
Casein Micelles as an Emerging Delivery System for Bioactive Food Components




Abstract
:1. Introduction
2. Casein Micelles and Its Structure
2.1. Casein Interactions
2.1.1. Self-Association of Caseins
2.1.2. Interactions with Other Caseins
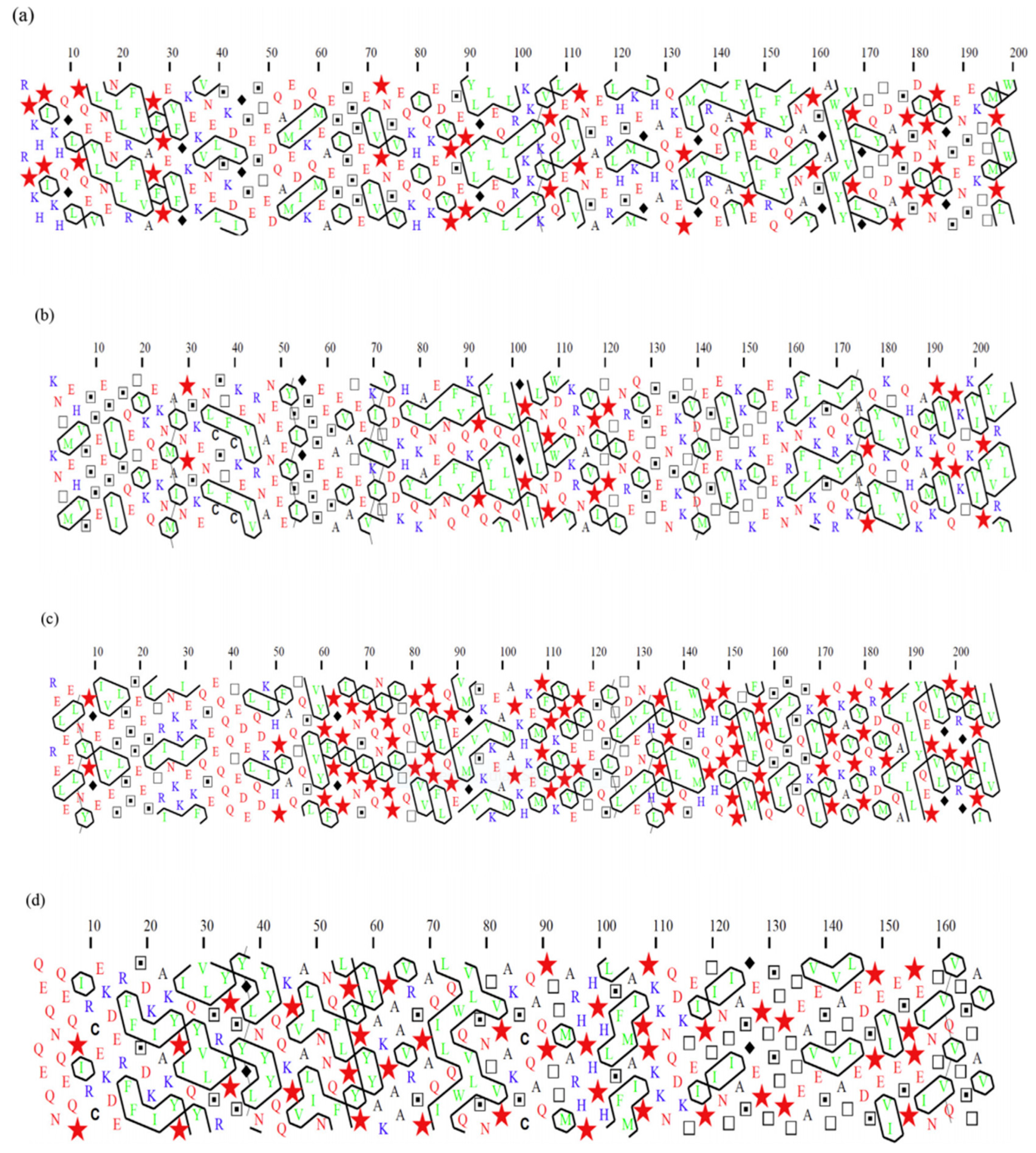
2.1.3. Hydrophobic Clustering of Caseins
2.1.4. Casein–Mineral Interactions
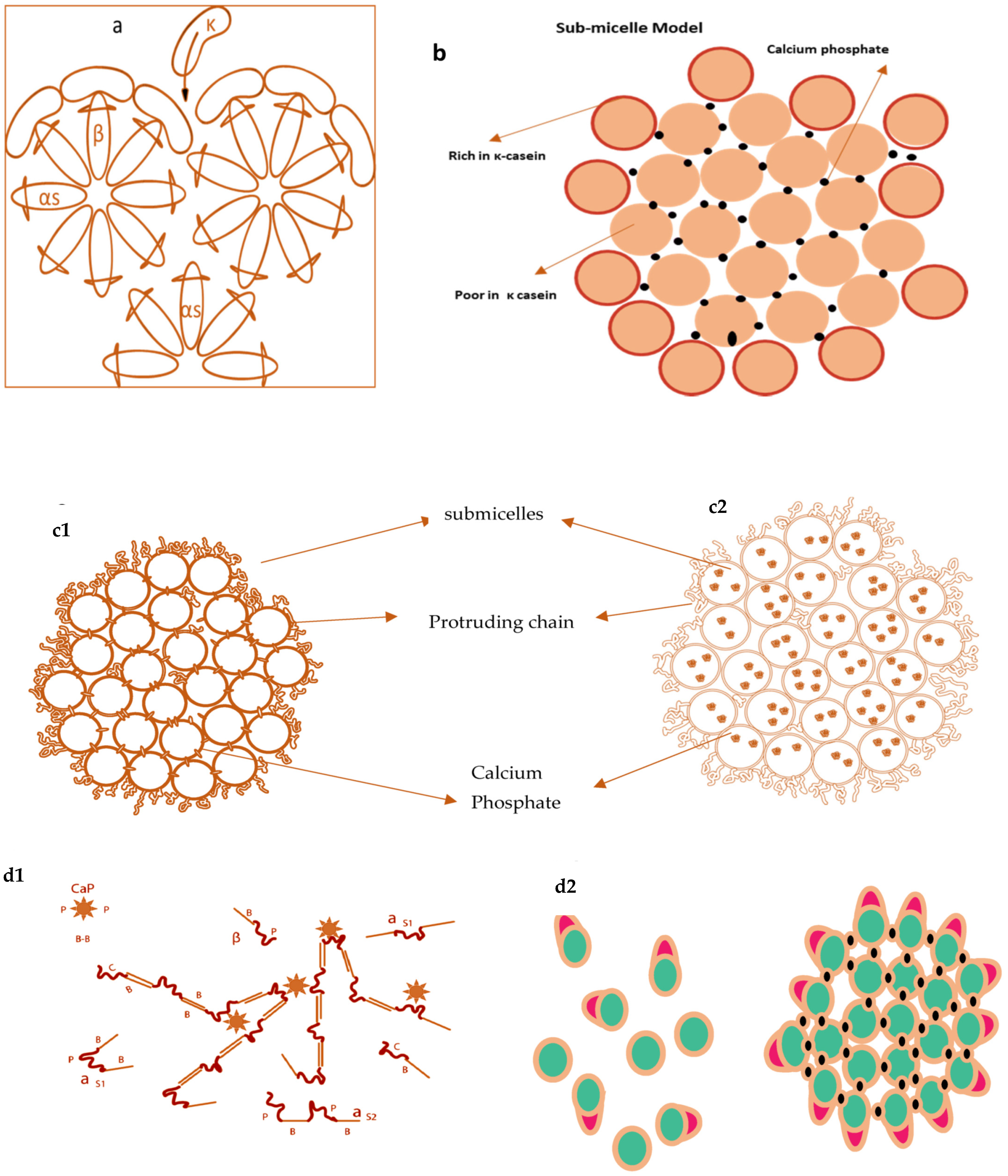
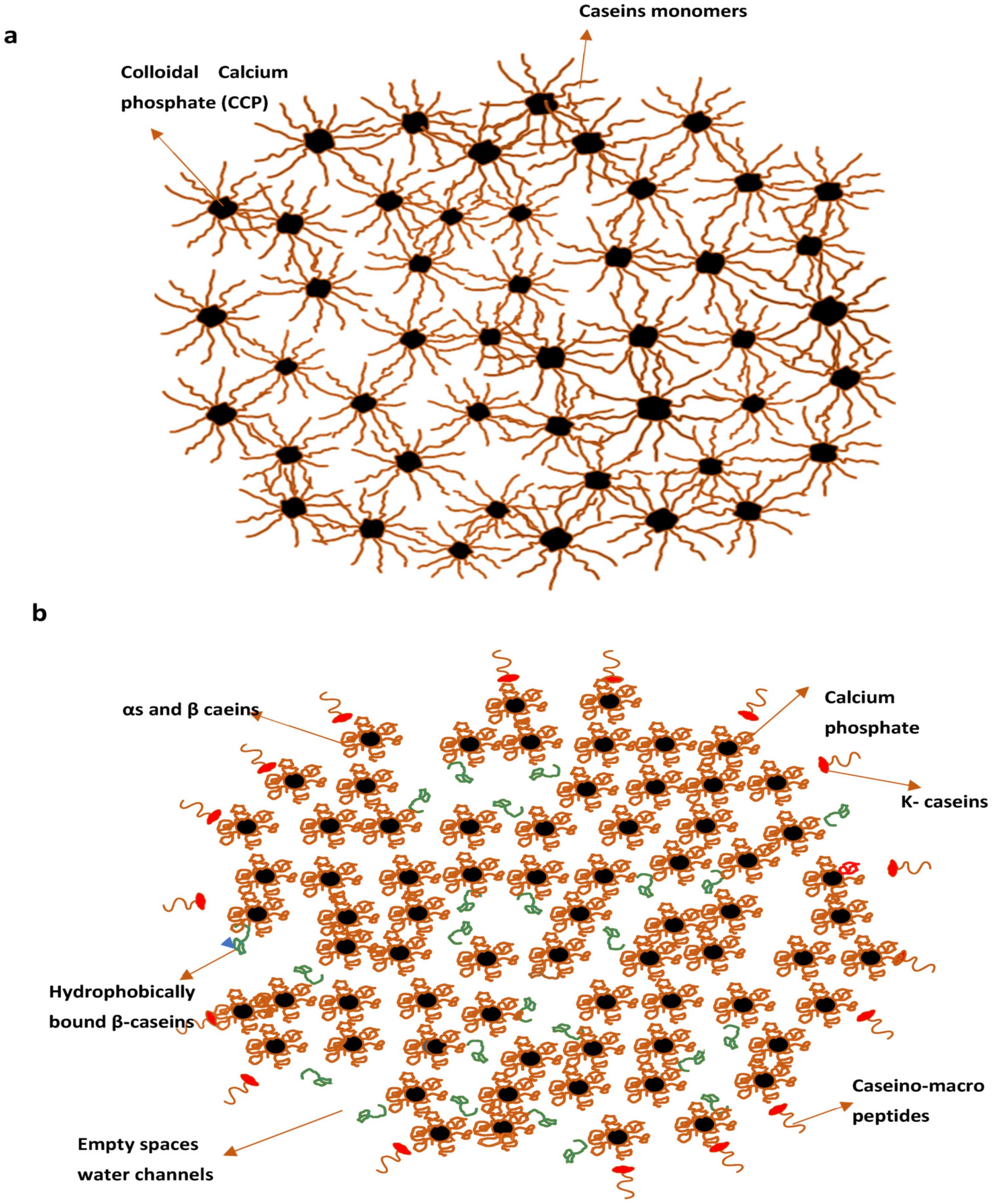
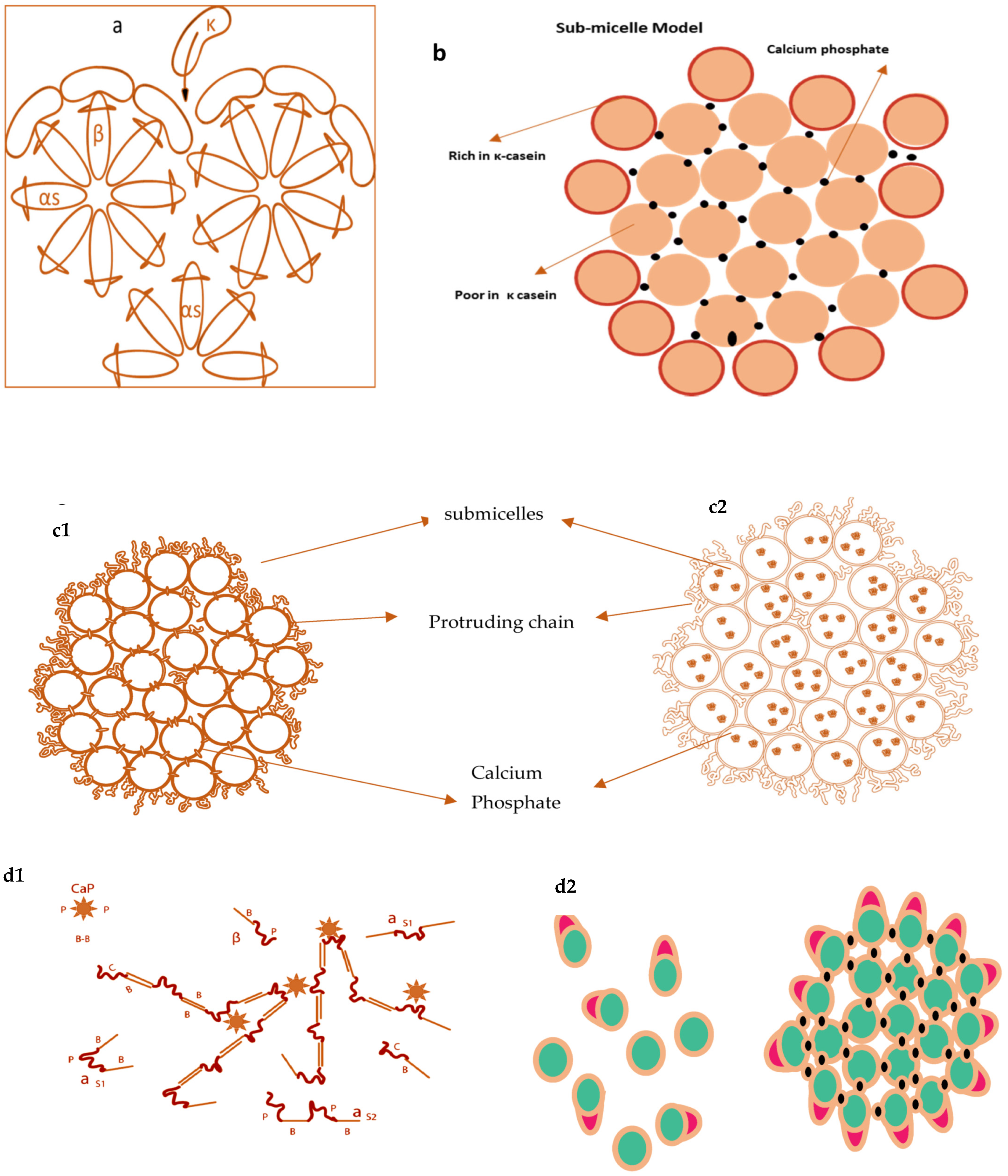
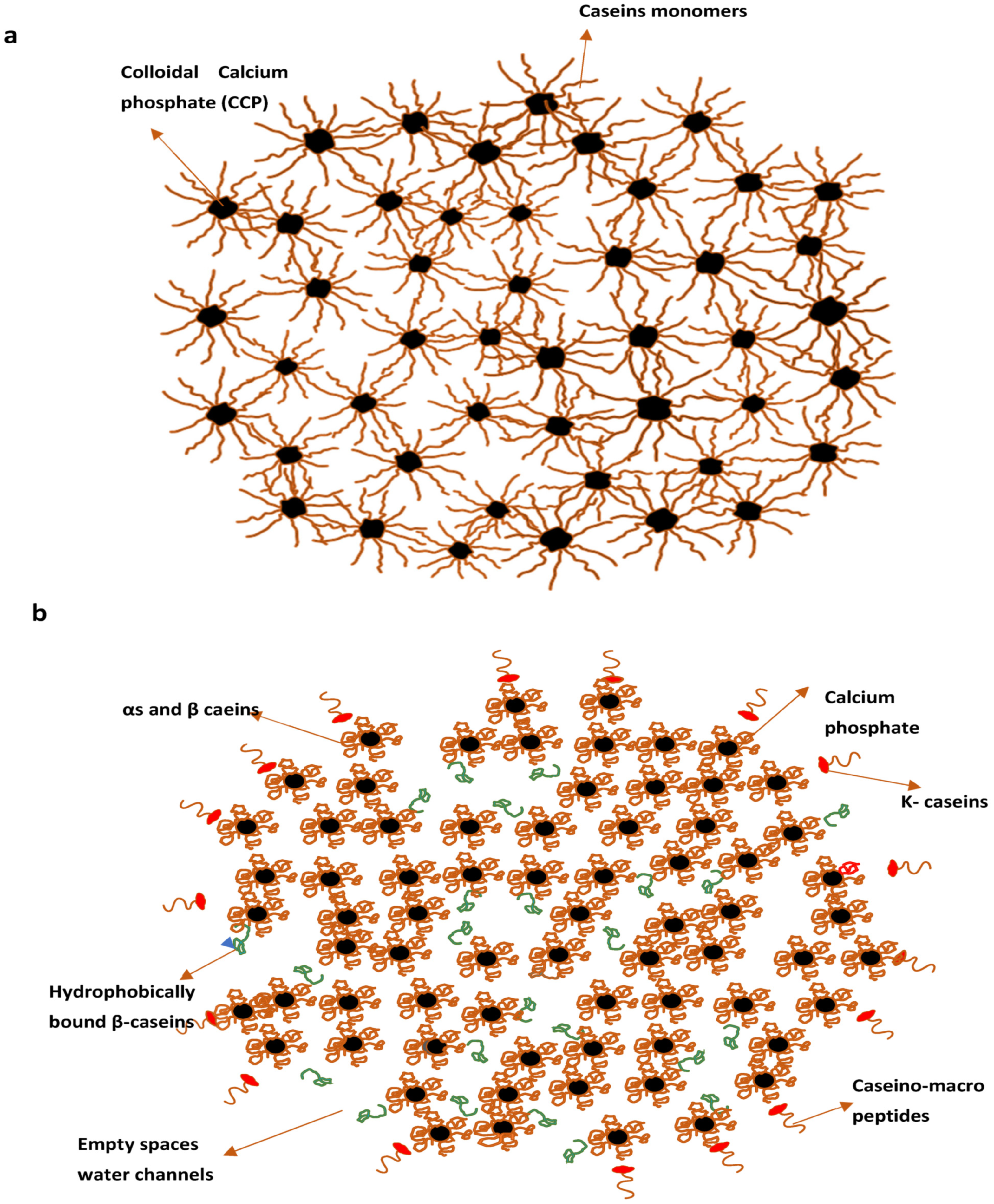
2.1.5. Models of Casein Micelles
3. Factors Affecting Techno-Functionalities of Casein Micelles
3.1. Effect of Temperature on Techno-Functionality of Casein Micelles
3.2. Effect of pH on Techno-Functionality of Casein Micelles
4. Casein Micelles–Based Delivery Systems
4.1. Caseins as Micro and Nanoparticles
4.2. Caseins and Nano Emulsions
4.3. Casein-Based Complexes as Delivery Systems
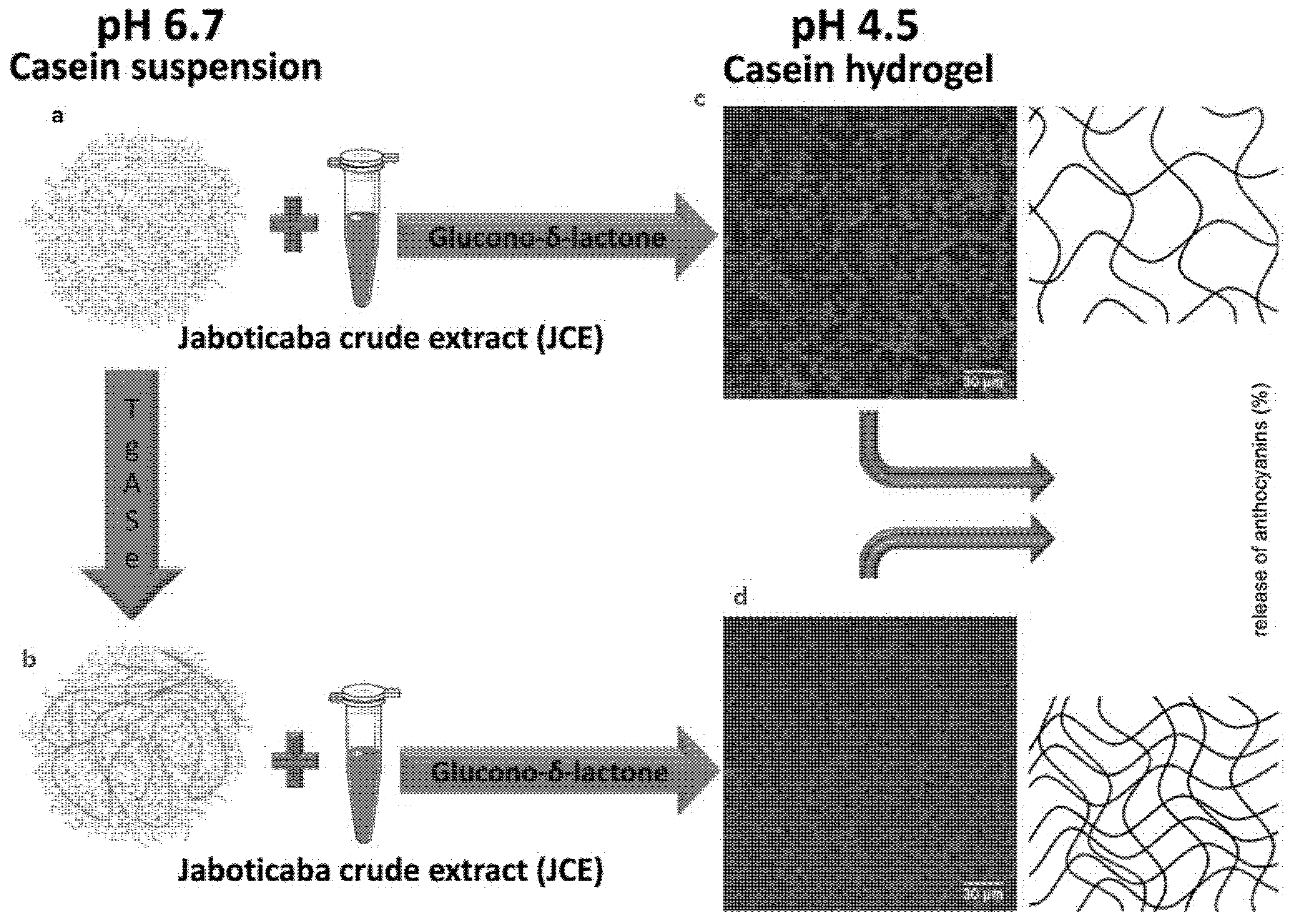
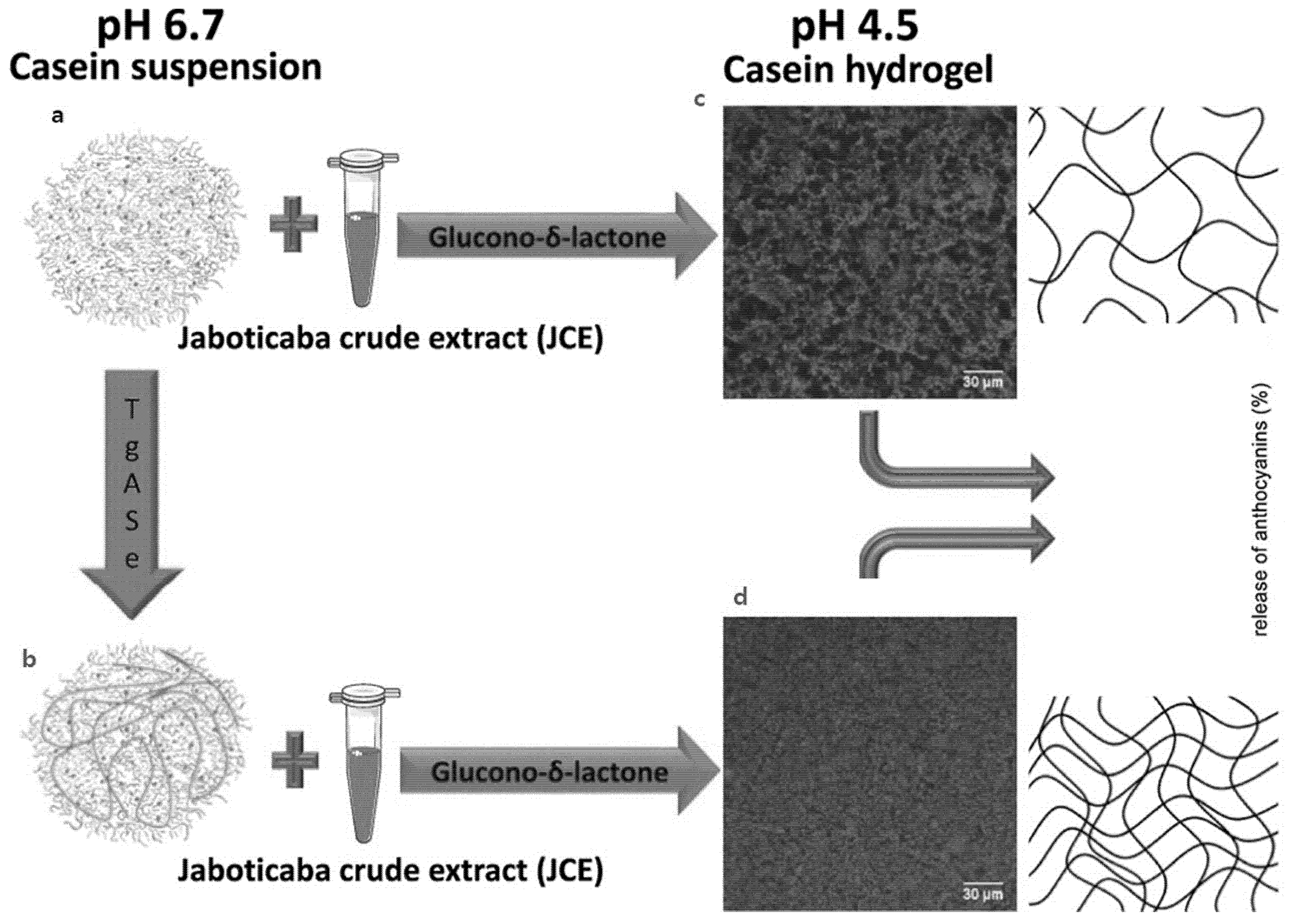
4.4. Casein Micelles as Hydrogels
5. Conclusions and Future Challenges
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gleeson, J.P.; Ryan, S.M.; Brayden, D. Oral delivery strategies for nutraceuticals: Delivery vehicles and absorption enhancers. Trends Food Sci. Technol. 2016, 53, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.S. Bioactive compounds and their neuroprotective effects in diabetic complications. Nutrients 2016, 8, 472. [Google Scholar] [CrossRef] [Green Version]
- Santiago, L.G.; Castro, G.R. Novel technologies for the encapsulation of bioactive food compounds. Curr. Opin. Food Sci. 2016, 7, 78–85. [Google Scholar] [CrossRef]
- De Vos, P.; Faas, M.M.; Spasojevic, M.; Sikkema, J. Encapsulation for preservation of functionality and targeted delivery of bioactive food components. Int. Dairy J. 2010, 20, 292–302. [Google Scholar] [CrossRef]
- Benshitrit, R.C.; Levi, C.S.; Tal, S.L.; Shimoni, E.; Lesmes, U. Development of oral food-grade delivery systems: Current knowledge and future challenges. Food Funct. 2012, 3, 10–21. [Google Scholar] [CrossRef] [PubMed]
- McClements, D.J.; Li, Y. Structured emulsion-based delivery systems: Controlling the digestion and release of lipophilic food components. Adv. Colloid Interface Sci. 2010, 159, 213–228. [Google Scholar] [CrossRef]
- McClements, D.J.; Decker, E.A.; Weiss, J. Emulsion-based delivery systems for lipophilic bioactive components. J. Food Sci. 2007, 72, 109–124. [Google Scholar] [CrossRef] [PubMed]
- Kosaraju, S.L. Colon targeted delivery systems: Review of polysaccharides for encapsulation and delivery. Crit. Rev. Food Sci. Nutr. 2005, 45, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, E. Interfacial structure and stability of food emulsions as affected by protein-polysaccharide interactions. Soft Matter 2008, 4, 932–942. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Y.; Remondetto, G.E.; Subirade, M. Food proteinbased materials as nutraceutical delivery systems. Trends Food Sci. Technol. 2006, 17, 272–283. [Google Scholar] [CrossRef]
- Tavares, G.M.; Croguennec, T.; Carvalho, A.F.; Bouhallab, S. Milk proteins as encapsulation devices and delivery vehicles: Applications and trends. Trend Food Sci. Technol. 2014, 37, 5–20. [Google Scholar] [CrossRef]
- Martins, J.T.; Bourbon, A.I.; Pinheiro, A.C.; Fasolin, L.H.; Vicente, A.A. Protein-based structures for food applications: From macro to nanoscale. Front. Sustain. Food Syst. 2018, 2, 77. [Google Scholar] [CrossRef]
- Rehan, F.; Ahemad, N.; Gupta, M. Casein nanomicelle as an emerging biomaterial—A comprehensive review. Colloids Surf. B Biointerfaces 2019, 179, 280–292. [Google Scholar] [CrossRef]
- Semo, E.; Kesselman, E.; Danino, D.; Livney, Y. Casein micelle as a natural nano-capsular vehicle for nutraceuticals. Food Hydrocoll. 2007, 21, 936–942. [Google Scholar] [CrossRef]
- Boisgard, R.; Chanat, E. Phospholipase D-dependent and -independent mechanisms are involved in milk protein secretion in rabbit mammary epithelial cells. Biochim. Biophys. Acta Mol. Cell Res. 2000, 1495, 281–296. [Google Scholar] [CrossRef] [Green Version]
- Broyard, C.; Gaucheron, F. Modifications of structures and functions of caseins: A scientific and technological challenge. Dairy Sci. Technol. 2015, 95, 831–862. [Google Scholar] [CrossRef]
- Gebhardt, R.; Doster, W.; Friedrich, J.; Kulozik, U. Size distribution of pressure-decomposed casein micelles studied by dynamic light scattering and AFM. Eur. Biophys. J. 2006, 35, 503–509. [Google Scholar] [CrossRef]
- Dalgleish, D.G. On the structural models of bovine casein micelles—Review and possible improvements. Soft Matter 2011, 7, 2265–2272. [Google Scholar] [CrossRef]
- Ye, R.; Harte, F. Casein maps: Effect of ethanol, pH, temperature, and CaCl2 on the particle size of reconstituted casein micelles. J. Dairy Sci. 2013, 96, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Juliano, P.; Williams, R.P.W.; Niere, J.; Augustin, M.A. Ultrasound effects on the assembly of casein micelles in reconstituted skim milk. J. Dairy Res. 2014, 81, 146–155. [Google Scholar] [CrossRef]
- Tuinier, R.; de Kruif, C.G. Stability of casein micelles in milk. J. Chem. Phys. 2002, 117, 1290–1295. [Google Scholar] [CrossRef] [Green Version]
- Suárez-Luque, S.; Mato, I.; Huidobro, J.F.; Simal-Lozano, J. Determination of major metal cations in milk by capillary zone electrophoresis. Int. Dairy J. 2007, 17, 896–901. [Google Scholar] [CrossRef]
- Livney, Y.D. Milk proteins as vehicles for bioactives. Curr. Opin. Colloid Interface Sci. 2010, 15, 73–83. [Google Scholar] [CrossRef]
- Liu, C.; Yao, W.; Zhang, L.; Qian, H.; Wu, W.; Jiang, X. Cell-penetrating hollow spheres based on milk protein. Chem. Commun. 2010, 46, 7566–7568. [Google Scholar] [CrossRef]
- Głąb, T.K.; Boratyński, J. Potential of Casein as a Carrier for Biologically Active Agents. Top. Curr. Chem. 2017, 375, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, Y.; Ish-Shalom, S.; Segal, E.; Nudelman, O.; Shpigelman, A.; Livney, Y.D. The bioavailability of vitamin D3, a model hydrophobic nutraceutical, in casein micelles, as model protein nanoparticles: Human clinical trial results. J. Funct. Foods 2017, 30, 321–325. [Google Scholar] [CrossRef]
