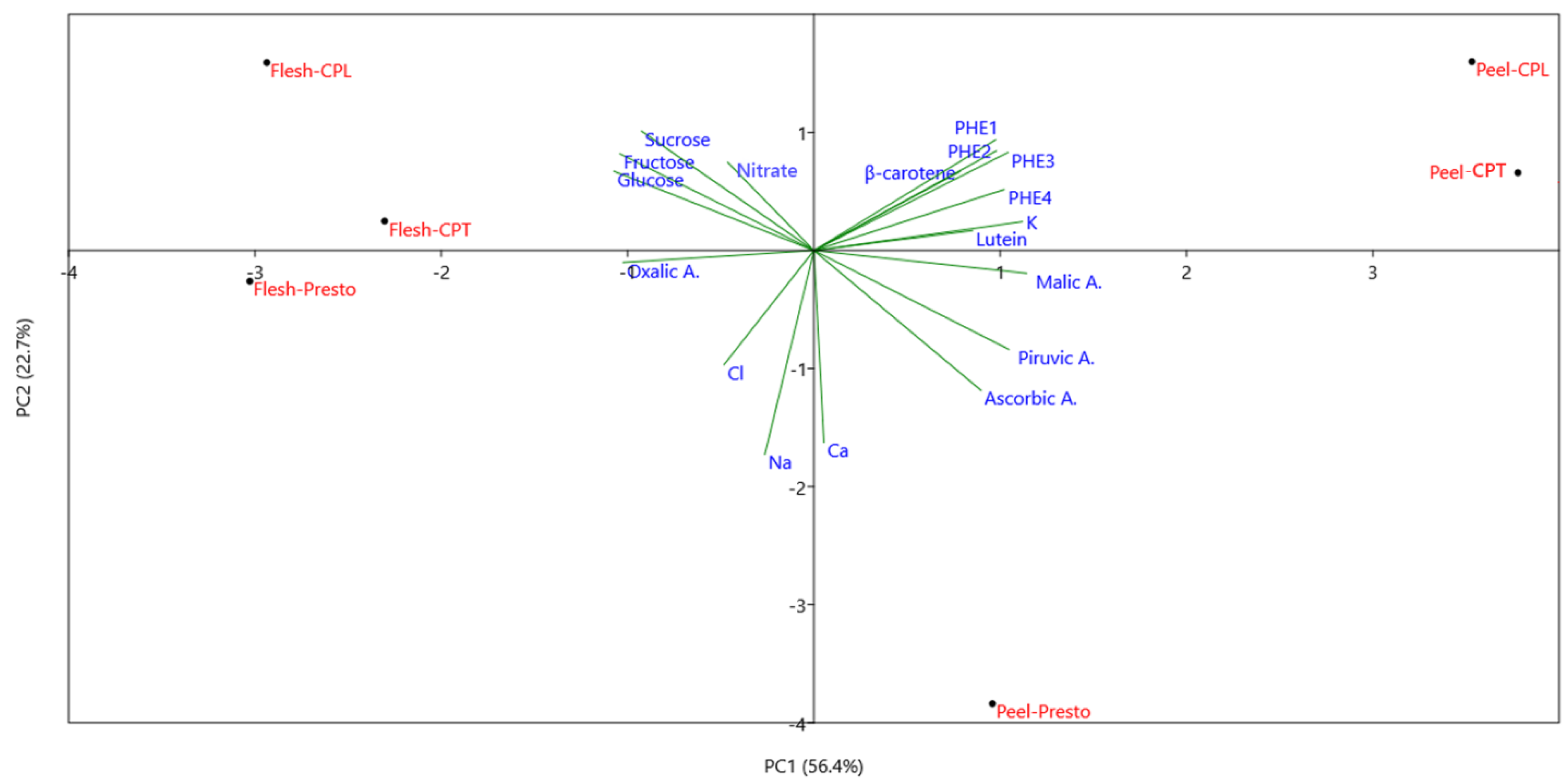
3.7. Principal Component Analysis and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Statistics Division Food and Agriculture Organization of the United Nations, Rome. Available online: https://www.fao.org/faostat/en/#data (accessed on 18 November 2021).
