
Pectin Influences the Absorption and Metabolism of Polyphenols from Blackcurrant and Green Tea in Rats

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Experiment
2.2. Nitrogen and Corticosterone
2.3. Preparation of Samples for Polyphenol Analysis
2.4. Liquid Chromatography–Mass Spectrometry
2.5. Total Peroxyl Radical-Trapping Antioxidant Parameter (TRAP) and Uric Acid
2.6. Statistical Analysis
3. Results and Discussion
3.1. Rat Food Intake, Body Weight and Fecal Nitrogen
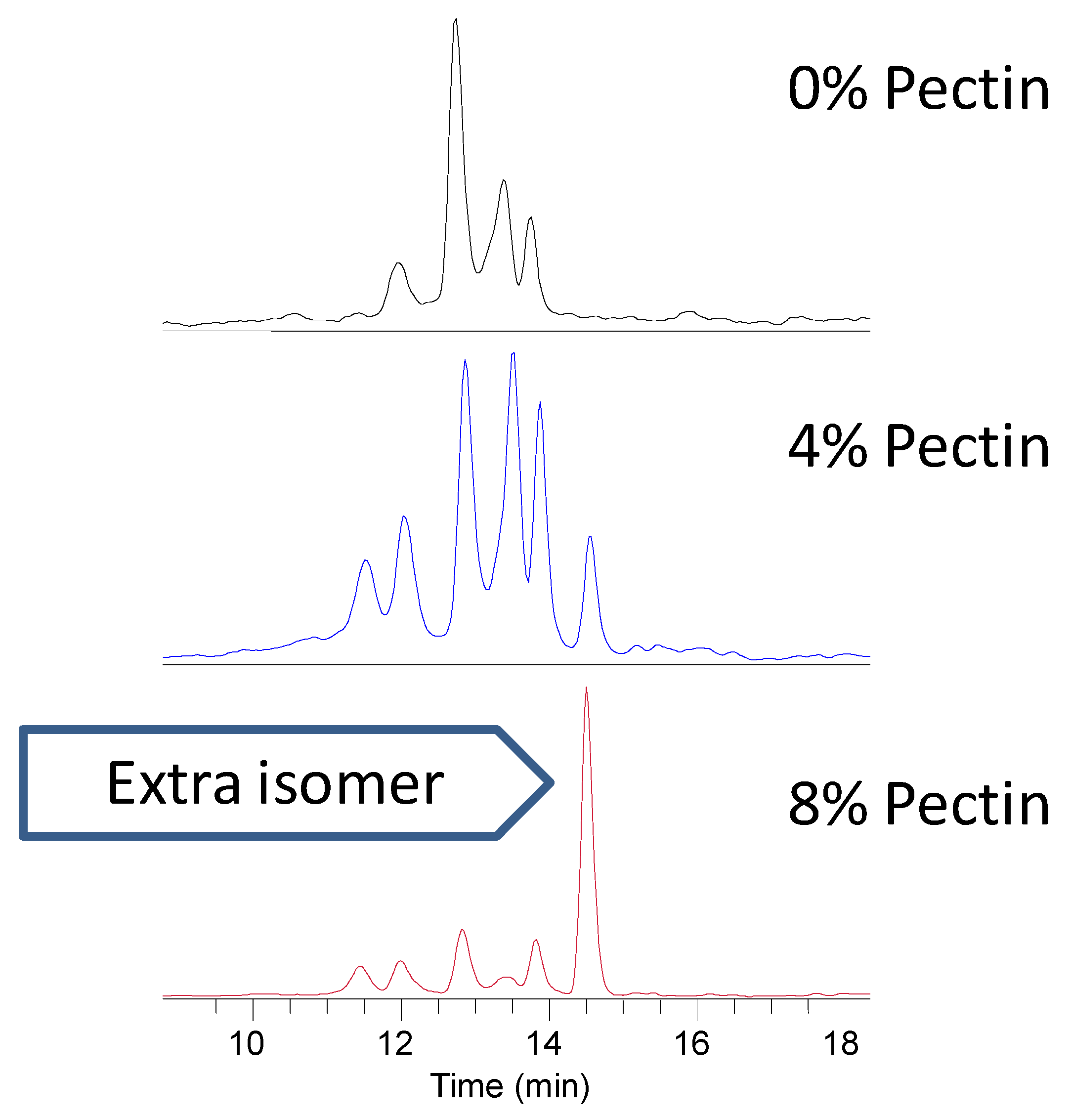
3.2. Polyphenols in Urine, Feces and Plasma of Rats
3.3. Plasma Corticosterone, TRAP and Uric Acid
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Serino, A.; Salazar, G. Protective role of polyphenols against vascular inflammation, aging and cardiovascular disease. Nutrients 2019, 11, 53. [Google Scholar] [CrossRef]
- Karaś, M.; Jakubczyk, A.; Szymanowska, U.; Złotek, U.; Zielińska, E. Digestion and bioavailability of bioactive phytochemicals. Int. J. Food Sci. Technol. 2017, 52, 291–305. [Google Scholar] [CrossRef]
- Teng, H.; Chen, L. Polyphenols and bioavailability: An update. Crit. Rev. Food Sci. Nutr. 2019, 59, 2040–2051. [Google Scholar] [CrossRef] [PubMed]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of dietary polyphenols: The role of metabolites. Crit. Rev. Food Sci. Nutr. 2020, 60, 626–659. [Google Scholar] [CrossRef] [PubMed]
- Eker, M.E.; Aaby, K.; Budic-Leto, I.; Rimac Brnčić, S.; El, S.N.; Karakaya, S.; Simsek, S.; Manach, C.; Wiczkowski, W.; de Pascual-Teresa, S. A review of factors affecting anthocyanin bioavailability: Possible implications for the inter-individual variability. Foods 2020, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Lavefve, L.; Howard, L.R.; Carbonero, F. Berry polyphenols metabolism and impact on human gut microbiota and health. Food Funct. 2020, 11, 45–65. [Google Scholar] [CrossRef]
- Li, Y.O.; Komarek, A.R. Dietary fibre basics: Health, nutrition, analysis, and applications. Food Qual. Saf. 2017, 1, 47–59. [Google Scholar] [CrossRef]
- Paturi, G.; Butts, C.A.; Monro, J.A.; Hedderley, D. Effects of blackcurrant and dietary fibers on large intestinal health biomarkers in rats. Plant Foods Hum. Nutr. 2018, 73, 54–60. [Google Scholar] [CrossRef]
- Bohn, T. Dietary factors affecting polyphenol bioavailability. Nutr. Rev. 2014, 72, 429–452. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, J.M. The food matrix: Implications in processing, nutrition and health. Crit. Rev. Food Sci. Nutr. 2019, 59, 3612–3629. [Google Scholar] [CrossRef] [PubMed]
