
Health Promoting Properties of Cereal Vinegars

 ,
, 
 ,
,
Abstract
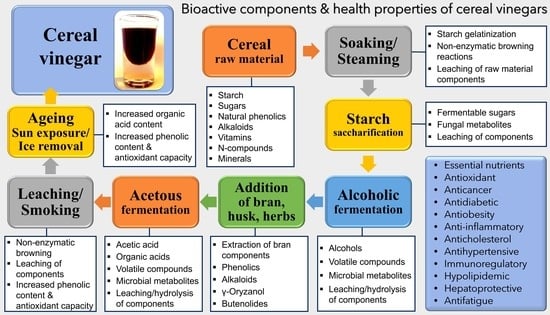
1. Introduction
2. Health Benefits of Cereal Vinegars
2.1. Antioxidant Properties
2.1.1. Antioxidant Components
2.1.2. Antioxidant Activity and Protection against Oxidative Stress In Vitro and In Vivo
2.2. Antimicrobial Properties
2.3. Antidiabetic Properties
2.4. Regulation of Lipid Metabolism and Antiobesity Properties
2.5. Anticancer Properties
2.6. Antihypertensive Properties
2.7. Effect on Neurodegenerative Diseases
2.8. Vinegar Side Effects
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lim, S.J.; Ho, C.W.; Lazim, A.M.; Fazry, S. History and current issues of vinegar. In Advances in Vinegar Production, 1st ed.; Bekatorou, A., Ed.; CRC Press: Boca Raton, FL, USA, 2019; pp. 1–18. [Google Scholar] [CrossRef]
- Lazim, A.M.; Lim, S.J.; Ho, C.W.; Fazry, S. Types of vinegars. In Advances in Vinegar Production, 1st ed.; Bekatorou, A., Ed.; CRC Press: Boca Raton, FL, USA, 2019; pp. 19–28. [Google Scholar] [CrossRef]
- Kandylis, P. Innovative vinegar products. In Advances in Vinegar Production, 1st ed.; Bekatorou, A., Ed.; CRC Press: Boca Raton, FL, USA, 2019; pp. 265–298. [Google Scholar] [CrossRef]
- Zhang, Q.; Fu, C.; Zhao, C.; Yang, S.; Zheng, Y.; Xia, M.; Yan, Y.; Lang, F.; Wang, M. Monitoring microbial succession and metabolic activity during manual and mechanical solid-state fermentation of Chinese cereal vinegar. LWT 2020, 133, 109868. [Google Scholar] [CrossRef]
- Li, W.; Fan, G.; Fu, Z.; Wang, W.; Xu, Y.; Teng, C.; Zhang, C.; Yang, R.; Sun, B.; Li, X. Effects of fortification of Daqu with various yeasts on microbial community structure and flavor metabolism. Food Res. Int. 2020, 129, 108837. [Google Scholar] [CrossRef] [PubMed]
- Nie, Z.; Zheng, Y.; Xie, S.; Zhang, X.; Song, J.; Xia, M.; Wang, M. Unraveling the correlation between microbiota succession and metabolite changes in traditional Shanxi aged vinegar. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Wu, L.H.; Lu, Z.M.; Zhang, X.J.; Wang, Z.M.; Yu, Y.J.; Shi, J.S.; Xu, Z.H. Metagenomics reveals flavour metabolic network of cereal vinegar microbiota. Food Microbiol. 2017, 62, 23–31. [Google Scholar] [CrossRef]
- Ling, J.W.A.; Mun, S.L.; Fazry, S.; Lazim, A.M.; Lim, S.J. Health benefits of vinegars. In Advances in Vinegar Production, 1st ed.; Bekatorou, A., Ed.; CRC Press: Boca Raton, FL, USA, 2019; pp. 379–408. [Google Scholar] [CrossRef]
- Budak, N.H.; Aykin, E.; Seydim, A.C.; Greene, A.K.; Guzel-Seydim, Z.B. Functional properties of vinegar. J. Food Sci. 2014, 79, R757–R764. [Google Scholar] [CrossRef]
- Martillanes, S.; Rocha-Pimienta, J.; Gil, M.V.; Ayuso-Yuste, M.C.; Delgado-Adámez, J. Antioxidant and antimicrobial evaluation of rice bran (Oryza sativa L.) extracts in a mayonnaise-type emulsion. Food Chem. 2020, 308, 125633. [Google Scholar] [CrossRef]
- Lemus, C.; Angelis, A.; Halabalaki, M.; Skaltsounis, A.L. γ-Oryzanol. An attractive bioactive component from rice bran. In Wheat and Rice in Disease Prevention and Health; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Academic Press: Waltham, MA, USA, 2014; pp. 409–430. [Google Scholar] [CrossRef]
- Castanho, A.; Lageiro, M.; Calhelha, R.C.; Ferreira, I.C.F.R.; Sokovic, M.; Cunha, L.M.; Brites, C. Exploiting the bioactive properties of γ-oryzanol from bran of different exotic rice varieties. Food Funct. 2019, 10, 2382–2389. [Google Scholar] [CrossRef] [PubMed]
