
Evaluation of Lipid Oxidation, Volatile Compounds and Vibrational Spectroscopy of Silver Carp (Hypophthalmichthys molitrix) during Ice Storage as Related to the Quality of Its Washed Mince


Abstract

1. Introduction
2. Materials and Methods
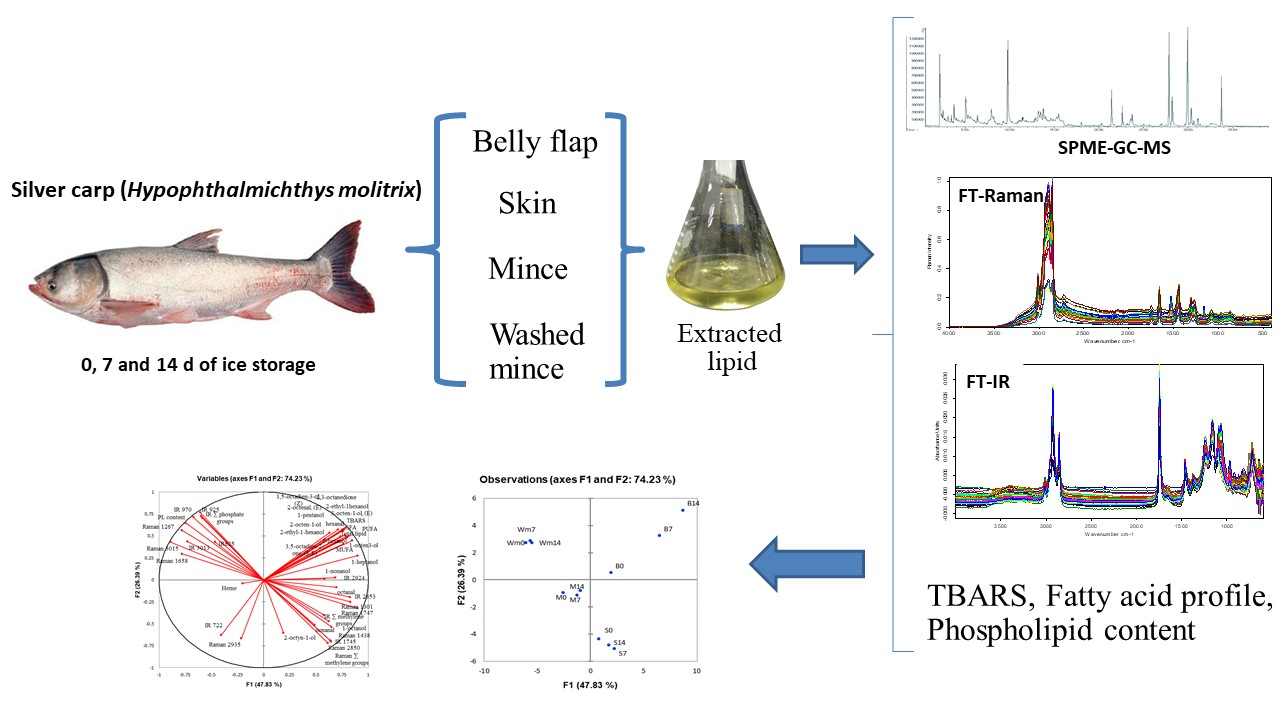
2.1. Sample Preparation
2.2. Analysis of Lipid and Fatty Acids
2.3. Heme Iron Content
2.4. Thiobarbituric Acid Reactive Substances (TBARS)
2.5. Determination of Volatile Compounds
2.6. Vibrational Spectroscopy
2.7. Statistical Analyses
3. Results and Discussion
3.1. Changes in Lipids
3.2. Heme Iron Content
3.3. TBARS Value
3.4. Volatile Compounds
3.5. FT—Raman Spectroscopy
3.6. FTIR
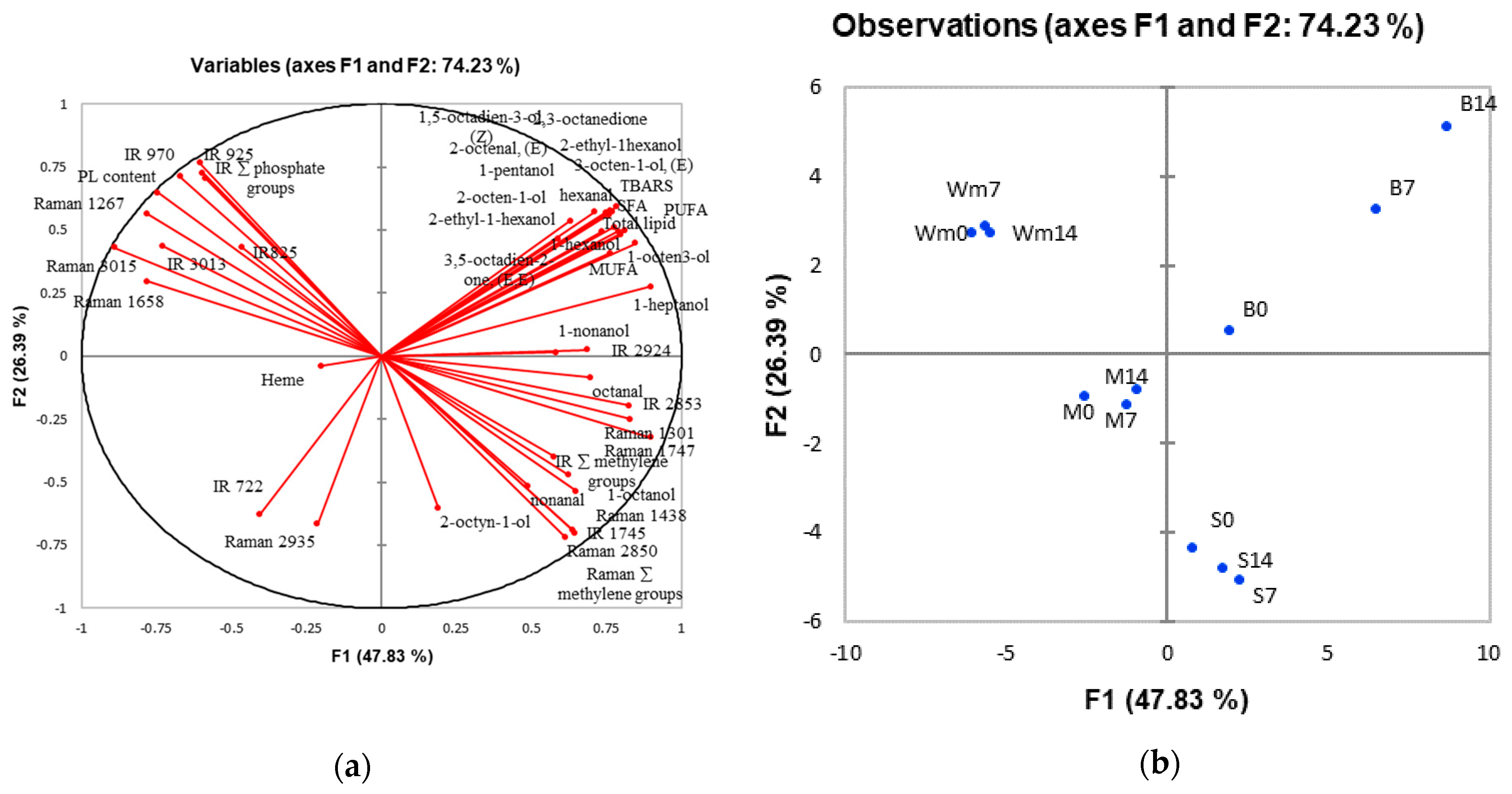
3.7. Principal Component Analysis (PCA)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Yearbook. In Fishery and Aquaculture Statistics 2017; Food and Agriculture Organization of the United Nations: Rome, Italy, 2019. [Google Scholar]
- Park, J.W.; Graves, D.; Draves, R.; Yongsawatdigul, J. Manufacture of Surimi. In Surimi and Surimi Seafood, 3rd ed.; Park, J.W., Ed.; CRC Press: Boca Raton, FL, USA, 2013; pp. 55–97. [Google Scholar]
- Weng, W.; Zheng, W. Silver carp (Hypophthalmichthys molitrix) surimi acid-induced gel extract characteristics: A Comparison with heat-induced gel. Int. J. Food Prop. 2015, 18, 821–832. [Google Scholar] [CrossRef]
- Reitznerová, A.; Šuleková, M.; Nagy, J.; Marcinčák, S.; Semjon, B.; Čertík, M.; Klempová, T. Lipid peroxidation process in meat and meat products: A comparison study of malondialdehyde determination between modified 2-thiobarbituric acid spectrophotometric method and reverse-phase high-performance liquid chromatography. Molecules 2017, 22, 1988. [Google Scholar] [CrossRef]
- Giménez, B.; Gómez-Guillén, M.C.; Pérez-Mateos, M.; Montero, P.; Márquez-Ruiz, G. Evaluation of lipid oxidation in horse mackerel patties covered with borage-containing film during frozen storage. Food Chem. 2011, 124, 1393–1403. [Google Scholar] [CrossRef]
- Iglesias, J.; Medina, I. Solid-phase microextraction method for the determination of volatile compounds associated to oxidation of fish muscle. J. Chromatogr. A 2008, 1192, 9–16. [Google Scholar] [CrossRef]
- Sánchez-Alonso, I.; Carmona, P.; Careche, M. Vibrational spectroscopic analysis of hake (Merluccius merluccius L.) lipids during frozen storage. Food Chem. 2012, 132, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Xie, Y.; Xi, J.; Guo, Y.; Qian, H.; Cheng, Y.; Chen, Y.; Yao, W. Characterization of lipid oxidation process of beef during repeated freeze-thaw by electron spin resonance technology and Raman spectroscopy. Food Chem. 2018, 243, 58–64. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Eymard, S.; Carcouët, E.; Rochet, M.J.; Dumay, J.; Chopin, C.; Genot, C. Development of lipid oxidation during manufacturing of horse mackerel surimi. J. Sci. Food Agric. 2005, 85, 1750–1756. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis of the Association of Official Analytical Chemists, 18th ed.; AOAC: Arlington, VA, USA, 2005. [Google Scholar]
