
Impact of Lacticaseibacillus rhamnosus GG on the Emulsion Stability of Raw Milk

 , , , and
, , , and
Abstract

1. Introduction
2. Materials and Methods
2.1. Material
2.2. Bacterial Strains and Culture Conditions
2.3. Preparation of Milk-Containing LGG Samples
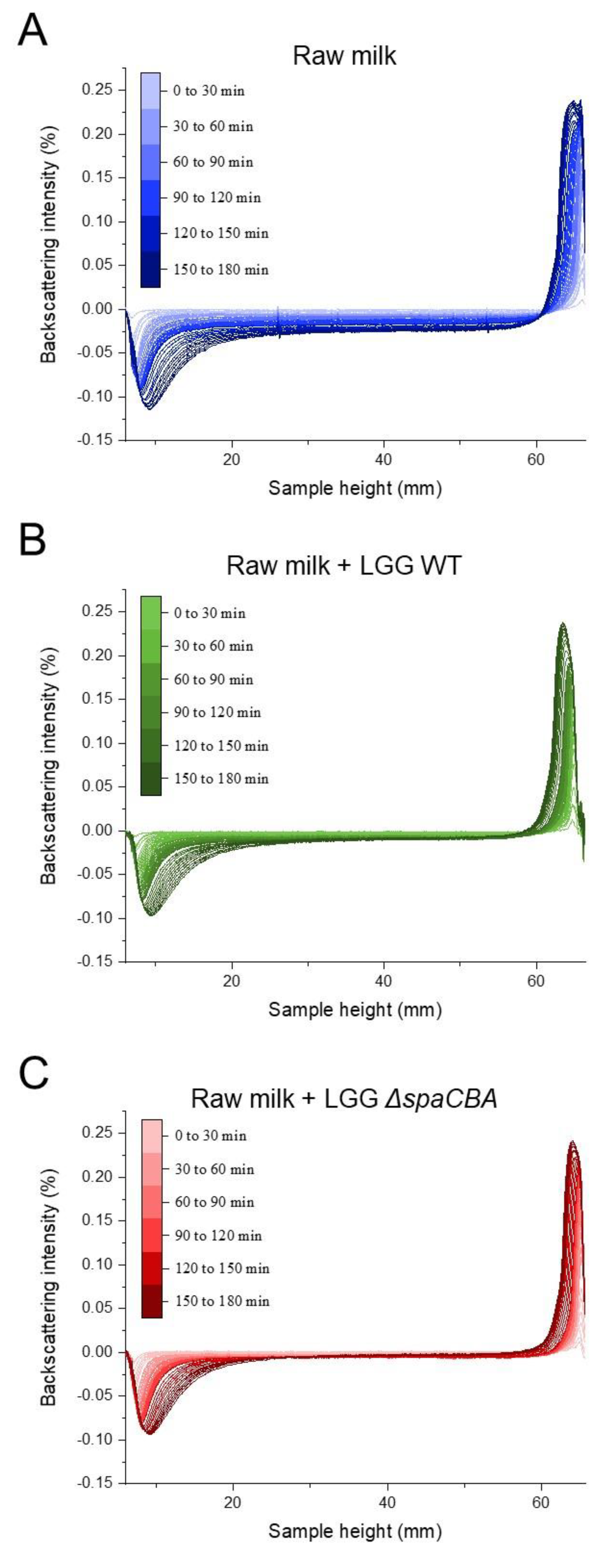
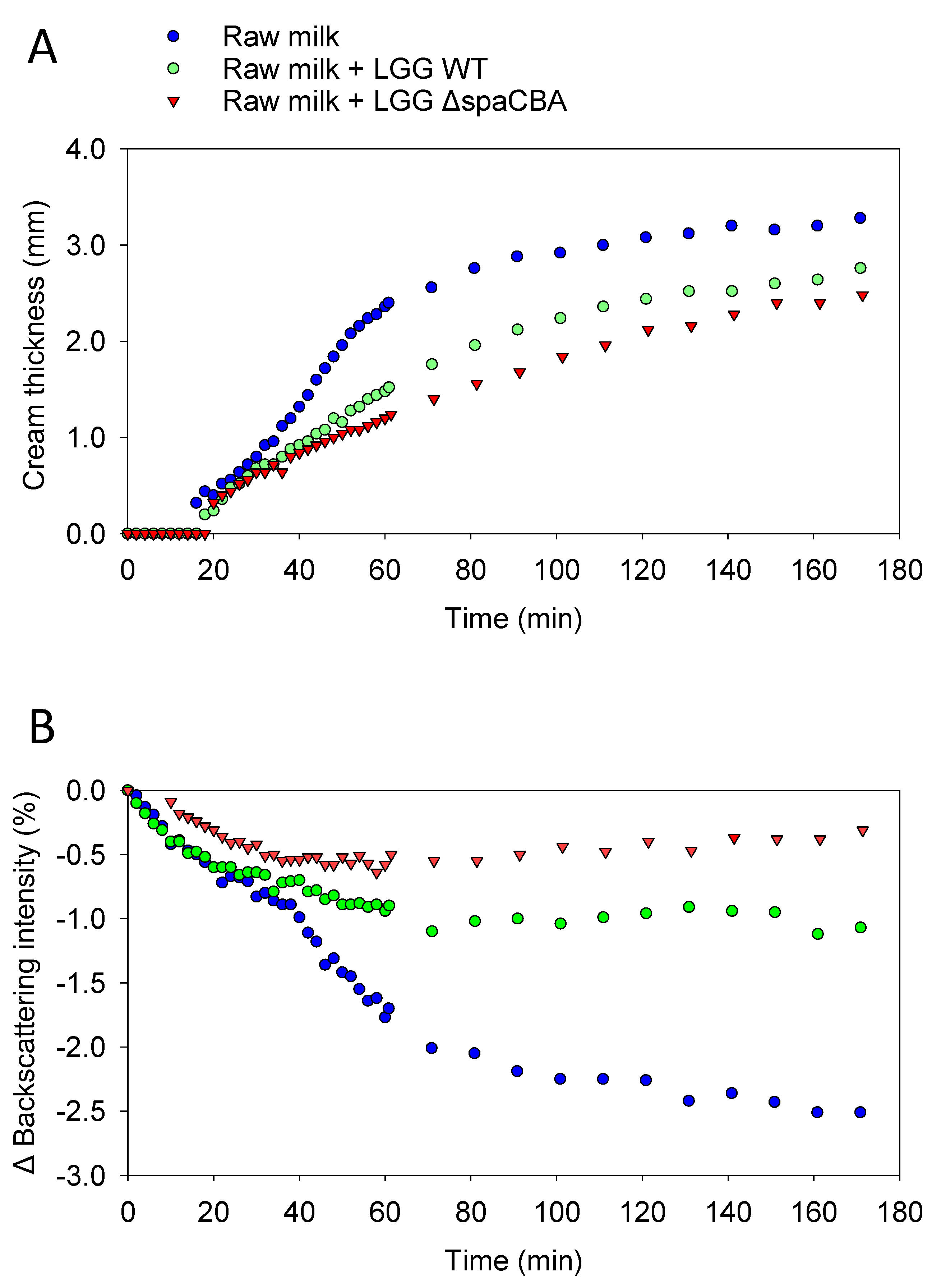
2.4. Emulsion Stability
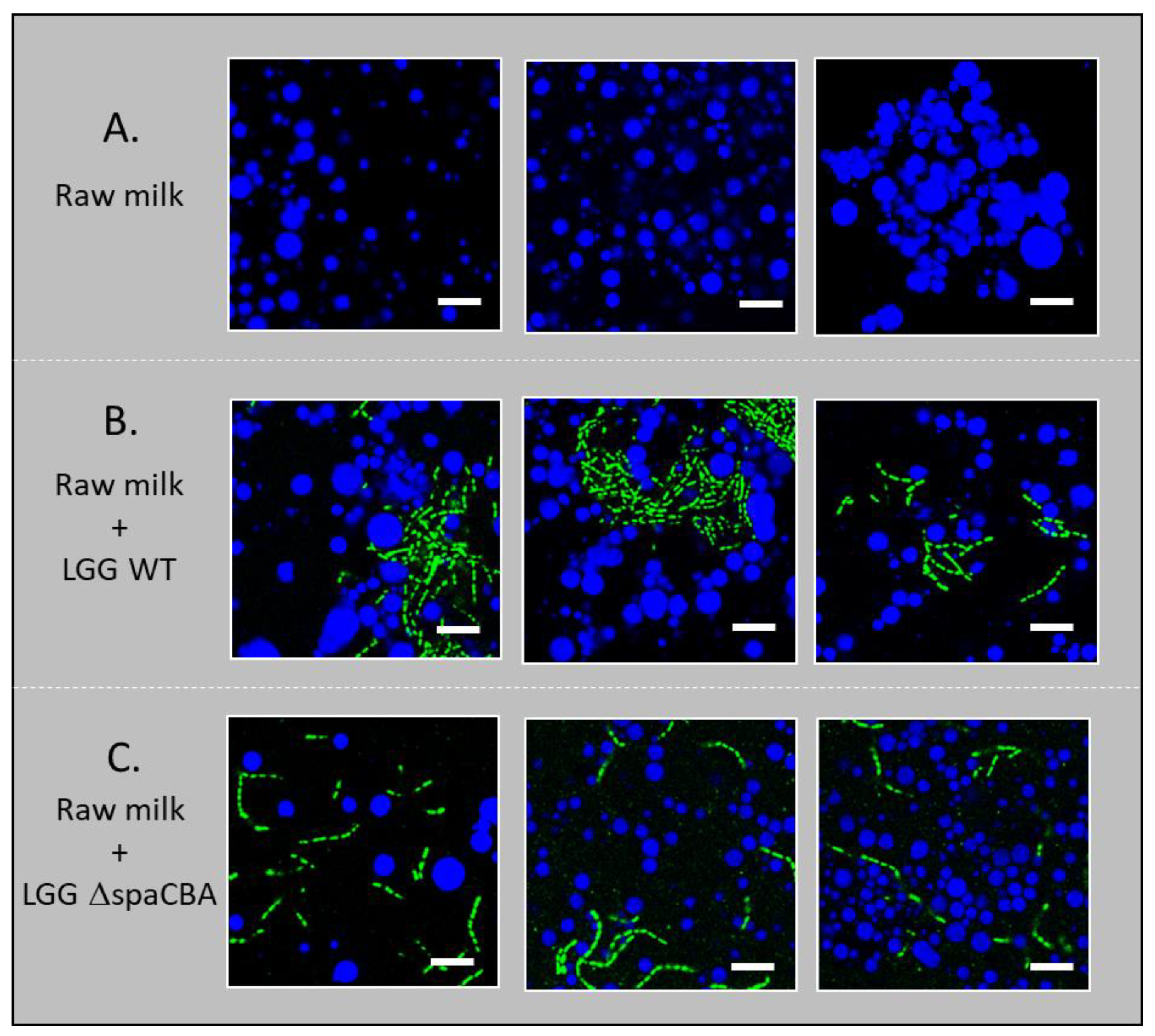
2.5. Microbial Distribution in the Milk Matrix
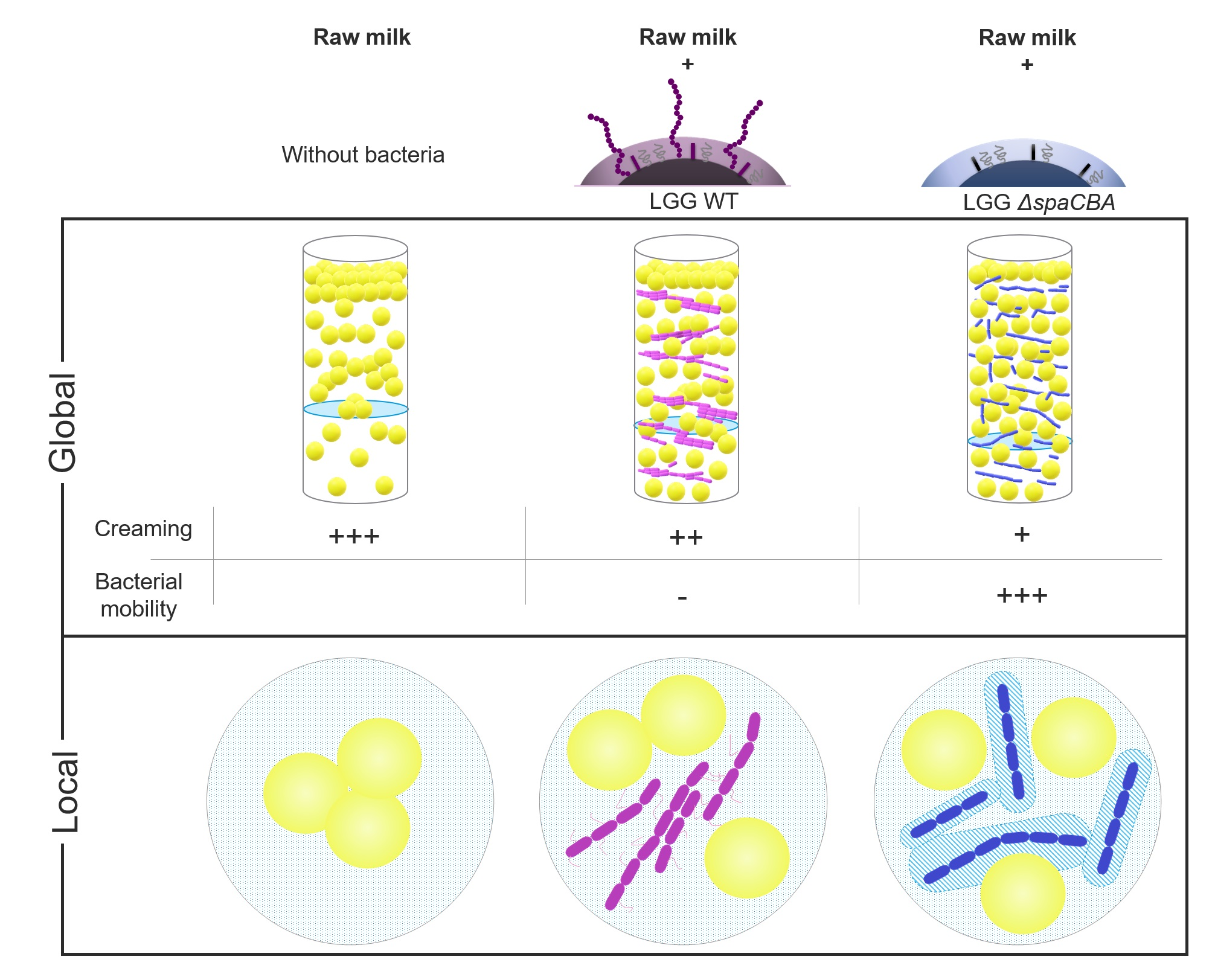
3. Results
3.1. Raw Milk Stability
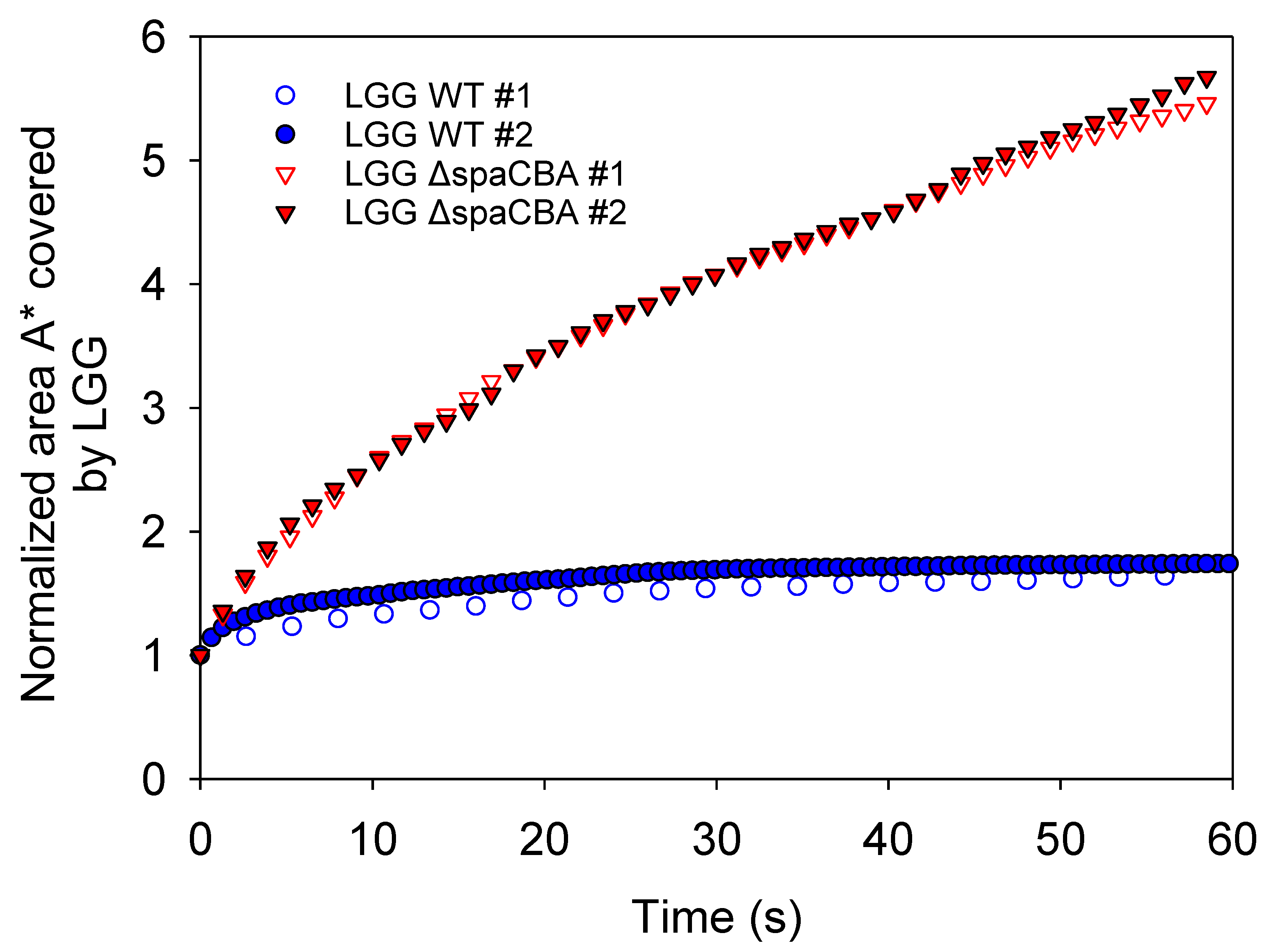
3.2. Microbial Distribution in Raw Milk
3.3. Matrix Structuration Dynamics
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lucey, J.A.; Otter, D.; Horne, D.S. A 100-Year Review: Progress on the Chemistry of Milk and Its Components. J. Dairy Sci. 2017, 100, 9916–9932. [Google Scholar] [CrossRef]
- Guerin, J.; Burgain, J.; Gomand, F.; Scher, J.; Gaiani, C. Milk Fat Globule Membrane Glycoproteins: Valuable Ingredients for Lactic Acid Bacteria Encapsulation? Crit. Rev. Food Sci. Nutr. 2019, 59, 639–651. [Google Scholar] [CrossRef]
