
Enzymatic Conversion of Mannan-Rich Plant Waste Biomass into Prebiotic Mannooligosaccharides




Abstract
1. Introduction
2. Mannan-Containing Waste Biomass Derived from Agro-Processing
3. Mannan
4. Endo-β-1,4-Mannanases
5. Enzymatic Production of MOS from Agro-Industrial Biomass
6. Identification and Quantification of Mannanase Produced MOS
7. MOS Generation Using Mannanases from Various GH Families
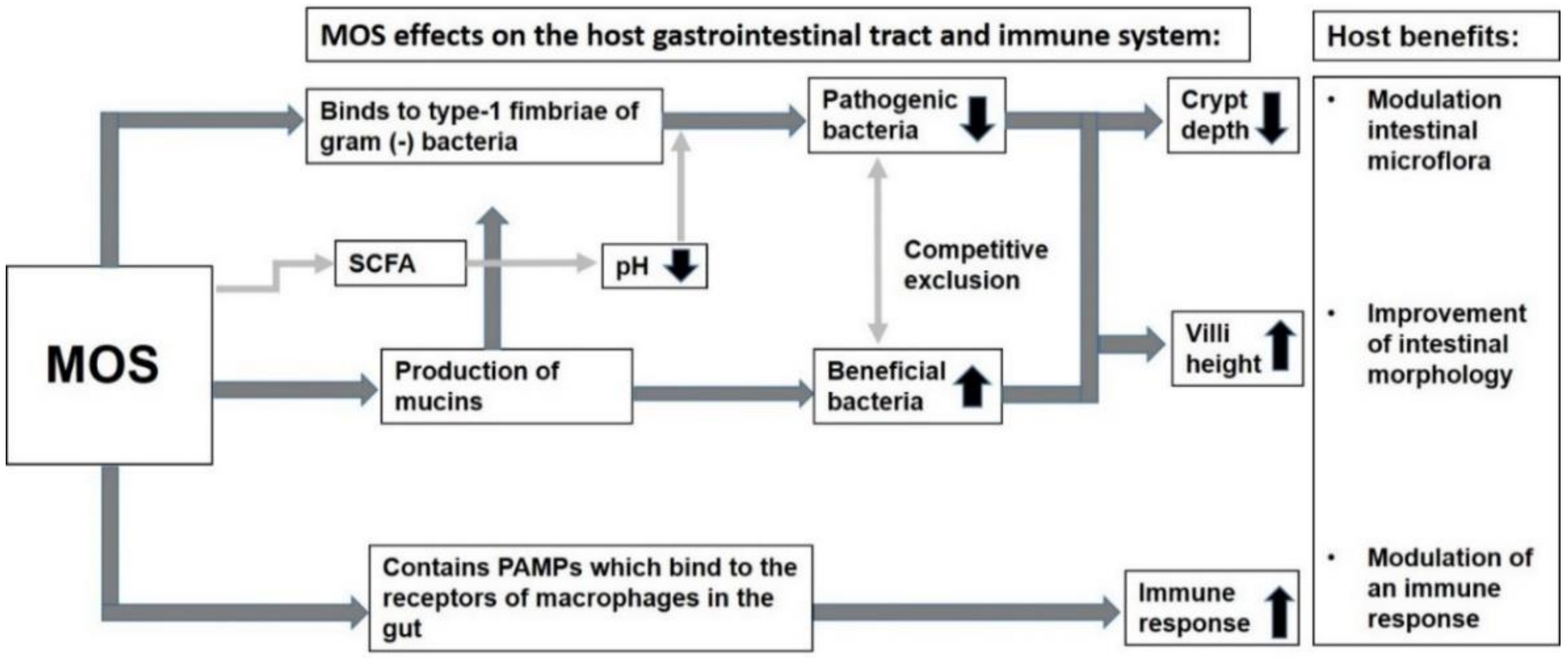
8. Properties and Biological Action of MOS
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Dyk, J.S.; Pletschke, B.I. A review of lignocellulose bioconversion using enzymatic hydrolysis and synergistic cooperation between enzymes-Factors affecting enzymes, conversion and synergy. Biotechnol. Adv. 2012, 30, 1458–1480. [Google Scholar] [CrossRef]
- Malgas, S.; Chandra, R.; Van Dyk, J.S.; Saddler, J.N.; Pletschke, B.I. Formulation of an optimized synergistic enzyme cocktail, HoloMix, for effective degradation of various pre-treated hardwoods. Bioresour. Technol. 2017, 245, 52–65. [Google Scholar] [CrossRef]
- Willför, S.; Sundberg, K.; Tenkanen, M.; Holmbom, B. Spruce-derived mannans—A potential raw material for hydrocolloids and novel advanced natural materials. Carbohydr. Polym. 2008, 72, 197–210. [Google Scholar] [CrossRef]
- Cerveró, J.M.; Skovgaard, P.A.; Felby, C.; Sørensen, H.R.; Jørgensen, H. Enzymatic hydrolysis and fermentation of palm kernel press cake for production of bioethanol. Enzym. Microb. Technol. 2010, 46, 177–184. [Google Scholar] [CrossRef]
- Jooste, T.; García-Aparicio, M.P.; Brienzo, M.; Van Zyl, W.H.; Görgens, J.F. Enzymatic hydrolysis of spent coffee ground. Appl. Biochem. Biotechnol. 2013, 169, 2248–2262. [Google Scholar] [CrossRef]
- Chapla, D.; Pandit, P.; Shah, A. Production of xylooligosaccharides from corncob xylan by fungal xylanase and their utilization by probiotics. Bioresour. Technol. 2012, 115, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Guarner, F.; Malagelada, J. Gut flora in health and disease. Lancet 2003, 360, 512–519. [Google Scholar] [CrossRef]
- Burokas, A.; Arboleya, S.; Moloney, R.D.; Peterson, V.L.; Murphy, K.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Targeting the microbiota-gut-brain Axis: Prebiotics have anxiolytic and antidepressant-like effects and reverse the impact of chronic stress in mice. Biol. Psychiatry 2016, 82, 472–487. [Google Scholar] [CrossRef]
- Bruce-keller, A.J.; Salbaum, J.M.; Berthoud, H. Harnessing Gut Microbes for Mental Health: Getting From Here to There. Intest. Microbiota Ment. Health 2018, 214–223. [Google Scholar] [CrossRef]
- Weina, L.; Yu, D.; Qian, C.; Peng, Z. Gut microbiome and cancer immunotherapy. Cancer Lett. 2019, 447, 41–47. [Google Scholar]
- Hekmatshoar, Y.; Rahbar Saadat, Y.; Hosseiniyan Khatibi, S.M.; Ozkan, T.; Zununi Vahed, F.; Nariman-Saleh-Fam, Z.; Pourghassem Gargari, B.; Sunguroglu, A.; Zununi Vahed, S. The impact of tumor and gut microbiotas on cancer therapy: Beneficial or detrimental? Life Sci. 2019, 233, 116680. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, A.; Butt, M.S.; Afzaal, M.; van Baak, M.; Nadeem, M.T.; Shahid, M.Z. Prebiotics, gut microbiota and metabolic risks: Unveiling the relationship. J. Funct. Foods 2015, 17, 189–201. [Google Scholar] [CrossRef]
- Vandenplas, Y. Probiotics and prebiotics in infectious gastroenteritis. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Charoensiddhi, S.; Conlon, M.A.; Franco, C.M.M.; Zhang, W. The development of seaweed-derived bioactive compounds for use as prebiotics and nutraceuticals using enzyme technologies. Trends Food Sci. Technol. 2017, 70, 20–33. [Google Scholar] [CrossRef]
- Hutkins, R.W.; Krumbeck, J.A.; Bindels, L.B.; Cani, P.D.; Fahey, G.; Goh, Y.J.; Hamaker, B.; Martens, E.C.; Mills, D.A.; Rastal, R.A.; et al. Prebiotics: Why definitions matter. Curr. Opin. Biotechnol. 2016, 37, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Moreno, F.J.; Corzo, N.; Montilla, A.; Villamiel, M.; Olano, A. Current state and latest advances in the concept, production and functionality of prebiotic oligosaccharides. Curr. Opin. Food Sci. 2017, 13, 50–55. [Google Scholar] [CrossRef]