- Scarano, A.; Gerardi, C.; d’Amico, L.; Accogli, R.; Santino, A. Phytochemical analysis and antioxidant properties in colored Tiggiano carrots. Agriculture 2018, 8, 102. [Google Scholar] [CrossRef]
- Arscott, S.A.; Tanumihardjo, S.A. Carrots of many colors provide basic nutrition and bioavailable phytochemicals acting as a functional food. Compr. Rev. Food Sci. Food Saf. 2010, 9, 223–239. [Google Scholar] [CrossRef]
- Deding, U.; Baatrup, G.; Christensen, L.P.; Kobaek-Larsen, M. Carrot intake and risk of colorectal cancer: A prospective cohort study of 57,053 danes. Nutrients 2020, 12, 332. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Jiang, H.; Yang, W.; Song, F.; Yan, S.; Wang, C.; Fu, W.; Li, H.; Lyu, C.; Gan, Y.; et al. Is carrot consumption associated with a decreased risk of lung cancer? A meta-analysis of observational studies. Br. J. Nutr. 2019, 122, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Shao, F.; Zhang, F.; Miao, Q. Association between dietary carrot intake and breast cancer: A meta-analysis. Medicine 2018, 97, e12164. [Google Scholar] [CrossRef] [PubMed]
- Fallahzadeh, H.; Jalali, A.; Momayyezi, M.; Bazm, S. Effect of carrot intake in the prevention of gastric cancer: A meta-analysis. J. Gastric Cancer 2015, 15, 256–261. [Google Scholar] [CrossRef][Green Version]
- Xu, X.; Cheng, Y.; Li, S.; Zhu, Y.; Xu, X.; Zheng, X.; Mao, Q.; Xie, L. Dietary carrot consumption and the risk of prostate cancer. Eur. J. Nutr. 2014, 53, 1615–1623. [Google Scholar] [CrossRef]
- Ahmad, T.; Cawood, M.; Iqbal, Q.; Ariño, A.; Batool, A.; Sabir Tariq, R.M.; Azam, M.; Akhtar, S. Phytochemicals in Daucus carota and their health benefits—Review article. Foods 2019, 8, 424. [Google Scholar] [CrossRef]
- Hellström, J.; Granato, D.; Mattila, P.H. Accumulation of phenolic acids during storage over differently handled fresh carrots. Foods 2020, 9, 1515. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Hamauzu, Y. Phenolic compounds and their antioxidant properties in different tissues of carrots (Daucus carota L.). Int. J. Food Agric. Environ. 2004, 2, 332. [Google Scholar]
- Kenny, O.; O’Beirne, D. Antioxidant phytochemicals in fresh-cut carrot disks as affected by peeling method. Postharvest Biol. Technol. 2010, 58, 247–253. [Google Scholar] [CrossRef]
- Talcott, S.T.; Howard, L.R.; Brenes, C.H. Antioxidant changes and sensory properties of carrot puree processed with and without periderm tissue. J. Agric. Food Chem. 2000, 48, 1315–1321. [Google Scholar] [CrossRef] [PubMed]
- Seljåsen, R.; Kristensen, H.L.; Lauridsen, C.; Wyss, G.S.; Kretzschmar, U.; Birlouez-Aragone, I.; Kahl, J. Quality of carrots as affected by pre- and postharvest factors and processing. J. Sci. Food Agric. 2013, 93, 2611–2626. [Google Scholar] [CrossRef]
- Korolev, A.V.; Tomos, A.D.; Bowtell, R.; Farrar, J.F. Spatial and temporal distribution of solutes in the developing carrot taproot measured at single-cell resolution. J. Exp. Bot. 2000, 51, 567–577. [Google Scholar] [CrossRef][Green Version]
- ISTAT. Italian National Institute of Statistics, Rome. 2017. Available online: http://agri.istat.it/ (accessed on 20 October 2021).