- McClements, D.J. Theoretical Analysis of Factors Affecting the Formation and Stability of Multilayered Colloidal Dispersions. Langmuir 2005, 21, 9777–9785. [Google Scholar] [CrossRef] [PubMed]
- Mcclements, D.J.; Decker, E.A. Lipid Oxidation in Oil-in-Water Emulsions: Impact of Molecular Environment on Chemical Reactions in Heterogeneous Food Systems. J. Food Sci. 2000, 65, 1270–1282. [Google Scholar] [CrossRef]
- Dalgleish, D.G. Sedimentation of Casein Micelles During the Storage of Ultra-High Temperature Milk Products—A Calculation. J. Dairy Sci. 1992, 75, 371–379. [Google Scholar] [CrossRef]
- Ma, Y.; Barbano, D.M. Gravity Separation of Raw Bovine Milk: Fat Globule Size Distribution and Fat Content of Milk Fractions. J. Dairy Sci. 2000, 83, 1719–1727. [Google Scholar] [CrossRef]
- Walstra, P.; Oortwijn, H. Effect of Globule Size and Concentration on Creaming in Pasteurized Milk. Neth. Milk Dairy J. 1975, 29, 263–278. [Google Scholar]
- Mills, P.; Snabre, P. Apparent Viscosity and Particle Pressure of a Concentrated Suspension of Non-Brownian Hard Spheres near the Jamming Transition. Eur. Phys. J. E Soft Matter 2009, 30, 309–316. [Google Scholar] [CrossRef]
- Tcholakova, S.; Denkov, N.D.; Ivanov, I.B.; Campbell, B. Coalescence Stability of Emulsions Containing Globular Milk Proteins. Adv. Colloid Interface Sci. 2006, 123–126, 259–293. [Google Scholar] [CrossRef]