- Saravanan, P.; Muthuvelayudham, R.; Viruthagiri, T. Enhanced production of cellulase from pineapple waste by responses surface methodology. J. Eng. 2013, 2013. [Google Scholar] [CrossRef]
- Daud, Z.; Hatta, M.Z.M.; Kassim, A.S.M.; Awang, H.; Aripin, A.M. Exploring of agro waste (pineapple leaf, corn stalk, and napier grass) by chemical composition and morphological study. BioResources 2014, 9, 872–880. [Google Scholar] [CrossRef]
- Tanaka, K.; Hilary, Z.D.; Ishizaki, A. Investigation of the utility of pineapple juice and pineapple waste material as low-cost substrate for ethanol fermentation by Zymomonas mobilis. J. Biosci. Bioeng. 1999, 87, 642–646. [Google Scholar] [CrossRef]
- Olver, B.; Dyk, J.S.; Beukes, N.; Pletschke, B.I. Synergy between EngE, XynA and ManA from Clostridium cellulovorans on corn stalk, grass and pineapple pulp substrates. 3 Biotech 2011, 1, 187–192. [Google Scholar] [CrossRef]
- Malgas, S.; Pletschke, B.I. Combination of CTec2 and GH5 or GH26 endo-mannanases for effective lignocellulosic biomass degradation. Catalysts 2020, 10, 1193. [Google Scholar] [CrossRef]
- Omwango, E.; Njagi, E.N.; Orinda, G.; Wanjau, R. Nutrient enrichment of pineapple waste using Aspergillus niger and Trichoderma viride by solid state fermentation. Afr. J. Biotechnol. 2013, 12, 6193–6196. [Google Scholar] [CrossRef]
- Sukruansuwan, V.; Napathorn, S.C. Use of agro-industrial residue from the canned pineapple industry for polyhydroxybutyrate production by Cupriavidus necator strain A-04. Biotechnol. Biofuels 2018, 11, 202. [Google Scholar] [CrossRef]
- Aruna, T.E. Production of value-added product from pineapple peels using solid state fermentation. Innov. Food Sci. Emerg. Technol. 2019, 57, 102193. [Google Scholar] [CrossRef]
- Clauser, N.M.; Gutiérrez, S.; Area, M.C.; Felissia, F.E.; Vallejos, M.E. Techno-economic assessment of carboxylic acids, furfural, and pellet production in a pine sawdust biorefinery. Biofuels Bioprod. Biorefin. 2018, 12, 997–1012. [Google Scholar] [CrossRef]
- Andrew, J.; Johakimu, J.; Lekha, P.; Gibril, M.; Sitholé, B. Beneficiation of sawdust waste in the context of an integrated forest biorefinery mill: Kraft and prehydrolysis kraft Pulping properties. In Opportunities for Biomass and Organic Waste Valorisation; Godfrey, L., Görgens, J.F., Roman, H., Eds.; Taylor & Francis: Oxfordshire, UK, 2020; pp. 123–139. [Google Scholar]
- Ferreira-Santos, P.; Zanuso, E.; Genisheva, Z.; Rocha, C.M.R.; Teixeira, J.A. Green and sustainable valorization of bioactive phenolic compounds from pinus by-products. Molecules 2020, 25, 2931. [Google Scholar] [CrossRef]
- Rodríguez, E.R.; Martín, J.D.; Romero, C.D. Aloe vera as a functional ingredient in foods. Crit. Rev. Food Sci. Nutr. 2010, 50, 305–326. [Google Scholar] [CrossRef] [PubMed]
- Tai-Nin Chow, J.; Williamson, D.A.; Yates, K.M.; Goux, W.J. Chemical characterization of the immunomodulating polysaccharide of Aloe vera L. Carbohydr. Res. 2005, 340, 1131–1142. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-González, V.M.; Femenia, A.; González-Laredo, R.F.; Rocha-Guzmán, N.E.; Gallegos-Infante, J.A.; Candelas-Cadillo, M.G.; Ramírez-Baca, P.; Simal, S.; Rosselló, C. Effects of pasteurization on bioactive polysaccharide acemannan and cell wall polymers from Aloe barbadensis Miller. Carbohydr. Polym. 2011, 86, 1675–1683. [Google Scholar] [CrossRef]
- Chang, X.L.; Chen, B.Y.; Feng, Y.M. Water-soluble polysaccharides isolated from skin juice, gel juice and flower of Aloe vera Miller. J. Taiwan Inst. Chem. Eng. 2011, 42, 197–203. [Google Scholar] [CrossRef]
- Minjares-Fuentes, R.; Femenia, A.; Comas-Serra, F.; Rodríguez-González, V.M. Compositional and structural features of the main bioactive polysaccharides present in the aloe vera plant. J. AOAC Int. 2018, 101, 1711–1719. [Google Scholar] [CrossRef]
- Simoes, J.; Nunes, F.M.; Coimbra, M.A.; Domingues, M.R. Mass spectrometry characterization of an Aloe vera mannan presenting immunostimulatory activity. Carbohydr. Res. 2012, 90, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Wongsiridetchai, C.; Chiangkham, W.; Khlaihiran, N.; Sawangwan, T.; Wongwathanarat, P.; Charoenrat, T.; Chantorn, S. Alkaline pretreatment of spent coffee grounds for oligosaccharides production by mannanase from Bacillus sp. GA2(1). Agric. Nat. Resour. 2018, 52, 222–227. [Google Scholar] [CrossRef]