- Conversa, G.; Lazzizera, C.; Bonasia, A.; Cifarelli, S.; Losavio, F.; Sonnante, G.; Elia, A. Exploring on-farm agro-biodiversity: A study case of vegetable landraces from Puglia region (Italy). Biodivers. Conserv. 2020, 29, 747–770. [Google Scholar] [CrossRef]
- Cefola, M.; Pace, B.; Renna, M.; Santamaria, P.; Signore, A.; Serio, F. Compositional analysis and antioxidant profile of yellow, orange and purple polignano carrots. Ital. J. Food Sci. 2012, 24, 284–291. [Google Scholar]
- Renna, M.; Serio, F.; Signore, A.; Santamaria, P. The yellow–purple Polignano carrot (Daucus carota L.): A multicoloured landrace from the Puglia region (Southern Italy) at risk of genetic erosion. Genet. Resour. Crop Evol. 2014, 61, 1611–1619. [Google Scholar] [CrossRef]
- Blando, F.; Marchello, S.; Maiorano, G.; Durante, M.; Signore, A.; Laus, M.N.; Soccio, M.; Mita, G. Bioactive compounds and antioxidant capacity in anthocyanin-rich carrots: A comparison between the black carrot and the Apulian landrace “Polignano” carrot. Plants 2021, 10, 564. [Google Scholar] [CrossRef] [PubMed]
- Negri, V.; Maxted, N.; Veteläinen, M. European Landrace Conservation: An Introduction. In European Landraces: On-Farm Conservation, Management and Use; Biodiversity Technical Bulletin No. 15; Veteläinen, V., Negri, N., Maxted, N., Eds.; Biodiversity International: Rome, Italy, 2009; pp. 1–22. [Google Scholar]
- Conversa, G.; Lazzizera, C.; Bonasia, A.; la Rotonda, P.; Elia, A. Nutritional characterization of two rare landraces of turnip (Brassica rapa var. rapa) tops and their on-farm conservation in Foggia province. Sustainability 2020, 12, 3842. [Google Scholar]
- Bonasia, A.; Conversa, G.; Lazzizera, C.; Loizzo, P.; Gambacorta, G.; Elia, A. Evaluation of garlic landraces from Foggia province (Puglia Region; Italy). Foods 2020, 9, 850. [Google Scholar] [CrossRef]
- Bonasia, A.; Conversa, G.; Lazzizera, C.; Gambacorta, G.; Elia, A. Morpho-biometrical, nutritional and phytochemical characterization of carrot landraces from Puglia region (Southern Italy). Sustainability 2021, 13, 3940. [Google Scholar] [CrossRef]
- Rohrer, J. Analysis of Carbohydrates by High-Performance Anion-Exchange Chromatography with Pulsed Amperometric Detection (HPAEPAD); Technical Note 20; Thermo Fisher Scientific: Sunnyvale, CA, USA, 2013. [Google Scholar]
- Magwaza, L.S.; Opara, U.L. Analytical methods for determination of sugars and sweetness of horticultural products—A review. Sci. Hortic. 2015, 184, 179–192. [Google Scholar] [CrossRef]
- Taungbodhitham, A.K.; Jones, G.P.; Wahlqvist, M.L.; Briggs, D.R. Evaluation of extraction method for the analysis of carotenoids in fruits and vegetables. Food Chem. 1998, 63, 577–584. [Google Scholar] [CrossRef]
- González-Castro, M.J.; Oruña-Concha, M.J.; López-Hernández, J.; Simal-Lozano, J. Effects of freezing on the organic acid content of frozen green beans and padrón peppers. Zeitschrift für Lebensmitteluntersuchung und-Forschung A 1997, 204, 365–368. [Google Scholar] [CrossRef]