- Clark, E.M.; Mahoney, A.W.; Carpenter, C.E. Heme and total iron in ready-to-eat chicken. J. Agric. Food Chem. 1997, 45, 124–126. [Google Scholar] [CrossRef]
- Lombardi-Boccia, G.; Martínez-Domínguez, B.; Aguzzia, A.; Rincón-León, F. Optimization of heme iron analysis in raw and cooked red meat. Food Chem. 2002, 78, 505–510. [Google Scholar] [CrossRef]
- Ackman, R. Seafood lipids. In Seafoods: Chemistry, Processing Technology and Quality; Shahidi, F., Botta, J.R., Eds.; Springer: Boston, MA, USA, 1994; pp. 34–48. [Google Scholar]
- Moradi, Y.; Bakar, J.; Motalebi, A.; Syed Muhamad, S.; Che Man, Y. A review on fish lipid: Composition and changes during cooking methods. J. Aquat. Food Prod. Technol. 2011, 20, 379–390. [Google Scholar] [CrossRef]
- Thilakarathne, L.; Attygalle, M. Lipid composition of skin and muscle of the Indo-Pacific sailfish, Istiophorus platypterus. Vidyodaya J. Sci. 2009, 14, 161–166. [Google Scholar]
- Aursand, M.; Bleivik, B.; Rainuzzo, J.R.; Leif, J.; Mohr, V. Lipid distribution and composition of commercially farmed Atlantic salmon (Salmosalar). J. Sci. Food Agric. 1994, 64, 239–248. [Google Scholar] [CrossRef]
- Tongnuanchan, P.; Benjakul, S.; Prodpran, T.; Songtipya, P. Characteristics of film based on protein isolate from red tilapia muscle with negligible yellow discoloration. Int. J. Biol. Macromol. 2011, 48, 758–767. [Google Scholar] [CrossRef]
- Domínguez, R.; Pateiro, M.; Gagaoua, M.; Barba, F.J.; Zhang, W.; Lorenzo, J.M. A comprehensive review on lipid oxidation in meat and meat products. Antioxidants 2020, 8, 429. [Google Scholar] [CrossRef]
- Chaijan, M.; Benjakul, S.; Visessanguan, W.; Faustman, C. Changes of lipids in sardine (Sardinella gibbosa) muscle during iced storage. Food Chem. 2006, 99, 83–91. [Google Scholar] [CrossRef]
- Chávez-Mendoza, C.; García-Macías, J.A.; Alarcón-Rojo, A.D.; Ortega-Gutiérrez, J.Á.; Holguín-Licón, C.; Corral-Flores, G. Comparison of fatty acid content of fresh and frozen fillets of rainbow trout (Oncorhynchus mykiss) Walbaum. Braz. Arch. Biol. Technol. 2014, 57, 103–109. [Google Scholar] [CrossRef]
- Šimat, V.; Bogdanović, T.; Poljak, V.; Petričević, S. Changes in fatty acid composition, atherogenic and thrombogenic health lipid indices and lipid stability of bogue (Boops boops Linnaeus, 1758) during storage on ice: Effect of fish farming activities. J. Food Compost. Anal. 2015, 40, 120–125. [Google Scholar] [CrossRef]
- Thiansilakul, Y.; Benjakul, S.; Richards, M.P. Changes in heme proteins and lipids associated with off-odour of seabass (Lates calcarifer) and red tilapia (Oreochromis mossambicus × O. niloticus) during iced storage. Food Chem. 2010, 121, 1109–1119. [Google Scholar]