- Ballesteros, L.F.; Cerqueira, M.A.; Teixeira, J.A.; Mussatto, S.I. Characterization of polysaccharides extracted from spent coffee grounds by alkali pretreatment. Carbohydr. Polym. 2015, 127, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.A.; Cho, E.J.; Lee, D.S.; Bae, H.J. Development of an advanced integrative process to create valuable biosugars including manno-oligosaccharides and mannose from spent coffee grounds. Bioresour. Technol. 2019, 272, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Simões, J.; Maricato, É.; Nunes, F.M.; Domingues, M.R.; Coimbra, M.A. Thermal stability of spent coffee ground polysaccharides: Galactomannans and arabinogalactans. Carbohydr. Polym. 2014, 101, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Favaro, C.P.; Baraldi, I.J.; Casciatori, F.P. β-Mannanase production using coffee industry waste for application in soluble coffee processing. Biomolecules 2020, 10, 227. [Google Scholar] [CrossRef]
- Asano, I.; Umemura, M.; Fujii, S.; Hoshino, H.; Iino, H. Effects of mannooligosaccharides from coffee mannan on fecal microflora and defecation in healthy volunteers. Food Sci. Technol. Res. 2004, 10, 93–97. [Google Scholar] [CrossRef]
- Gu, J.; Pei, W.; Tang, S.; Yan, F.; Peng, Z.; Huang, C.; Yang, J.; Yong, Q. Procuring biologically active galactomannans from spent coffee ground (SCG) by autohydrolysis and enzymatic hydrolysis. Int. J. Biol. Macromol. 2020, 149, 572–580. [Google Scholar] [CrossRef]
- Abdeltawab, A.M.; Khattab, M.S.A. Utilization of palm kernel cake as a ruminant feed for animal: A review. Asian J. Biol. Sci. 2018, 11, 157–164. [Google Scholar] [CrossRef]
- Sharmila, A.; Alimon, A.R.; Azhar, K.; Noor, H. Improving nutritional values of palm kernel cake (PKC) as poultry feeds: A review. Malays. J. Anim. Sci. 2014, 17, 1–18. [Google Scholar]
- Aguzey, H.A.; Gao, Z.H.; Wu, H.H.; Cheng, G.L.; Wu, Z.M.; Chen, J.H.; Niu, Z.L. Enzymatic treatment of palm kernel cake improves intestinal health, gut microbiota and immune system of broilers. J. Anim. Plant Sci. 2020, 30, 517–526. [Google Scholar] [CrossRef]
- Ariandi; Yopi; Meryandini, A. Enzymatic hydrolysis of copra meal by mannanase from Streptomyces sp. BF3.1 for the production of mannooligosaccharides. HAYATI J. Biosci. 2015, 22, 79–86. [Google Scholar] [CrossRef]
- Kangas, H.; Felissia, F.E.; Filgueira, D.; Ehman, N.V.; Vallejos, M.E.; Imlauer, C.M.; Lahtinen, P.; Area, M.C.; Chinga-Carrasco, A.G. 3D printing high-consistency enzymatic nanocellulose obtained from a soda-ethanol-O2 pine sawdust pulp. Bioengineering 2019, 6, 60. [Google Scholar] [CrossRef] [PubMed]
- Rättö, M.; Siika-aho, M.; Buchert, J.; Valkeajävi, A.; Viikari, L. Enzymatic hydrolosis of isolated and fibre-bound galactoglucomannans from pine-wood and pine kraft pulp. Appl. Microbiol. Biotechnol. 1993, 40, 449–454. [Google Scholar] [CrossRef]
- Samanta, A.K.; Jayapal, N.; Jayaram, C.; Roy, S.; Kolte, A.P.; Senani, S.; Sridhar, M. Xylooligosaccharides as prebiotics from agricultural by-products: Production and applications. Bioact. Carbohydr. Diet. Fibre 2015, 5, 62–71. [Google Scholar] [CrossRef]
- Malgas, S.; van Dyk, J.S.; Pletschke, B.I. A review of the enzymatic hydrolysis of mannans and synergistic interactions between β-mannanase, β-mannosidase and α-galactosidase. World J. Microbiol. Biotechnol. 2015, 31, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, G.; Arya, S.K. Mannans: An overview of properties and application in food products. Int. J. Biol. Macromol. 2018, 119, 79–95. [Google Scholar] [CrossRef]
- Dhawan, S.; Kaur, J. Microbial mannanases: An overview of production and applications. Crit. Rev. Biotechnol. 2007, 27, 197–216. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.R.S.; Filho, E.X.F. An overview of mannan structure and mannan-degrading enzyme systems. Appl. Microbiol. Biotechnol. 2008, 79, 165–178. [Google Scholar] [CrossRef]
- Malgas, S.; van Dyk, S.J.; Pletschke, B.I. β-Mannanase (Man26A) and α-galactosidase (Aga27A) synergism—A key factor for the hydrolysis of galactomannan substrates. Enzym. Microb. Technol. 2015, 70, 1–8. [Google Scholar] [CrossRef]