- Hardham, J.F.; Imison, B.W.; French, H.M. Effect of Homogenisation and Microfluidisation on the Extent of Fat Separation during Storage of UHT Milk. Aust. J. Dairy Technol. 2000, 55, 16–22. [Google Scholar]
- Boode, K.; Walstra, P. Partial Coalescence in Oil-in-Water Emulsions 1. Nature of the Aggregation. Colloids Surf. A Physicochem. Eng. Asp. 1993, 81, 121–137. [Google Scholar] [CrossRef]
- Britten, M.; Giroux, H.J. Coalescence Index of Protein-Stabilized Emulsions. J. Food Sci. 1991, 56, 792–795. [Google Scholar] [CrossRef]
- Das, K.P.; Kinsella, J.E. Droplet Size and Coalescence Stability of Whey Protein Stabilized Milkfat Peanut Oil Emulsions. J. Food Sci. 1993, 58, 439–444. [Google Scholar] [CrossRef]
- Dickinson, E.; Stainsby, G. Advances in Food Emulsions and Foams; Sole Distributor in the USA and Canada, Elsevier Science Publishers: Amsterdam, The Netherlands, 1988; ISBN 978-1-85166-200-5. [Google Scholar]
- Jeanson, S.; Chadœuf, J.; Madec, M.N.; Aly, S.; Floury, J.; Brocklehurst, T.F.; Lortal, S. Spatial Distribution of Bacterial Colonies in a Model Cheese. Appl. Environ. Microbiol. 2011, 77, 1493–1500. [Google Scholar] [CrossRef]