- Jakobek, L.; Matić, P. Non-covalent dietary fiber—Polyphenol interactions and their influence on polyphenol bioaccessibility. Trends Food Sci. Technol. 2019, 83, 235–247. [Google Scholar] [CrossRef]
- Mullen, W.; Edwards, C.A.; Crozier, A. Absorption, excretion and metabolite profiling of methyl-, glucuronyl-, glucosyl- and sulpho-conjugates of quercetin in human plasma and urine after ingestion of onions. Br. J. Nutr. 2006, 96, 107–116. [Google Scholar] [CrossRef]
- Gee, J.M.; Wroblewska, M.A.; Bennett, R.N.; Mellon, F.A.; Johnson, I.T. Absorption and twenty-four-hour metabolism time-course of quercetin-3-O-glucoside in rats, in vivo. J. Sci. Food Agric. 2004, 84, 1341–1348. [Google Scholar] [CrossRef]
- Valkonen, M.; Kuusi, T. Spectrophotometric assay for total peroxyl radical-trapping antioxidant potential in human serum. J. Lipid Res. 1997, 38, 823–833. [Google Scholar] [CrossRef]
- Pires, M.A.; Pastrana, L.M.; Fuciños, P.; Abreu, C.S.; Oliveira, S.M. Sensorial perception of astringency: Oral mechanisms and current analysis methods. Foods 2020, 9, 1124. [Google Scholar] [CrossRef] [PubMed]
- Van Hul, M.; Cani, P.D. Targeting carbohydrates and polyphenols for a healthy microbiome and healthy weight. Curr. Nutr. Rep. 2019, 8, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Cirkovic Velickovic, T.D.; Stanic-Vucinic, D.J. The role of dietary phenolic compounds in protein digestion and processing technologies to improve their antinutritive properties. Compr. Rev. Food Sci. Food Saf. 2018, 17, 82–103. [Google Scholar] [CrossRef]
- Cai, Z.-Y.; Li, X.-M.; Liang, J.-P.; Xiang, L.-P.; Wang, K.-R.; Shi, Y.-L.; Yang, R.; Shi, M.; Ye, J.-H.; Lu, J.-L.; et al. Bioavailability of tea catechins and its improvement. Molecules 2018, 23, 2346. [Google Scholar] [CrossRef]
- Nishijima, T.; Iwai, K.; Saito, Y.; Takida, Y.; Matsue, H. Chronic ingestion of apple pectin can enhance the absorption of quercetin. J. Agric. Food Chem. 2009, 57, 2583–2587. [Google Scholar] [CrossRef]
- Zhu, F. Interactions between cell wall polysaccharides and polyphenols. Crit. Rev. Food Sci. Nutr. 2018, 58, 1808–1831. [Google Scholar] [CrossRef]
- Haider, K.; Wilde, P. Digestion and metabolism of pectin. In Pectin: Technological and Physiological Properties; Kontogiorgos, V., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 149–164. [Google Scholar]