- Ghayour, N.; Hosseini, S.M.H.; Eskandari, M.H.; Esteghlal, S.; Nekoei, A.-R.; Hashemi Gahruie, H.; Tatar, M.; Naghibalhossaini, F. Nanoencapsulation of quercetin and curcumin in casein-based delivery systems. Food Hydrocoll. 2019, 87, 394–403. [Google Scholar] [CrossRef]
- Nascimento, L.G.L.; Casanova, F.; Silva, N.F.N.; Teixeira, Á.V.N.d.C.; Júnior, P.P.d.S.P.; Vidigal, M.C.T.R.; Stringheta, P.C.; Carvalho, A.F. Use of a crosslinked casein micelle hydrogel as a carrier for jaboticaba (Myrciaria cauliflora) extract. Food Hydrocoll. 2020, 106, 105872. [Google Scholar] [CrossRef]
- Marreto, R.N.; Ramos, M.F.S.; Silva, E.J.; de Freitas, O.; de Freitas, L.A.P. Impact of Cross-linking and Drying Method on Drug Delivery Performance of Casein–Pectin Microparticles. AAPS PharmSciTech 2013, 14, 1227–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nascimento, L.G.L.; Casanova, F.; Silva, N.F.N.; Teixeira, A.V.N.C.; Carvalho, A.F. Casein-based hydrogels: A mini-review. Food Chem. 2020, 314, 126063. [Google Scholar] [CrossRef] [PubMed]
- Ranadheera, C.; Liyanaarachchi, W.; Chandrapala, J.; Dissanayake, M.; Vasiljevic, T. Utilizing unique properties of caseins and the casein micelle for delivery of sensitive food ingredients and bioactives. Trends Food Sci. Technol. 2016, 57, 178–187. [Google Scholar] [CrossRef]
- McSweeney, P.L.; Fox, P.F. Advanced Dairy Chemistry; Springer: Boston, MA, USA, 2003; Volume 1. [Google Scholar]
- McSweeney, P.L.; Fox, P.F. Advanced Dairy Chemistry: Proteins: Basic Aspects, 3rd ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2013; Volume 1A. [Google Scholar]
- Holt, C.; Carver, J.A.; Ecroyd, H.; Thorn, D.C. Invited review: Caseins and the casein micelle: Their biological functions, structures, and behavior in foods. J. Dairy Sci. 2013, 96, 6127–6146. [Google Scholar] [CrossRef]
- De Kruif, C.G.; Huppertz, T. Casein Micelles: Size Distribution in Milks from Individual Cows. J. Agric. Food Chem. 2012, 60, 4649–4655. [Google Scholar] [CrossRef] [Green Version]
- Huppertz, T.; Gazi, I.; Luyten, H.; Nieuwenhuijse, H.; Alting, A.; Schokker, E. Hydration of casein micelles and caseinates: Implications for casein micelle structure. Int. Dairy J. 2017, 74, 1–11. [Google Scholar] [CrossRef]
- Swaisgood, H.E. Chemistry of the caseins. In Advanced Dairy Chemistry—1 Proteins; Springer: Boston, MA, USA, 2003; pp. 139–201. [Google Scholar]
- Farrell, H.M.; Malin, E.L.; Brown, E.M.; Qi, P.X. Casein micelle structure: What can be learned from milk synthesis and structural biology? Curr. Opin. Colloid Interface Sci. 2006, 11, 135–147. [Google Scholar] [CrossRef]
- Treweek, T.M. Alpha-casein as a molecular chaperone. Milk Proteins 2012, 85–119. [Google Scholar]
- Shapira, A.; Assaraf, Y.G.; Livney, Y.D. Beta-casein nanovehicles for oral delivery of chemotherapeutic drugs. Nanomed. Nanotechnol. Biol. Med. 2010, 6, 119–126. [Google Scholar] [CrossRef] [PubMed]
- De Kruif, C.G.; Zhulina, E.B. κ-casein as a polyelectrolyte brush on the surface of casein micelles. Colloids Surf. A Physicochem. Eng. Asp. 1996, 117, 151–159. [Google Scholar] [CrossRef]
- Dalgleish, D.G.; Corredig, M. The Structure of the Casein Micelle of Milk and Its Changes During Processing. Annu. Rev. Food Sci. Technol. 2012, 3, 449–467. [Google Scholar] [CrossRef]
- Farrell, H.M.; Jimenez-Flores, R.; Bleck, G.T.; Brown, E.M.; Butler, J.E.; Creamer, L.K.; Hicks, C.L.; Hollar, C.M.; Ng-Kwai-Hang, K.F.; Swaisgood, H.E. Nomenclature of the Proteins of Cows’ Milk—Sixth Revision. J. Dairy Sci. 2004, 87, 1641–1674. [Google Scholar] [CrossRef] [Green Version]
- Creamer, L.K.; Plowman, J.E.; Liddell, M.J.; Smith, M.H.; Hill, J.P. Micelle Stability: κ-Casein Structure and Function. J. Dairy Sci. 1998, 81, 3004–3012. [Google Scholar] [CrossRef]
- Huppertz, T.; Fox, P.F.; Kelly, A.L. The caseins: Structure, stability, and functionality. In Proteins in Food Processing; Yada, R.Y., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 49–92. [Google Scholar]
- Horne, D.S. A balanced view of casein interactions. Curr. Opin. Colloid Interface Sci. 2017, 28, 74–86. [Google Scholar] [CrossRef]
- Schmidt, D.G. Differences between the association of the genetic variants B, C and D of αs1-casein. Biochim. Biophys. Acta Protein 1970, 221, 140–142. [Google Scholar] [CrossRef]
- Von Hippel, P.H.; Waugh, D.F. Casein: Monomers and Polymers1. J. Am. Chem. Soc. 1955, 77, 4311–4319. [Google Scholar] [CrossRef]
- Schmidt, D.G.; Payens, T.A. The evaluation of positive and negative contributions to the second virial coefficient of some milk proteins. J. Colloid Interface Sci. 1972, 39, 655–662. [Google Scholar] [CrossRef]
- Schmidt, D.G. The association of αs1-casein B at pH 6.6. Biochim. Biophys. Acta Protein Struct. 1970, 207, 130–138. [Google Scholar] [CrossRef]
- Swaisgood, H.E.; Timasheff, S.N. Association of αs-casein C in the alkaline pH range. Arch. Biochem. Biophys. 1968, 125, 344–361. [Google Scholar] [CrossRef]
- Malin, E.L.; Brown, E.M.; Wickham, E.D.; Farrell, H.M. Contributions of Terminal Peptides to the Associative Behavior of αs1-Casein. J. Dairy Sci. 2005, 88, 2318–2328. [Google Scholar] [CrossRef] [Green Version]
- Snoeren, T.H.M.; Van Markwijk, B.; Van Montfort, R. Some physicochemical properties of bovine αs2-casein. Biochim. Biophys. Protein Struct. 1980, 622, 268–276. [Google Scholar] [CrossRef]
- Thorn, D.C.; Ecroyd, H.; Sunde, M.; Poon, S.; Carver, J.A. Amyloid Fibril Formation by Bovine Milk αs2-Casein Occurs under Physiological Conditions Yet Is Prevented by Its Natural Counterpart, αs1-Casein. Biochemistry 2008, 47, 3926–3936. [Google Scholar] [CrossRef]
- O’Connell, J.; Grinberg, V.Y.; De Kruif, C. Association behavior of β-casein. J. Colloid Interface Sci. 2003, 258, 33–39. [Google Scholar] [CrossRef]
- Qi, P.X.; Wickham, E.D.; Piotrowski, E.G.; Fagerquist, C.K.; Farrell Jr, H.M. Implication of C-Terminal Deletion on the Structure and Stability of Bovine β-casein. Protein J. 2005, 24, 431–444. [Google Scholar] [CrossRef]
- Mikheeva, L.M.; Grinberg, N.V.; Grinberg, V.Y.; Khokhlov, A.R.; de Kruif, C.G. Thermodynamics of Micellization of Bovine β-Casein Studied by High-Sensitivity Differential Scanning Calorimetry. Langmuir 2003, 19, 2913–2921. [Google Scholar] [CrossRef]
- Swaisgood, H.E.; Brunner, J.R.; Lillevik, H.A. Physical Parameters of K-Casein from Cow’s Milk. Biochemistry 1964, 3, 1616–1623. [Google Scholar] [CrossRef]