- Rezaei, M.; Hosseini, S. Quality assessment of farmed rainbow trout (Oncorhynchus mykiss) during chilled storage. J. Food Sci. 2008, 73, H93–H96. [Google Scholar] [CrossRef] [PubMed]
- Mei, J.; Ma, X.; Xie, J. Review on natural preservatives for extending fish shelf life. Foods 2019, 8, 490. [Google Scholar] [CrossRef] [PubMed]
- Addeen, A.; Benjakul, S.; Maqsood, S. Haemoglobin-mediated lipid oxidation in washed chicken mince. Indian J. Sci. Technol. 2016, 9, 1–8. [Google Scholar] [CrossRef]
- Hoke, M.E.; Jahncke, M.L.; Silva, J.L.; Hearnsberger, J.O.; Suriyaphan, O. Stability of washed frozen mince from channel catfish frames. J. Food Sci. 2000, 65, 1083–1086. [Google Scholar] [CrossRef]
- Iglesias, J.; Medina, I.; Bianchi, F.; Careri, M.; Mangia, A.; Musci, M. Study of the volatile compounds useful for the characterisation of fresh and frozen-thawed cultured gilthead sea bream fish by solid-phase microextraction gas chromatography–mass spectrometry. Food Chem. 2009, 115, 1473–1478. [Google Scholar] [CrossRef]
- Choe, E.; Min, D.B. Mechanisms and factors for edible oil oxidation. Compr. Rev. Food Sci. Food Saf. 2006, 5, 169–186. [Google Scholar] [CrossRef]
- Buettner, A. Springer Handbook of Odor; Springer: Berlin/Heidelberg, Germany, 2017; p. VI. [Google Scholar] [CrossRef]
- Zhang, Q.; Ding, Y.; Gu, S.; Zhu, S.; Zhou, X.; Ding, Y. Identification of changes in volatile compounds in dry-cured fish during storage using HS-GC-IMS. Food Res. Int. 2020, 137, 109339. [Google Scholar] [CrossRef]
- Qian, M.C.; An, Y.; Qian, Y.L.; Alcazar Magana, A.; Xiong, S. Comparative characterization of aroma compounds in silver carp (Hypophthalmichthys molitrix), Pacific whiting (Merluccius productus) and Alaska pollock (Theragra chalcogramma) surimi by aroma etract dilution analysis, odor activity value, and aroma recombination studies. J. Agric. Food Chem. 2020, 68, 10403–10413. [Google Scholar]
- Cebi, N.; Yilmaz, M.T.; Sagdic, O.; Yuce, H.; Yelboga, E. Prediction of peroxide value in omega-3 rich microalgae oil by ATR-FTIR spectroscopy combined with chemometrics. Food Chem. 2017, 225, 188–196. [Google Scholar] [CrossRef]
- Rohman, A.; Che Man, Y.B. Quantification and classification of corn and sunflower oils as adulterants in olive oil using chemometrics and FTIR spectra. Sci. World J. 2012, 2012, 1–6. [Google Scholar] [CrossRef]
- Volpe, M.; Coccia, E.; Siano, F.; Di Stasio, M.; Paolucci, M. Rapid evaluation methods for quality of Trout (Oncorhynchus mykiss) fresh fillet preserved in an active edible coating. Foods 2019, 8, 113. [Google Scholar] [CrossRef] [PubMed]