- Hajilou, H.; Farahpour, M.R.; Hamishehkar, H. Polycaprolactone nanofiber coated with chitosan and gamma oryzanol functionalized as a novel wound dressing for healing infected wounds. Int. J. Biol. Macromol. 2020, 164, 2358–2369. [Google Scholar] [CrossRef] [PubMed]
- Lomthong, T.; Saithong, P. Feasibility of Leum Pua glutinous rice substrate for sugar syrup and vinegar production by raw starch degrading enzyme hydrolysis. Int. Food Res. J. 2019, 26, 1515–1523. [Google Scholar]
- Liu, M.E.; Chou, C.H.; Li, L.; Wu, Y.H.S.; Lin, Y.L.; Tu, D.G.; Chen, Y.C. Modulation effects of black-vinegar-based supplement against a high-fat dietary habit: Antiobesity/hypolipidemic, antioxidative, and energy-metabolism effects. J. Sci. Food Agric. 2020, 100, 2380–2388. [Google Scholar] [CrossRef] [PubMed]
- Gong, M.; Zhou, Z.; Yu, Y.; Liu, S.; Zhu, S.; Jian, D.; Cui, P.; Zhong, F.; Mao, J. Investigation of the 5-hydroxymethylfurfural and furfural content of Chinese traditional fermented vinegars from different regions and its correlation with the saccharide and amino acid content. LWT 2020, 124, 109175. [Google Scholar] [CrossRef]
- Ren, M.; Wang, X.; Tian, C.; Li, X.; Zhang, B.; Song, X.; Zhang, J. Characterization of organic acids and phenolic compounds of cereal vinegars and fruit vinegars in China. J. Food Process. Pres. 2017, 41, e12937. [Google Scholar] [CrossRef]
- Zhao, C.; Xia, T.; Du, P.; Duan, W.; Zhang, B.; Zhang, J.; Zhu, S.; Zheng, Y.; Wang, M.; Yu, Y. Chemical composition and antioxidant characteristic of traditional and industrial Zhenjiang aromatic vinegars during the aging process. Molecules 2018, 23, 2949. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Xia, T.; Duan, W.; Zhang, Z.; Li, Y.; Fang, B.; Xia, M.; Wang, M. Effects of organic acids, amino acids and phenolic compounds on antioxidant characteristic of Zhenjiang aromatic vinegar. Molecules 2019, 24, 3799. [Google Scholar] [CrossRef]
- Zhao, G.; Kuang, G.; Li, J.; Hadiatullah, H.; Chen, Z.; Wang, X.; Yao, Y.; Pan, Z.H.; Wang, Y. Characterization of aldehydes and hydroxy acids as the main contribution to the traditional Chinese rose vinegar by flavor and taste analyses. Food Res. Int. 2020, 129, 108879. [Google Scholar] [CrossRef]
- Zheng, Y.; Cheng, C.; Liu, J.; Liu, X.; Xie, X.; Xu, Y.; Song, J.; Wang, M. Analysis of main characteristic flavor components in Chinese traditional solid-state fermented vinegars. J. Chin. Inst. Food Sci. Tech. 2020, 20, 237–247. [Google Scholar]
- Xia, T.; Zhang, B.; Duan, W.; Zhang, J.; Wang, M. Nutrients and bioactive components from vinegar: A fermented and functional food. J. Funct. Foods 2020, 64, 103681. [Google Scholar] [CrossRef]
- Fan, J.; Zhang, Y.; Zhou, L.; Li, Z.; Zhang, B.; Saito, M.; Wang, X. Nutritional composition and α-glucosidase inhibitory activity of five Chinese vinegars. Jpn. Agr. Res. Q. JARQ 2011, 45, 445–456. [Google Scholar] [CrossRef]
- Xie, X.; Zheng, Y.; Liu, X.; Cheng, C.; Zhang, X.; Xia, T.; Yu, S.; Wang, M. Antioxidant activity of Chinese Shanxi aged vinegar and its correlation with polyphenols and flavonoids during the brewing process. J. Food Sci. 2017, 82, 2479–2486. [Google Scholar] [CrossRef]
