
Plant Proteins for Future Foods: A Roadmap


Abstract
1. Introduction
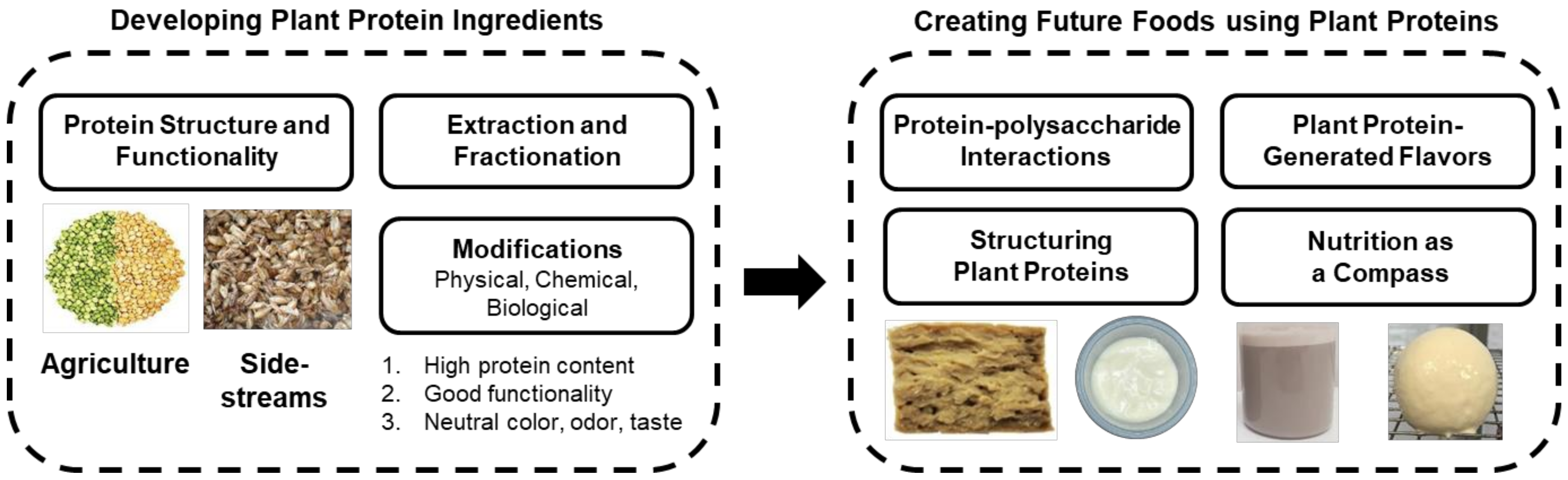
2. Developing Plant Protein Ingredients
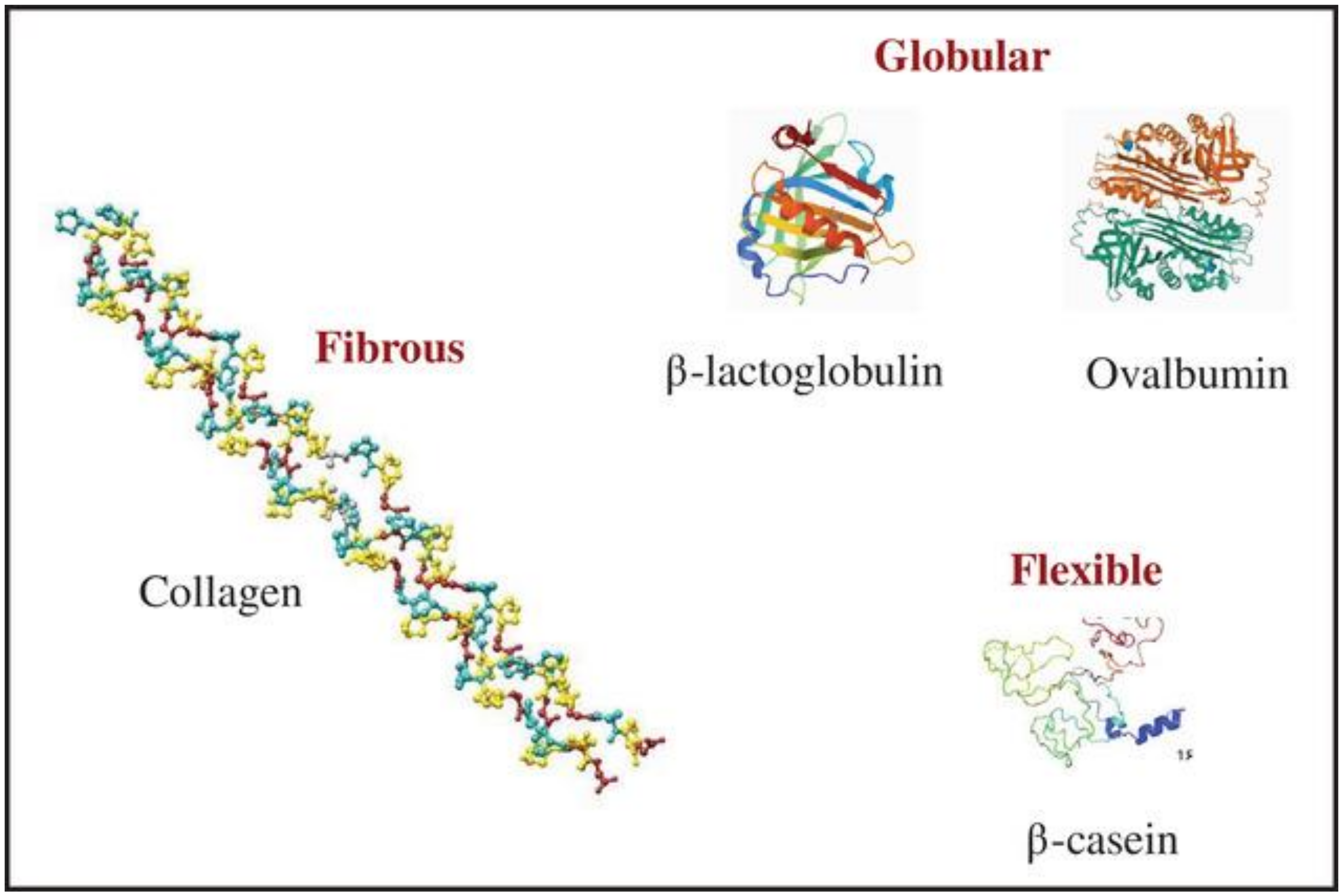
2.1. Protein Structure and Functionality
2.1.1. Globular Proteins
2.1.2. Fibrous Proteins
2.1.3. Flexible Proteins
2.2. Plant Protein Extraction and Fractionation
2.2.1. Plant Protein Extraction
2.2.2. Plant Protein Fractionation
2.3. Modification of Plant Proteins
2.3.1. Physical Modifications
High-Pressure Processing
High-Pressure Homogenization
Extrusion
Sonication
2.3.2. Chemical Modifications
Glycation
pH Shifting
2.3.3. Biological Modifications
3. Creating Future Foods Using Plant Proteins
3.1. Protein–Polysaccharide Interactions
3.2. Structuring Plant Proteins
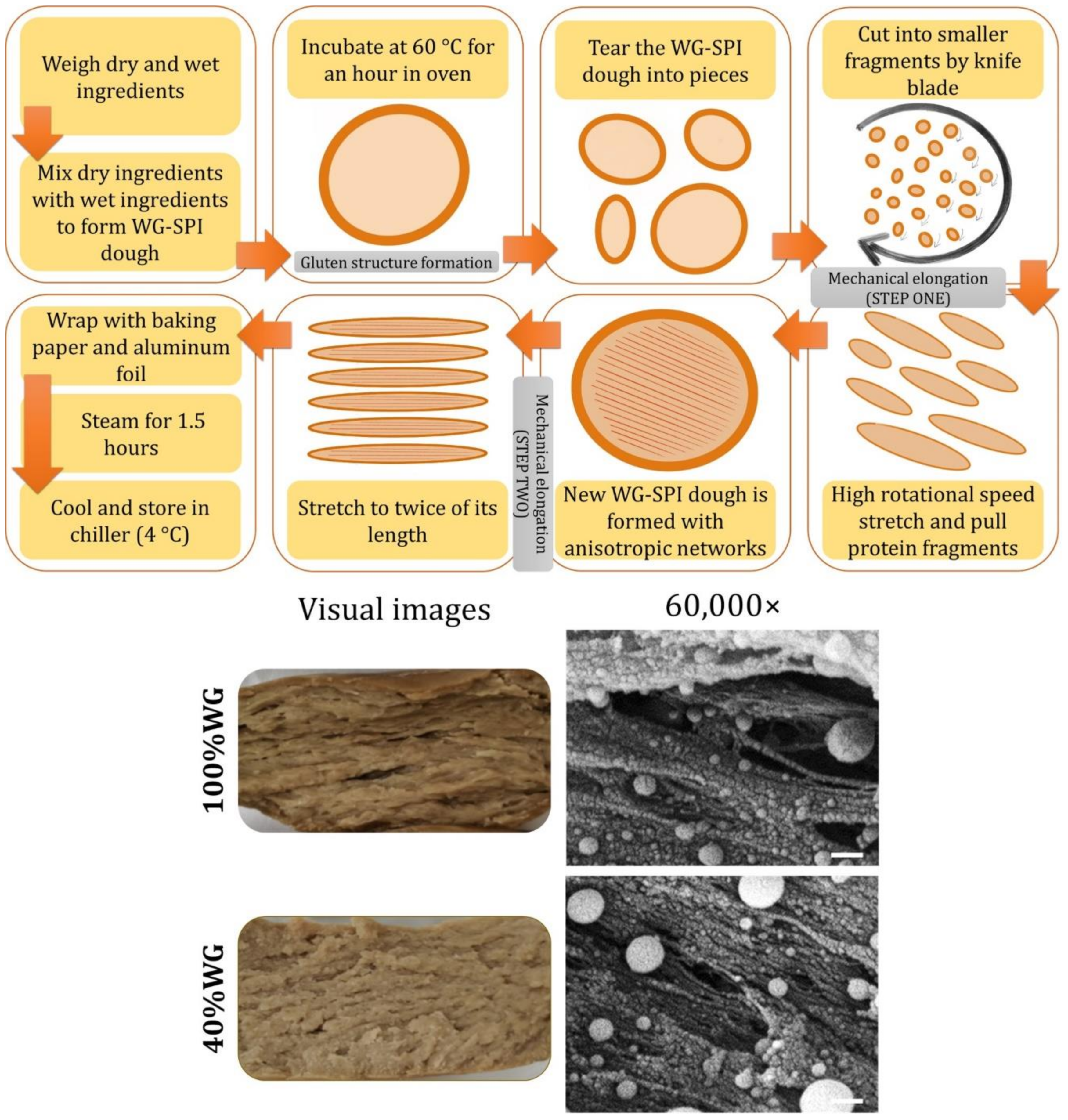
3.2.1. Formation of Fibrous Structures
3.2.2. Formation of Gels
3.3. Flavors Generated from Plant Proteins
3.4. Nutrition as a Compass
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- U.N. Growing at a Slower Pace, World Population is Expected to Reach 9.7 Billion in 2050 and Could Peak at Nearly 11 Billion around 2100: UN Report. Available online: https://www.un.org/sustainabledevelopment/blog/2019/06/growing-at-a-slower-pace-world-population-is-expected-to-reach-9-7-billion-in-2050-and-could-peak-at-nearly-11-billion-around-2100-un-report/ (accessed on 22 July 2021).