- Laloy, E.; Vuillemard, J.-C.; El Soda, M.; Simard, R.E. Influence of the Fat Content of Cheddar Cheese on Retention and Localization of Starters. Int. Dairy J. 1996, 6, 729–740. [Google Scholar] [CrossRef]
- Burgain, J.; Scher, J.; Francius, G.; Borges, F.; Corgneau, M.; Revol-Junelles, A.M.; Cailliez-Grimal, C.; Gaiani, C. Lactic Acid Bacteria in Dairy Food: Surface Characterization and Interactions with Food Matrix Components. Adv. Colloid Interface Sci. 2014, 213, 21–35. [Google Scholar] [CrossRef]
- Burgain, J.; Scher, J.; Lebeer, S.; Vanderleyden, J.; Cailliez-Grimal, C.; Corgneau, M.; Francius, G.; Gaiani, C. Significance of Bacterial Surface Molecules Interactions with Milk Proteins to Enhance Microencapsulation of Lactobacillus rhamnosus GG. Food Hydrocoll. 2014, 41, 60–70. [Google Scholar] [CrossRef]
- Dos Santos Morais, R.; Gaiani, C.; Borges, F.; Burgain, J. Interactions Microbe-Matrix in Dairy Products. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2020; ISBN 978-0-08-100596-5. [Google Scholar]
- Tarazanova, M.; Huppertz, T.; Kok, J.; Bachmann, H. Altering Textural Properties of Fermented Milk by Using Surface-Engineered Lactococcus lactis. Microb. Biotechnol. 2018, 11, 770–780. [Google Scholar] [CrossRef]