- Tailford, L.E.; Ducros, V.M.A.; Flint, J.E.; Roberts, S.M.; Morland, C.; Zechel, D.L.; Smith, N.; Bjørnvad, M.E.; Borchert, T.V.; Wilson, K.S.; et al. Understanding how diverse β-mannanases recognize heterogeneous substrates. Biochemistry 2009, 48, 7009–7018. [Google Scholar] [CrossRef] [PubMed]
- Shallom, D.; Shoham, Y. Microbial hemicellulases. Curr. Opin. Microbiol. 2003, 6, 219–228. [Google Scholar] [CrossRef]
- Shoseyov, O.; Shani, Z.; Levy, I. Carbohydrate binding modules: Biochemical properties and novel applications. Microbiol. Mol. Biol. Rev. 2006, 70, 283–295. [Google Scholar] [CrossRef]
- Van Zyl, W.H.; Rose, S.H.; Trollope, K.; Görgens, J.F. Fungal β-mannanases: Mannan hydrolysis, heterologous production and biotechnological applications. Process Biochem. 2010, 45, 1203–1213. [Google Scholar] [CrossRef]
- Ghosh, A.; Luís, A.S.; Brás, J.L.A.; Fontes, C.M.G.A.; Goyal, A. Thermostable recombinant β-(1→4)-mannanase from C. thermocellum: Biochemical characterization and manno-oligosaccharides production. J. Agric. Food Chem. 2013, 61, 12333–12344. [Google Scholar] [CrossRef] [PubMed]
- Yamabhai, M.; Sak-Ubol, S.; Srila, W.; Haltrich, D. Mannan biotechnology: From biofuels to health. Crit. Rev. Biotechnol. 2016, 36, 30–42. [Google Scholar] [CrossRef]
- Suzuki, K.; Michikawa, M.; Sato, H.; Yuki, M.; Kamino, K.; Ogasawara, W.; Fushinobu, S.; Kaneko, S. Purification, cloning, functional expression, structure, and characterization of a thermostable β-mannanase from Talaromyces trachyspermus B168 and its efficiency in production of mannooligosaccharides from coffee wastes. J. Appl. Glycosci. 2018, 65, 13–21. [Google Scholar] [CrossRef]
- Kumar Suryawanshi, R.; Kango, N. Production of mannooligosaccharides from various mannans and evaluation of their prebiotic potential. Food Chem. 2021, 334, 127428. [Google Scholar] [CrossRef]
- Li, Y.; Yi, P.; Liu, J.; Yan, Q.; Jiang, Z. High-level expression of an engineered β-mannanase (mRmMan5A) in Pichia pastoris for manno-oligosaccharide production using steam explosion pretreated palm kernel cake. Bioresour. Technol. 2018, 256, 30–37. [Google Scholar] [CrossRef]
- Bååth, J.A.; Abad, A.M.; Berglund, J.; Larsbrink, J.; Vilaplana, F.; Olsson, L. Mannanase hydrolysis of Spruce galactoglucomannan focusing on the influence of acetylation on enzymatic mannan degradation. Biotechnol. Biofuels 2018, 11, 114. [Google Scholar] [CrossRef]
- Patel, S.; Goyal, A. Functional oligosaccharides: Production, properties and applications. World J. Microbiol. Biotechnol. 2011, 27, 1119–1128. [Google Scholar] [CrossRef]
- Nimptsch, K.; Süß, R.; Riemer, T.; Nimptsch, A.; Schnabelrauch, M.; Schiller, J. Differently complex oligosaccharides can be easily identified by matrix-assisted laser desorption and ionization time-of-flight mass spectrometry directly from a standard thin-layer chromatography plate. J. Chromatogr. A 2010, 1217, 3711–3715. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, T.; Bortolini, O.; Massi, A.; Sacchetti, G.; Tacchini, M.; De Risi, C. Exploring the synergy between HPTLC and HPLC-DAD for the investigation of wine-making by-products. Molecules 2019, 24, 3416. [Google Scholar] [CrossRef] [PubMed]
- Starr, C.M.; Masada, R.I.; Hague, C.; Skop, E.; Klock, J.C. Fluorophore-assisted carbohydrate electrophoresis in the separation, analysis, and sequencing of carbohydrates. J. Chromatogr. A 1996, 720, 295–321. [Google Scholar] [CrossRef]
- O’Shea, M.G.; Samuel, M.S.; Konik, C.M.; Morell, M.K. Fluorophore-assisted carbohydrate electrophoresis (FACE) of oligosaccharides: Efficiency of labelling and high-resolution separation. Carbohydr. Res. 1998, 307, 1–12. [Google Scholar] [CrossRef]
- Goins, T.L.; Cutler, J.I.M.E. Relative abundance of oligosaccharides in Candida species as determined by fluorophore-assisted carbohydrate electrophoresis. J. Clin. Microbiol. 2000, 38, 2862–2869. [Google Scholar] [CrossRef]
- Srivastava, P.K.; Panwar, D.; Prashanth, K.V.H.; Kapoor, M. Structural characterization and in vitro fermentation of β-mannooligosaccharides produced from locust bean gum by GH-26 endo-β-1,4-mannanase (ManB-1601). J. Agric. Food Chem. 2017, 65, 2827–2838. [Google Scholar] [CrossRef]