- Maurer, N.E.; Hatta-Sakoda, B.; Pascual-Chagman, G.; Rodriguez-Saona, L.E. Characterization and authentication of a novel vegetable source of omega-3 fatty acids, sacha inchi (Plukenetia volubilis L.) oil. Food Chem. 2012, 134, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Henna Lu, F.S.; Tan, P.P. A comparative study of storage stability in virgin coconut oil and extra virgin olive oil upon thermal treatment. Int. Food Res. J. 2009, 16, 343–354. [Google Scholar]
- Setiowaty, G.; Che Man, Y.B.; Jinap, S.; Moh, M.H. Quantitative determination of peroxide value in thermally oxidized palm olein by Fourier transform infrared spectroscopy. Phytochem. Anal. 2000, 11, 74–78. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Storage Time (Days) | Skin | Belly | Mince | Washed Mince | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 7 | 14 | 0 | 7 | 14 | 0 | 7 | 14 | 0 | 7 | 14 | |
| Total lipid (g/100 g) | 8.78 ± 0.34 a | 8.12 ± 0.55 a | 7.07 ± 0.60 b | 40.52 ± 1.86 a | 38.60 ± 1.98 a | 33.86 ± 2.74 b | 11.45 ± 0.15 a | 10.74 ± 0.33 b | 9.00 ± 0.14 c | 6.40 ± 0.52 | 6.29 ± 0.85 | 5.83 ± 0.56 |
| Phospholipid (g PC/100 g) | 0.52 ± 0.08 | 0.51 ± 0.09 | 0.47 ± 0.12 | 1.15 ± 0.18 | 1.10 ± 0.21 | 0.99 ± 0.22 | 1.67 ± 0.27 a | 1.38 ± 0.17 b | 1.19 ± 0.20 b | 2.49 ± 0.23 | 2.35 ± 0.41 | 2.17 ± 0.36 |
| Thiobarbituric acid reactive substances (TBARS) (ng of malondialdehyde (MDA)/kg) | 0.69 ± 0.05 b | 0.83 ± 0.11 ab | 0.94 ± 0.06 a | 2.08 ± 0.10 c | 10.12 ± 0.90 b | 12.51 ± 0.21 a | 0.59 ± 0.03 c | 0.70 ± 0.02 b | 0.86 ± 0.06 a | 0.77 ± 0.08 b | 0.96 ± 0.05 a | 1.06 ± 0.05 a |
| Heme iron (mg/100 g) | 0.092 ± 0.01 | 0.090 ± 0.01 | 0.071 ± 0.01 | 0.176 ± 0.02 | 0.155 ± 0.04 | 0.156 ± 0.05 | 0.772 ± 0.08 a | 0.590 ± 0.03 b | 0.294 ± 0.05 c | 0.193 ± 0.04 a | 0.118 ± 0.03 b | 0.102 ± 0.02 b |
| Fatty Acids | Skin | Belly | Mince | Washed Mince | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D0 | D7 | D14 | D0 | D7 | D14 | D0 | D7 | D14 | D0 | D7 | D14 | |
| C10:0 | 0.20 | 0.25 | 0.27 | 1.15 | 1.21 | 1.17 | 0.15 | 0.14 | 0.34 | 0.10 | 0.09 | 0.09 |
| C12:0 | 0.08 | 0.10 | 0.08 | 0.50 | 0.52 | 0.60 | 0.05 | 0.03 | 0.08 | 0.03 | 0.03 | 0.04 |
| C13:0 | 0.04 | 0.07 | 0.05 | 0.32 | 0.39 | 0.43 | 0.03 | 0.04 | 0.06 | 0.00 | 0.02 | 0.03 |
| C14:0 | 1.30 | 2.06 | 1.59 | 9.55 | 11.28 | 12.08 | 0.92 | 1.17 | 2.04 | 0.56 | 0.65 | 1.20 |
| C15:0 | 0.29 | 0.39 | 0.29 | 0.88 | 0.87 | 0.93 | 0.37 | 0.32 | 0.74 | 0.23 | 0.28 | 0.44 |
| C16:0 | 8.16 | 8.53 | 10.90 | 58.59 | 63.75 | 65.59 | 7.87 | 8.37 | 10.63 | 6.00 | 6.33 | 10.51 |
| C18:0 | 1.49 | 2.20 | 1.90 | 10.74 | 11.71 | 11.94 | 1.86 | 1.61 | 3.36 | 1.18 | 1.19 | 1.31 |
| C20:0 | 0.14 | 0.28 | 0.15 | 0.89 | 0.90 | 1.09 | 0.11 | 0.11 | 0.23 | 0.08 | 0.08 | 0.16 |
| C21:0 | 0.02 | 0.02 | 0.04 | 0.18 | 0.25 | 0.31 | 0.00 | 0.02 | 0.10 | 0.00 | 0.02 | 0.03 |
| C22:0 | 0.00 | 0.04 | 0.00 | 0.08 | 0.05 | 0.05 | 0.00 | 0.03 | 0.12 | 0.00 | 0.02 | 0.03 |