- Tarazanova, M.; Huppertz, T.; Kok, J.; Bachmann, H. Influence of Lactococcal Surface Properties on Cell Retention and Distribution in Cheese Curd. Int. Dairy J. 2018, 85, 73–78. [Google Scholar] [CrossRef]
- Dos Santos Morais, R.; El-Kirat-Chatel, S.; Burgain, J.; Simard, B.; Barrau, S.; Paris, C.; Borges, F.; Gaiani, C. A Fast, Efficient and Easy to Implement Method to Purify Bacterial Pili From Lacticaseibacillus rhamnosus GG Based on Multimodal Chromatography. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Gomand, F.; Borges, F.; Salim, D.; Burgain, J.; Guerin, J.; Gaiani, C. High-Throughput Screening Approach to Evaluate the Adhesive Properties of Bacteria to Milk Biomolecules. Food Hydrocoll. 2018, 84, 537–544. [Google Scholar] [CrossRef]
- Guerin, J.; Burgain, J.; Francius, G.; El-Kirat-Chatel, S.; Beaussart, A.; Scher, J.; Gaiani, C. Adhesion of Lactobacillus rhamnosus GG Surface Biomolecules to Milk Proteins. Food Hydrocoll. 2018, 82, 296–303. [Google Scholar] [CrossRef]