- Williamson, G.; Clifford, M.N. Role of the small intestine, colon and microbiota in determining the metabolic fate of polyphenols. Biochem. Pharmacol. 2017, 139, 24–39. [Google Scholar] [CrossRef] [PubMed]
- Paturi, G.; Butts, C.A.; Stoklosinski, H.; Herath, T.D.; Monro, J.A. Short-Term feeding of fermentable dietary fibres influences the gut microbiota composition and metabolic activity in rats. Int. J. Food Sci. Technol. 2017, 52, 2572–2581. [Google Scholar] [CrossRef]
- Donoso, F.; Egerton, S.; Bastiaanssen, T.F.S.; Fitzgerald, P.; Gite, S.; Fouhy, F.; Ross, R.P.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Polyphenols selectively reverse early-life stress-induced behavioural, neurochemical and microbiota changes in the rat. Psychoneuroendocrinology 2020, 116, 104673. [Google Scholar] [CrossRef]
- Van de Wouw, M.; Boehme, M.; Lyte, J.M.; Wiley, N.; Strain, C.; O’Sullivan, O.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Short-chain fatty acids: Microbial metabolites that alleviate stress-induced brain–gut axis alterations. J. Physiol. 2018, 596, 4923–4944. [Google Scholar] [CrossRef] [PubMed]
- Gulcin, İ. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef]
- Pellegrini, N.; Serafini, M.; Colombi, B.; Del Rio, D.; Salvatore, S.; Bianchi, M.; Brighenti, F. Total antioxidant capacity of plant foods, beverages and oils consumed in italy assessed by three different in vitro assays. J. Nutr. 2003, 133, 2812–2819. [Google Scholar] [CrossRef]
- Denev, P.; Ciz, M.; Ambrozova, G.; Lojek, A.; Yanakieva, I.; Kratchanova, M. Solid-phase extraction of berries’ anthocyanins and evaluation of their antioxidative properties. Food Chem. 2010, 123, 1055–1061. [Google Scholar] [CrossRef]
- Mikami, T.; Sorimachi, M. Uric acid contributes greatly to hepatic antioxidant capacity besides protein. Physiol. Res. 2017, 66, 1001–1007. [Google Scholar] [CrossRef]
- Koguchi, T.; Tadokoro, T. Beneficial effect of dietary fiber on hyperuricemia in rats and humans: A review. Int. J. Vitam. Nutr. Res. 2019, 89, 89–108. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Control | Blackcurrant and Green Tea | Blackcurrant, Green Tea and Pectin (4%) | Blackcurrant, Green Tea and Pectin (8%) | |
|---|---|---|---|---|
| Lactic casein 1 | 299.4 | 299.4 | 299.4 | 299.4 |
| Vitamin mix 2 | 50 | 50 | 50 | 50 |
| Mineral mix 3 | 50 | 50 | 50 | 50 |
| Corn oil 4 | 97.6 | 97.6 | 97.6 | 97.6 |
| Starch 5 | 413 | 372.8 | 332.8 | 292.8 |
| Sucrose 6 | 40 | 40 | 40 | 40 |
| Cellulose 7 | 50 | 50 | 50 | 50 |
| Blackcurrant extract 8 | 40 | 40 | 40 | |
| Green tea extract 9 | 0.2 | 0.2 | 0.2 | |