| C23:0 | 0.00 | 0.00 | 0.03 | 0.07 | 0.20 | 0.22 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| C24:0 | 0.86 | 0.95 | 0.90 | 4.26 | 2.52 | 2.52 | 2.43 | 2.30 | 2.43 | 2.03 | 1.82 | 2.29 |
| C14:1 | 0.64 | 0.73 | 0.38 | 3.66 | 4.30 | 3.35 | 0.21 | 0.14 | 0.03 | 0.03 | 0.02 | 0.02 |
| C16:1 | 3.77 | 3.33 | 3.43 | 24.09 | 24.56 | 23.19 | 2.49 | 2.13 | 2.16 | 1.60 | 1.66 | 1.54 |
| C17:1 | 0.74 | 0.64 | 0.58 | 3.72 | 3.48 | 4.11 | 0.48 | 0.40 | 0.41 | 0.34 | 0.32 | 0.39 |
| C18:1n9t | 0.28 | 0.06 | 0.17 | 1.08 | 1.59 | 1.41 | 0.16 | 0.14 | 0.14 | 0.12 | 0.11 | 0.12 |
| C18:1n9c | 15.13 | 14.14 | 10.01 | 85.16 | 70.87 | 67.39 | 7.12 | 6.46 | 6.22 | 4.93 | 4.44 | 3.95 |
| C20:1 | 0.92 | 1.56 | 0.87 | 5.38 | 4.72 | 6.21 | 0.58 | 0.57 | 0.60 | 0.38 | 0.33 | 0.59 |
| C22:1n9 | 0.07 | 0.04 | 0.03 | 0.16 | 0.25 | 0.26 | 0.03 | 0.03 | 0.03 | 0.02 | 0.02 | 0.02 |
| C18:2n6t | 0.15 | 0.10 | 0.12 | 1.20 | 1.20 | 1.15 | 0.06 | 0.06 | 0.04 | 0.04 | 0.05 | 0.09 |
| C18:2n6c | 2.80 | 1.80 | 2.17 | 13.12 | 16.74 | 15.15 | 1.91 | 1.68 | 1.74 | 1.61 | 1.22 | 1.18 |
| C18:3n6 | 0.29 | 0.10 | 0.19 | 1.04 | 1.59 | 1.45 | 0.15 | 0.12 | 0.13 | 0.13 | 0.12 | 0.19 |
| C18:3n3 | 3.80 | 2.38 | 2.92 | 24.74 | 23.65 | 22.61 | 2.18 | 1.90 | 2.02 | 1.57 | 1.45 | 1.04 |
| C20:2 | 0.35 | 0.23 | 0.25 | 1.42 | 2.02 | 1.91 | 0.24 | 0.20 | 0.21 | 0.21 | 0.18 | 0.15 |
| C20:3n6 | 0.81 | 0.38 | 0.58 | 2.38 | 2.71 | 2.52 | 0.75 | 0.63 | 0.59 | 0.65 | 0.54 | 0.54 |
| C20:3n3 | 0.45 | 0.36 | 0.42 | 2.74 | 2.90 | 2.63 | 0.35 | 0.31 | 0.34 | 0.24 | 0.29 | 0.41 |
| C20:4n6 | 1.15 | 1.41 | 0.91 | 5.20 | 5.05 | 5.54 | 1.82 | 1.42 | 1.45 | 1.64 | 1.57 | 1.53 |
| C22:2 | 0.98 | 0.00 | 0.00 | 4.48 | 4.30 | 4.37 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| C20:5n3 | 1.02 | 0.00 | 0.00 | 6.40 | 5.71 | 5.20 | 0.97 | 0.95 | 0.58 | 1.25 | 1.18 | 1.17 |
| C22:6n3 | 1.90 | 2.63 | 1.39 | 13.88 | 9.83 | 9.70 | 3.47 | 2.66 | 2.58 | 3.07 | 2.39 | 2.34 |
| Saturated fatty acid (SFA) | 12.58 ± 1.05 b | 14.89 ± 0.92 ab | 16.20 ± 0.94 a | 87.20 ± 1.89 b | 93.65 ± 2.25 ab | 96.93 ± 2.53 a | 13.78 ± 0.98 b | 14.14 ± 1.23 b | 20.13 ± 0.62 a | 10.21 ± 0.53 b | 10.55 ± 0.67 b | 16.11 ± 1.08 a |
| Monounsaturated fatty acid (MUFA) | 21.55 ± 0.93 a | 21.49 ± 0.85 a | 15.47 ± 0.84 b | 123.25 ± 2.91 a | 109.77 ± 1.58 b | 105.93 ± 2.82 b | 11.09 ± 0.31 a | 9.87 ± 0.57 ab | 9.59 ± 0.41 b | 7.42 ± 0.69 | 6.91 ± 0.76 | 6.62 ± 0.26 |
| Polyunsaturated fatty acid (PUFA) | 13.70 ± 0.92 a | 9.39 ± 0.83 b | 8.97 ± 0.70 b | 76.59 ± 1.18 a | 75.70 ± 1.09 ab | 72.23 ± 1.41 b | 11.91 ± 0.32 a | 9.94 ± 0.29 b | 9.69 ± 0.34 b | 10.41 ± 0.45 a | 8.99 ± 0.39 b | 8.64 ± 0.46 b |
| RI | Compounds | Skin | Belly | Mince | Washed Mince | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D0 | D7 | D14 | D0 | D7 | D14 | D0 | D7 | D14 | D0 | D7 | D14 | ||
| Alcohols | |||||||||||||
| 1256 | 1-pentanol | 0.014 ± 0.004 a | 0.022 ± 0.005 a | N.D. | 0.057 ± 0.014 b | 0.259 ± 0.052 a | 0.190 ± 0.064 a | 0.019 ± 0.009 | 0.025 ± 0.005 | 0.056 ± 0.034 | N.D. | N.D. | N.D. |