- Guerin, J.; Soligot, C.; Burgain, J.; Huguet, M.; Francius, G.; El-Kirat-Chatel, S.; Gomand, F.; Lebeer, S.; Le Roux, Y.; Borges, F.; et al. Adhesive Interactions between Milk Fat Globule Membrane and Lactobacillus rhamnosus GG Inhibit Bacterial Attachment to Caco-2 TC7 Intestinal Cell. Colloids Surf. B Biointerfaces 2018, 167, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Claes, I.; Tytgat, H.L.P.; Verhoeven, T.L.A.; Marien, E.; von Ossowski, I.; Reunanen, J.; Palva, A.; de Vos, W.M.; Keersmaecker, S.C.J.D.; et al. Functional Analysis of Lactobacillus rhamnosus GG Pili in Relation to Adhesion and Immunomodulatory Interactions with Intestinal Epithelial Cells. Appl. Environ. Microbiol. 2012, 78, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Mengual, O.; Meunier, G.; Cayre, I.; Puech, K.; Snabre, P. Characterisation of Instability of Concentrated Dispersions by a New Optical Analyser: The TURBISCAN MA 1000. Colloids Surf. A Physicochem. Eng. Asp. 1999, 152, 111–123. [Google Scholar] [CrossRef]
- Mengual, O.; Meunier, G.; Cayré, I.; Puech, K.; Snabre, P. TURBISCAN MA 2000: Multiple Light Scattering Measurement for Concentrated Emulsion and Suspension Instability Analysis. Talanta 1999, 50, 445–456. [Google Scholar] [CrossRef]