- Pasqualone, A.; Punzi, R.; Trani, A.; Summo, C.; Paradiso, V.M.; Caponio, F.; Gambacorta, G. Enrichment of fresh pasta with antioxidant extracts obtained from artichoke canning by-products by ultrasound-assisted technology and quality characterisation of the end product. Int. J. Food Sci. Technol. 2017, 52, 2078–2087. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9–18. [Google Scholar]
- Nielsen, S.S. Food Analysis Laboratory Manual, 2nd ed.; Springer International Publishing: Cham, Switzerland, 2010. [Google Scholar]
- Nicolle, C.; Simon, G.; Rock, E.; Amouroux, P.; Rémésy, C. Genetic variability influences carotenoid, vitamin, phenolic, and mineral content in white, yellow, purple, orange, and dark-orange carrot cultivars. J. Am. Soc. Hortic. Sci. 2004, 129, 523–529. [Google Scholar] [CrossRef]
- Park, J.; Kwock, C.K.; Yang, Y.J. The Effect of the sodium to potassium ratio on hypertension prevalence: A propensity score matching approach. Nutrients 2016, 8, 482. [Google Scholar] [CrossRef] [PubMed]
- Sekara, A.; Pohl, A.; Kalisz, A.; Grabowska, A.; Cebula, S. Evaluation of selected Polish carrot cultivars for nutritive value and processing—A preliminary study. Ann. Warsaw Univ. Life Sci. SGGW. Hortic. Landsc. Archit. 2014, 35, 3–14. [Google Scholar]
- Kmecl, V.; Knap, T.; Žnidarčič, D. Evaluation of the nitrate and nitrite content of vegetables commonly grown in Slovenia. Ital. J. Agron. 2017, 12, 79–84. [Google Scholar] [CrossRef]
- Santamaria, P.; Elia, A.; Serio, F.; Todaro, E. A survey of nitrate and oxalate content in retail fresh vegetables. J. Sci. Food Agric. 1999, 79, 1882–1888. [Google Scholar] [CrossRef]
- Song, P.; Wu, L.; Guan, W. Dietary nitrates, nitrites, and nitrosamines intake and the risk of gastric cancer: A meta-analysis. Nutrients 2015, 7, 9872–9895. [Google Scholar] [CrossRef] [PubMed]
- Kreutzmann, S.; Christensen, L.P.; Edelenbos, M. Investigation of bitterness in carrots (Daucus carota L.) based on quantitative chemical and sensory analyses. LWT Food Sci. Technol. 2008, 41, 193–205. [Google Scholar] [CrossRef]
- Baranski, R.; Allender, C.; Klimek-Chodacka, M. Towards better tasting and more nutritious carrots: Carotenoid and sugar content variation in carrot genetic resources. Food Res. Int. 2012, 47, 182–187. [Google Scholar] [CrossRef]
- Benamor, J.; Mezghani, N.; Spooner, D.; Hamdi, K.; Ghezal, A.; Tarchoun, N. Assessment of phenotypic diversity in tunisian carrot (Daucus carota subsp. sativus) landraces. Int. J. Agric. Environ. Bioresearch 2019, 4, 75–91. [Google Scholar] [CrossRef]
- Šink, N.; Mikulic-Petkovsek, M.; Veberic, R.; Kacjan Marsic, N. Chemical composition and morphometric traits and yield of carrots grown in organic and integrated farming systems. Turkish J. Agric. For. 2017, 41, 452–462. [Google Scholar] [CrossRef]
- Priecina, L.; Karklina, D. Influence of steam treatment and drying on carrots composition and concentration of phenolics, organic acids and carotenoids. Proc. Latv. Acad. Sci. Sect. B Nat. Exact. Appl. Sci. 2018, 72, 103–112. [Google Scholar] [CrossRef]
- Yusuf, E.; Tkacz, K.; Turkiewicz, I.P.; Wojdyło, A.; Nowicka, P. Analysis of chemical compounds’ content in different varieties of carrots, including qualification and quantification of sugars, organic acids, minerals, and bioactive compounds by UPLC. Eur. Food Res. Technol. 2021, 247, 3053–3062. [Google Scholar] [CrossRef]