- Calicioglu, O.; Flammini, A.; Bracco, S.; Bellù, L.; Sims, R. The Future Challenges of Food and Agriculture: An Integrated Analysis of Trends and Solutions. Sustainability 2019, 11, 222. [Google Scholar] [CrossRef]
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT–Lancet Commission on healthy diets from sustainable food systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef]
- McClements, D.J.; Grossmann, L. The science of plant-based foods: Constructing next-generation meat, fish, milk, and egg analogs. Compr. Rev. Food Sci. Food Saf. 2021, 20, 4049–4100. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Munoz, L.M.; Tavares, G.M.; Corredig, M. Design future foods using plant protein blends for best nutritional and technological functionality. Trends Food Sci. Technol. 2021, 113, 139–150. [Google Scholar] [CrossRef]
- Schweiggert-Weisz, U.; Eisner, P.; Bader-Mittermaier, S.; Osen, R. Food proteins from plants and fungi. Curr. Opin. Food Sci. 2020, 32, 156–162. [Google Scholar] [CrossRef]
- Loveday, S.M. Plant protein ingredients with food functionality potential. Nutr. Bull. 2020, 45, 321–327. [Google Scholar] [CrossRef]
- Akharume, F.U.; Aluko, R.E.; Adedeji, A.A. Modification of plant proteins for improved functionality: A review. Compr. Rev. Food Sci. Food Saf. 2021, 20, 198–224. [Google Scholar] [CrossRef] [PubMed]
- Nikbakht Nasrabadi, M.; Sedaghat Doost, A.; Mezzenga, R. Modification approaches of plant-based proteins to improve their techno-functionality and use in food products. Food Hydrocoll. 2021, 118, 106789. [Google Scholar] [CrossRef]
- World Health Organization. The State of Food Security and Nutrition in the World 2020: Transforming Food Systems for Affordable Healthy Diets; Food and Agriculture Organization: Rome, Italy, 2020; Volume 2020. [Google Scholar]
- Arif, M.; Pauls, K.P. Properties of plant proteins. In Plant Bioproducts; Chen, G., Weselake, R.J., Singer, S.D., Eds.; Springer: New York, NY, USA, 2018; pp. 121–142. [Google Scholar] [CrossRef]
- Ismail, B.P.; Senaratne-Lenagala, L.; Stube, A.; Brackenridge, A. Protein demand: Review of plant and animal proteins used in alternative protein product development and production. Anim. Front. 2020, 10, 53–63. [Google Scholar] [CrossRef]
- Abd El-Salam, M.H.; El-Shibiny, S.; Salem, A. Factors affecting the functional properties of whey protein products: A review. Food Rev. Int. 2009, 25, 251–270. [Google Scholar] [CrossRef]
- O’Sullivan, J.; Murray, B.; Flynn, C.; Norton, I. The effect of ultrasound treatment on the structural, physical and emulsifying properties of animal and vegetable proteins. Food Hydrocoll. 2016, 53, 141–154. [Google Scholar] [CrossRef]
- Wilding, P.; Lillford, P.J.; Regenstein, J.M. Functional properties of proteins in foods. J. Chem. Technol. Biotechnol. Biotechnol. 1984, 34, 182–189. [Google Scholar] [CrossRef]
- Großmann, L.; Weiss, J. Alternative protein sources as technofunctional food ingredients. Annu. Rev. Food Sci. Technol. 2021, 12, 93–117. [Google Scholar] [CrossRef] [PubMed]
- Boye, J.; Zare, F.; Pletch, A. Pulse proteins: Processing, characterization, functional properties and applications in food and feed. Food Res. Int. 2010, 43, 414–431. [Google Scholar] [CrossRef]
- Veraverbeke, W.S.; Delcour, J.A. Wheat protein composition and properties of wheat glutenin in relation to breadmaking functionality. Crit. Rev. Food Sci. Nutr. 2002, 42, 179–208. [Google Scholar] [CrossRef]
- Delcour, J.A.; Joye, I.J.; Pareyt, B.; Wilderjans, E.; Brijs, K.; Lagrain, B. Wheat gluten functionality as a quality determinant in cereal-based food products. Annu. Rev. Food Sci. Technol. 2012, 3, 469–492. [Google Scholar] [CrossRef] [PubMed]
- González-Pérez, S.; Arellano, J.B. 15—Vegetable protein isolates. In Handbook of Hydrocolloids, 2nd ed.; Phillips, G.O., Williams, P.A., Eds.; Woodhead Publishing: Sawston, UK, 2009; pp. 383–419. [Google Scholar] [CrossRef]
- Molina, M.I.; Petruccelli, S.; Añón, M.C. Effect of pH and ionic strength modifications on thermal denaturation of the 11S globulin of sunflower (Helianthus annuus). J. Agric. Food Chem. 2004, 52, 6023–6029. [Google Scholar] [CrossRef] [PubMed]
- Lakemond, C.M.M.; de Jongh, H.H.J.; Hessing, M.; Gruppen, H.; Voragen, A.G.J. Soy Glycinin: Influence of pH and Ionic Strength on Solubility and Molecular Structure at Ambient Temperatures. J. Agric. Food Chem. 2000, 48, 1985–1990. [Google Scholar] [CrossRef]
- Mundi, S.; Aluko, R.E. Physicochemical and functional properties of kidney bean albumin and globulin protein fractions. Food Res. Int. 2012, 48, 299–306. [Google Scholar] [CrossRef]
- Pérez, S.G.; Vereijken, J.M.; van Koningsveld, G.A.; Gruppen, H.; Voragen, A.G.J. Physicochemical Properties of 2S Albumins and the Corresponding Protein Isolate from Sunflower (Helianthus annuus). J. Food Sci. 2005, 70, C98–C103. [Google Scholar] [CrossRef]
- Nwachukwu, I.D.; Aluko, R.E. Chapter 1 food protein structures, functionality and product development. In Food Proteins and Peptides: Emerging Biofunctions, Food and Biomaterial Applications; The Royal Society of Chemistry: London, UK, 2021; pp. 1–33. [Google Scholar] [CrossRef]
- Nwachukwu, I.D.; Aluko, R.E. Physicochemical and emulsification properties of flaxseed (Linum usitatissimum) albumin and globulin fractions. Food Chem. 2018, 255, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Kimura, A.; Fukuda, T.; Zhang, M.; Motoyama, S.; Maruyama, N.; Utsumi, S. Comparison of physicochemical properties of 7S and 11S globulins from pea, fava bean, cowpea, and French bean with those of soybean-French bean 7S globulin exhibits excellent properties. J. Agric. Food Chem. 2008, 56, 10273–10279. [Google Scholar] [CrossRef]
- Majzoobi, M.; Abedi, E.; Farahnaky, A.; Aminlari, M. Functional properties of acetylated glutenin and gliadin at varying pH values. Food Chem. 2012, 133, 1402–1407. [Google Scholar] [CrossRef]
- Lu, B.-Y.; Quillien, L.; Popineau, Y. Foaming and emulsifying properties of pea albumin fractions and partial characterisation of surface-active components. J. Sci. Food Agric. 2000, 80, 1964–1972. [Google Scholar] [CrossRef]
- Papalamprou, E.M.; Doxastakis, G.I.; Kiosseoglou, V. Chickpea protein isolates obtained by wet extraction as emulsifying agents. J. Sci. Food Agric. 2010, 90, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Nishinari, K.; Fang, Y.; Nagano, T.; Guo, S.; Wang, R. Soy as a food ingredient. In Proteins in Food Processing; Elsevier: Amsterdam, The Netherlands, 2018; pp. 149–186. [Google Scholar]
- Ruíz-Henestrosa, V.P.; Sánchez, C.C.; Escobar, M.d.M.Y.; Jiménez, J.J.P.; Rodríguez, F.M.; Patino, J.M.R. Interfacial and foaming characteristics of soy globulins as a function of pH and ionic strength. Colloids Surf. A Physicochem. Eng. Asp. 2007, 309, 202–215. [Google Scholar] [CrossRef]
- Keerati-u-rai, M.; Corredig, M. Heat-Induced Changes Occurring in Oil/Water Emulsions Stabilized by Soy Glycinin and β-Conglycinin. J. Agric. Food Chem. 2010, 58, 9171–9180. [Google Scholar] [CrossRef]
- Chen, M.; Lu, J.; Liu, F.; Nsor-Atindana, J.; Xu, F.; Goff, H.D.; Ma, J.; Zhong, F. Study on the emulsifying stability and interfacial adsorption of pea proteins. Food Hydrocoll. 2019, 88, 247–255. [Google Scholar] [CrossRef]
- Liang, H.-N.; Tang, C.-H. pH-dependent emulsifying properties of pea [Pisum sativum (L.)] proteins. Food Hydrocoll. 2013, 33, 309–319. [Google Scholar] [CrossRef]
- Thewissen, B.G.; Celus, I.; Brijs, K.; Delcour, J.A. Foaming Properties of Wheat Gliadin. J. Agric. Food Chem. 2011, 59, 1370–1375. [Google Scholar] [CrossRef]
- Uruakpa, F. Gelling Behavior of Plant Proteins and Polysaccharides in Food Systems. J. Food Sci. Eng. 2012, 2, 247. [Google Scholar] [CrossRef]
- Dissanayake, M.; Vasiljevic, T. Functional properties of whey proteins affected by heat treatment and hydrodynamic high-pressure shearing. J. Dairy Sci. 2009, 92, 1387–1397. [Google Scholar] [CrossRef]
- Patel, M.T.; Kilara, A. Studies on whey protein concentrates. 2. Foaming and emulsifying properties and their relationships with physicochemical properties. J. Dairy Sci. 1990, 73, 2731–2740. [Google Scholar] [CrossRef]
- Sha, L.; Xiong, Y.L. Plant protein-based alternatives of reconstructed meat: Science, technology, and challenges. Trends Food Sci. Technol. 2020, 102, 51–61. [Google Scholar] [CrossRef]
- McClements, D.J.; Grossmann, L. A brief review of the science behind the design of healthy and sustainable plant-based foods. NPJ Sci. Food 2021, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Tornberg, E. Engineering processes in meat products and how they influence their biophysical properties. Meat Sci. 2013, 95, 871–878. [Google Scholar] [CrossRef]