- Kim, H.; Lee, H.; Shin, K.S. Intestinal immunostimulatory activity of neutral polysaccharide isolated from traditionally fermented Korean brown rice vinegar. Biosci. Biotechnol. Biochem. 2016, 80, 2383–2390. [Google Scholar] [CrossRef]
- Li, J.; Yu, G.; Fan, J. Alditols and monosaccharides from sorghum vinegar can attenuate platelet aggregation by inhibiting cyclooxygenase-1 and thromboxane-A2 synthase. J. Ethnopharmacol. 2014, 155, 285–292. [Google Scholar] [CrossRef]
- Chiu, H.F.; Cheng, Y.; Lu, Y.Y.; Han, Y.C.; Shen, Y.C.; Venkatakrishnan, K.; Wang, C.K. Anti-mutagenicity, hypouricemic and antioxidant activities of alkaloids from vinegar and mei vinegar. J. Food Biochem. 2017, 41, e12373. [Google Scholar] [CrossRef]
- Chou, C.H.; Liu, C.W.; Yang, D.J.; Wu, Y.H.S.; Chen, Y.C. Amino acid, mineral, and polyphenolic profiles of black vinegar, and its lipid lowering and antioxidant effects in vivo. Food Chem. 2015, 168, 63–69. [Google Scholar] [CrossRef]
- Phuapaiboon, P. Gamma-aminobutyric acid, total anthocyanin content and antioxidant activity of vinegar brewed from germinated pigmented rice. Pakistan J. Nutr. 2017, 16, 109–118. [Google Scholar] [CrossRef][Green Version]
- Xia, T.; Duan, W.; Zhang, Z.; Fang, B.; Zhang, B.; Xu, B.; de la Cruz, C.B.V.; El-Seedi, H.; Simal-Gandara, J.; Wang, S.; et al. Polyphenol-rich extract of Zhenjiang aromatic vinegar ameliorates high glucose-induced insulin resistance by regulating JNK-IRS-1 and PI3K/Akt signaling pathways. Food Chem. 2021, 335, 127513. [Google Scholar] [CrossRef]
- Duan, W.; Xia, T.; Zhang, B.; Li, S.; Zhang, C.; Zhao, C.; Song, J.; Wang, M. Changes of physicochemical, bioactive compounds and antioxidant capacity during the brewing process of Zhenjiang aromatic vinegar. Molecules 2019, 24, 3935. [Google Scholar] [CrossRef]
- Chen, H.; Zhou, Y.; Shao, Y.; Chen, F. Free phenolic acids in Shanxi aged vinegar: Changes during aging and synergistic antioxidant activities. Int. J. Food Prop. 2016, 19, 1183–1193. [Google Scholar] [CrossRef]
- Xia, T.; Zhang, B.; Duan, W.; Li, Y.; Zhang, J.; Song, J.; Zheng, Y.; Wang, M. Hepatoprotective efficacy of Shanxi aged vinegar extract against oxidative damage in vitro and in vivo. J. Funct. Foods 2019, 60. [Google Scholar] [CrossRef]
- Ho, C.W.; Lazim, A.M.; Fazry, S.; Zaki, U.K.H.H.; Lim, S.J. Varieties, production, composition and health benefits of vinegars: A review. Food Chem. 2017, 221, 1621–1630. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.T.; Huang, Q.; Wu, G.L.; Chen, C.G.; Li, Z.J. Evaluation of the antioxidant property and effects in Caenorhabditis elegans of Xiangxi flavor vinegar, a Hunan local traditional vinegar. J. Zhejiang Univ. Sci. B 2017, 18, 324–333. [Google Scholar] [CrossRef]
- Bakir, S.; Devecioglu, D.; Kayacan, S.; Toydemir, G.; Karbancioglu-Guler, F.; Capanoglu, E. Investigating the antioxidant and antimicrobial activities of different vinegars. Eur. Food Res. Technol. 2017, 243, 2083–2094. [Google Scholar] [CrossRef]
- Sangngern, N.; Puangnark, T.; Nguansangiam, W.; Saithong, P.; Kitpreechavanich, V.; Lomthong, T. Production and development of vinegar fermentation from broken Riceberry rice using raw starch-degrading enzyme hydrolysis. 3 Biotech. 2020, 10, 515. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Zhou, J.; Zhang, L.; Zhang, J.; Li, N.; Zhao, C.; Tu, L.; Zheng, Y.; Xia, T.; Luo, J.; et al. GC × GC-MS analysis and hypolipidemic effects of polyphenol extracts from Shanxi-aged vinegar in rats under a high fat diet. Food Funct. 2020, 11, 7468–7480. [Google Scholar] [CrossRef]