| 1360 | 1-hexanol | 0.040 ± 0.012 b | 0.174 ± 0.047 a | 0.198 ± 0.027 a | 0.491 ± 0.259 b | 1.642 ± 0.065 a | 1.432 ± 0.055 a | 0.199 ± 0.154 b | 0.319 ± 0.049 ab | 0.524 ± 0.039 a | 0.039 ± 0.010 | 0.050 ± 0.017 | 0.053 ± 0.017 |
| 1456 | 1-octen-3-ol | 0.029 ± 0.017 c | 0.225 ± 0.004 b | 0.285 ± 0.012 a | 0.131 ± 0.059 b | 0.926 ± 0.119 a | 1.107 ± 0.520 a | 0.048 ± 0.006 b | 0.099 ± 0.013 ab | 0.175 ± 0.073 a | 0.041 ± 0.013 | 0.052 ± 0.028 | 0.048 ± 0.022 |
| 1460 | 1-heptanol | 0.018 ± 0.001 b | 0.095 ± 0.031 a | 0.065 ± 0.015 ab | 0.049 ± 0.031 b | 0.162 ± 0.010 a | 0.175 ± 0.042 a | 0.020 ± 0.008 b | 0.029 ± 0.003 b | 0.064 ± 0.009 a | 0.012 ± 0.007 | 0.016 ± 0.002 | 0.018 ± 0.001 |
| 1488 | 1,5-octadien-3-ol, (Z)- | N.D. | N.D. | N.D. | N.D. | 0.171 ± 0.022 a | 0.279 ± 0.139 a | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. |
| 1492 | 2-Ethyl-1-hexanol | 0.062 ± 0.015 ab | 0.073 ± 0.020 a | 0.033 ± 0.014 b | 0.388 ± 0.232 | 0.354 ± 0.0353 | 0.396 ± 0.068 | N.D. | 0.111 ± 0.049 a | 0.042 ± 0.035 b | 0.230 ± 0.039 a | 0.037 ± 0.015 b | N.D. |
| 1562 | 1-octanol | 0.038 ± 0.009 b | 0.153 ± 0.029 a | 0.101 ± 0.023 a | 0.018 ± 0.013 b | 0.073 ± 0.004 a | 0.068 ± 0.028 a | N.D. | 0.028 ± 0.015 a | 0.021 ± 0.006 a | 0.007 ± 0.001 a | 0.003 ± 0.001 b | 0.006 ± 0.001 a |
| 1621 | 2-octenol | 0.007 ± 0.002 b | 0.031 ± 0.002 a | 0.025 ± 0.011 a | 0.021 ± 0.004 | 0.048 ± 0.008 | 0.087 ± 0.012 | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. |
| 1665 | 1-nonanol | 0.024 ± 0.008 b | 0.149 ± 0.035 a | 0.091 ± 0.034 ab | 0.041 ± 0.003 | 0.371 ± 0.005 | 0.027 ± 0.011 | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. |
| 1689 | 2-octyn-1-ol | N.D. | 0.039 ± 0.003 a | 0.014 ± 0.002 b | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. |
| 1777 | 2-ethyl-1-hexanol | N.D. | N.D. | N.D. | N.D. | 0.044 ± 0.004 | 0.041 ± 0.003 | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. |
| 1842 | 3-octen-1-ol, (E)- | N.D. | N.D. | N.D. | N.D. | 0.053 ± 0.010 | 0.064 ± 0.007 | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. |
| Aldehydes | |||||||||||||
| 1081 | hexanal | 0.019 ± 0.024 | 0.029 ± 0.007 | 0.027 ± 0.003 | 0.087 ± 0.034 b | 0.144 ± 0.054 b | 0.440 ± 0.135 a | N.D. | N.D. | N.D. | 0.014 ± 0.012 b | 0.033 ± 0.005 b | 0.055 ± 0.005 a |
| 1281 | octanal | N.D. | 0.070 ± 0.009 a | 0.026 ± 0.004 b | N.D. | 0.027 ± 0.001 b | 0.061 ± 0.018 a | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. |
| 1392 | nonanal | 0.032 ± 0.008 c | 0.161 ± 0.036 b | 0.242 ± 0.029 a | 0.041 ± 0.008 b | 0.063 ± 0.007 b | 0.102 ± 0.017 a | 0.022 ± 0.007 b | 0.039 ± 0.003 ab | 0.064 ± 0.014 a | 0.013 ± 0.012 | 0.019 ± 0.003 | 0.020 ± 0.008 |
| 1430 | 2-octenal, (E)- | N.D. | N.D. | N.D. | N.D. | 0.035 ± 0.006 b | 0.073 ± 0.010 a | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. |