- Kalidas, N.R.; Saminathan, M.; Ismail, I.S.; Abas, F.; Maity, P.; Islam, S.S.; Manshoor, N.; Shaari, K. Structural characterization and evaluation of prebiotic activity of oil palm kernel cake mannanoligosaccharides. Food Chem. 2017, 234, 348–355. [Google Scholar] [CrossRef]
- Morrill, J.; Kulcinskaja, E.; Sulewska, A.M.; Lahtinen, S.; Stålbrand, H.; Svensson, B.; Abou Hachem, M. The GH5 1,4-β-mannanase from Bifidobacterium animalis subsp. lactis Bl-04 possesses a low-affinity mannan-binding module and highlights the diversity of mannanolytic enzymes Protein and enzyme biochemistry. BMC Biochem. 2015, 16, 26. [Google Scholar] [CrossRef]
- Couturier, M.; Roussel, A.; Rosengren, A.; Leone, P.; Stålbrand, H.; Berrin, J.G. Structural and biochemical analyses of glycoside hydrolase families 5 and 26 β-(1,4)-mannanases from Podospora anserina reveal differences upon manno-oligosaccharide catalysis. J. Biol. Chem. 2013, 288, 14624–14635. [Google Scholar] [CrossRef]
- Liu, Z.; Ning, C.; Yuan, M.; Yang, S.; Wei, X.; Xiao, M.; Fu, X.; Zhu, C.; Mou, H. High-level expression of a thermophilic and acidophilic β-mannanase from Aspergillus kawachii IFO 4308 with significant potential in mannooligosaccharide preparation. Bioresour. Technol. 2020, 295, 122257. [Google Scholar] [CrossRef]
- Aguilera-Gálvez, C.; Vásquez-Ospina, J.J.; Gutiérrez-Sanchez, P.; Acuña-Zornosa, R. Cloning and biochemical characterization of an endo-1,4-β-mannanase from the coffee berry borer Hypothenemus hampei. BMC Res. Notes 2013, 6, 333. [Google Scholar] [CrossRef]
- Zang, H.; Xie, S.; Wu, H.; Wang, W.; Shao, X.; Wu, L.; Rajer, F.U.; Gao, X. A novel thermostable GH5_7 β-mannanase from Bacillus pumilus GBSW19 and its application in manno-oligosaccharides (MOS) production. Enzym. Microb. Technol. 2015, 78, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Harnpicharnchai, P.; Pinngoen, W.; Teanngam, W.; Sornlake, W.; Sae-Tang, K.; Manitchotpisit, P.; Tanapongpipat, S. Production of high activity Aspergillus niger BCC4525 β-mannanase in Pichia pastoris and its application for mannooligosaccharides production from biomass hydrolysis. Biosci. Biotechnol. Biochem. 2016, 80, 2298–2305. [Google Scholar] [CrossRef] [PubMed]
- Arunrattanamook, N.; Wansuksri, R.; Uengwetwanit, T.; Champreda, V. Engineering of β-mannanase from Aspergillus niger to increase product selectivity towards medium chain length mannooligosaccharides. J. Biosci. Bioeng. 2020, 130, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, P.S.; Sharma, P.; Puri, N.; Gupta, N. Purification and characterization of an alkali-thermostable β-mannanase from Bacillus nealsonii PN-11 and its application in mannooligosaccharides preparation having prebiotic potential. Eur. Food Res. Technol. 2014, 238, 927–936. [Google Scholar] [CrossRef]
- Chauhan, P.S.; Sharma, P.; Puri, N.; Gupta, N. A process for reduction in viscosity of coffee extract by enzymatic hydrolysis of mannan. Bioprocess Biosyst. Eng. 2014, 37, 1459–1467. [Google Scholar] [CrossRef]
- Zhang, R.; Li, X.Y.; Cen, X.L.; Gao, Q.H.; Zhang, M.; Li, K.Y.; Wu, Q.; Mu, Y.L.; Tang, X.H.; Zhou, J.P.; et al. Enzymatic preparation of manno-oligosaccharides from locust bean gum and palm kernel cake, and investigations into its prebiotic activity. Electron. J. Biotechnol. 2021, 49, 64–71. [Google Scholar] [CrossRef]
- Bågenholm, V.; Wiemann, M.; Reddy, S.K.; Bhattacharya, A.; Logan, D.T.; Stålbrand, H. A surface exposed GH26 β-mannanase from Bacteroides ovatus: Structure, role and phylogenetic analysis of BoMan26B. J. Biol. Chem. 2019, 294, 9100–9117. [Google Scholar] [CrossRef]
- Piwpankaew, Y.; Sakulsirirat, S.; Nitisinprasert, S.; Nguyen, T.H.; Haltrich, D.; Keawsompong, S. Cloning, secretory expression and characterization of recombinant β-mannanase from Bacillus circulans NT 6.7. Springerplus 2014, 3, 430. [Google Scholar] [CrossRef][Green Version]