- Vreeman, H.J.; Brinkhuis, J.A.; Van Der Spek, C.A. Some association properties of bovine SH-k-casein. Biophys. Chem. 1981, 14, 185–193. [Google Scholar] [CrossRef]
- Vreeman, H.J.; Visser, S.; Slangen, C.J.; Van Riel, J.A. Characterization of bovine kappa-casein fractions and the kinetics of chymosin-induced macropeptide release from carbohydrate-free and carbohydrate-containing fractions determined by high-performance gel-permeation chromatography. J. Biochem. 1986, 240, 87–97. [Google Scholar] [CrossRef]
- Ecroyd, H.; Thorn, D.C.; Liu, Y.; Carver, J.A. The dissociated form of κ-casein is the precursor to its amyloid fibril formation. J. Biochem. 2010, 429, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Leonil, J.; Henry, G.; Jouanneau, D.; Delage, M.-M.; Forge, V.; Putaux, J.-L. Kinetics of Fibril Formation of Bovine κ-Casein Indicate a Conformational Rearrangement as a Critical Step in the Process. J. Mol. Biol. 2008, 381, 1267–1280. [Google Scholar] [CrossRef]
- Creamer, L.; Berry, G.; Mills, O. A study of the dissociation of b-casein from the bovine casein micelle at low temperature [milk and cream]. N. Z. J. Dairy Sci. Technol. 1977, 12, 58–66. [Google Scholar]
- Bigelow, C.C. On the average hydrophobicity of proteins and the relation between it and protein structure. J. Theor. Biol. 1967, 16, 187–211. [Google Scholar] [CrossRef]
- Arunachalam, J.; Gautham, N. Hydrophobic clusters in protein structures. Proteins 2008, 71, 2012–2202. [Google Scholar] [CrossRef] [PubMed]
- Callebaut, I.; Labesse, G.; Durand, P.; Poupon, A.; Canard, L.; Chomilier, J.; Henrissat, B.; Mornon, J.P. Deciphering protein sequence information through hydrophobic cluster analysis (HCA): Current status and perspectives. Cell. Mol. Life Sci. 1997, 53, 621–645. [Google Scholar] [CrossRef] [PubMed]
- Farrell, H., Jr.; Qi, P.; Uversky, V. New views of protein structure: Applications to the caseins: Protein structure and functionality. In Advances in Biopolymers; Fishman, M.L., Qi, P.X., Wicker, L., Eds.; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2006; Volume 935, pp. 52–70. [Google Scholar]
- Farrell, H.M.; Brown, E.M.; Malin, E.L. Higher Order Structures of the Caseins: A Paradox? In Advanced Dairy Chemistry, 3rd ed.; Fox, P.F., McSweeney, P.L.H., Eds.; Proteins Part A; Kluwer Academic: New York, NY, USA, 2012; Volume 1, pp. 203–231. [Google Scholar]
- Dickson, I.R.; Perkins, D.J. Studies on the interactions between purified bovine caseins and alkaline-earth-metal ions. Biochemistry 1971, 124, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Lucey, J.A.; Horne, D.S. Perspectives on casein interactions. Int. Dairy J. 2018, 85, 56–65. [Google Scholar] [CrossRef]
- Holt, C.; de Kruif, C.G.; Tuinier, R.; Timmins, P.A. Substructure of bovine casein micelles by small-angle X-ray and neutron scattering. Colloids Surf. A Physicochem. Eng. Asp. 2003, 213, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, J.K.; Jakobsen, H.J.; Nielsen, N.C.; Petersen, T.E.; Rasmussen, L.K. Solid-state Magic-angle Spinning 31P-NMR Studies of Native Casein Micelles. Eur. J. Biochem. 1995, 230, 454–459. [Google Scholar] [CrossRef]
- Aoki, T.; Toyooka, K.; Kako, Y. Role of phosphate groups in the calcium sensitivity of αS2-casein. J. Dairy Sci. 1985, 68, 1624–1629. [Google Scholar] [CrossRef]
- Van Kemenade, M.; De Bruyn, P.L. The influence of casein on the kinetics of hydroxyapatite precipitation. J. Colloid Interface Sci. 1989, 129, 1–14. [Google Scholar] [CrossRef]
- Holt, C. Casein micelle substructure and calcium phosphate interactions studied by sephacryl column chromatography. J. Dairy Sci. 1998, 81, 2994–3003. [Google Scholar] [CrossRef]
- Waugh, D.F. The interactions of αs-β- and κ-caseins in micelle formation. Discuss. Faraday Soc. 1958, 25, 186–192. [Google Scholar] [CrossRef]
- Horne, D.S. Casein structure, self-assembly and gelation. Curr. Opin. Colloid Interface Sci. 2002, 7, 456–461. [Google Scholar] [CrossRef]
- Walstra, P. Casein sub-micelles: Do they exist? Int. Dairy J. 1999, 9, 189–192. [Google Scholar] [CrossRef]
- Holt, C. Structure and stability of bovine casein micelles. Adv. Protein Chem. 1992, 43, 63–151. [Google Scholar]
- Horne, D.S. Casein micelles as hard spheres: Limitations of the model in acidified gel formation. Colloids Surf. A Physicochem. Eng. Asp. 2003, 213, 255–263. [Google Scholar] [CrossRef]
- Horne, D.S. Casein micelle structure: Models and muddles. Curr. Opin. Colloid Interface Sci. 2006, 11, 148–153. [Google Scholar] [CrossRef]
- De Kruif, C.G.; Huppertz, T.; Urban, V.S.; Petukhov, A.V. Casein micelles and their internal structure. Adv. Colloid Interface Sci. 2012, 171–172, 36–52. [Google Scholar] [CrossRef]
- McMahon, D.J.; Oommen, B.S. Casein Micelle Structure, Functions, and Interactions. In Advanced Dairy Chemistry, 2nd ed.; Fox, P., McSweeney, L., Eds.; Proteins Basic Aspects 1A; Springer: New York, NY, USA, 2012; pp. 185–210. [Google Scholar]
- Gaucheron, F.; MollÉ, D.; Pannetier, R. Influence of pH on the heat-induced proteolysis of casein molecules. J. Dairy Res. 2001, 68, 71–80. [Google Scholar] [CrossRef]
- Davies, D.T.; White, J.C.D. The use of ultrafiltration and dialysis in isolating the aqueous phase of milk and in determining the partition of milk constituents between the aqueous and disperse phases. J. Dairy Res. 1960, 27, 171–190. [Google Scholar] [CrossRef]
- Van Boekel, M.A.J.S. Heat-induced deamidation, dephosphorylation and breakdown of caseinate. Int. Dairy J. 1999, 9, 237–241. [Google Scholar] [CrossRef]
- Koutina, G.; Knudsen, J.C.; Andersen, U.; Skibsted, L.H. Temperature effect on calcium and phosphorus equilibria in relation to gel formation during acidification of skim milk. Int. Dairy J. 2014, 36, 65–73. [Google Scholar] [CrossRef]
- Liu, D.Z.; Weeks, M.G.; Dunstan, D.E.; Martin, G.J.O. Temperature-dependent dynamics of bovine casein micelles in the range 10–40 °C. Food Chem. 2013, 141, 4081–4086. [Google Scholar] [CrossRef] [PubMed]
- Van Hekken, D.L.; Holsinger, V.H. Use of cold microfiltration to produce unique β-casein enriched milk gels. Le Lait 2000, 80, 69–76. [Google Scholar] [CrossRef]