- Xia, T.; Zhang, J.; Yao, J.; Zhang, B.; Duan, W.; Xia, M.; Song, J.; Zheng, Y.; Wang, M. Shanxi aged vinegar prevents alcoholic liver injury by inhibiting CYP2E1 and NADPH oxidase activities. J. Funct. Foods 2018, 47, 575–584. [Google Scholar] [CrossRef]
- Suzuki, E.; Otake, S.; Hamadate, N.; Hasumi, K. Kurozu melanoidin, a novel oligoglucan-melanoidin complex from Japanese black vinegar, suppresses adipogenesis in vitro. J. Funct. Foods 2020, 72, 104046. [Google Scholar] [CrossRef]
- Chen, H.; Wang, S.; Fu, H.; Chen, F.; Zhang, L.; Lan, W.; Yang, J.; Yang, X.; She, Y. A colorimetric sensor array for recognition of 32 Chinese traditional cereal vinegars based on “turn-off/on” fluorescence of acid-sensitive quantum dots. Spectrochim. Acta-Part A Mol. Biomol. Spectrosc. 2020, 227, 117683. [Google Scholar] [CrossRef]
- Chai, L.J.; Qiu, T.; Lu, Z.M.; Deng, Y.J.; Zhang, X.J.; Shi, J.S.; Xu, Z.H. Modulating microbiota metabolism via bioaugmentation with Lactobacillus casei and Acetobacter pasteurianus to enhance acetoin accumulation during cereal vinegar fermentation. Food Res. Int. 2020, 138, 109737. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Xu, Q.; Chen, J.; Lu, Z.; Xia, R.; Li, G.; Xu, Z.; Ma, Y. Ligustrazine formation in Zhenjiang aromatic vinegar: Changes during fermentation and storing process. J. Sci. Food Agr. 2011, 91, 1612–1617. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tian, J.; Ge, H.; Liu, R.; Xiao, J. Effects of tetramethylpyrazine from Chinese black vinegar on antioxidant and hypolipidemia activities in HepG2 cells. Food Chem. Toxicol. 2017, 109, 930–940. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, J.; Zhou, R.; Wu, C. Evaluating the feasibility of fermentation starter inoculated with Bacillus amyloliquefaciens for improving acetoin and tetramethylpyrazine in Baoning bran vinegar. Int. J. Food Microbiol. 2017, 255, 42–50. [Google Scholar] [CrossRef]
- Zhou, Z.; Jian, D.; Gong, M.; Zhu, S.; Li, G.; Zhang, S.; Zhong, F.; Mao, J. Characterization of the key aroma compounds in aged Zhenjiang aromatic vinegar by gas chromatography-olfactometry-mass spectrometry, quantitative measurements, aroma recombination and omission experiments. Food Res. Int. 2020, 136, 109434. [Google Scholar] [CrossRef]
- Yatmaz, A.H.; Kinoshita, T.; Miyazato, A.; Takagi, M.; Tsujino, Y.; Beppu, F.; Gotoh, N. Quantification of Fraglide-1, a new functional ingredient, in vinegars. J. Oleo Sci. 2017, 66, 1381–1386. [Google Scholar] [CrossRef]
- Zhang, C.; Li, D.; Zhang, C.; Li, H.; Yin, K.; Zhang, X.; Zhang, D. γ-Oryzanol content and ferulic acid ester composition in brown rice from Heilongjiang Province. Food Sci. 2020, 41, 234–241. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A.; Iordache, F.; Stanca, L.; Predoi, G.; Serban, A.I. Oxidative stress mitigation by antioxidants—An overview on their chemistry and influences on health status. Eur. J. Med. Chem. 2021, 209, 112891. [Google Scholar] [CrossRef]
- Maes, M.; Galecki, P.; Chang, Y.S.; Berk, M. A review on the oxidative and nitrosative stress (OandNS) pathways in major depression and their possible contribution to the (neuro) degenerative processes in that illness. Prog. Neuro Psychoph. 2011, 35, 676–692. [Google Scholar] [CrossRef]