- Tornberg, E.; Andersson, K.; Andersson, A.; Josell, A. The texture of comminuted meat products. Food Aust. 2000, 52, 519–524. [Google Scholar]
- Feher, J. 3.5-contractile mechanisms in skeletal muscle. In Quantitative Human Physiology; Elsevier: Amsterdam, The Netherlands, 2012; pp. 249–258. [Google Scholar]
- Farrell, H.M.; Qi, P.X.; Brown, E.M.; Cooke, P.H.; Tunick, M.H.; Wickham, E.D.; Unruh, J.J. Molten globule structures in milk proteins: Implications for potential new structure-function relationships. J. Dairy Sci. 2002, 85, 459–471. [Google Scholar] [CrossRef]
- Broyard, C.; Gaucheron, F. Modifications of structures and functions of caseins: A scientific and technological challenge. Dairy Sci. Technol. 2015, 95, 831–862. [Google Scholar] [CrossRef]
- Bhat, M.; Dar, T.; Singh, L. Casein proteins: Structural and functional aspects. In Milk Proteins—From Structure to Biological Properties and Health Aspects; InTech: Rijeka, Croatia, 2016. [Google Scholar] [CrossRef]
- Lin, L.; Regenstein, J.; Lv, S.; Lu, J.; Jiang, S. An overview of gelatin derived from aquatic animals: Properties and modification. Trends Food Sci. Technol. 2017, 68, 102–112. [Google Scholar] [CrossRef]
- Hou, F.; Ding, W.; Qu, W.; Oladejo, A.O.; Xiong, F.; Zhang, W.; He, R.; Ma, H. Alkali solution extraction of rice residue protein isolates: Influence of alkali concentration on protein functional, structural properties and lysinoalanine formation. Food Chem. 2017, 218, 207–215. [Google Scholar] [CrossRef]
- Pelgrom, P.J.M.; Vissers, A.M.; Boom, R.M.; Schutyser, M.A.I. Dry fractionation for production of functional pea protein concentrates. Food Res. Int. 2013, 53, 232–239. [Google Scholar] [CrossRef]
- Hinderink, E.B.A.; Schröder, A.; Sagis, L.; Schroën, K.; Berton-Carabin, C.C. Physical and oxidative stability of food emulsions prepared with pea protein fractions. LWT 2021, 146, 111424. [Google Scholar] [CrossRef]
- Ozdal, T.; Capanoglu, E.; Altay, F. A review on protein–phenolic interactions and associated changes. Food Res. Int. 2013, 51, 954–970. [Google Scholar] [CrossRef]
- Loveday, S.M. Food proteins: Technological, nutritional, and sustainability attributes of traditional and emerging proteins. Annu. Rev. Food Sci. Technol. 2019, 10, 311–339. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Tomar, M.; Potkule, J.; Verma, R.; Punia, S.; Mahapatra, A.; Belwal, T.; Dahuja, A.; Joshi, S.; Berwal, M.K.; et al. Advances in the plant protein extraction: Mechanism and recommendations. Food Hydrocoll. 2021, 115, 106595. [Google Scholar] [CrossRef]
- Watchararuji, K.; Goto, M.; Sasaki, M.; Shotipruk, A. Value-added subcritical water hydrolysate from rice bran and soybean meal. Bioresour. Technol. 2008, 99, 6207–6213. [Google Scholar] [CrossRef]
- Chen, J.; Li, Y.; Wang, X.; Liu, W. Application of deep eutectic solvents in food analysis: A review. Molecules 2019, 24, 4594. [Google Scholar] [CrossRef] [PubMed]
- Grudniewska, A.; de Melo, E.M.; Chan, A.; Gniłka, R.; Boratyński, F.; Matharu, A.S. Enhanced protein extraction from oilseed cakes using glycerol–choline chloride deep eutectic solvents: A biorefinery approach. ACS Sustain. Chem. Eng. 2018, 6, 15791–15800. [Google Scholar] [CrossRef]
- Yue, J.; Zhu, Z.; Yi, J.; Lan, Y.; Chen, B.; Rao, J. Structure and functionality of oat protein extracted by choline chloride‒dihydric alcohol deep eutectic solvent and its water binary mixtures. Food Hydrocoll. 2021, 112, 106330. [Google Scholar] [CrossRef]
- Rozzi, N.L.; Singh, R.K. Supercritical fluids and the food industry. Compr. Rev. Food Sci. Food Saf. 2002, 1, 33–44. [Google Scholar] [CrossRef]
- Samaranayake, C.P.; Sastry, S.K. In-situ pH measurement of selected liquid foods under high pressure. Innov. Food Sci. Emerg. Technol. 2013, 17, 22–26. [Google Scholar] [CrossRef]
- Boye, J.I.; Aksay, S.; Roufik, S.; Ribéreau, S.; Mondor, M.; Farnworth, E.; Rajamohamed, S.H. Comparison of the functional properties of pea, chickpea and lentil protein concentrates processed using ultrafiltration and isoelectric precipitation techniques. Food Res. Int. 2010, 43, 537–546. [Google Scholar] [CrossRef]
- She, Q.; Tang, C.Y.; Wang, Y.-N.; Zhang, Z. The role of hydrodynamic conditions and solution chemistry on protein fouling during ultrafiltration. Desalination 2009, 249, 1079–1087. [Google Scholar] [CrossRef]
- Su, Y.P.; Sim, L.N.; Li, X.; Coster, H.G.L.; Chong, T.H. Anti-fouling piezoelectric PVDF membrane: Effect of morphology on dielectric and piezoelectric properties. J. Membr. Sci. 2021, 620, 118818. [Google Scholar] [CrossRef]
- Boretti, A.; Al-Zubaidy, S.; Vaclavikova, M.; Al-Abri, M.; Castelletto, S.; Mikhalovsky, S. Outlook for graphene-based desalination membranes. NPJ Clean Water 2018, 1, 5. [Google Scholar] [CrossRef]
- Lam, A.C.Y.; Can Karaca, A.; Tyler, R.T.; Nickerson, M.T. Pea protein isolates: Structure, extraction, and functionality. Food Rev. Int. 2018, 34, 126–147. [Google Scholar] [CrossRef]
- Yang, J.; Zamani, S.; Liang, L.; Chen, L. Extraction methods significantly impact pea protein composition, structure and gelling properties. Food Hydrocoll. 2021, 117, 106678. [Google Scholar] [CrossRef]
- Kornet, R.; Shek, C.; Venema, P.; Jan van der Goot, A.; Meinders, M.; van der Linden, E. Substitution of whey protein by pea protein is facilitated by specific fractionation routes. Food Hydrocoll. 2021, 117, 106691. [Google Scholar] [CrossRef]
- Emkani, M.; Oliete, B.; Saurel, R. Pea protein extraction assisted by lactic fermentation: Impact on protein profile and thermal properties. Foods 2021, 10, 549. [Google Scholar] [CrossRef]
- Peng, Y.; Kersten, N.; Kyriakopoulou, K.; van der Goot, A.J. Functional properties of mildly fractionated soy protein as influenced by the processing pH. J. Food Eng. 2020, 275, 109875. [Google Scholar] [CrossRef]
- Sara, A.S.; Mathé, C.; Basselin, M.; Fournier, F.; Aymes, A.; Bianeis, M.; Galet, O.; Kapel, R. Optimization of sunflower albumin extraction from oleaginous meal and characterization of their structure and properties. Food Hydrocoll. 2020, 99, 105335. [Google Scholar] [CrossRef]
- Flory, P.J. Principles of Polymer Chemistry; Cornell University Press: Ithaca, NY, USA, 1953. [Google Scholar]
- Gonzalez-Coloma, A.; Martín, L.; Mainar, A.M.; Urieta, J.S.; Fraga, B.M.; Rodríguez-Vallejo, V.; Díaz, C.E. Supercritical extraction and supercritical antisolvent fractionation of natural products from plant material: Comparative results on Persea indica. Phytochem. Rev. 2012, 11, 433–446. [Google Scholar] [CrossRef]
- Zydney, A.L. Protein separations using membrane filtration: New opportunities for whey fractionation. Int. Dairy J. 1998, 8, 243–250. [Google Scholar] [CrossRef]
- Arrutia, F.; Binner, E.; Williams, P.; Waldron, K.W. Oilseeds beyond oil: Press cakes and meals supplying global protein requirements. Trends Food Sci. Technol. 2020, 100, 88–102. [Google Scholar] [CrossRef]
- Naibaho, J.; Korzeniowska, M. Brewers’ spent grain in food systems: Processing and final products quality as a function of fiber modification treatment. J. Food Sci. 2021, 86, 1532–1551. [Google Scholar] [CrossRef]
- Kamal, H.; Le, C.F.; Salter, A.M.; Ali, A. Extraction of protein from food waste: An overview of current status and opportunities. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2455–2475. [Google Scholar] [CrossRef]
- Fernando, S. Production of protein-rich pulse ingredients through dry fractionation: A review. LWT 2021, 141, 110961. [Google Scholar] [CrossRef]
- Assatory, A.; Vitelli, M.; Rajabzadeh, A.R.; Legge, R.L. Dry fractionation methods for plant protein, starch and fiber enrichment: A review. Trends Food Sci. Technol. 2019, 86, 340–351. [Google Scholar] [CrossRef]
- Zhu, H.-G.; Tang, H.-Q.; Cheng, Y.-Q.; Li, Z.-G.; Tong, L.-T. Electrostatic separation technology for obtaining plant protein concentrates: A review. Trends Food Sci. Technol. 2021, 113, 66–76. [Google Scholar] [CrossRef]
- Kornet, R.; Penris, S.; Venema, P.; van der Goot, A.J.; Meinders, M.B.J.; van der Linden, E. How pea fractions with different protein composition and purity can substitute WPI in heat-set gels. Food Hydrocoll. 2021, 120, 106891. [Google Scholar] [CrossRef]
- Mession, J.-L.; Chihi, M.L.; Sok, N.; Saurel, R. Effect of globular pea proteins fractionation on their heat-induced aggregation and acid cold-set gelation. Food Hydrocoll. 2015, 46, 233–243. [Google Scholar] [CrossRef]
- Irache, J.M.; Bergougnoux, L.; Ezpeleta, I.; Gueguen, J.; Orecchioni, A.-M. Optimization and in vitro stability of legumin nanoparticles obtained by a coacervation method. Int. J. Pharm. 1995, 126, 103–109. [Google Scholar] [CrossRef]