| Ketones | |||||||||||||
| 1320 | 2,3-octanedione | N.D. | 0.023 ± 0.002 b | 0.035 ± 0.007 a | 0.049 ± 0.003 b | 0.234 ± 0.053 a | 0.291 ± 0.089 a | N.D. | 0.037 ± 0.008 b | 0.142 ± 0.028 a | N.D. | N.D. | N.D. |
| 1576 | 3,5-octadien-2-one, (E,E)- | N.D. | N.D. | N.D. | N.D. | N.D. | 0.024 ± 0.003 | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. |
| Wavenumber (cm−1) | Band Assignment | Skin | Belly | Mince | Washed Mince | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D0 | D7 | D14 | D0 | D7 | D14 | D0 | D7 | D14 | D0 | D7 | D14 | ||
| 3015 | cis-olefinic group =CH stretching | 7.374 ± 0.14 | 7.260 ± 0.11 | 7.279 ± 0.06 | 7.763 ± 0.30 a | 7.525 ± 0.22 b | 7.151 ± 0.18 c | 8.463 ± 0.11 a | 8.098 ± 0.23 ab | 7.964 ± 0.23 b | 9.187 ± 0.14 a | 9.044 ± 0.10 ab | 8.840 ± 0.10 b |
| ∑methylene groups (2935, 2850,1438, 1301) | 56.450 ± 0.51 | 56.003 ± 0.21 | 55.856 ± 0.79 | 55.157 ± 0.45 | 54.306 ± 0.84 | 54.133 ± 0.31 | 54.496 ± 0.41 | 53.930 ± 0.53 | 53.546 ± 0.64 | 51.845 ± 0.34 a | 50.632 ± 0.50 b | 50.645 ± 0.79 b | |
| 2935 | νas CH2 | 9.805 | 9.827 | 9.783 | 9.545 | 9.510 | 9.401 | 9.538 | 9.543 | 9.422 | 9.753 | 9.579 | 9.560 |
| 2850 | νs CH2 | 32.890 | 32.591 | 32.487 | 31.604 | 31.140 | 31.398 | 31.717 | 31.504 | 31.306 | 29.675 | 28.614 | 28.648 |
| 1438 | δ(CH2) | 9.380 | 9.227 | 9.229 | 9.461 | 9.073 | 8.931 | 8.856 | 8.666 | 8.676 | 8.468 | 8.501 | 8.504 |
| 1301 | t CH2 in-phase twisting | 4.375 | 4.358 | 4.357 | 4.547 | 4.583 | 4.403 | 4.384 | 4.217 | 4.143 | 3.949 | 3.938 | 3.933 |
| 1747 | ν(C=O) carbonyl compounds | 1.421 ± 0.01 b | 1.526 ± 0.07 ab | 1.576 ± 0.03 a | 1.519 ± 0.04 b | 1.575 ± 0.14 b | 1.606 ± 0.07 a | 1.255 ± 0.12 b | 1.455 ± 0.03 a | 1.449 ± 0.04 a | 0.991 ± 0.04 b | 1.078 ± 0.09 ab | 1.152 ± 0.04 a |
| 1658 | ν(C=C) conjugated double bonds | 11.119 ± 0.52 | 11.175 ± 0.60 | 10.901 ± 0.29 | 11.606 ± 0.41 a | 11.210 ± 0.11 ab | 10.739 ± 0.07 b | 11.790 ± 0.38 a | 11.287 ± 0.07 ab | 10.994 ± 0.12 b | 11.853 ± 0.22 | 11.874 ± 0.28 | 11.725 ± 0.11 |
| 1267 | δ(=CH) symmetric rock (cis) | 2.541 ± 0.10 | 2.517 ± 0.23 | 2.497 ± 0.19 | 3.503 ± 07 a | 3.231 ± 0.09 ab | 3.051 ± 0.17 b | 3.549 ± 0.02 a | 3.346 ± 0.08 ab | 3.072 ± 0.04 b | 3.895 ± 0.09 | 3.867 ± 0.07 | 3.701 ± 0.07 |
| Wavenumber (cm−1) | Band Assignment | Skin | Belly | Mince | Washed Mince | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D0 | D7 | D14 | D0 | D7 | D14 | D0 | D7 | D14 | D0 | D7 | D14 | ||
| 3013 | Olefinic ν(=C-H) (cis) | 2.685 ± 0.10 a | 2.538 ± 0.04 ab | 2.508 ± 0.10 b | 2.980 ± 0.12 a | 2.743 ± 0.15 ab | 2.635 ± 0.09 b | 3.203 ± 0.12 a | 3.035 ± 0.35 a | 2.895 ± 0.31 a | 3.558 ± 0.20 a | 3.243 ± 0.17 ab | 2.913 ± 0.22 b |