- Sinha, K.; Khare, V. Review on: Antinutritional factors in vegetable crops. Pharma Innov. J. 2017, 6, 353–358. [Google Scholar]
- Guil, J.L.; Torija, M.E.; Giménez, J.J.; Rodríguez-García, I.; Giménez, A. Oxalic acid and calcium determination in wild edible plants. J. Agric. Food Chem. 1996, 44, 1821–1823. [Google Scholar] [CrossRef]
- Tsuchida, H.; Cheng, D.H.; Inoue, K.; Kozukue, N.; Mizuno, S. Changes in pyruvic acid content and in GPT activity in chilling-sensitive and nonsensitive crops. HortScience 1990, 25, 952–953. [Google Scholar] [CrossRef]
- Phan, C.T.; Hsu, H. Physiological and chemical changes occurring in the carrot root during growth. Can. J. Plant Sci. 1973, 53, 629–634. [Google Scholar] [CrossRef]
- Abbey, L.; Ofoe, R.; Gunupuru, L.R.; Ijenyo, M. Variation in frequency of CQA-tested municipal solid waste compost can alter metabolites in vegetables. Food Res. Int. 2021, 143, 110225. [Google Scholar]
- Leahu, A.; Damian, C.; Carpiuc, N.; Oroian, M.; Avramiuc, M. Change in colour and physicochemical quality of carrot juice mixed with other fruits. J. Agroaliment. Process. Technol. 2013, 19, 241–246. [Google Scholar]
- Gutiérrez-Grijalva, E.P.; Ambriz-Pére, D.L.; Leyva-López, N.; Castillo-López, R.I.; Heredia, J.B. Review: Dietary phenolic compounds, health benefits and bioaccessibility. Arch. Latinoam. Nutr. 2016, 66, 87–100. [Google Scholar] [PubMed]
- Alarcón-Flores, M.I.; Romero-González, R.; Vidal, J.L.M.; González, F.J.E.; Frenich, A.G. Monitoring of phytochemicals in fresh and fresh-cut vegetables: A comparison. Food Chem. 2014, 142, 392–399. [Google Scholar] [CrossRef]
- Mattila, P.; Hellström, J. Phenolic acids in potatoes, vegetables, and some of their products. J. Food Compos. Anal. 2007, 20, 152–160. [Google Scholar] [CrossRef]
- Alasalvar, C.; Grigor, J.M.; Zhang, D.; Quantick, P.C.; Shahidi, F. Comparison of volatiles, phenolics, sugars, antioxidant vitamins, and sensory quality of different colored carrot varieties. J. Agric. Food Chem. 2001, 49, 1410–1416. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Genotype 1 | Leaves | Root | ||||||
|---|---|---|---|---|---|---|---|---|
| Fresh Weight (g) | Number (no.) | Fresh Weight (g) | Length (mm) | Equatorial Diameter (mm) | Dry Mass Concentration | Peel/Total Fresh Weight (%) | ||
| Root Flesh | Peel | |||||||
| (g kg−1 fw) | ||||||||
| CPL | 48.2 (4.9) a 2 | 8.2 (0.5) a | 248.8 (21.2) a | 231.1 (4.8) a | 38.7 (1.2) a | 102.8 (0.7) b | 92.8 (0.7) b | 9.8 (1.0) b |
| CPT | 17.4 (1.9) b | 6.5 (0.3) b | 110.4 (7.50) b | 182.7 (2.5) c | 30.3 (0.3) b | 93.9 (0.9) c | 81.6 (1.5) c | 14.6 (1.5) a |
| Presto | 11.7 (1.0) b | 5.7 (0.3) b | 135.4 (10.0) b | 210.8 (6.7) b | 31.4 (0.9) b | 117.1 (0.6) a | 103.3 (0.5) a | 12.1 (0.6) b |
| Significance 3 | *** | *** | *** | *** | *** | *** | *** | *** |
| Genotype 1 | Na | K | Mg | Ca | Total Cations | Na/K |
|---|---|---|---|---|---|---|
| Root flesh | ||||||
| CPL | 21.2 (0.9) b 2 | 94.6 (3.4) a | 2.1 (0.3) b | 7.3 (0.5) b | 125.2 (4.0) a | 0.2 (0.013) b |