- Auty, M.A.E.; Twomey, M.; Guinee, T.P.; Mulvihill, D.M. Development and Application of Confocal Scanning Laser Microscopy Methods for Studying the Distribution of Fat and Protein in Selected Dairy Products. J. Dairy Res. 2001, 68, 417–427. [Google Scholar] [CrossRef]
- Gomand, F.; Borges, F.; Guerin, J.; El-Kirat-Chatel, S.; Francius, G.; Dumas, D.; Burgain, J.; Gaiani, C. Adhesive Interactions Between Lactic Acid Bacteria and β-Lactoglobulin: Specificity and Impact on Bacterial Location in Whey Protein Isolate. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Guerin, J.; Bacharouche, J.; Burgain, J.; Lebeer, S.; Francius, G.; Borges, F.; Scher, J.; Gaiani, C. Pili of Lactobacillus rhamnosus GG Mediate Interaction with β-Lactoglobulin. Food Hydrocoll. 2016, 58, 35–41. [Google Scholar] [CrossRef]
- Hickey, C.D.; Sheehan, J.J.; Wilkinson, M.G.; Auty, M.A.E. Growth and Location of Bacterial Colonies within Dairy Foods Using Microscopy Techniques: A Review. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ΔBackscattering Intensity (%) | Creaming Thickness (mm) | Creaming Rate (µm/min) | |
|---|---|---|---|
| Raw Milk | 2.6 | 3.3 | 62.1 |
| +LGG WT | 1.1 | 2.8 | 29.6 |