- Corzo-Martínez, M.; Moreno, F.J.; Villamiel, M.; Harte, F.M. Characterization and improvement of rheological properties of sodium caseinate glycated with galactose, lactose and dextran. Food Hydrocoll. 2010, 24, 88–97. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, H.; Cucheval, A.; Coker, C.; Patel, H.; Carr, A.; Bennett, R. Effect of lactosylation on plasmin-induced hydrolysis of β-casein. Int. Dairy J. 2014, 38, 213–218. [Google Scholar] [CrossRef]
- McCarthy, N.A.; Kelly, A.L.; O’Mahony, J.A.; Fenelon, M.A. The physical characteristics and emulsification properties of partially dephosphorylated bovine β-casein. Food Chem. 2013, 138, 1304–1311. [Google Scholar] [CrossRef] [PubMed]
- Tezcucano Molina, A.C.; Alli, I.; Konishi, Y.; Kermasha, S. Effect of dephosphorylation on bovine casein. Food Chem. 2007, 101, 1263–1271. [Google Scholar] [CrossRef]
- De Kruif, C.G. Skim Milk Acidification. J. Colloid Interface Sci. 1997, 185, 19–25. [Google Scholar] [CrossRef] [PubMed]
- McMahon, D.J.; Du, H.; McManus, W.R.; Larsen, K.M. Microstructural changes in casein supramolecules during acidification of skim milk. J. Dairy Sci. 2010, 93, 1783. [Google Scholar] [CrossRef]
- Dalgleish, D.G.; Law, A.J.R. pH-Induced dissociation of bovine casein micelles. II. Mineral solubilization and its relation to casein release. J. Dairy Res. 1989, 56, 727–735. [Google Scholar] [CrossRef]
- Silva, N.N.; Piot, M.; de Carvalho, A.F.; Violleau, F.; Fameau, A.-L.; Gaucheron, F. pH-induced demineralization of casein micelles modifies their physico-chemical and foaming properties. Food Hydrocoll. 2013, 32, 322–330. [Google Scholar] [CrossRef]
- Ahmad, S.; Piot, M.; Rousseau, F.; Grongnet, J.F.; Gaucheron, F. Physico-chemical changes in casein micelles of buffalo and cow milks as a function of alkalinisation. Dairy Sci. Technol. 2009, 89, 387–403. [Google Scholar] [CrossRef] [Green Version]
- Huppertz, T.; Vaia, B.; Smiddy, M.A. Reformation of casein particles from alkaline-disrupted casein micelles. J. Dairy Res. 2008, 75, 44–47. [Google Scholar] [CrossRef] [Green Version]
- Silva, N.F.N.; Saint-Jalmes, A.; de Carvalho, A.F.; Gaucheron, F. Development of Casein Microgels from Cross-Linking of Casein Micelles by Genipin. Langmuir 2014, 30, 10167–10175. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Shi, Y.; Wang, P.; Liu, H.; Wen, P.; Ren, F. Effect of succinylation on the functional properties of yak caseins: A comparison with cow caseins. Dairy Sci. Technol. 2014, 94, 359–372. [Google Scholar] [CrossRef]
- Miwa, N.; Yokoyama, K.; Wakabayashi, H.; Nio, N. Effect of deamidation by protein-glutaminase on physicochemical and functional properties of skim milk. Int. Dairy J. 2010, 20, 393–399. [Google Scholar] [CrossRef]
- Miwa, N.; Nio, N.; Sonomoto, K. Effect of enzymatic deamidation by protein-glutaminase on the textural and microstructural properties of set yoghurt. Int. Dairy J. 2014, 36, 1–5. [Google Scholar] [CrossRef]
- Huppertz, T.; de Kruif, C.G. Disruption and reassociation of casein micelles during high pressure treatment: Influence of whey proteins. J. Dairy Res. 2007, 74, 194–197. [Google Scholar] [CrossRef]
- Huppertz, T.; Fox, P.F.; Kelly, A.L. High pressure treatment of bovine milk: Effects on casein micelles and whey proteins. J. Dairy Res. 2004, 71, 97–106. [Google Scholar] [CrossRef]
- Menéndez-Aguirre, O.; Kessler, A.; Stuetz, W.; Grune, T.; Weiss, J.; Hinrichs, J. Increased loading of vitamin D 2 in reassembled casein micelles with temperature-modulated high pressure treatment. Food Res. Int. 2014, 64, 74–80. [Google Scholar] [CrossRef]
- Menéndez-Aguirre, O.; Stuetz, W.; Grune, T.; Kessler, A.; Weiss, J.; Hinrichs, J. High pressure-assisted encapsulation of vitamin D2 in reassembled casein micelles. High Press. Res. 2011, 31, 265–274. [Google Scholar] [CrossRef]
- Medina, A.L.; Colas, B.; Le Meste, M.; Renaudet, I.; Lorient, D. Physicochemical and dynamic properties of caseins modified by chemical phosphorylation. J. Food Sci. 1992, 57, 617–621. [Google Scholar] [CrossRef]
- Moon, J.-H.; Hong, Y.-H.; Huppertz, T.; Fox, P.F.; Kelly, A.L. Properties of casein micelles cross-linked by transglutaminase. Int. J. Dairy Technol. 2009, 62, 27–32. [Google Scholar] [CrossRef]
- Huppertz, T.; de Kruif, C.G. Structure and stability of nanogel particles prepared by internal cross-linking of casein micelles. Int. Dairy J. 2008, 18, 556–565. [Google Scholar] [CrossRef]
- Madadlou, A.; Mousavi, M.E.; Emam-djomeh, Z.; Ehsani, M.; Sheehan, D. Sonodisruption of re-assembled casein micelles at different pH values. Ultrason. Sonochem. 2009, 16, 644–648. [Google Scholar] [CrossRef]
- Madadlou, A.; Emam-Djomeh, Z.; Mousavi, M.E.; Mohamadifar, M.; Ehsani, M. Acid-induced gelation behavior of sonicated casein solutions. Ultrason. Sonochem. 2010, 17, 153–158. [Google Scholar] [CrossRef]
- Zheng, L.I.U.; Juliano, P.; Williams, R.P.W.; Niere, J.; Augustin, M.A. Ultrasound improves the renneting properties of milk. Ultrason. Sonochem. 2014, 21, 2131–2137. [Google Scholar]
- Cases, E.; Vidal, V.; Cuq, J.L. Effect of K-Casein Deglycosylation on the Acid Coagulability of Milk. J. Food Sci. 2003, 68, 2406–2410. [Google Scholar] [CrossRef]
- Jacobsen, J.; Wind, S.L.; Rasholt, E.L.; van den Brink, J.M. N-Glycosidase F improves gel firmness in fermented milk products. Int. Dairy J. 2014, 38, 169–173. [Google Scholar] [CrossRef]
- Mekmene, O.; Le Graët, Y.; Gaucheron, F. Theoretical Model for Calculating Ionic Equilibria in Milk as a Function of pH: Comparison to Experiment. J. Agric. Food Chem. 2010, 58, 4440–4447. [Google Scholar] [CrossRef]
- Omoarukhe, E.D.; On-Nom, N.; Grandison, A.S.; Lewis, M.J. Effects of different calcium salts on properties of milk related to heat stability. Int. J. Dairy Technol. 2010, 63, 504–511. [Google Scholar] [CrossRef]
- Vidal, V.; Marchesseau, S.; Cuq, J.L. Physicochemical Properties of Acylated Casein Micelles in Milk. J. Food Sci. 2002, 67, 42–47. [Google Scholar] [CrossRef]
- Wang, J.; Su, Y.; Jia, F.; Jin, H. Characterization of casein hydrolysates derived from enzymatic hydrolysis. Chem. Cent. J. 2013, 7, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Pan, K.; Zhong, Q. Physical, chemical and biochemical properties of casein hydrolyzed by three proteases: Partial characterizations. Food Chem. 2014, 155, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Huppertz, T.; Fox, P.F. Effect of NaCl on some physico-chemical properties of concentrated bovine milk. Int. Dairy J. 2006, 16, 1142–1148. [Google Scholar] [CrossRef]