| CPT | 18.3 (1.4) b | 59.2 (4.4) b | 1.2 (0.1) b | 7.4 (0.6) b | 86.1 (5.9) b | 0.3 (0.008) b |
| Presto | 34.5 (2.5) a | 40.3 (3.7) c | 8.6 (4.5) a | 14.6 (4.1) a | 98.0 (13.2) b | 0.9 (0.093) a |
| Significance 3 | *** | *** | * | * | ** | *** |
| Peel | ||||||
| CPL | 15.4 (0.8) b | 178.4 (5.2) a | 1.3 (0.1) b | 10.6 (0.8) b | 205.6 (5.6) a | 0.1 (0.006) b |
| CPT | 12.5 (1.3) b | 161.4 (10.4) b | 1.0 (0.4) b | 8.0 (0.7) c | 182.8(11.0) b | 0.1 (0.010) b |
| Presto | 54.2 (1.4) a | 123.6 (4.9) c | 7.7 (0.6) a | 19.3 (1.1) a | 204.9 (5.7) a | 0.4 (0.017) a |
| Significance 3 | *** | *** | *** | *** | ** | *** |
| Genotype 1 | Cl | NO3 | PO4 | SO4 | Total |
|---|---|---|---|---|---|
| Root flesh | |||||
| CPL | 42.0 (0.9) b 2 | 15.4 (0.5) a | 26.0 (0.8) a | 18.8 (0.6) b | 102.2 (1.8) a |
| CPT | 44.7 (0.8) a | 1.0 (0.5) b | 6.9 (0.6) c | 10.1 (0.3) c | 62.8 (1.3) c |
| Presto | 42.6 (1.0) b | 2.0 (0.6) b | 19.0 (1.7) b | 21.0 (0.3) a | 84.6 (2.2) b |
| Significance 3 | * | *** | *** | *** | *** |
| Peel | |||||
| CPL | 29.1 (0.8) b | 4.6 (0.3) a | 23.9 (0.6) a | 17.9 (0.4) b | 75.5 (0.8) c |
| CPT | 44.7 (0.9) a | 0.0 (0.0) c | 25.6 (1.1) a | 16.4 (0.4) c | 86.6 (1.7) b |
| Presto | 45.6 (1.2) a | 2.3 (0.6) b | 23.1 (0.7) a | 20.4 (0.4) a | 91.4 (2.0) a |
| Significance 3 | *** | *** | ns | *** | *** |
| Genotype 1 | Glucose | Fructose | Sucrose | Total Sugars | Sucrose/ Total Sugars | Sweetness Index | Lutein | ß-Carotene |
|---|---|---|---|---|---|---|---|---|
| (g 100 g−1 fw) | (µg 100 g−1 fw) | |||||||
| Root flesh | ||||||||
| CPL | 2.1 (0.06) a 2 | 2.1 (0.06) a | 3.9 (0.16) a | 8.1 (0.21) a | 0.48 (0.01) ab | 12.1 (0.31) a | 307 (46) a | 28,430 (1304) a |
| CPT | 1.8 (0.04) b | 1.8 (0.03) b | 3.1 (0.05) b | 6.8 (0.09) b | 0.46 (0.01) b | 10.2 (0.13) b | 122 (14) b | 12,336 (471) c |
| Presto | 2.1 (0.03) a | 2.0 (0.02) a | 4.0 (0.05) a | 8.0 (0.08) a | 0.49 (0.01) a | 12.0 (0.13) a | 270 (73) a | 18,368 (2348) b |
| Significance 3 | *** | *** | *** | *** | ** | *** | * | *** |
| Peel | ||||||||
| CPL | 1.5 (0.05) a | 1.5 (0.03) a | 2.4 (0.07) a | 5.4 (0.10) a | 0.45 (0.01) a | 8.2 (0.15) a | 349 (24) b | 31,849 (1060) a |
| CPT | 1.4 (0.07) b | 1.5 (0.04) a | 2.5 (0.05) a | 5.3 (0.13) a | 0.46 (0.01) a | 8.1 (0.19) a | 433 (32) a | 29,629 (1689) a |
| Presto | 1.4 (0.05) b | 1.4 (0.04) a | 1.7 (0.07) b | 4.6 (0.14) b | 0.37 (0.01) b | 7.0 (0.21) b | 312 (38) b | 22,682 (2150) b |
| Significance 3 | *** | ns | *** | *** | *** | *** | ** | *** |
| Genotype 1 | Oxalic Acid | Pyruvic Acid | Ascorbic Acid | Citric Acid | Malic Acid | Succinic Acid | Total | Oxalic Acid/Ca |
|---|---|---|---|---|---|---|---|---|
| Root flesh | ||||||||
| CPL | 3.8 (0.1) c 2 | 1.4 (0.1) a | 2.6 (0.1) c | 1.9 (0.3) a | 311.4 (7.5) a | 19.9 (1.3) a | 341.0 (8.5) a | 0.6 (0.1) b |
| CPT | 12.6 (1.3) b | 0.9 (0.1) b | 3.0 (0.1) b | 0.4 (0.1) a | 274.3 (20.8) b | 6.0 (0.2) b | 297.3 (13.8) b | 1.9 (0.2) a |
| Presto | 16.4 (0.3) a | 0.4 (0.0) c | 4.1 (0.1) a | 1.4 (0.5) a | 246.4 (11.2) c | 1.9 (0.2) c | 270.7(11.0) c | 1.9 (0.2) a |
| Significance 3 | *** | *** | *** | *** | *** | *** | *** | *** |
| Peel | ||||||||