| Pectin 10 | 40 | 80 |
| Final Body Weight (g) | Body Weight Gain (g) | Food Intake (g/day) | |
|---|---|---|---|
| Control | 309 a | 59 a | 16 b |
| Blackcurrant and green tea | 303 a | 41 b | 12 a,b |
| Blackcurrant, green tea and pectin (4%) | 316 a | 44 a,b | 17 b |
| Blackcurrant, green tea and pectin (8%) | 317 a | 50 a,b | 11 a |
| Tukey HSD | 30 | 16 | 4 |
| p value | 0.552 | 0.025 | < 0.001 |
| Protein Intake (g) | Protein Efficiency Ratio 1 | Fecal Nitrogen Output (mg/5 Days) | Apparent Nitrogen Balance (mg/5 Days) 2 | Protein Digestibility (%) 3 | |
|---|---|---|---|---|---|
| Control | 42.8 a | 1.4 a | 202 c | 152 c | 78 a |
| Blackcurrant and green tea | 38.3 b | 1.0 a | 244 c | 220 b,c | 76 a |
| Blackcurrant, green tea and pectin (4%) | 40.6 a,b | 1.1 a | 307 b | 554 a | 78 a |
| Blackcurrant, green tea and pectin (8%) | 41.0 a,b | 1.2 a | 368 a | 356 b | 70 b |
| Tukey HSD | 3.7 | 0.4 | 56 | 170 | 4 |
| p value | 0.021 | 0.089 | <0.001 | <0.001 | <0.001 |
| Diets 1 | Urine | Feces | Plasma | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Pectin (%) | Pectin (%) | Pectin (%) | ||||||||
| 0 | 4 | 8 | 0 | 4 | 8 | 0 | 4 | 8 | ||
| Catechins | ||||||||||
| Catechin | ● | ● | ● | ● | ||||||
| Epicatechin | ● | ● | ● | |||||||
| Gallocatechin | ● | ● | ● | ● | ||||||
| Epigallocatechin | ● | ● | ● | ● | ||||||
| Catechin gallate | ● | ● | ● | |||||||
| Epicatechin gallate | ● | ● | ● | ● | ||||||
| Gallocatechin gallate | ● | ● | ● | ● | ||||||
| Epigallocatechin gallate | ● | ● | ● | ● | ||||||
| Catechin diglucuronide sulfate | ● | ● | ||||||||
| Epicatechin glucuronide | ● | ● | ● | ● | ● | ● | ||||
| Methyl epicatechin glucuronide | ● | ● | ● | ● | ● | ● | ||||
| Gallocatechin glucuronide | ● | ● | ● | ● | ||||||
| Methyl gallocatechin glucuronide | ● | ● | ● | ● | ● | ● | ||||
| Trimethyl catechin diglucuronide | ● | ● | ● | ● | ● | ● | ||||
| Anthocyanins | ||||||||||
| Cyanidin glucoside | ● | ● | ● | ● | ● | ● | ● | |||
| Cyanidin rutinoside | ● | ● | ● | ● | ● | ● | ● | |||
| Delphinidin glucoside | ● | ● | ● | |||||||
| Delphinidin rutinoside | ● | ● | ● | ● | ● | ● | ● | |||
| Petunidin glucoside | ● | ● | ● | ● | ● | |||||
| Petunidin rutinoside | ● | ● | ● | ● | ||||||
| Peonidin rutinoside | ● | ● | ● | ● | ● | |||||
| Methyl cyanidin glucuronide | ● | ● | ||||||||
| Methyl delphinidin diglucuronide | ● | ● | ● | |||||||
| Methyl delphinidin glucuronide | ● | ● | ||||||||
| Flavonols | ||||||||||