- Moll, P.; Salminen, H.; Schmitt, C.; Weiss, J. Impact of microfluidization on colloidal properties of insoluble pea protein fractions. Eur. Food Res. Technol. 2021, 247, 545–554. [Google Scholar] [CrossRef]
- Bahrami, N.; Bayliss, D.; Chope, G.; Penson, S.; Perehinec, T.; Fisk, I.D. Cold plasma: A new technology to modify wheat flour functionality. Food Chem. 2016, 202, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Cai, M.-j.; Cheng, J.-H.; Sun, D.-W. Effects of electric fields and electromagnetic wave on food protein structure and functionality: A review. Trends Food Sci. Technol. 2018, 75, 1–9. [Google Scholar] [CrossRef]
- Dhalleine, C.; Passe, D. Process for Manufacturing Soluble and Functional Plant Proteins, Products Obtained and Uses. U.S. Patent No. 9,259,017, 16 February 2016. [Google Scholar]
- Djemaoune, Y.; Cases, E.; Saurel, R. the effect of high-pressure microfluidization treatment on the foaming properties of pea albumin aggregates. J. Food Sci. 2019, 84, 2242–2249. [Google Scholar] [CrossRef]
- Devi, A.F.; Buckow, R.; Hemar, Y.; Kasapis, S. Structuring dairy systems through high pressure processing. J. Food Eng. 2013, 114, 106–122. [Google Scholar] [CrossRef]
- Mozhaev, V.V.; Heremans, K.; Frank, J.; Masson, P.; Balny, C. High pressure effects on protein structure and function. Proteins Struct. Funct. Bioinform. 1996, 24, 81–91. [Google Scholar] [CrossRef]
- Balny, C.; Masson, P. Effects of high pressure on proteins. Food Rev. Int. 1993, 9, 611–628. [Google Scholar] [CrossRef]
- Balasubramaniam, V.; Barbosa-Cánovas, G.V.; Lelieveld, H. High. Pressure Processing of Food: Principles, Technology and Applications; Springer: Berlin, Germany, 2016. [Google Scholar]
- Queirós, R.P.; Saraiva, J.A.; da Silva, J.A.L. Tailoring structure and technological properties of plant proteins using high hydrostatic pressure. Crit. Rev. Food Sci. Nutr. 2018, 58, 1538–1556. [Google Scholar] [CrossRef]
- Gharibzahedi, S.M.T.; Smith, B. Effects of high hydrostatic pressure on the quality and functionality of protein isolates, concentrates, and hydrolysates derived from pulse legumes: A review. Trends Food Sci. Technol. 2021, 107, 466–479. [Google Scholar] [CrossRef]
- Sim, S.Y.J.; Karwe, M.V.; Moraru, C.I. High pressure structuring of pea protein concentrates. J. Food Process. Eng. 2019, 42, e13261. [Google Scholar] [CrossRef]
- Dos Santos Aguilar, J.G.; Cristianini, M.; Sato, H.H. Modification of enzymes by use of high-pressure homogenization. Food Res. Int. 2018, 109, 120–125. [Google Scholar] [CrossRef]
- Saricaoglu, F.T.; Gul, O.; Besir, A.; Atalar, I. Effect of high pressure homogenization (HPH) on functional and rheological properties of hazelnut meal proteins obtained from hazelnut oil industry by-products. J. Food Eng. 2018, 233, 98–108. [Google Scholar] [CrossRef]
- Saricaoglu, F.T. Application of high-pressure homogenization (HPH) to modify functional, structural and rheological properties of lentil (Lens culinaris) proteins. Int. J. Biol. Macromol. 2020, 144, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Porto, B.C.; Tribst, A.A.L.; Cristianini, M. Chapter 10—Dynamic high pressure effects on biopolymers: Polysaccharides and proteins. In Biopolymers for Food Design; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 313–350. [Google Scholar] [CrossRef]
- Yang, J.; Liu, G.; Zeng, H.; Chen, L. Effects of high pressure homogenization on faba bean protein aggregation in relation to solubility and interfacial properties. Food Hydrocoll. 2018, 83, 275–286. [Google Scholar] [CrossRef]
- Guo, Z.; Huang, Z.; Guo, Y.; Li, B.; Yu, W.; Zhou, L.; Jiang, L.; Teng, F.; Wang, Z. Effects of high-pressure homogenization on structural and emulsifying properties of thermally soluble aggregated kidney bean (Phaseolus vulgaris L.) proteins. Food Hydrocoll. 2021, 119, 106835. [Google Scholar] [CrossRef]
- Rossen, J.L.; Miller, R.C. Food extrusion. Food Technol. 1973, 27, 46–53. [Google Scholar]
- Chiang, J.H.; Loveday, S.M.; Hardacre, A.K.; Parker, M.E. Effects of soy protein to wheat gluten ratio on the physicochemical properties of extruded meat analogues. Food Struct. 2019, 19, 100102. [Google Scholar] [CrossRef]
- Osen, R.; Toelstede, S.; Wild, F.; Eisner, P.; Schweiggert-Weisz, U. High moisture extrusion cooking of pea protein isolates: Raw material characteristics, extruder responses, and texture properties. J. Food Eng. 2014, 127, 67–74. [Google Scholar] [CrossRef]
- Strahm, B. Meat alternatives. In Soy Applications in Food, 1st ed.; Riaz, M., Ed.; CRC Press: Boca Raton, FL, USA, 2005; pp. 135–154. [Google Scholar]
- Beck, S.M.; Knoerzer, K.; Arcot, J. Effect of low moisture extrusion on a pea protein isolate’s expansion, solubility, molecular weight distribution and secondary structure as determined by Fourier Transform Infrared Spectroscopy (FTIR). J. Food Eng. 2017, 214, 166–174. [Google Scholar] [CrossRef]
- Samard, S.; Gu, B.-Y.; Ryu, G.-H. Effects of extrusion types, screw speed and addition of wheat gluten on physicochemical characteristics and cooking stability of meat analogues. J. Sci. Food Agric. 2019, 99, 4922–4931. [Google Scholar] [CrossRef] [PubMed]
- Nosworthy, M.G.; Medina, G.; Franczyk, A.J.; Neufeld, J.; Appah, P.; Utioh, A.; Frohlich, P.; Tar’an, B.; House, J.D. Thermal processing methods differentially affect the protein quality of Chickpea (Cicer arietinum). Food Sci. Nutr. 2020, 8, 2950–2958. [Google Scholar] [CrossRef] [PubMed]
- Omosebi, M.O.; Osundahunsi, O.F.; Fagbemi, T.N. Effect of extrusion on protein quality, antinutritional factors, and digestibility of complementary diet from quality protein maize and soybean protein concentrate. J. Food Biochem. 2018, 42, e12508. [Google Scholar] [CrossRef]
- Alfaro-Diaz, A.; Urías-Silvas, J.E.; Loarca-Piña, G.; Gaytan-Martínez, M.; Prado-Ramirez, R.; Mojica, L. Techno-functional properties of thermally treated black bean protein concentrate generated through ultrafiltration process. LWT 2021, 136, 110296. [Google Scholar] [CrossRef]
- Chen, L.; Chen, J.; Yu, L.; Wu, K.; Zhao, M. Emulsification performance and interfacial properties of enzymically hydrolyzed peanut protein isolate pretreated by extrusion cooking. Food Hydrocoll. 2018, 77, 607–616. [Google Scholar] [CrossRef]
- Zhou, Q.-C.; Liu, N.; Feng, C.-X. Research on the effect of papain co-extrusion on pea protein and enzymolysis antioxidant peptides. J. Food Process. Preserv. 2017, 41, e13301. [Google Scholar] [CrossRef]
- Berovič, M. 2.12—Sterilization in biotechnology. In Comprehensive Biotechnology, 2nd ed.; Moo-Young, M., Ed.; Academic Press: Burlington, MA, USA, 2011; pp. 135–150. [Google Scholar] [CrossRef]
- Chung, D.D.L. 6—Cement-matrix composites. In Carbon Composites, 2nd ed.; Chung, D.D.L., Ed.; Butterworth-Heinemann: Oxford, UK, 2017; pp. 333–386. [Google Scholar] [CrossRef]
- Ghauri, A.; Ghauri, I.; Elhissi, A.M.A.; Ahmed, W. Chapter 14—Characterization of cochleate nanoparticles for delivery of the anti-asthma drug beclomethasone dipropionate. In Advances in Medical and Surgical Engineering; Ahmed, W., Phoenix, D.A., Jackson, M.J., Charalambous, C.P., Eds.; Academic Press: Burlington, MA, USA, 2020; pp. 267–277. [Google Scholar] [CrossRef]
- O’Sullivan, J.; Park, M.; Beevers, J. The effect of ultrasound upon the physicochemical and emulsifying properties of wheat and soy protein isolates. J. Cereal Sci. 2016, 69, 77–84. [Google Scholar] [CrossRef]
- O’Sullivan, J.J.; Park, M.; Beevers, J.; Greenwood, R.W.; Norton, I.T. Applications of ultrasound for the functional modification of proteins and nanoemulsion formation: A review. Food Hydrocoll. 2017, 71, 299–310. [Google Scholar] [CrossRef]
- Arzeni, C.; Martínez, K.; Zema, P.; Arias, A.; Pérez, O.E.; Pilosof, A.M.R. Comparative study of high intensity ultrasound effects on food proteins functionality. J. Food Eng. 2012, 108, 463–472. [Google Scholar] [CrossRef]
- Xiong, T.; Xiong, W.; Ge, M.; Xia, J.; Li, B.; Chen, Y. Effect of high intensity ultrasound on structure and foaming properties of pea protein isolate. Food Res. Int. 2018, 109, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhu, W.; Yi, J.; Liu, N.; Cao, Y.; Lu, J.; Decker, E.A.; McClements, D.J. Effects of sonication on the physicochemical and functional properties of walnut protein isolate. Food Res. Int. 2018, 106, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Okagu, O.D.; Yagoub, A.E.A.; Udenigwe, C.C. Effects of sonication on the in vitro digestibility and structural properties of buckwheat protein isolates. Ultrason. Sonochem. 2021, 70, 105348. [Google Scholar] [CrossRef] [PubMed]
- Gharibzahedi, S.M.T.; Smith, B. The functional modification of legume proteins by ultrasonication: A review. Trends Food Sci. Technol. 2020, 98, 107–116. [Google Scholar] [CrossRef]