- Bågenholm, V.; Reddy, S.K.; Bouraoui, H.; Morrill, J.; Kulcinskaja, E.; Bahr, C.M.; Aurelius, O.; Rogers, T.; Xiao, Y.; Logan, D.T.; et al. Galactomannan catabolism conferred by a polysaccharide utilisation locus of Bacteroides ovatus: Enzyme synergy and crystal structure of a β-mannanase. J. Biol. Chem. 2017, 292, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Cartmell, A.; Topakas, E.; Ducros, V.M.A.; Suits, M.D.L.; Davies, G.J.; Gilbert, H.J. The Cellvibrio japonicus mannanase CjMan26C displays a unique exo-mode of action that is conferred by subtle changes to the distal region of the active site. J. Biol. Chem. 2008, 283, 34403–34413. [Google Scholar] [CrossRef]
- Tsukagoshi, H.; Nakamura, A.; Ishida, T.; Otagiri, M.; Moriya, S.; Samejima, M.; Igarashi, K.; Kitamoto, K.; Arioka, M. The GH26 β-mannanase RsMan26H from a symbiotic protist of the termite Reticulitermes speratus is an endo-processive mannobiohydrolase: Heterologous expression and characterization. Biochem. Biophys. Res. Commun. 2014, 452, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ju, J.; Peng, H.; Gao, F.; Zhou, C.; Zeng, Y.; Xue, Y.; Li, Y.; Henrissat, B.; Gao, G.F.; et al. Biochemical and structural characterization of the intracellular mannanase AaManA of Alicyclobacillus acidocaldarius reveals a novel glycoside hydrolase family belonging to clan GH-A. J. Biol. Chem. 2008, 283, 31551–31558. [Google Scholar] [CrossRef]
- Liu, W.; Ma, C.; Liu, W.; Zheng, Y.; Chen, C.-C.; Liang, A.; Luo, X.; Li, Z.; Ma, W.; Song, Y.; et al. Functional and structural investigation of a novel β-mannanase BaMan113A from Bacillus sp. N16-5. Int. J. Biol. Macromol. 2021, 182, 899–909. [Google Scholar] [CrossRef]
- Shimizu, M.; Kaneko, Y.; Ishihara, S.; Mochizuki, M.; Sakai, K.; Yamada, M.; Murata, S.; Itoh, E.; Yamamoto, T.; Sugimura, Y.; et al. Novel β-1,4-mannanase belonging to a new glycoside hydrolase family in Aspergillus nidulans. J. Biol. Chem. 2015, 290, 27914–27927. [Google Scholar] [CrossRef]
- Jana, U.K.; Suryawanshi, R.K.; Prajapati, B.P.; Soni, H.; Kango, N. Production optimization and characterization of mannooligosaccharide generating β-mannanase from Aspergillus oryzae. Bioresour. Technol. 2018, 268, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Jana, U.K.; Kango, N. Characteristics and bioactive properties of mannooligosaccharides derived from agro-waste mannans. Int. J. Biol. Macromol. 2020, 149, 931–940. [Google Scholar] [CrossRef]
- Purnawan, A.; Yopi, Y.; Irawadi, T.T. Production of manooligomannan from palm kernel cake by mannanase produced from Streptomyces cyaenus. Biosaintifika J. Biol. Biol. Educ. 2017, 9, 73–80. [Google Scholar] [CrossRef]
- Pangestu, R.; Rahmani, N.; Palar, R.; Lisdiyanti, P. Yopi The effect of biomass particle size and chemical structure on the enzymatic hydrolysis reaction of galactomannan from sugar palm fruit by β-mannanase from Kitasatospora sp. KY576672. IOP Conf. Ser. Earth Environ. Sci. 2019, 251, 012008. [Google Scholar] [CrossRef]
- Yopi; Rahmani, N.; Amanah, S.; Santoso, P.; Lisdiyanti, P. The production of β-mannanase from Kitasatospora sp. strain using submerged fermentation: Purification, characterization and its potential in mannooligosaccharides production. Biocatal. Agric. Biotechnol. 2020, 24, 101532. [Google Scholar] [CrossRef]
- Bangoria, P.; Divecha, J.; Shah, A. Production of mannooligosaccharides producing b-mannanase by newly isolated Penicillium aculeatum APS1 using oil seed residues under solid state fermentation. Biocatal. Agric. Biotechnol. 2021, 34, 102023. [Google Scholar] [CrossRef]
- Chacher, M.F.A.; Kamran, Z.; Ahsan, U.; Ahmad, S.; Koutoulis, K.C.; Qutab Ud DIn, H.G.; Cengiz, O. Use of mannan oligosaccharide in broiler diets: An overview of underlying mechanisms. Worlds. Poult. Sci. J. 2017, 73, 831–844. [Google Scholar] [CrossRef]
- Baurhoo, B.; Letellier, A.; Zhao, X.; Ruiz-Feria, C. Cecal populations of Lactobacilli and Bifidobacteria and Escherichia coli populations after in vivo Escherichia coli challenge in birds fed diets with purified lignin or mannanoligoccharides. Poult. Sci. 2007, 86, 2509–2516. [Google Scholar]
- Bland, E.J.; Keshavarz, T.; Bucke, C. The influence of small oligosaccharides on the immune system. Carbohydr. Res. 2004, 339, 1673–1678. [Google Scholar] [CrossRef]
- Saeed, M.; Ahmad, F.; Arain, M.A.; Abd El-Hack, M.E.; Emam, M.; Bhutto, Z.A.; Moshaveri, A. Use of mannan-oligosaccharides (MOS) as a feed additive in poultry nutrition. J. World’s Poult. Res. 2017, 7, 94–103. [Google Scholar]