- Hussain, R.; Gaiani, C.; Aberkane, L.; Scher, J. Characterization of high-milk-protein powders upon rehydration under various salt concentrations. J. Dairy Sci. 2011, 94, 14–23. [Google Scholar] [CrossRef]
- De Kort, E.; Minor, M.; Snoeren, T.; van Hooijdonk, T.; van der Linden, E. Effect of calcium chelators on physical changes in casein micelles in concentrated micellar casein solutions. Int. Dairy J. 2011, 21, 907–913. [Google Scholar] [CrossRef]
- Schokker, E.P.; Church, J.S.; Mata, J.P.; Gilbert, E.P.; Puvanenthiran, A.; Udabage, P. Reconstitution properties of micellar casein powder: Effects of composition and storage. Int. Dairy J. 2011, 21, 877–886. [Google Scholar] [CrossRef]
- Haratifar, S.; Corredig, M. Interactions between tea catechins and casein micelles and their impact on renneting functionality. Food Chem. 2014, 143, 27–32. [Google Scholar] [CrossRef]
- Ouanezar, M.; Guyomarc’h, F.; Bouchoux, A. AFM Imaging of Milk Casein Micelles: Evidence for Structural Rearrangement upon Acidification. Langmuir 2012, 28, 4915–4919. [Google Scholar] [CrossRef]
- Marchin, S.; Putaux, J.-L.; Pignon, F.; Leonil, J. Effects of the environmental factors on the casein micelle structure studied by Cryo transmission microscopy and small-angle X-ray scattering/ultrasmall-angle X-ray scattering. J. Chem. Phys. 2007, 126, 045101. [Google Scholar] [CrossRef] [PubMed]
- Vaia, B.; Smiddy, M.A.; Kelly, A.L.; Huppertz, T. Solvent-Mediated Disruption of Bovine Casein Micelles at Alkaline pH. J. Agric. Food Chem. 2006, 54, 8288–8293. [Google Scholar] [CrossRef]
- Pan, K.; Luo, Y.; Gan, Y.; Baek, S.J.; Zhong, Q.J.S.M. pH-driven encapsulation of curcumin in self-assembled casein nanoparticles for enhanced dispersibility and bioactivity. Soft Matter 2014, 10, 6820–6830. [Google Scholar] [CrossRef] [PubMed]
- De Kruif, C.; Anema, S.G.; Zhu, C.; Havea, P.; Coker, C. Water holding capacity and swelling of casein hydrogels. Food Hydrocoll. 2015, 44, 372–379. [Google Scholar] [CrossRef]
- Song, F.; Zhang, L.-M.; Shi, J.-F.; Li, N.-N. Novel casein hydrogels: Formation, structure and controlled drug release. Colloids Surf. B Biointerfaces 2010, 79, 142–148. [Google Scholar] [CrossRef]
- Bonnaillie, L.; Zhang, H.; Akkurt, S.; Yam, K.; Tomasula, P. Casein Films: The Effects of Formulation, Environmental Conditions and the Addition of Citric Pectin on the Structure and Mechanical Properties. Polymers 2014, 6, 2018–2036. [Google Scholar] [CrossRef]
- Lesmes, U.; Sandra, S.; Decker, E.A.; McClements, D.J. Impact of surface deposition of lactoferrin on physical and chemical stability of omega-3 rich lipid droplets stabilised by caseinate. Food Chem. 2010, 123, 99–106. [Google Scholar] [CrossRef]
- Yang, M.; Wei, Y.; Ashokkumar, M.; Qin, J.; Han, N.; Wang, Y. Effect of ultrasound on binding interaction between emodin and micellar casein and its microencapsulation at various temperatures. Ultrason. Sonochem. 2020, 62, 104861. [Google Scholar] [CrossRef]
- Singh Chauhan, P.; Abutbul Ionita, I.; Moshe Halamish, H.; Sosnik, A.; Danino, D. Multidomain drug delivery systems of β-casein micelles for the local oral co-administration of antiretroviral combinations. J. Colloid Interface Sci. 2021, 592, 156–166. [Google Scholar] [CrossRef]
- Tan, S.; Ebrahimi, A.; Langrish, T. Controlled release of caffeine from tablets of spray-dried casein gels. Food Hydrocoll. 2019, 88, 13–20. [Google Scholar] [CrossRef]
- Tan, S.; Ebrahimi, A.; Langrish, T. Smart release-control of microencapsulated ingredients from milk protein tablets using spray drying and heating. Food Hydrocoll. 2019, 92, 181–188. [Google Scholar] [CrossRef]
- Cheng, H.; Dong, H.; Liang, L. A comparison of β-casein complexes and micelles as vehicles for trans-/cis-resveratrol. Food Chem. 2020, 330, 127209. [Google Scholar] [CrossRef]
- Consoli, L.; Dias, R.A.; Rabelo, R.S.; Furtado, G.F.; Sussulini, A.; Cunha, R.L.; Hubinger, M.D. Sodium caseinate-corn starch hydrolysates conjugates obtained through the Maillard reaction as stabilizing agents in resveratrol-loaded emulsions. Food Hydrocoll. 2018, 84, 458–472. [Google Scholar] [CrossRef]
- Gorji, E.G.; Rocchi, E.; Schleining, G.; Bender-Bojalil, D.; Furtmüller, P.G.; Piazza, L.; Iturri, J.J.; Toca-Herrera, J.L. Characterization of resveratrol–milk protein interaction. J. Food Eng. 2015, 167, 217–225. [Google Scholar] [CrossRef]
- Duerasch, A.; Herrmann, P.; Hogh, K.; Henle, T. Study on β-Casein Depleted Casein Micelles: Micellar Stability, Enzymatic Cross-Linking, and Suitability as Nanocarriers. J. Agric. Food Chem. 2020, 68, 13940–13949. [Google Scholar] [CrossRef]
- Xu, G.; Li, L.; Bao, X.; Yao, P. Curcumin, casein and soy polysaccharide ternary complex nanoparticles for enhanced dispersibility, stability and oral bioavailability of curcumin. Food Biosci. 2020, 35, 100569. [Google Scholar] [CrossRef]
- Perrechil, F.A.; Maximo, G.J.; Sato, A.C.K.; Cunha, R.L. Microbeads of Sodium Caseinate and κ-Carrageenan as a β-Carotene Carrier in Aqueous Systems. Food Bioprocess Technol. 2020, 13, 661–669. [Google Scholar] [CrossRef]
- Yi, J.; Lam, T.I.; Yokoyama, W.; Cheng, L.W.; Zhong, F. Beta-carotene encapsulated in food protein nanoparticles reduces peroxyl radical oxidation in Caco-2 cells. Food Hydrocoll. 2015, 43, 31–40. [Google Scholar] [CrossRef]
- Haham, M.; Ish-Shalom, S.; Nodelman, M.; Duek, I.; Segal, E.; Kustanovich, M.; Livney, Y.D. Stability and bioavailability of vitamin D nanoencapsulated in casein micelles. Food Funct. 2012, 3, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Levinson, Y.; Ish-Shalom, S.; Segal, E.; Livney, Y.D. Bioavailability, rheology and sensory evaluation of fat-free yogurt enriched with VD3 encapsulated in re-assembled casein micelles. Food Funct. 2016, 7, 1477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malekhosseini, P.; Alami, M.; Khomeiri, M.; Esteghlal, S.; Nekoei, A.R.; Hosseini, S.M.H. Development of casein-based nanoencapsulation systems for delivery of epigallocatechin gallate and folic acid. Food Sci. Nutr. 2019, 7, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Lee, J.; Han, S.S.; Oh, K.H.; Nam, K.T.; Sun, J.-Y. Highly Stretchable and Notch-Insensitive Hydrogel Based on Polyacrylamide and Milk Protein. ACS Appl. Mater. Interfaces 2016, 8, 29220–29226. [Google Scholar] [CrossRef]