- Zink, J.; Wyrobnik, T.; Prinz, T.; Schmid, M. Physical, chemical and biochemical modifications of protein-based films and coatings: An extensive review. Int. J. Mol. Sci. 2016, 17, 1376. [Google Scholar] [CrossRef]
- Van Boekel, M.A.J.S. Kinetic aspects of the Maillard reaction: A critical review. Food Nahr. 2001, 45, 150–159. [Google Scholar] [CrossRef]
- De Oliveira, F.C.; Coimbra, J.S.d.R.; de Oliveira, E.B.; Zuñiga, A.D.G.; Rojas, E.E.G. Food Protein-polysaccharide conjugates obtained via the Maillard reaction: A review. Crit. Rev. Food Sci. Nutr. 2016, 56, 1108–1125. [Google Scholar] [CrossRef]
- Kutzli, I.; Weiss, J.; Gibis, M. Glycation of plant proteins via maillard reaction: Reaction chemistry, technofunctional properties, and potential food application. Foods 2021, 10, 376. [Google Scholar] [CrossRef]
- Oliver, C.M.; Melton, L.D.; Stanley, R.A. Creating proteins with novel functionality via the Maillard reaction: A review. Crit. Rev. Food Sci. Nutr. 2006, 46, 337–350. [Google Scholar] [CrossRef]
- Zhu, D.; Damodaran, S.; Lucey, J.A. Formation of Whey Protein Isolate (WPI)−dextran conjugates in aqueous solutions. J. Agric. Food Chem. 2008, 56, 7113–7118. [Google Scholar] [CrossRef]
- Dickinson, E.; Galazka, V.B. Emulsion stabilization by ionic and covalent complexes of β-lactoglobulin with polysaccharides. Food Hydrocoll. 1991, 5, 281–296. [Google Scholar] [CrossRef]
- Lam, R.S.H.; Nickerson, M.T. Food proteins: A review on their emulsifying properties using a structure-function approach. Food Chem. 2013, 141, 975–984. [Google Scholar] [CrossRef]
- Saatchi, A.; Kiani, H.; Labbafi, M. A new functional protein-polysaccharide conjugate based on protein concentrate from sesame processing by-products: Functional and physico-chemical properties. Int. J. Biol. Macromol. 2019, 122, 659–666. [Google Scholar] [CrossRef]
- Li, R.; Cui, Q.; Wang, G.; Liu, J.; Chen, S.; Wang, X.; Wang, X.; Jiang, L. Relationship between surface functional properties and flexibility of soy protein isolate-glucose conjugates. Food Hydrocoll. 2019, 95, 349–357. [Google Scholar] [CrossRef]
- Wen, C.; Zhang, J.; Qin, W.; Gu, J.; Zhang, H.; Duan, Y.; Ma, H. Structure and functional properties of soy protein isolate-lentinan conjugates obtained in Maillard reaction by slit divergent ultrasonic assisted wet heating and the stability of oil-in-water emulsions. Food Chem. 2020, 331, 127374. [Google Scholar] [CrossRef]
- Zha, F.; Yang, Z.; Rao, J.; Chen, B. Gum arabic-mediated synthesis of glyco-pea protein hydrolysate via Maillard reaction improves solubility, flavor profile, and functionality of plant protein. J. Agric. Food Chem. 2019, 67, 10195–10206. [Google Scholar] [CrossRef] [PubMed]
- Yongling, S.; Ruiqi, Q.I.N.; Shaoming, Y.; Jianghe, L.I.; Ruolan, W. Improvement of foaming and emulsifying properties of gluten by conjugation with fructose through Maillard reaction. Grain Oil Sci. Technol. 2018, 1, 119–125. [Google Scholar] [CrossRef][Green Version]
- Alavi, F.; Chen, L.; Wang, Z.; Emam-Djomeh, Z. Consequences of heating under alkaline pH alone or in the presence of maltodextrin on solubility, emulsifying and foaming properties of faba bean protein. Food Hydrocoll. 2021, 112, 106335. [Google Scholar] [CrossRef]
- Wang, Q.; Jin, Y.; Xiong, Y.L. Heating-aided pH shifting modifies hemp seed protein structure, cross-linking, and emulsifying properties. J. Agric. Food Chem. 2018, 66, 10827–10834. [Google Scholar] [CrossRef]
- Hirose, M. Molten globule state of food proteins. Trends Food Sci. Technol. 1993, 4, 48–51. [Google Scholar] [CrossRef]
- Christensen, H.; Pain, R. Molten globule intermediates and protein folding. Eur. Biophys. J. 1991, 19, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, C.; Zhao, J.; Ma, T.; He, Z.; Huang, M.; Wang, Y. Modification of structure and functionalities of ginkgo seed proteins by pH-shifting treatment. Food Chem. 2021, 358, 129862. [Google Scholar] [CrossRef] [PubMed]
- Silventoinen, P.; Sozer, N. Impact of ultrasound treatment and pH-shifting on physicochemical properties of protein-enriched barley fraction and barley protein isolate. Foods 2020, 9, 1055. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, G.; Ding, J.; Andrade, J.; Engeseth, N.J.; Feng, H. Effect of plant protein-polysaccharide complexes produced by mano-thermo-sonication and pH-shifting on the structure and stability of oil-in-water emulsions. Innov. Food Sci. Emerg. Technol. 2018, 47, 317–325. [Google Scholar] [CrossRef]
- Yildiz, G.; Andrade, J.; Engeseth, N.E.; Feng, H. Functionalizing soy protein nano-aggregates with pH-shifting and mano-thermo-sonication. J. Colloid Interface Sci. 2017, 505, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Muneer, F.; Johansson, E.; Hedenqvist, M.S.; Plivelic, T.S.; Kuktaite, R. Impact of pH modification on protein polymerization and structure–function relationships in potato protein and wheat gluten composites. Int. J. Mol. Sci 2019, 20, 58. [Google Scholar] [CrossRef]
- Li, J.; Wu, M.; Wang, Y.; Li, K.; Du, J.; Bai, Y. Effect of pH-shifting treatment on structural and heat induced gel properties of peanut protein isolate. Food Chem. 2020, 325, 126921. [Google Scholar] [CrossRef]
- Jiang, J.; Xiong, Y.L.; Chen, J. pH shifting alters solubility characteristics and thermal stability of soy protein isolate and its globulin fractions in different pH, salt concentration, and temperature conditions. J. Agric. Food Chem. 2010, 58, 8035–8042. [Google Scholar] [CrossRef]
- Lee, H.; Yildiz, G.; Dos Santos, L.; Jiang, S.; Andrade, J.; Engeseth, N.; Feng, H. Soy protein nano-aggregates with improved functional properties prepared by sequential pH treatment and ultrasonication. Food Hydrocoll. 2016, 55, 200–209. [Google Scholar] [CrossRef]
- Zhang, L.; Pan, Z.; Shen, K.; Cai, X.; Zheng, B.; Miao, S. Influence of ultrasound-assisted alkali treatment on the structural properties and functionalities of rice protein. J. Cereal Sci. 2018, 79, 204–209. [Google Scholar] [CrossRef]
- Jiang, S.; Ding, J.; Andrade, J.; Rababah, T.M.; Almajwal, A.; Abulmeaty, M.M.; Feng, H. Modifying the physicochemical properties of pea protein by pH-shifting and ultrasound combined treatments. Ultrason. Sonochem. 2017, 38, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Z.; Ren, W.; Wang, Y.; Mintah, B.K.; Dabbour, M.; Hou, Y.; He, R.; Cheng, Y.; Ma, H. Inhibition effect of ultrasound on the formation of lysinoalanine in rapeseed protein isolates during pH shift treatment. J. Agric. Food Chem. 2021, 69, 8536–8545. [Google Scholar] [CrossRef] [PubMed]
- Villamil, O.; Váquiro, H.; Solanilla, J.F. Fish viscera protein hydrolysates: Production, potential applications and functional and bioactive properties. Food Chem. 2017, 224, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.D.; Arntfield, S.D. Gelation properties of salt-extracted pea protein isolate catalyzed by microbial transglutaminase cross-linking. Food Hydrocoll. 2011, 25, 25–31. [Google Scholar] [CrossRef]
- Nivala, O.; Mäkinen, O.E.; Kruus, K.; Nordlund, E.; Ercili-Cura, D. Structuring colloidal oat and faba bean protein particles via enzymatic modification. Food Chem. 2017, 231, 87–95. [Google Scholar] [CrossRef]
- Zang, X.; Yue, C.; Wang, Y.; Shao, M.; Yu, G. Effect of limited enzymatic hydrolysis on the structure and emulsifying properties of rice bran protein. J. Cereal Sci. 2019, 85, 168–174. [Google Scholar] [CrossRef]
- Beaubier, S.; Albe-Slabi, S.; Aymes, A.; Bianeis, M.; Galet, O.; Kapel, R. A rational approach for the production of highly soluble and functional sunflower protein hydrolysates. Foods 2021, 10, 664. [Google Scholar] [CrossRef]
- Klost, M.; Drusch, S. Functionalisation of pea protein by tryptic hydrolysis—Characterisation of interfacial and functional properties. Food Hydrocoll. 2019, 86, 134–140. [Google Scholar] [CrossRef]
- Schlegel, K.; Sontheimer, K.; Eisner, P.; Schweiggert-Weisz, U. Effect of enzyme-assisted hydrolysis on protein pattern, technofunctional, and sensory properties of lupin protein isolates using enzyme combinations. Food Sci. Nutr. 2020, 8, 3041–3051. [Google Scholar] [CrossRef]
- Aguilar, J.G.d.S.; Granato Cason, V.; de Castro, R.J.S. Improving antioxidant activity of black bean protein by hydrolysis with protease combinations. Int. J. Food Sci. Technol. 2019, 54, 34–41. [Google Scholar] [CrossRef]
- Guan, H.; Diao, X.; Jiang, F.; Han, J.; Kong, B. The enzymatic hydrolysis of soy protein isolate by Corolase PP under high hydrostatic pressure and its effect on bioactivity and characteristics of hydrolysates. Food Chem. 2018, 245, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Venuste, M.; Zhang, X.; Shoemaker, C.F.; Karangwa, E.; Abbas, S.; Kamdem, P.E. Influence of enzymatic hydrolysis and enzyme type on the nutritional and antioxidant properties of pumpkin meal hydrolysates. Food Funct. 2013, 4, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, L.; Yuan, D.; Zhao, X.; Cui, S.; Hu, J.; Wang, J. Reduction of the allergenic protein in soybean meal by enzymatic hydrolysis. Food Agric. Immunol. 2014, 25, 301–310. [Google Scholar] [CrossRef]