| +LGG ΔspaCBA | 0.7 | 2.5 | 21.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dos Santos Morais, R.; Louvet, N.; Borges, F.; Dumas, D.; Cvetkovska-Ben Mohamed, L.; Barrau, S.; Scher, J.; Gaiani, C.; Burgain, J. Impact of Lacticaseibacillus rhamnosus GG on the Emulsion Stability of Raw Milk. Foods 2021, 10, 991. https://doi.org/10.3390/foods10050991
Dos Santos Morais R, Louvet N, Borges F, Dumas D, Cvetkovska-Ben Mohamed L, Barrau S, Scher J, Gaiani C, Burgain J. Impact of Lacticaseibacillus rhamnosus GG on the Emulsion Stability of Raw Milk. Foods. 2021; 10(5):991. https://doi.org/10.3390/foods10050991
Chicago/Turabian StyleDos Santos Morais, Raphael, Nicolas Louvet, Frederic Borges, Dominique Dumas, Loubiana Cvetkovska-Ben Mohamed, Sarah Barrau, Joël Scher, Claire Gaiani, and Jennifer Burgain. 2021. "Impact of Lacticaseibacillus rhamnosus GG on the Emulsion Stability of Raw Milk" Foods 10, no. 5: 991. https://doi.org/10.3390/foods10050991
APA StyleDos Santos Morais, R., Louvet, N., Borges, F., Dumas, D., Cvetkovska-Ben Mohamed, L., Barrau, S., Scher, J., Gaiani, C., & Burgain, J. (2021). Impact of Lacticaseibacillus rhamnosus GG on the Emulsion Stability of Raw Milk. Foods, 10(5), 991. https://doi.org/10.3390/foods10050991