- Turner, M.W. The role of mannose-binding lectin in health and disease. Mol. Immunol. 2003, 40, 423–429. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef]
- Ismail, S.A.; Hassan, A.A.; Emran, M.A. Economic production of thermo-active endo β-mannanase for the removal of food stain and production of antioxidant manno-oligosaccharides. Biocatal. Agric. Biotechnol. 2019, 22, 101387. [Google Scholar] [CrossRef]
- Thambiraj, S.R.; Phillips, M.; Koyyalamudi, S.R.; Reddy, N. Yellow lupin (Lupinus luteus L.) polysaccharides: Antioxidant, immunomodulatory and prebiotic activities and their structural characterisation. Food Chem. 2018, 267, 319–328. [Google Scholar] [CrossRef]
- Aliakbarlu, J.; Mohammadi, S.; Khalili, S. A study on antioxidant potency and antibacterial activity of water extracts of some spices widely consumed in Iranian diet. J. Food Biochem. 2014, 38, 159–166. [Google Scholar] [CrossRef]
- Pereira, D.I.A.; Gibson, G.R. Effects of consumption of probiotics and prebiotics on serum lipid levels in humans. Crit. Rev. Biochem. Mol. Biol. 2002, 37, 259–281. [Google Scholar] [CrossRef] [PubMed]
- Van Loo, J.; Cummings, J.; Delzenne, N.; Englyst, H.; Franck, A.; Hopkins, M.; Kok, N.; Macfarlane, G.; Newton, D.; Quigley, M.; et al. Functional food properties of non-digestible oligosaccharides: A consensus report from the ENDO project (DGXII AIRII-CT94-1095). Br. J. Nutr. 1999, 81, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Lim, W.; Kim, S.; Khil, H.; Cheon, E.; An, S.; Hong, S.; Lee, D.H.; Kang, S.; Oh, H.; et al. Coffee Intake and Obesity: A Meta-Analysis. Nutrients 2019, 11, 1274. [Google Scholar] [CrossRef] [PubMed]
- Maki, C.; Funakoshi-Tago, M.; Aoyagi, R.; Ueda, F.; Kimura, M.; Kobata, K.; Tago, K.; Tamura, H. Coffee extract inhibits adipogenesis in 3T3-L1 preadipocyes by interrupting insulin signaling through the downregulation of IRS1. PLoS ONE 2017, 12, e0173264. [Google Scholar] [CrossRef]
- Van Dam, R.M.; Feskens, E.J.M. Coffee consumption and risk of type 2 diabetes mellitus. Lancet 2002, 360, 1477–1478. [Google Scholar] [CrossRef]
- Ghosh, A.; Verma, A.K.; Tingirikari, J.R.; Shukla, R.; Goyal, A. Recovery and purification of oligosaccharides from copra meal by recombinant endo-β-mannanase and deciphering molecular mechanism involved and its role as potent therapeutic agent. Mol. Biotechnol. 2014, 57, 111–127. [Google Scholar] [CrossRef]
- Li, T.; Yan, Q.; Wen, Y.; Liu, J.; Sun, J.; Jiang, Z. Synbiotic yogurt containing konjac mannan oligosaccharides and Bifidobacterium animalis ssp. lactis BB12 alleviates constipation in mice by modulating the stem cell factor (SCF)/c-Kit pathway and gut microbiota. J. Dairy Sci. 2021, 104, 5239–5255. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Biomass Waste | Mannan Composition (%), Dry Mass Basis | Type of Mannan | References |
|---|---|---|---|
| SCG | 20 | Galactomannan | [5,34,40] |
| PKC | 60 | Linear mannan | [43] |
| CM | 61 | Linear mannan | [44] |
| PP | 1.5 | Galactomannan | [21] |
| PSD (softwoods) | 12 | Acetyl-galactoglucomannan | [45,46] |
| A. vera | 50 | Acetyl-galactomannan | [32] |
| Enzyme Name and Source | Mannan Substrate | Parameters (pH, °C, Hours) | MOS Produced | Analytical Technique | References |
|---|---|---|---|---|---|
| GH5 mannanases | |||||
| CjMan5A, Cellvibrio japonicas | GGM KG LBG | 7, 35, 24 h | GGM: M1–M4 KG: M1–M4 LBG: M1–M3 | HPAEC-PAD: Carbopac PA200 column | [62] |
| PaMan5A, Podospora anserine | INM KG LBG | 5.2, 40, 0.5 h | M1–M3 | HPAEC-PAD: Carbo-PacPA-1 column | [72] |
| ManAK, Aspergillus kawachii IFO 4308 | LBG KG GG | 3.0, 60, 12 h | LBG: M1–M4, M6 KG: M1–M6 GG: M1–M4 | TLC, HPLC: TSKgel PWXL column | [73] |
| HhMan, Hypothenemis hampei | LBG GG | 5.5, 30, 24 h | M1–M4 | TLC | [74] |
| BlMan5_8, Bifidobacterium animalis | INM LBG | 6.0, 37, 0.0083 h (30 s) | INM: M3–M5 LBG: M2–M5 | HPAEC-PAD: CarboPac PA200 column | [71] |
| mRmMan5A, Rhizomucor miechei | PKC | 4.5, 50, 8 h | M1–M4 | HPLC-ELSD, Sugar KS-802 column | [61] |
| TtMan5A, Talaromyces trachyspermus B168 | DCW | 4.5, 50, 69 h | M1–M4 | HPAEC-PAD DX-500 | [59] |