| CPL | 10.4 (0.5) b | 15.3 (1.5) b | 6.8 (0.3) b | 1.1 (0.1) a | 531.6 (8.0) b | 9.9 (0.2) b | 575.1 (9.2) b | 1.1 (0.1) a |
| CPT | 3.5 (0.3) c | 18.2 (1.2) ab | 6.7 (0.2) b | 0.0 (0.0) b | 672.6 (25.1) a | 8.8 (0.5) b | 709.7 (25.0) a | 0.5 (0.1) b |
| Presto | 12.4 (0.2) a | 21.5 (1.4) a | 9.3 (0.2) a | 0.0 (0.0) b | 513.4 (9.8) b | 4.4 (0.1) c | 560.9 (10.0) b | 0.7 (0.1) b |
| Significance 3 | *** | * | *** | *** | *** | *** | *** | *** |
| Genotype 1 | Chlorogenic Acid 4 | Caffeic Acid 4 | 5-p-Coumaroyl-quinic Acid | 5-Feruloyl-quinic Acid | Caffeic Acid Derivate 1 | Caffeic Acid-o-hexoside | Ferulic Acid Derivative | Di-caffeoyl-quinic Acid 4 | Di-caffeic Acid Derivative | Caffeic Acid Derivative 2 | Caffeic/ferulic Acid Derivative | Sum of Un-identified Phenols | Total Phenols |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Root flesh | |||||||||||||
| CPL | 1.7 (1.1) b 2 | 0.002 (0.0003) b | 0.09 (0.03) b | 0.10 (0.04) b | 0.28 (0.22) a | 0.07 (0.02) a | 0.03 (0.02) a | 0.002 (0.001) a | 0.58 (0.46) a | 0.01 (0.001) a | 0.03 (0.02) a | 0.33 (0.12) a | 3.2 (1.9) b |
| CPT | 15.9 (1.3) a | 0.009 (0.0003) a | 3.10 (1.1) a | 0.84 (0.25) a | 0.90 (0.45) a | 0.07 (0.02) a | 0.11 (0.05) a | 0.007 (0.003) a | 1.34 (1.06) a | 0.02 (0.003) a | 0.06 (0.03) a | 0.21 (0.12) a | 22.6 (2.6) a |
| Presto | 9.1 (4.8) ab | 0.014 (0.002) a | 1.59 (0.50) ab | 0.71 (0.26) ab | 0.63 (0.35) a | 0.06 (0.01) a | 0.10 (0.03) a | 0.072 (0.011) a | 0.73 (0.64) a | 0.05 (0.01) a | 0.05 (0.02) a | 0.30 (0.13) a | 13.4 (6.3) ab |
| Significance 3 | * | ** | * | * | ns | ns | ns | ns | ns | ns | ns | ns | * |
| Peel | |||||||||||||
| CPL | 54.5 (8.5) a | 0.005 (0.002) a | 5.8 (1.0) a | 2.3 (0.5) a | 23.6 (5.1) a | 0.25 (0.03) a | 2.3 (0.3) a | 0.253 (0.107) a | 55.4 (13.1) a | 1.1 (0.6) a | 2.7 (0.7) a | 0.30 (0.05) a | 149.5 (28.4) a |
| CPT | 51.1 (7.2) a | 0.003 (0.0003) a | 7.8 (0.9) a | 3.2 (1.3) a | 17.1 (5.1) ab | 0.31 (0.05) a | 2.2 (1.3) a | 0.288 (0.114) a | 28.5 (9.0) ab | 0.5 (0.2) a | 2.0 (1.2) a | 0.18 (0.04) a | 113.2 (25.5) a |
| Presto | 0.7 (0.3) b | 0.004 (0.0003) a | 1.8 (0.2) b | 0.4 (0.09) a | 0.9 (0.4) b | 0.01 (0.001) b | 0.1 (0.05) a | 0.002 (0.001) a | 1.4 (0.7) b | 0.1 (0.02) a | 0.08 (0.03) a | 0.25 (0.03) a | 5.7 (1.4) b |
| Significance | * | ns | * | ns | * | *** | ns | ns | * | ns | ns | ns | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conversa, G.; Bonasia, A.; Natrella, G.; Lazzizera, C.; Elia, A. Peeling Affects the Nutritional Properties of Carrot Genotypes. Foods 2022, 11, 45. https://doi.org/10.3390/foods11010045
Conversa G, Bonasia A, Natrella G, Lazzizera C, Elia A. Peeling Affects the Nutritional Properties of Carrot Genotypes. Foods. 2022; 11(1):45. https://doi.org/10.3390/foods11010045
Chicago/Turabian StyleConversa, Giulia, Anna Bonasia, Giuseppe Natrella, Corrado Lazzizera, and Antonio Elia. 2022. "Peeling Affects the Nutritional Properties of Carrot Genotypes" Foods 11, no. 1: 45. https://doi.org/10.3390/foods11010045
APA StyleConversa, G., Bonasia, A., Natrella, G., Lazzizera, C., & Elia, A. (2022). Peeling Affects the Nutritional Properties of Carrot Genotypes. Foods, 11(1), 45. https://doi.org/10.3390/foods11010045