- Zhao, J.; Xiong, Y.L. Interfacial peptide partitioning and undiminished antioxidative and emulsifying activity of oxidatively stressed soy protein hydrolysate in an O/W emulsion. LWT Food Sci. Technol. 2015, 61, 322–329. [Google Scholar] [CrossRef]
- Borsook, H.; Wasteneys, H. The enzymatic synthesis of protein. II. The effect of temperature on the synthesizing action of pepsin. J. Biol. Chem. 1925, 62, 633–639. [Google Scholar]
- Andrews, A.T.; Alichanidis, E. The plastein reaction revisited: Evidence for a purely aggregation reaction mechanism. Food Chem. 1990, 35, 243–261. [Google Scholar] [CrossRef]
- Stevenson, D.E.; Ofman, D.J.; Morgan, K.R.; Stanley, R.A. Protease-catalyzed condensation of peptides as a potential means to reduce the bitter taste of hydrophobic peptides found in protein hydrolysates. Enzym. Microb. Technol. 1998, 22, 100–110. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Wu, S.; Drummond, K.; Gong, M. Revisiting the prospects of plastein: Thermal and simulated gastric stability in relation to the antioxidative capacity of casein plastein. J. Agric. Food Chem. 2014, 62, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Arai, S.; Kokubo, S.; Aso, K.; Fujimaki, M. A plastein with an extremely high amount of glutamic acid. Agric. Biol. Chem. 1974, 38, 1269–1271. [Google Scholar] [CrossRef]
- Gong, M.; Mohan, A.; Gibson, A.; Udenigwe, C.C. Mechanisms of plastein formation, and prospective food and nutraceutical applications of the peptide aggregates. Biotechnol. Rep. 2015, 5, 63–69. [Google Scholar] [CrossRef]
- Stephen, A.M.; Phillips, G.O. Food Polysaccharides and Their Applications; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Le, X.T.; Rioux, L.-E.; Turgeon, S.L. Formation and functional properties of protein–polysaccharide electrostatic hydrogels in comparison to protein or polysaccharide hydrogels. Adv. Colloid Interface Sci. 2017, 239, 127–135. [Google Scholar] [CrossRef]
- Schmitt, C.; Sanchez, C.; Desobry-Banon, S.; Hardy, J. Structure and technofunctional properties of protein-polysaccharide complexes: A review. Crit. Rev. Food Sci. Nutr. 1998, 38, 689–753. [Google Scholar] [CrossRef]
- Stounbjerg, L.; Andreasen, B.; Ipsen, R. Microparticles formed by heating potato protein—Polysaccharide electrostatic complexes. J. Food Eng. 2019, 263, 79–86. [Google Scholar] [CrossRef]
- Kutzli, I.; Gibis, M.; Baier, S.K.; Weiss, J. Electrospinning of whey and soy protein mixed with maltodextrin—Influence of protein type and ratio on the production and morphology of fibers. Food Hydrocoll. 2019, 93, 206–214. [Google Scholar] [CrossRef]
- Lan, Y.; Chen, B.; Rao, J. Pea protein isolate–high methoxyl pectin soluble complexes for improving pea protein functionality: Effect of pH, biopolymer ratio and concentrations. Food Hydrocoll. 2018, 80, 245–253. [Google Scholar] [CrossRef]
- Liu, S.; Elmer, C.; Low, N.H.; Nickerson, M.T. Effect of pH on the functional behaviour of pea protein isolate-gum Arabic complexes. Food Res. Int. 2010, 43, 489–495. [Google Scholar] [CrossRef]
- Li, C.; Xue, H.; Chen, Z.; Ding, Q.; Wang, X. Comparative studies on the physicochemical properties of peanut protein isolate–polysaccharide conjugates prepared by ultrasonic treatment or classical heating. Food Res. Int. 2014, 57, 1–7. [Google Scholar] [CrossRef]
- Zhang, X.; Qi, J.-R.; Li, K.-K.; Yin, S.-W.; Wang, J.-M.; Zhu, J.-H.; Yang, X.-Q. Characterization of soy β-conglycinin-dextran conjugate prepared by Maillard reaction in crowded liquid system. Food Res. Int. 2012, 49, 648–654. [Google Scholar] [CrossRef]
- Qu, W.; Zhang, X.; Han, X.; Wang, Z.; He, R.; Ma, H. Structure and functional characteristics of rapeseed protein isolate-dextran conjugates. Food Hydrocoll. 2018, 82, 329–337. [Google Scholar] [CrossRef]
- Cai, Y.; Huang, L.; Tao, X.; Su, J.; Chen, B.; Zhou, F.; Zhao, M.; Zhao, Q.; Van der Meeren, P. Effect of pH on okara protein-carboxymethyl cellulose interactions in aqueous solution and at oil-water interface. Food Hydrocoll. 2021, 113, 106529. [Google Scholar] [CrossRef]
- Mession, J.L.; Assifaoui, A.; Lafarge, C.; Saurel, R.; Cayot, P. Protein aggregation induced by phase separation in a pea proteins–sodium alginate-water ternary system. Food Hydrocoll. 2012, 28, 333–343. [Google Scholar] [CrossRef]
- Mohanan, A.; Nickerson, M.T.; Ghosh, S. Utilization of pulse protein-xanthan gum complexes for foam stabilization: The effect of protein concentrate and isolate at various pH. Food Chem. 2020, 316, 126282. [Google Scholar] [CrossRef] [PubMed]
- Dawa, Q.; Hua, Y.; Chamba, M.V.M.; Masamba, K.G.; Zhang, C. Effect of Xanthan and Arabic gums on foaming properties of pumpkin (Cucurbita pepo) seed protein isolate. J. Food Res. 2014, 3, 87. [Google Scholar] [CrossRef]
- Naderi, B.; Keramat, J.; Nasirpour, A.; Aminifar, M. Complex coacervation between oak protein isolate and gum Arabic: Optimization & functional characterization. Int. J. Food Prop. 2020, 23, 1854–1873. [Google Scholar] [CrossRef]
- Yin, B.; Zhang, R.; Yao, P. Influence of pea protein aggregates on the structure and stability of pea protein/soybean polysaccharide complex emulsions. Molecules 2015, 20, 5165–5183. [Google Scholar] [CrossRef] [PubMed]
- Yin, B.; Deng, W.; Xu, K.; Huang, L.; Yao, P. Stable nano-sized emulsions produced from soy protein and soy polysaccharide complexes. J. Colloid Interface Sci. 2012, 380, 51–59. [Google Scholar] [CrossRef]
- Kasran, M.; Cui, S.W.; Goff, H.D. Covalent attachment of fenugreek gum to soy whey protein isolate through natural Maillard reaction for improved emulsion stability. Food Hydrocoll. 2013, 30, 552–558. [Google Scholar] [CrossRef]
- Zha, F.; Dong, S.; Rao, J.; Chen, B. Pea protein isolate-gum Arabic Maillard conjugates improves physical and oxidative stability of oil-in-water emulsions. Food Chem. 2019, 285, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Su, Z.; Meng, X.; Zhang, X.; Kennedy, J.F.; Liu, B. Fabrication and characterization of Pickering emulsion stabilized by soy protein isolate-chitosan nanoparticles. Carbohydr. Polym. 2020, 247, 116712. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Wang, X.; Wang, X.; Zhang, X.; Gu, Y.; Xia, S.; Huang, Q. High internal phase pickering emulsions stabilized by pea protein isolate-high methoxyl pectin-EGCG complex: Interfacial properties and microstructure. Food Chem. 2021, 350, 129251. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Li, Y. Acylation modification and/or guar gum conjugation enhanced functional properties of pea protein isolate. Food Hydrocoll. 2021, 117, 106686. [Google Scholar] [CrossRef]
- Uruakpa, F.O.; Arntfield, S.D. Emulsifying characteristics of commercial canola protein–hydrocolloid systems. Food Res. Int. 2005, 38, 659–672. [Google Scholar] [CrossRef]
- Dong, D.; Hua, Y. Emulsifying behaviors and interfacial properties of different protein/gum arabic complexes: Effect of pH. Food Hydrocoll. 2018, 74, 289–295. [Google Scholar] [CrossRef]
- Zárate-Ramírez, L.S.; Romero, A.; Bengoechea, C.; Partal, P.; Guerrero, A. Thermo-mechanical and hydrophilic properties of polysaccharide/gluten-based bioplastics. Carbohydr. Polym. 2014, 112, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Wee, M.S.M.; Yusoff, R.; Lin, L.; Xu, Y.Y. Effect of polysaccharide concentration and charge density on acid-induced soy protein isolate-polysaccharide gels using HCl. Food Struct. 2017, 13, 45–55. [Google Scholar] [CrossRef]
- Nieto Nieto, T.V.; Wang, Y.; Ozimek, L.; Chen, L. Improved thermal gelation of oat protein with the formation of controlled phase-separated networks using dextrin and carrageenan polysaccharides. Food Res. Int. 2016, 82, 95–103. [Google Scholar] [CrossRef]
- Wang, Y.-R.; Yang, Q.; Li-Sha, Y.-J.; Chen, H.-Q. Structural, gelation properties and microstructure of rice glutelin/sugar beet pectin composite gels: Effects of ionic strengths. Food Chem. 2021, 346, 128956. [Google Scholar] [CrossRef]
- Sim, S.Y.J.; Moraru, C.I. High-pressure processing of pea protein-starch mixed systems: Effect of starch on structure formation. J. Food Process. Eng. 2020, 43, e13352. [Google Scholar] [CrossRef]
- He, Z.; Liu, C.; Zhao, J.; Li, W.; Wang, Y. Physicochemical properties of a ginkgo seed protein-pectin composite gel. Food Hydrocoll. 2021, 118, 106781. [Google Scholar] [CrossRef]