- Sahu, A.; Kasoju, N.; Bora, U. Fluorescence study of the curcumin–casein micelle complexation and its application as a drug nanocarrier to cancer cells. Biomacromolecules 2008, 9, 2905–2912. [Google Scholar] [CrossRef] [PubMed]
- Rahimi Yazdi, S.; Corredig, M. Heating of milk alters the binding of curcumin to casein micelles. A fluorescence spectroscopy study. Food Chem. 2012, 132, 1143–1149. [Google Scholar] [CrossRef]
- Elzoghby, A.O.; Helmy, M.W.; Samy, W.M.; Elgindy, N.A. Micellar Delivery of Flutamide Via Milk Protein Nanovehicles Enhances its Anti-Tumor Efficacy in Androgen-Dependent Prostate Cancer Rat Model. Pharm. Res. 2013, 30, 2654–2663. [Google Scholar] [CrossRef]
- Elzoghby, A.O.; Helmy, M.W.; Samy, W.M.; Elgindy, N.A. Spray-dried casein-based micelles as a vehicle for solubilization and controlled delivery of flutamide: Formulation, characterization, and in vivo pharmacokinetics. Eur. J. Pharm. Biopharm. 2013, 84, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Khanji, A.N.; Michaux, F.; Petit, J.; Salameh, D.; Rizk, T.; Jasniewski, J.; Banon, S. Structure, gelation, and antioxidant properties of curcumin-doped casein micelle powder produced by spray-drying. Food Funct. 2018, 9, 971–981. [Google Scholar] [CrossRef]
- Khanji, A.N.; Michaux, F.; Salameh, D.; Rizk, T.; Banon, S.; Jasniewski, J. The study of curcumin interaction with micellar casein and lactic acid bacteria cell envelope. LWT 2018, 91, 293–302. [Google Scholar] [CrossRef]
- Chen, F.; Liang, L.; Zhang, Z.; Deng, Z.; Decker, E.A.; McClements, D.J. Inhibition of lipid oxidation in nanoemulsions and filled microgels fortified with omega-3 fatty acids using casein as a natural antioxidant. Food Hydrocoll. 2017, 63, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Loewen, A.; Chan, B.; Li-Chan, E.C. Optimization of vitamins A and D3 loading in re-assembled casein micelles and effect of loading on stability of vitamin D3 during storage. Food Chem. 2018, 240, 472–481. [Google Scholar] [CrossRef]
- Bahri, A.; Henriquet, C.; Pugnière, M.; Marchesseau, S.; Chevalier-Lucia, D. Binding analysis between monomeric β-casein and hydrophobic bioactive compounds investigated by surface plasmon resonance and fluorescence spectroscopy. Food Chem. 2019, 286, 289–296. [Google Scholar] [CrossRef]
- Ghasemi, S.; Abbasi, S. Formation of natural casein micelle nanocapsule by means of pH changes and ultrasound. Food Hydrocoll. 2014, 42, 42–47. [Google Scholar] [CrossRef]
- Burgain, J.; Scher, J.; Lebeer, S.; Vanderleyden, J.; Cailliez-Grimal, C.; Corgneau, M.; Francius, G.; Gaiani, C. Significance of bacterial surface molecules interactions with milk proteins to enhance microencapsulation of Lactobacillus rhamnosus GG. Food Hydrocoll. 2014, 41, 60–70. [Google Scholar] [CrossRef]
- Perlstein, H.; Bavli, Y.; Turovsky, T.; Rubinstein, A.; Danino, D.; Stepensky, D.; Barenholz, Y. Beta-casein nanocarriers of celecoxib for improved oral bioavailability. Eur. J. Nanomed. 2014, 6, 217–226. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.; Su, R.; Qi, W.; He, Z. A casein-polysaccharide hybrid hydrogel cross-linked by transglutaminase for drug delivery. J. Mater. Sci. 2012, 47, 2045–2055. [Google Scholar] [CrossRef]
- Mohan, M.S.; Jurat-Fuentes, J.L.; Harte, F. Binding of vitamin A by casein micelles in commercial skim milk. J. Dairy Sci. 2013, 96, 790–798. [Google Scholar] [CrossRef] [Green Version]
- Arranz, E.; Santoyo, S.; Jaime, L.; Fornari, T.; Reglero, G.; Guri, A.; Corredig, M. Improved Bioavailability of Supercritical Rosemary Extract Through Encapsulation in Different Delivery Systems after In Vitro Digestion. Food Dig. Res. Curr. Opin. 2015, 6, 30–37. [Google Scholar]
- Anema, S.G.; de Kruif, C.G. Lactoferrin binding to transglutaminase cross-linked casein micelles. Int. Dairy J. 2012, 26, 83–87. [Google Scholar] [CrossRef]
- Elzoghby, A.O.; Samy, W.M.; Elgindy, N.A. Novel Spray-Dried Genipin-Crosslinked Casein Nanoparticles for Prolonged Release of Alfuzosin Hydrochloride. Pharm. Res. 2013, 30, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Mirpoor, S.F.; Hosseini, S.M.H.; Yousefi, G.H. Mixed biopolymer nanocomplexes conferred physicochemical stability and sustained release behavior to introduced curcumin. Food Hydrocoll. 2017, 71, 216–224. [Google Scholar] [CrossRef]
- Cohen, Y.; Levi, M.; Lesmes, U.; Margier, M.; Reboul, E.; Livney, Y.D. Re-assembled casein micelles improve in vitro bioavailability of vitamin D in a Caco-2 cell model. Food Funct. 2017, 8, 2133–2141. [Google Scholar] [CrossRef] [PubMed]
- Haratifar, S.; Guri, A. Nanocapsule formation by caseins. In Nanoencapsulation Technologies for the Food and Nutraceutical Industries; Academic Press: San Diego, CA, USA, 2017; pp. 140–164. [Google Scholar]
- Haratifar, S.; Meckling, K.A.; Corredig, M. Bioefficacy of tea catechins encapsulated in casein micelles tested on a normal mouse cell line (4D/WT) and its cancerous counterpart (D/v-src) before and after in vitro digestion. Food Funct. 2014, 5, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Haratifar, S.; Meckling, K.A.; Corredig, M. Antiproliferative activity of tea catechins associated with casein micelles, using HT29 colon cancer cells. J. Dairy Sci. 2014, 97, 672–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimet, P.; Rosenberg, D.; Livney, Y.D. Re-assembled casein micelles and casein nanoparticles as nano-vehicles for ω-3 polyunsaturated fatty acids. Food Hydrocoll. 2011, 25, 1270–1276. [Google Scholar] [CrossRef]
- Moeller, H.; Martin, D.; Schrader, K.; Hoffmann, W.; Pargmann, S.; Kurz, J.; Lorenzen, P.C. Comparative studies of loading lipophilic substances into casein micelles and investigating the influence of whey proteins and heat treatment on loading stability. Int. J. Dairy Technol. 2018, 71, 954–965. [Google Scholar] [CrossRef]
- Sahlan, M.; Evelyn, K.; Pratami, D.K.; Mulia, K. In Vitro release study of sambiloto (Andrographis paniculata) extract encapsulated by casein micelle as anti-diabetic herbal drug. In Proceedings of the 5th International Symposium on Applied Chemistry 2019, ISAC 2019, Tangerang, Indonesia, 23–24 October 2019; American Institute of Physics Inc.: Melville, NY, USA, 2019; Volume 2175, p. 020042. [Google Scholar]