| Bpman5, Bacillus pumilus CBSW19 | LBG | 6.5, 50, 24 h | M1–M3, M5, M6 | TLC, HPLC: C18 reverse phase column | [75] |
| ManF3, Aspergillus niger BCC4525 | CM PKC | 5.5, 50, 24 h | CM: M1–M5 PKC: M1–M3 | TLC | [76] |
| Mannan | 5.5, 50, 0.17–3 h | 0.17 h: M2–M5 3 h: M2–M7 | HPAEC-PAD | [77] | |
| ManPN11, Bacillus nealsonii PN-11 | LBG | 8.8, 55, 6 h | M1–M5, >M5 | HPAEC: CarboPac PA100 column | [78] |
| ManPN11, Bacillus nealsonii PN-11 | CB | 8.8, 37, 24 h | M1–M5 | TLC, HPLC: Aminex HPX-87P column | [79] |
| Man5HJ14, Bacillus sp. HJ14 | LBG PKC | 6.5, 60, 10 h | LBG: M1–M7 PKC: M1–M4 | TLC, ESI-MS | [80] |
| GH26 mannanases | |||||
| PaMan26A, Podospora anserine | INM KG LBG | 5.2, 40, 0.5 h | M1–M4 | HPAEC-PAD: Carbo-PacPA-1 column | [72] |
| BoMan26B, Bacteroides ovatus | GG LBG | 6.5, 37, 24 h | GG: M1–M6 LBG: M1–M6 | HPAEC-PAD: CarboPac PA200 and PA20 columns | [81] |
| CjMan26A, Cellvibrio japonicas | GGM KG LBG | 7.0, 37, 24 h | GGM: M1–M4 KG: M1–M3 LBG: M1, M2 | HPAEC-PAD: CarboPac PA200 column | [62] |
| Mannanase, Bacillus circulans | LBG | 6.0, 50, 24 h | M2–M6 | HPLC: Aminex-HPX42C column | [82] |
| BoMan26B, Bacteroides ovatus | GG INM LBG | 6.5, 37, 24 h | GG: M2–M6, >M6 INM: M2 LBG: M2–M6, >M6 | HPAEC-PAD: CarboPac PA200 column | [83] |
| ManAJB13, Sphingomonas sp. JB13 | LBG PKC | 6.5, 37, 10 h | LBG: M1–M6 PKC: M1–M4 | TLC, ESI-MS | [80] |
| GH26 mannobiohydrolases | |||||
| BoMan26A, Bacteroides ovatus | GG INM LBG | 6.5, 37, 24 h | GG: M2, GM3, M5 INM: M2 LBG: M2, GM3 | HPAEC-PAD: CarboPac PA200 column | [83] |
| CjMan26C, Cellvibrio japonicas | INM LBG | 37, 7.0, 0.5 h | INM: M2 LBG: M2, GM2 | HPLC: CarboPac PA100 column | [84] |
| RSMan26H, Reticulitermes speratus | INM | 5.5, 30, 1 h | M2–M5 | HPLC: Shodex Asahipak NH2P-50 4E column | [85] |
| GH113 mannanases | |||||
| AaManA, Alicyclobacillus acidocaldarius | KG LBG | 5.5, 65, 3 h | KG: M1–M6 LBG: M1–M6, >M6 | TLC | [86] |
| BaMan113A, Bacillus sp. N16- 5 | KG LBG | 7.0, 30, 2 h | KG: M1 LBG: G, GM, GM2, M1, M2, M4 | TLC | [87] |
| GH134 mannanases | |||||
| AnMan134A, Aspergillus nidulans | INM KG LBG | 6.0, 37, 0.25 h | INM: M2–M4 KG: M2–M6, >M6 LBG: M2–M6, >M6 | TLC, MALDI-TOF-MS, HPLC: Shim-pack ISA-07/S2504 column | [88] |
| Unidentified mannanases | |||||
| Mannanase, Bacillus sp. GA2(1) | SCG | 6.0, 50, 5 h | M2, M3 | TLC | [34] |
| ManAo, Aspergillus oryzae | GG KG LBG | 5.0, 60, 24 h | GG: M1–M4, >M4 KG: M1, M2 LBG: M1–M3, >M4 | FACE | [89] |
| ManAo, Aspergillus oryzae | CM GG KG LBG PKC | 5.0, 50, 12 h | CM: M1–M3 GG: M1–M3 KG: M1–M4 LBG: M1–M3 PKC: M1–M3 | HPLC: Sugar-Pak column, 2D NMR | [90] |
| rHhMan, Hypothenemus hampei | GG LBG | 5.5, 30, 24 h | M1–M4 | TLC | [74] |
| Mannanase, Streptomyces cyaenus | PKC | 6.0, 37, 8 h | M2–M6 | TLC, HPLC: Zorbax carbohydrate column | [91] |
| Mannanase, Kitasatospora sp. KY576672 | SPF | 6.0, 40, 32 h | M1–M6 | TLC, HPLC: Hi-PlexCa (Duo) column | [92] |
| Mannanase, Kitasatospora sp. | INM LBG PP SPF | 6.0, 40, 2–6 h | INM: M1–M5 LBG: M1–M6, >M6 PP: M1–M6, >M6 SPF: M1, M2, >M6 | TLC | [93] |
| Mannanase, Penicillium aculeatum AP S1 | GG KG LBG | 5.3, 50, 3 h | GG: M1–M3, >M4 KG & LBG: M1–M4, >M4 | TLC, HPLC: Hamilton RCX-30 column | [94] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hlalukana, N.; Magengelele, M.; Malgas, S.; Pletschke, B.I. Enzymatic Conversion of Mannan-Rich Plant Waste Biomass into Prebiotic Mannooligosaccharides. Foods 2021, 10, 2010. https://doi.org/10.3390/foods10092010
Hlalukana N, Magengelele M, Malgas S, Pletschke BI. Enzymatic Conversion of Mannan-Rich Plant Waste Biomass into Prebiotic Mannooligosaccharides. Foods. 2021; 10(9):2010. https://doi.org/10.3390/foods10092010
Chicago/Turabian StyleHlalukana, Nosipho, Mihle Magengelele, Samkelo Malgas, and Brett Ivan Pletschke. 2021. "Enzymatic Conversion of Mannan-Rich Plant Waste Biomass into Prebiotic Mannooligosaccharides" Foods 10, no. 9: 2010. https://doi.org/10.3390/foods10092010
APA StyleHlalukana, N., Magengelele, M., Malgas, S., & Pletschke, B. I. (2021). Enzymatic Conversion of Mannan-Rich Plant Waste Biomass into Prebiotic Mannooligosaccharides. Foods, 10(9), 2010. https://doi.org/10.3390/foods10092010