| Quercetin glucoside | ● | ● | ● | ● | ||||||
| Quercetin rutinoside | ● | ● | ● | ● | ||||||
| Myricetin glucoside | ● | ● | ● | ● | ||||||
| Myricetin rutinoside | ● | |||||||||
| Quercetin glucuronide | ● | ● | ● | |||||||
| Methyl quercetin glucuronide | ● | ● | ● | |||||||
| Quercetin diglucuronide | ● | ● | ● | ● | ● | ● | ||||
| Methyl quercetin diglucuronide | ● | ● | ● | ● | ● | ● | ||||
| Myricetin glucuronide | ● | ● | ● | ● | ● | |||||
| Myricetin diglucuronide | ● | ● | ● | ● | ● | ● | ||||
| Quercetin triglucuronide | ● | ● | ||||||||
| Kaempferol diglucuronide | ● | ● | ● | |||||||
| Myricetin triglucuronide | ● | ● | ● | |||||||
| Phenolic acids | ||||||||||
| 3,4 dihydroxybenzoic acid | ● | ● | ● | ● | ● | ● | ||||
| 2,4 dihydroxybenzoic acid | ● | ● | ● | ● | ● | ● | ||||
| Corticosterone (ng/mL) | |
|---|---|
| Control | 69 a |
| Blackcurrant and green tea | 58 a |
| Blackcurrant, green tea and pectin (4%) | 48 a |
| Blackcurrant, green tea and pectin (8%) | 40 a |
| Tukey HSD 1 | 713% |
| p value | 0.369 |
| TRAP (µmol/L) | Uric Acid (µmol/L) | |
|---|---|---|
| Control | 1085 a | 150 a |
| Blackcurrant and green tea | 1139 a | 148 a |
| Blackcurrant, green tea and pectin (4%) | 1189 a | 120 a |
| Blackcurrant, green tea and pectin (8%) | 1189 a | 124 a |
| Tukey HSD | 109 | 52 |
| p value | 0.046 | 0.267 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paturi, G.; Butts, C.A.; Joyce, N.I.; Rippon, P.E.; Morrison, S.C.; Hedderley, D.I.; Lister, C.E. Pectin Influences the Absorption and Metabolism of Polyphenols from Blackcurrant and Green Tea in Rats. Foods 2021, 10, 813. https://doi.org/10.3390/foods10040813
Paturi G, Butts CA, Joyce NI, Rippon PE, Morrison SC, Hedderley DI, Lister CE. Pectin Influences the Absorption and Metabolism of Polyphenols from Blackcurrant and Green Tea in Rats. Foods. 2021; 10(4):813. https://doi.org/10.3390/foods10040813
Chicago/Turabian StylePaturi, Gunaranjan, Christine A. Butts, Nigel I. Joyce, Paula E. Rippon, Sarah C. Morrison, Duncan I. Hedderley, and Carolyn E. Lister. 2021. "Pectin Influences the Absorption and Metabolism of Polyphenols from Blackcurrant and Green Tea in Rats" Foods 10, no. 4: 813. https://doi.org/10.3390/foods10040813
APA StylePaturi, G., Butts, C. A., Joyce, N. I., Rippon, P. E., Morrison, S. C., Hedderley, D. I., & Lister, C. E. (2021). Pectin Influences the Absorption and Metabolism of Polyphenols from Blackcurrant and Green Tea in Rats. Foods, 10(4), 813. https://doi.org/10.3390/foods10040813