- Xu, H.; Lu, Y.; Zhang, T.; Liu, K.; Liu, L.; He, Z.; Xu, B.; Wu, X. Characterization of binding interactions of anthraquinones and bovine β-lactoglobulin. Food Chem. 2019, 281, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Liu, Z.; Hu, G.; Qu, L.; Yang, R. Investigation on the binding of aloe-emodin with tyrosinase by spectral analysis and molecular docking. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2019, 211, 79–85. [Google Scholar] [CrossRef]
- Madan, J.R.; Ansari, I.N.; Dua, K.; Awasthi, R. Formulation and In Vitro Evaluation of Casein Nanoparticles as Carrier for Celecoxib. Adv. Pharm. Bull. 2020, 10, 408. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, D.; Murphy, P.C. Surface active and emulsifying properties of caseins/caseinates as influenced by state of aggregation. Int. Dairy J. 1991, 1, 13–37. [Google Scholar] [CrossRef]
- Ries, D.; Ye, A.; Haisman, D.; Singh, H. Antioxidant properties of caseins and whey proteins in model oil-in-water emulsions. Int. Dairy J. 2010, 20, 72–78. [Google Scholar] [CrossRef]
- Zhang, Y.; He, S.; Li, Y.; Tang, H. The physical stability and digestibility of β-carotene in oil-in-water sodium caseinate nanoemulsion. J. Food Meas. Charact. 2017, 11, 864–871. [Google Scholar] [CrossRef]
- Mora-Gutierrez, A.; Attaie, R.; de Gonzalez, M.T.N.; Jung, Y.; Marquez, S.A. Interface Compositions as Determinants of Resveratrol Stability in Nanoemulsion Delivery Systems. Foods 2020, 9, 1394. [Google Scholar] [CrossRef]
- Li, N.-N.; Fu, C.-P.; Zhang, L.-M. Using casein and oxidized hyaluronic acid to form biocompatible composite hydrogels for controlled drug release. Mater. Sci. Eng. C 2014, 36, 287–293. [Google Scholar] [CrossRef]
- Zhang, F.; Cai, X.; Ding, L.; Wang, S. Effect of pH, ionic strength, chitosan deacetylation on the stability and rheological properties of O/W emulsions formulated with chitosan/casein complexes. Food Hydrocoll. 2021, 111, 106211. [Google Scholar] [CrossRef]
- Gallo, M.; Vinci, G.; Graziani, G.; De Simone, C.; Ferranti, P. The interaction of cocoa polyphenols with milk proteins studied by proteomic techniques. Food Res. Int. 2013, 54, 406–415. [Google Scholar] [CrossRef]
- Hasni, I.; Bourassa, P.; Hamdani, S.; Samson, G.; Carpentier, R.; Tajmir-Riahi, H.-A. Interaction of milk α- and β-caseins with tea polyphenols. Food Chem. 2011, 126, 630–639. [Google Scholar] [CrossRef]
- Baxter, N.J.; Lilley, T.H.; Haslam, E.; Williamson, M.P. Multiple Interactions between Polyphenols and a Salivary Proline-Rich Protein Repeat Result in Complexation and Precipitation. Biochemistry 1997, 36, 5566–5577. [Google Scholar] [CrossRef]
- Zhou, S.; Seo, S.; Alli, I.; Chang, Y.-W. Interactions of caseins with phenolic acids found in chocolate. Food Res. Int. 2015, 74, 177–184. [Google Scholar] [CrossRef]
- Yi, J.; Fan, Y.; Yokoyama, W.; Zhang, Y.; Zhao, L. Characterization of milk proteins–lutein complexes and the impact on lutein chemical stability. Food Chem. 2016, 200, 91–97. [Google Scholar] [CrossRef]
- Ali, A.; Ahmed, S. Recent Advances in Edible Polymer Based Hydrogels as a Sustainable Alternative to Conventional Polymers. J. Agric. Food Chem. 2018, 66, 6940–6967. [Google Scholar] [CrossRef]
- Abbate, V.; Kong, X.; Bansal, S.S. Photocrosslinked bovine serum albumin hydrogels with partial retention of esterase activity. Enzyme Microb. Technol. 2012, 50, 130–136. [Google Scholar] [CrossRef]
- Farran, A.J.E.; Teller, S.S.; Jha, A.K.; Jiao, T.; Hule, R.A.; Clifton, R.J.; Pochan, D.P.; Duncan, R.L.; Jia, X. Effects of matrix composition, microstructure, and viscoelasticity on the behaviors of vocal fold fibroblasts cultured in three-dimensional hydrogel networks. (Report). Tissue Eng. Part A 2010, 16, 1247–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casanova, F.; Nogueira Silva, N.F.; Gaucheron, F.; Nogueira, M.H.; Teixeira, A.V.N.C.; Perrone, I.T.; Alves, M.P.; Fidelis, P.C.; Carvalho, A.F. Stability of casein micelles cross-linked with genipin: A physicochemical study as a function of pH. Int. Dairy J. 2017, 68, 70–74. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Caseins Types | |||
|---|---|---|---|---|
| αS1 | αS2 | β | κ | |
| Natural conformation | Unfolded structure | Unfolded structure | Unfolded structure | Unfolded structure |
| Percentage in milk | 1.2–1.5 | 0.3–0.4 | 0.9–1.1 | 0.3–0.4 |
| Amino acid residues = Hydrophilic peptides + Hydrophobic regions | 199 = 63 + 136 | 207 = 118 + 89 | 209 = 42 + 167 | 169 = 64 + 105 |
| Molecular weight (Da) | 23,000 | 25,000 | 24,000 | 19,000 |
| Setting in milk at room temperature | Inside Micelle | Inside Micelle | Inside Micelle | Micelle surface |
| Number of proline resides/number of cysteine residues | 17/0 | 10/2 | 34–35/0 | 20/2 |
| No. of Phosphate groups | 8 | 10–13 | 5 | 1 |
| No. of S-S groups/No of S-H groups | 0/0 | 1/0 | 0/0 | 1/0 |
| Analyzed charge at pH 6.6 (mV) | (−21)–(−23.5) | (−12.2)–(−17.1) | (−11.8)–(−13.8) | (−2.0)–(−3.0) |
| primary structure IP versus IP after phosphorylation | 4.91/4.42 | 8.34/4.95 | 5.13/4.65 | 5.93/5.6 |
| Physical Methods | Biochemical Effect | Charge on Casein | Chemical Methods | Biochemical Effect | Charge on Casein | Enzymatic Methods | Biochemical Effect | Charge on Casein |
|---|---|---|---|---|---|---|---|---|
| Temperature (High) [42,84,86] Temperature (Low) [87,88] | Blockage of lysyl residues by lactose β-lactoglobulin covalent association Calcium phosphate precipitation and solubilisation Β casein solubilisation | Reduced negative charge Not determined | Reaction with sugar Glycation [90] Lactosylation [91] | Blockage of lysyl residues Blockage of lysyl residues | More negative More negative | Dephosphorylation [92,93] | organic phosphate removal from phosphoseryl residues | Reduced negativity |
| pH (Acid) [94,95,96,97] pH (alkaline) [98,99] | Protonation of casein Decrease of cations casein interactions Increase of the casein ionisation Insolubilisation of calcium phosphate | Reduced negativity More negative | Chemical Reticulation [100,101] | Blockage of lysyl residues | More negative | Deamidation [102,103] | - Release of ammonia from glutamine transformed into glutamic residues | More negative |