| ∑methylene groups (2924, 2853, 722) | 32.507 ± 0.57 | 32.057 ± 0.56 | 32.065 ± 0.79 | 32.636 ± 0.23 | 32.141 ± 0.24 | 32.201 ± 0.42 | 32.516 ± 0.40 | 32.665 ± 0.76 | 31.583 ± 0.90 | 31.453 ± 0.51 a | 30.818 ± 0.39 ab | 29.991 ± 0.96 b | |
| 2924 | νas(CH2) | 15.472 | 15.146 | 15.301 | 16.093 | 15.672 | 15.777 | 15.587 | 15.483 | 15.209 | 14.966 | 15.091 | 14.640 |
| 2853 | νs(CH2) | 12.019 | 11.968 | 11.745 | 12.038 | 12.000 | 12.322 | 11.457 | 12.221 | 11.457 | 11.294 | 11.217 | 11.002 |
| 722 | -(CH2)- rocking | 5.017 | 4.943 | 5.020 | 4.504 | 4.469 | 4.101 | 5.472 | 4.961 | 4.917 | 5.193 | 4.510 | 4.349 |
| 1745 | Ester ν(C=O) | 27.268 ± 0.11 c | 27.712 ± 0.11 b | 28.103 ± 0.05 a | 23.014 ± 0.23 b | 25.175 ± 0.86 ab | 25.455 ± 0.85 a | 23.292 ± 1.21 b | 24.068 ± 1.01 ab | 25.685 ± 1.16 a | 21.069 ± 0.32 b | 21.687 ± 0.26 ab | 22.029 ± 0.11 a |
| 970 | -HC=CH-(trans) isolated double bonds | 0.735 ± 0.04 | 0.755 ± 0.07 | 0.728 ± 0.09 | 1.693 ± 0.08 b | 1.827 ± 0.12 b | 2.188 ± 0.11 a | 2.074 ± 0.23 | 2.345 ± 0.13 | 2.396 ± 0.21 | 4.105 ± 0.19 | 4.111 ± 0.22 | 4.605 ± 0.19 |
| ∑phosphate (925, 825) | 1.104 ± 0.05 | 1.169 ± 0.08 | 1.128 ± 0.17 | 2.050 ± 0.35 | 1.866 ± 0.13 | 1.883 ± 0.23 | 1.598 ± 0.05 a | 1.453 ± 0.04 b | 1.458 ± 0.04 b | 3.815 ± 0.12 a | 3.203 ± 0.09 b | 2.962 ± 0.13 b | |
| 925 | νs(P-O-C) | 0.717 | 0.889 | 0.605 | 1.326 | 1.213 | 1.282 | 1.296 | 1.183 | 1.171 | 1.957 | 1.934 | 1.947 |
| 825 | νas(P-O-C) | 0.386 | 0.280 | 1.047 | 0.724 | 0.653 | 0.601 | 0.302 | 0.270 | 0.287 | 1.858 | 1.269 | 1.014 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kunyaboon, S.; Thumanu, K.; Park, J.W.; Khongla, C.; Yongsawatdigul, J. Evaluation of Lipid Oxidation, Volatile Compounds and Vibrational Spectroscopy of Silver Carp (Hypophthalmichthys molitrix) during Ice Storage as Related to the Quality of Its Washed Mince. Foods 2021, 10, 495. https://doi.org/10.3390/foods10030495
Kunyaboon S, Thumanu K, Park JW, Khongla C, Yongsawatdigul J. Evaluation of Lipid Oxidation, Volatile Compounds and Vibrational Spectroscopy of Silver Carp (Hypophthalmichthys molitrix) during Ice Storage as Related to the Quality of Its Washed Mince. Foods. 2021; 10(3):495. https://doi.org/10.3390/foods10030495
Chicago/Turabian StyleKunyaboon, Sasinee, Kanjana Thumanu, Jae W. Park, Chompoonuch Khongla, and Jirawat Yongsawatdigul. 2021. "Evaluation of Lipid Oxidation, Volatile Compounds and Vibrational Spectroscopy of Silver Carp (Hypophthalmichthys molitrix) during Ice Storage as Related to the Quality of Its Washed Mince" Foods 10, no. 3: 495. https://doi.org/10.3390/foods10030495
APA StyleKunyaboon, S., Thumanu, K., Park, J. W., Khongla, C., & Yongsawatdigul, J. (2021). Evaluation of Lipid Oxidation, Volatile Compounds and Vibrational Spectroscopy of Silver Carp (Hypophthalmichthys molitrix) during Ice Storage as Related to the Quality of Its Washed Mince. Foods, 10(3), 495. https://doi.org/10.3390/foods10030495