- Joshi, M.; Aldred, P.; Panozzo, J.F.; Kasapis, S.; Adhikari, B. Rheological and microstructural characteristics of lentil starch-lentil protein composite pastes and gels. Food Hydrocoll. 2014, 35, 226–237. [Google Scholar] [CrossRef]
- Li, J.-Y.; Yeh, A.-I.; Fan, K.-L. Gelation characteristics and morphology of corn starch/soy protein concentrate composites during heating. J. Food Eng. 2007, 78, 1240–1247. [Google Scholar] [CrossRef]
- Li, G.-Y.; Chen, Q.-H.; Su, C.-R.; Wang, H.; He, S.; Liu, J.; Nag, A.; Yuan, Y. Soy protein-polysaccharide complex coacervate under physical treatment: Effects of pH, ionic strength and polysaccharide type. Innov. Food Sci. Emerg. Technol. 2021, 68, 102612. [Google Scholar] [CrossRef]
- Stone, A.K.; Teymurova, A.; Dang, Q.; Abeysekara, S.; Karalash, A.; Nickerson, M.T. Formation and functional attributes of electrostatic complexes involving napin protein isolate and anionic polysaccharides. Eur. Food Res. Technol. 2014, 238, 773–780. [Google Scholar] [CrossRef]
- Ducel, V.; Saulnier, P.; Richard, J.; Boury, F. Plant protein–polysaccharide interactions in solutions: Application of soft particle analysis and light scattering measurements. Colloids Surf. B Biointerfaces 2005, 41, 95–102. [Google Scholar] [CrossRef]
- Baeza, R.I.; Carp, D.J.; Pérez, O.E.; Pilosof, A.M.R. κ -Carrageenan—Protein interactions: Effect of proteins on polysaccharide gelling and textural properties. Food Sci. Technol. 2002, 35, 741–747. [Google Scholar] [CrossRef]
- Timilsena, Y.P.; Wang, B.; Adhikari, R.; Adhikari, B. Preparation and characterization of chia seed protein isolate–chia seed gum complex coacervates. Food Hydrocoll. 2016, 52, 554–563. [Google Scholar] [CrossRef]
- Chang, P.G.; Gupta, R.; Timilsena, Y.P.; Adhikari, B. Optimisation of the complex coacervation between canola protein isolate and chitosan. J. Food Eng. 2016, 191, 58–66. [Google Scholar] [CrossRef]
- Busu, N.M.; Amonsou, E.O. Fractionation pH of bambara groundnut (Vigna subterranea) protein impacts the degree of complexation with gum arabic. Food Hydrocoll. 2019, 87, 653–660. [Google Scholar] [CrossRef]
- Hasanvand, E.; Rafe, A. Characterization of flaxseed gum/rice bran protein complex coacervates. Food Biophys. 2018, 13, 387–395. [Google Scholar] [CrossRef]
- Hasanvand, E.; Rafe, A. Rheological and structural properties of rice bran protein-flaxseed (Linum usitatissimum L.) gum complex coacervates. Food Hydrocoll. 2018, 83, 296–307. [Google Scholar] [CrossRef]
- Lan, Y.; Ohm, J.-B.; Chen, B.; Rao, J. Phase behavior and complex coacervation of concentrated pea protein isolate-beet pectin solution. Food Chem. 2020, 307, 125536. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Dong, H.; Gao, J.; Chen, L.; Vasanthan, T. Field pea protein isolate/chitosan complex coacervates: Formation and characterization. Carbohydr. Polym. 2020, 250, 116925. [Google Scholar] [CrossRef] [PubMed]
- Varankovich, N.V.; Khan, N.H.; Nickerson, M.T.; Kalmokoff, M.; Korber, D.R. Evaluation of pea protein–polysaccharide matrices for encapsulation of acid-sensitive bacteria. Food Res. Int. 2015, 70, 118–124. [Google Scholar] [CrossRef]
- Huang, X.; Huang, X.; Gong, Y.; Xiao, H.; McClements, D.J.; Hu, K. Enhancement of curcumin water dispersibility and antioxidant activity using core–shell protein–polysaccharide nanoparticles. Food Res. Int. 2016, 87, 1–9. [Google Scholar] [CrossRef]
- Yang, Y.; Cui, S.; Gong, J.; Miller, S.S.; Wang, Q.; Hua, Y. Stability of citral in oil-in-water emulsions protected by a soy protein–polysaccharide Maillard reaction product. Food Res. Int. 2015, 69, 357–363. [Google Scholar] [CrossRef]
- Santos, J.; Alcaide-González, M.A.; Trujillo-Cayado, L.A.; Carrillo, F.; Alfaro-Rodríguez, M.C. Development of food-grade Pickering emulsions stabilized by a biological macromolecule (xanthan gum) and zein. Int. J. Biol. Macromol. 2020, 153, 747–754. [Google Scholar] [CrossRef]
- Xiong, Y.L. Structure-function relationships of muscle proteins. In Food Proteins and Their Applications; CRC Press: Boca Raton, FL, USA, 2017; pp. 341–392. [Google Scholar]
- Brishti, F.; Zarei, M.; Muhammad, K.; Ismail-Fitry, M.R.; Shukri, R.; Saari, N. Evaluation of the functional properties of mung bean protein isolate for development of textured vegetable protein. Int. Food Res. J. 2017, 24, 1595–1605. [Google Scholar]
- Kyriakopoulou, K.; Keppler, J.K.; van der Goot, A.J. Functionality of ingredients and additives in plant-based meat analogues. Foods 2021, 10, 600. [Google Scholar] [CrossRef]
- Beniwal, A.S.; Singh, J.; Kaur, L.; Hardacre, A.; Singh, H. Meat analogs: Protein restructuring during thermomechanical processing. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1221–1249. [Google Scholar] [CrossRef]
- Dekkers, B.L.; Boom, R.M.; van der Goot, A.J. Structuring processes for meat analogues. Trends Food Sci. Technol. 2018, 81, 25–36. [Google Scholar] [CrossRef]
- Chiang, J.H.; Tay, W.; Ong, D.S.M.; Liebl, D.; Ng, C.P.; Henry, C.J. Physicochemical, textural and structural characteristics of wheat gluten-soy protein composited meat analogues prepared with the mechanical elongation method. Food Struct. 2021, 28, 100183. [Google Scholar] [CrossRef]
- Dekkers, B.L.; Emin, M.A.; Boom, R.M.; van der Goot, A.J. The phase properties of soy protein and wheat gluten in a blend for fibrous structure formation. Food Hydrocoll. 2018, 79, 273–281. [Google Scholar] [CrossRef]
- Krintiras, G.A.; Göbel, J.; Bouwman, W.G.; van der Goot, A.J.; Stefanidis, G.D. On characterization of anisotropic plant protein structures. Food Funct. 2014, 5, 3233–3240. [Google Scholar] [CrossRef]
- Mattice, K.D.; Marangoni, A.G. Comparing methods to produce fibrous material from zein. Food Res. Int. 2020, 128, 108804. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, L.; Liu, H.; Yoon, A.; Rizvi, S.S.H.; Wang, Q. Changes in conformation and quality of vegetable protein during texturization process by extrusion. Crit. Rev. Food Sci. Nutr. 2019, 59, 3267–3280. [Google Scholar] [CrossRef]
- Qi, P.X.; Onwulata, C.I. Physical properties, molecular structures, and protein quality of texturized whey protein isolate: Effect of extrusion moisture content. J. Dairy Sci. 2011, 94, 2231–2244. [Google Scholar] [CrossRef]
- Chiang, J.H.; Hardacre, A.K.; Parker, M.E. Effects of Maillard-reacted beef bone hydrolysate on the physicochemical properties of extruded meat alternatives. J. Food Sci. 2020, 85, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Mezzenga, R. Design principles of food gels. Nat. Food 2020, 1, 106–118. [Google Scholar] [CrossRef]
- DePalma, K.; Smith, B.; McDonald, A.G. Synergistic effects of processing parameters on the biochemical and physical properties of tofu made from yellow field pea (Pisum sativum), as determined by response surface methodology. Food Sci. Nutr. 2021, 9, 1132–1142. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.; Johan, Z.; Bakar, J. Chickpea, mungbean, cowpea and peanuts as substitutes for soybean curds. Int. J. Food Sci. Technol. 1989, 24, 385–394. [Google Scholar] [CrossRef]
- Cai, R.; Klamczynska, B.; Baik, B.K. Preparation of bean curds from protein fractions of six legumes. J. Agric. Food Chem. 2001, 49, 3068–3073. [Google Scholar] [CrossRef] [PubMed]
- Jeske, S.; Zannini, E.; Arendt, E.K. Past, present and future: The strength of plant-based dairy substitutes based on gluten-free raw materials. Food Res. Int. 2018, 110, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Li, L.; Yang, X.-Q.; Ning, Z.-X.; Randhawa, M.A. Improvements in the flavour of soy cheese. Food Technol. Biotechnol. 2008, 46, 252–261. [Google Scholar]
- Mattice, K.D.; Marangoni, A.G. Physical properties of plant-based cheese products produced with zein. Food Hydrocoll. 2020, 105, 105746. [Google Scholar] [CrossRef]
- Margolis, G.; Myers, S.; Newbold, D. Manufacturing of Plant-Based Yogurt. U.S. Patent 16/293,282, 12 September 2019. [Google Scholar]
- Grasso, N.; Alonso-Miravalles, L.; O’Mahony, J.A. Composition, physicochemical and sensorial properties of commercial plant-based yogurts. Foods 2020, 9, 252. [Google Scholar] [CrossRef]
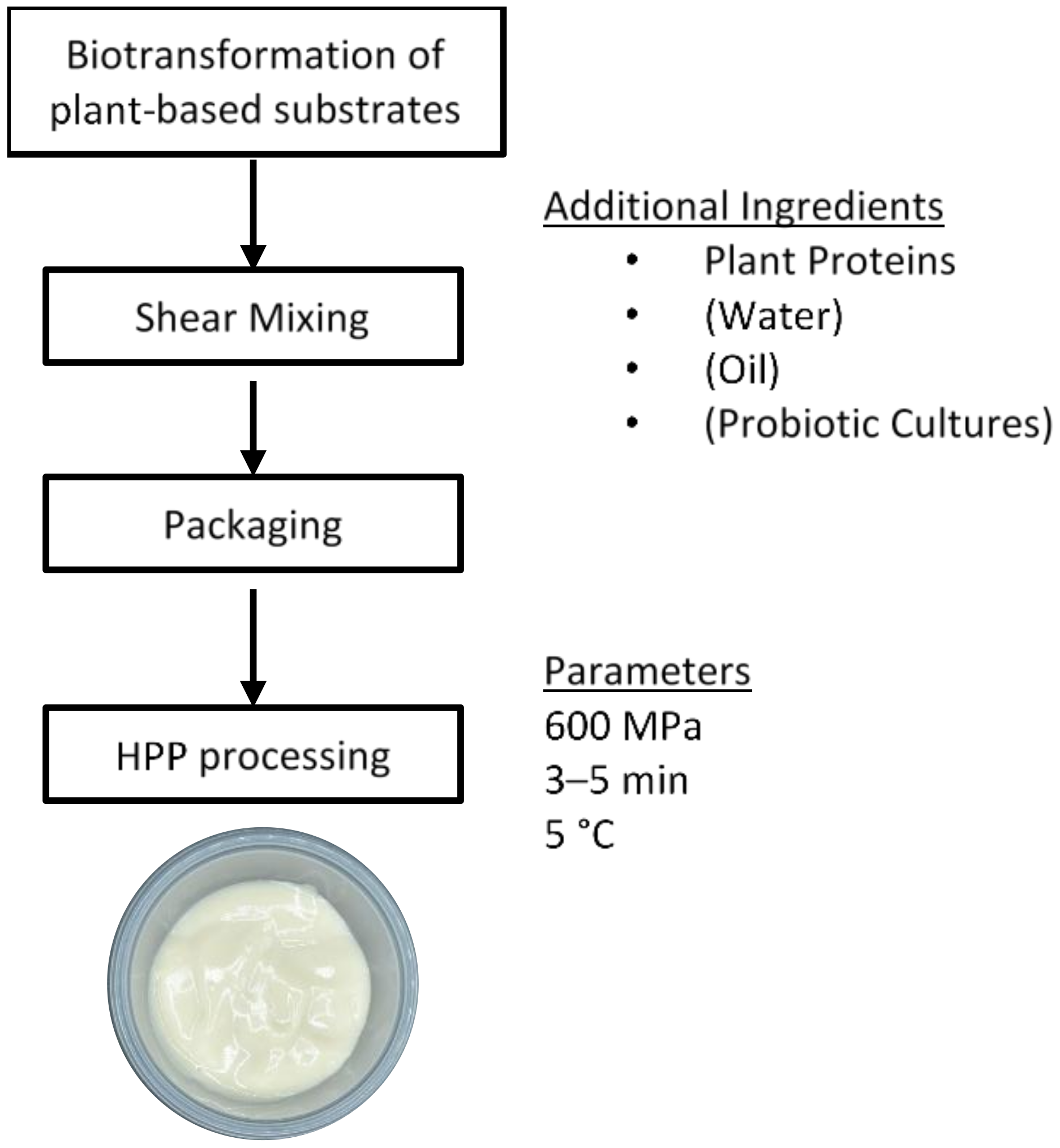
- Sim, S.Y.J.; Hua, X.Y.; Henry, C.J. A novel approach to structure plant-based yogurts using high pressure processing. Foods 2020, 9, 1126. [Google Scholar] [CrossRef]
- Tan, S.-Y.; Peh, E.; Siow, P.C.; Henry, C.J. Chapter 17—The role of ethylcellulose oleogel in human health and its potential applications. In Edible Oleogels, 2nd ed.; Marangoni, A.G., Garti, N., Eds.; AOCS Press: Urbana, IL, USA, 2018; pp. 401–414. [Google Scholar] [CrossRef]