| Pressure [104,105,106,107] | Casein micelles distruptions | Not determined | Phosphorylation [108] | Reticulation [109,110] | Lysyl and glutamine crosslinking | Enhanced negativity | ||
| Ultrasound [20,111,112,113] | Casein micelles disruptions | Not determined | Glycosylation | Deglycosylation [114,115] | - Release of NANA | No effect | ||
| Addition of cations (di & trivalent) [116,117] | Direct association of added cation to casein Association of cation-inorganic phosphate to casein micelles Increase in ionic strength | Less negative | Succinylation [103] Acetylation [118] | Lysyl residues inhibition | More negative More negative | Proteolysis [119,120] | - Release of caseino macropeptide negatively charged between 106 to 169 peptides | Reduced negativity between 1–105 peptides |
| Adding salt [121,122] | Micellar calcium solubilisation Ionic strength enhancement | No change | ||||||
| Removal of diffusible ions | Diffusible ions removal | More negative ions | ||||||
| Calcium chelatants addition [123,124] | Casein and calcium association reductions Micellar calcium phosphate solubilisation | More negative ions | ||||||
| External ligands addition [125] | Hydrophobic and hydrogen interactions to caseins | ND |
| Casein Type | The Technique Used for Preparing Loaded Reassembled Casein Micelles | Bioactive | Encapsulation Mechanism | References |
|---|---|---|---|---|
| Micellar casein | • Casein–emodin complex formation by vortex • Heat and Ultrasound treatments • Spray-drying microencapsulation • In Vitro digestion evaluation | Emodin | Microencapsulation | [134] |
| β casein micelle | • Drug loaded β caseins dispersion • Freeze drying • Making and description of gastro-resistant Nanoparticle in Microparticle Delivery Systems • pH 2 and 6.5 • In Vitro drug release | Antiretroviral (ARV) combinations of Darunavir, efavirenz and ritonavir encapsulation in β caseins and further within Eudragit L100 | Co-encapsulation, Nanoparticle-in-microparticle delivery system (NiMDS) | [135] |
| Casein gels | • Casein gel production at pH 1 and 9 • Spray-dried gel and tablet • Oven-dried gel and tablets • Controlled release under various compression methods | Caffeine | Gels | [136,137] |
| β casein micelle Sodium Caseinate | • β casein preparation in 7.4 phosphate buffer • Blending of protein and resveratrol • Production of polysaccharide conjugates by Millard reaction Resveratrol loading at pH 7.5 | Resveratrol | Encapsulation Emulsions | [138,139,140] |
| β casein depleted Casein micelles | • Centrifugation • Lyophilisation • Mixing by shaker • Ultracentrifugation • Enzymatic crosslinking | Linoleic acid | Nanoencapsulation | [141] |
| Caseins | • Acidification • Homogenisation at high pressure • Curcumin/casein/soy polysaccharide complex at pH 10.0 • In Vitro digestion evaluation • CUR pharmacokinetics of CUR/CN/SSPS in mice | Curcumin | Nanoencapsulation | [142] |
| Casein Micelle | • Chemical acidification • Crosslinking by transglutaminase | Jaboticaba extract | Hydrogels | [28] |
| Sodium casienate/Carrageenan | • Primary and multilayered emulsion preparations • Microbeads preparation by gelation in an atomiser | β carotene | Emulsions/Gels | [143] |
| Casein micelles | • Mineral arrangement restoration and spray-drying • Homogenisation at high pressure • pH and temperature-induced opening | β carotene | Nanoencapsulation | [25,144,145,146] |
| Re-assembled casein micelles (r-CM) Sodium caseinate (CNP) | • Binding at pH 7.4 and temperature 74 °C • Centrifugation • EGGC binding r-CM and CNP • Encapsulation efficiency determination | Epigallocatechin gallate (EGGC), folic acid | Nanoencapsulation | [147] |
| Casein micelles | • Preparation of casein-PAAm hydrogels by free radical polymerisation | Polyacrylamide | Hydrogels | [148] |
| Casein micelles | • Spray-drying pH-shifting • High-pressure treatment | curcumin | Nanoencapsulation | [149,150,151,152,153,154] |
| Reassembled Casein micelles | • Restoration of mineral composition and ultrahigh-pressure homogenisation | Vitamin D3 | Nanoencapsulation | [145,146,155,156,157] |
| Micellar Casein | • A shift in pH and ultrasonication | Fish oil | Emulsions | [158] |
| Micellar casei Re-assembled casein micelle from micellar casein | • A shift in pH and ultrasonication | Vegetable oil (Lactobacillus and Bifidobacteria | Nanoencapsulation Microencapsulation | [158,159] |
| Casein micelles | • Mineral composition restoration • Homogenisation with high pressure | Omega-3 | Nanoencapsulation | [158] |
| β Casein micelles | • Lyophilization | Celecoxib | Nanoencapsulation | [160] |
| Casein micelles + konjac glucomannan (KGM) | • Enzyme-induced casein KGM hydrogels preparation • Ageing in refrigeration | Docetaxel | Gel | [161] |
| Casein micelles | • Skim milk natural conditions • Thermally treated commercial skim milk | Vitamin A | Nanoencapsulation | [156,162] |
| Casein micelles | • Mineral composition restoration and homogenisation at high pressure • Re-assembly of casein micelles | Vitamin D2 | Nanoencapsulation | [155] |
| Casein micelles | Rosemary Extract | Nanoencapsulation | [163] | |
| Casein micelle | Lactoferrin | Nanoencapsulation | [164] | |
| Casein micelle | • Spray-drying crosslinked with genipin | Alfuzosin | suspension | [165] |
| Casein micelle | • Spray-drying crosslinked with genipin | Flutamide | Microencapsulation | [165] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadiq, U.; Gill, H.; Chandrapala, J. Casein Micelles as an Emerging Delivery System for Bioactive Food Components. Foods 2021, 10, 1965. https://doi.org/10.3390/foods10081965
Sadiq U, Gill H, Chandrapala J. Casein Micelles as an Emerging Delivery System for Bioactive Food Components. Foods. 2021; 10(8):1965. https://doi.org/10.3390/foods10081965
Chicago/Turabian StyleSadiq, Uzma, Harsharn Gill, and Jayani Chandrapala. 2021. "Casein Micelles as an Emerging Delivery System for Bioactive Food Components" Foods 10, no. 8: 1965. https://doi.org/10.3390/foods10081965
APA StyleSadiq, U., Gill, H., & Chandrapala, J. (2021). Casein Micelles as an Emerging Delivery System for Bioactive Food Components. Foods, 10(8), 1965. https://doi.org/10.3390/foods10081965