- Tan, S.-Y.; Wan-Yi Peh, E.; Marangoni, A.G.; Henry, C.J. Effects of liquid oil vs. oleogel co-ingested with a carbohydrate-rich meal on human blood triglycerides, glucose, insulin and appetite. Food Funct. 2017, 8, 241–249. [Google Scholar] [CrossRef]
- Tan, S.-Y.; Peh, E.; Siow, P.C.; Marangoni, A.G.; Henry, C.J. Effects of the physical-form and the degree-of-saturation of oil on postprandial plasma triglycerides, glycemia and appetite of healthy Chinese adults. Food Funct. 2017, 8, 4433–4440. [Google Scholar] [CrossRef]
- Tan, S.-Y.; Peh, E.; Lau, E.; Marangoni, A.G.; Henry, C.J. Physical form of dietary fat alters postprandial substrate utilization and glycemic response in healthy Chinese men. J. Nutr. 2017, 147, 1138–1144. [Google Scholar] [CrossRef]
- Quek, R.Y.; Peh, E.W.; Henry, C.J. Effects of cocoa butter and cocoa butter equivalent in a chocolate confectionery on human blood triglycerides, glucose and insulin. Foods 2020, 9, 455. [Google Scholar] [CrossRef] [PubMed]
- Co, E.D.; Marangoni, A.G. Chapter 1—Oleogels: An introduction. In Edible Oleogels, 2nd ed,; Marangoni, A.G., Garti, N., Eds.; AOCS Press: Urbana, IL, USA, 2018; pp. 1–29. [Google Scholar] [CrossRef]
- Feichtinger, A.; Scholten, E. Preparation of protein oleogels: Effect on structure and functionality. Foods 2020, 9, 1745. [Google Scholar] [CrossRef] [PubMed]
- Van Ba, H.; Touseef, A.; Jeong, D.; Hwang, I. Principle of meat aroma flavors and future prospect. In Latest Research into Quality Control; Akyar, I., Ed.; Intech Open Access Publisher: London, UK, 2012. [Google Scholar]
- Reineccius, G. Changes in food flavor due to processing. In Flavor Chemistry and Technology, 2nd ed.; Reineccius, G., Ed.; CRC Press: Boca Raton, USA, 2005; pp. 517–558. [Google Scholar]
- Kerler, J.; Winkel, C.; Davidek, T.; Blank, I. Basic chemistry and process conditions for reaction flavours with particular focus on Maillard-type reactions. In Food Flavour Technology; Taylor, A.J., Linforth, R.S.T., Eds.; Blackwell Publishing Ltd.: West Sussex, UK, 2010; Volume 2, pp. 51–88. [Google Scholar]
- De Roos, K.B. Meat flavor generation from cysteine and sugars. In ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1992. [Google Scholar]
- Zhou, X.; Cui, H.; Zhang, Q.; Hayat, K.; Yu, J.; Hussain, S.; Tahir, M.U.; Zhang, X.; Ho, C.-T. Taste improvement of Maillard reaction intermediates derived from enzymatic hydrolysates of pea protein. Food Res. Int. 2021, 140, 109985. [Google Scholar] [CrossRef]
- Lotfy, S.N.; Saad, R.; El-Massrey, K.F.; Fadel, H.H.M. Effects of pH on headspace volatiles and properties of Maillard reaction products derived from enzymatically hydrolyzed quinoa protein-xylose model system. LWT 2021, 145, 111328. [Google Scholar] [CrossRef]
- Wei, C.-K.; Ni, Z.-J.; Thakur, K.; Liao, A.-M.; Huang, J.-H.; Wei, Z.-J. Color and flavor of flaxseed protein hydrolysates Maillard reaction products: Effect of cysteine, initial pH, and thermal treatment. Int. J. Food Prop. 2019, 22, 84–99. [Google Scholar] [CrossRef]
- Fadel, H.H.; Samad, A.A.; Kobeasy, M.; Mageed, M.A.A.; Lotfy, S.N. Flavour quality and stability of an encapsulated meat-like process flavouring prepared from soybean based acid hydrolyzed protein. Int. J. Food Process. Technol. 2015, 2, 17–25. [Google Scholar] [CrossRef]
- Yu, M.; He, S.; Tang, M.; Zhang, Z.; Zhu, Y.; Sun, H. Antioxidant activity and sensory characteristics of Maillard reaction products derived from different peptide fractions of soybean meal hydrolysate. Food Chem. 2018, 243, 249–257. [Google Scholar] [CrossRef]
- Marinangeli, C.P.F.; House, J.D. Potential impact of the digestible indispensable amino acid score as a measure of protein quality on dietary regulations and health. Nutr. Rev. 2017, 75, 658–667. [Google Scholar] [CrossRef]
- Protein quality evaluation. Joint FAO/WHO. FAO Food Nutr. Pap. 1991, 51, 1–66.
- Berrazaga, I.; Micard, V.; Gueugneau, M.; Walrand, S. The role of the anabolic properties of plant- versus animal-based protein sources in supporting muscle mass maintenance: A critical review. Nutrients 2019, 11, 1825. [Google Scholar] [CrossRef]
- Phillips, S.M. Current concepts and unresolved questions in dietary protein requirements and supplements in adults. Front. Nutr. 2017, 4, 13. [Google Scholar] [CrossRef] [PubMed]
- Gilani, G.S.; Cockell, K.A.; Sepehr, E. Effects of antinutritional factors on protein digestibility and amino acid availability in foods. J. AOAC Int. 2005, 88, 967–987. [Google Scholar] [CrossRef] [PubMed]
- Carbonaro, M.; Maselli, P.; Nucara, A. Relationship between digestibility and secondary structure of raw and thermally treated legume proteins: A Fourier transform infrared (FT-IR) spectroscopic study. Amino Acids 2012, 43, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Duodu, K.; Taylor, J.; Belton, P.; Hamaker, B. Factors affecting sorghum protein digestibility. J. Cereal Sci. 2003, 38, 117–131. [Google Scholar] [CrossRef]
- Boirie, Y.; Dangin, M.; Gachon, P.; Vasson, M.P.; Maubois, J.L.; Beaufrère, B. Slow and fast dietary proteins differently modulate postprandial protein accretion. Proc. Natl. Acad. Sci. USA 1997, 94, 14930–14935. [Google Scholar] [CrossRef] [PubMed]
- Dangin, M.; Guillet, C.; Garcia-Rodenas, C.; Gachon, P.; Bouteloup-Demange, C.; Reiffers-Magnani, K.; Fauquant, J.; Ballèvre, O.; Beaufrère, B. The rate of protein digestion affects protein gain differently during aging in humans. J. Physiol. 2003, 549, 635–644. [Google Scholar] [CrossRef]
- Tang, J.E.; Moore, D.R.; Kujbida, G.W.; Tarnopolsky, M.A.; Phillips, S.M. Ingestion of whey hydrolysate, casein, or soy protein isolate: Effects on mixed muscle protein synthesis at rest and following resistance exercise in young men. J. Appl. Physiol. 2009, 107, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Churchward-Venne, T.A.; Burd, N.A.; Breen, L.; Tarnopolsky, M.A.; Phillips, S.M. Myofibrillar protein synthesis following ingestion of soy protein isolate at rest and after resistance exercise in elderly men. Nutr. Metab. 2012, 9, 57. [Google Scholar] [CrossRef]
- Suryawan, A.; Orellana, R.A.; Fiorotto, M.L.; Davis, T.A. Triennial Growth Symposium: Leucine acts as a nutrient signal to stimulate protein synthesis in neonatal pigs. J. Anim. Sci. 2011, 89, 2004–2016. [Google Scholar] [CrossRef]
- Norton, L.E.; Layman, D.K.; Bunpo, P.; Anthony, T.G.; Brana, D.V.; Garlick, P.J. The leucine content of a complete meal directs peak activation but not duration of skeletal muscle protein synthesis and mammalian target of rapamycin signaling in rats. J. Nutr. 2009, 139, 1103–1109. [Google Scholar] [CrossRef]
- Gorissen, S.H.; Horstman, A.M.; Franssen, R.; Crombag, J.J.; Langer, H.; Bierau, J.; Respondek, F.; van Loon, L.J. Ingestion of wheat protein increases in vivo muscle protein synthesis rates in healthy older men in a randomized trial. J. Nutr. 2016, 146, 1651–1659. [Google Scholar] [CrossRef] [PubMed]
- Engelen, M.P.; Rutten, E.P.; De Castro, C.L.; Wouters, E.F.; Schols, A.M.; Deutz, N.E. Supplementation of soy protein with branched-chain amino acids alters protein metabolism in healthy elderly and even more in patients with chronic obstructive pulmonary disease. Am. J. Clin. Nutr. 2007, 85, 431–439. [Google Scholar] [CrossRef]
- Hertzler, S.R.; Lieblein-Boff, J.C.; Weiler, M.; Allgeier, C. Plant proteins: Assessing their nutritional quality and effects on health and physical function. Nutrients 2020, 12, 3704. [Google Scholar] [CrossRef] [PubMed]
- Day, L. Proteins from land plants—Potential resources for human nutrition and food security. Trends Food Sci. Technol. 2013, 32, 25–42. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food | PDCAAS | DIAAS | Limiting Amino Acids |
|---|---|---|---|
| Soy protein isolate | 0.98 | 0.90 | Met + Cys |
| Pea protein isolate | 0.89 | 0.82 | Met + Cys |
| Rice protein concentrate | 0.42 | 0.37 | Lys |
| Cooked Peas | 0.60 | 0.58 | Met + Cys |
| Cooked Rice | 0.62 | 0.59 | Lys |
| Almonds | 0.39 | 0.40 | Lys |
| Chickpeas | 0.74 | 0.83 | Met + Cys |
| Tofu | 0.56 | 0.52 | Met + Cys |
| Whole Milk | 1.00 | 1.14 | Met + Cys |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sim, S.Y.J.; SRV, A.; Chiang, J.H.; Henry, C.J. Plant Proteins for Future Foods: A Roadmap. Foods 2021, 10, 1967. https://doi.org/10.3390/foods10081967
Sim SYJ, SRV A, Chiang JH, Henry CJ. Plant Proteins for Future Foods: A Roadmap. Foods. 2021; 10(8):1967. https://doi.org/10.3390/foods10081967
Chicago/Turabian StyleSim, Shaun Yong Jie, Akila SRV, Jie Hong Chiang, and Christiani Jeyakumar Henry. 2021. "Plant Proteins for Future Foods: A Roadmap" Foods 10, no. 8: 1967. https://doi.org/10.3390/foods10081967
APA StyleSim, S. Y. J., SRV, A., Chiang, J. H., & Henry, C. J. (2021). Plant Proteins for Future Foods: A Roadmap. Foods, 10(8), 1967. https://doi.org/10.3390/foods10081967