- Liu, J.; Gan, J.; Nirasawa, S.; Zhou, Y.; Xu, J.; Zhu, S.; Cheng, Y. Cellular uptake and trans-enterocyte transport of phenolics bound to vinegar melanoidins. J. Funct. Foods 2017, 37, 632–640. [Google Scholar] [CrossRef]
- Shimoji, Y.; Tamura, Y.; Nakamura, Y.; Nanda, K.; Nishidai, S.; Nishikawa, Y.; Ohigashi, H. Isolation and identification of DPPH radical scavenging compounds in Kurosu (Japanese unpolished rice vinegar). J. Agr. Food Chem. 2002, 50, 6501–6503. [Google Scholar] [CrossRef] [PubMed]
- Shimoji, Y.; Kohno, H.; Nanda, K.; Nishikawa, Y.; Ohigashi, H.; Uenakai, K.; Tanaka, T. Extract of Kurosu, a vinegar from unpolished rice, inhibits azoxymethane-induced colon carcinogenesis in male F344 rats. Nutr. Cancer 2004, 49, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Shibayama, Y.; Kanouchi, H.; Fujii, A.; Nagano, M. A review of Kurozu, amber rice vinegar made in pottery jars. Funct. Foods Health Dis. 2020, 10, 254–264. [Google Scholar] [CrossRef]
- Nanda, K.; Miyoshi, N.; Nakamura, Y.; Shimoji, Y.; Tamura, Y.; Nishikawa, Y.; Uenakai, K.; Kohno, H.; Tanaka, T. Extract of vinegar “Kurosu” from unpolished rice inhibits the proliferation of human cancer cells. J. Exp. Clin. Cancer Res. 2004, 23, 69–76. [Google Scholar] [PubMed]
- Nagashima, M.; Saito, K. Antioxidant activity of the new black vinegar “iZUMI”. J. Nutr. Health Aging 2010, 14, 845–849. [Google Scholar] [CrossRef]
- Xia, T.; Yao, J.H.; Zhang, J.; Duan, W.H.; Zhang, B.; Xie, X.L.; Xia, M.L.; Song, J.; Zheng, Y.; Wang, M. Evaluation of nutritional compositions, bioactive compounds, and antioxidant activities of Shanxi aged vinegars during the aging process. J. Food Sci. 2018, 83, 2638–2644. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Jo, Y.; Chung, N.; Gu, S.-Y.; Jeong, Y.-J.; Kwon, J.-H. Physicochemical qualities and flavor patterns of traditional Chinese vinegars manufactured by different fermentation methods and aging periods. Prev. Nutr. Food Sci. 2017, 22, 30–36. [Google Scholar] [CrossRef]
- Choi, H.; Gwak, G.; Choi, D.; Park, J.; Cheong, H. Antimicrobial efficacy of fermented dark vinegar from unpolished rice. Microbiol. Biotechnol. Lett. 2015, 43, 97–104. [Google Scholar] [CrossRef]
- Yu, X.; Yang, M.; Dong, J.; Shen, R. Comparative analysis of the antioxidant capacities and phenolic compounds of oat and buckwheat vinegars during production processes. J. Food Sci. 2018, 83, 844–853. [Google Scholar] [CrossRef] [PubMed]
- Pazuch, C.M.; Kalschne, D.L.; Siepmann, F.B.; Marx, I.M.G.; de Oliveira, T.C.G.; Spinosa, W.A.; Canan, C.; Colla, E. Optimization and characterization of vinegar produced from rice bran. Food Sci. Technol. 2020, 40, 608–613. [Google Scholar] [CrossRef]
- Li, Y.; He, Y.; Hu, H.; Li, H.; Hu, J.; Cheng, Z. Study of antioxidant, thrombolytic and antithrombus activities of Tartary buckwheat vinegar. J. Chin. Inst. Food Sci. Technol. 2018, 18, 46–51. [Google Scholar] [CrossRef]
- Mudura, E.; Coldea, T.E.; Socaciu, C.; Ranga, F.; Pop, C.R.; Rotar, A.M.; Pasqualone, A. Brown beer vinegar: A potentially functional product based on its phenolic profile and antioxidant activity. J. Serb. Chem. Soc. 2018, 83, 19–30. [Google Scholar] [CrossRef]
- Ho, C.W.; Lazim, A.M.; Fazry, S.; Hussain Zaki, U.M.K.H.; Lim, S.J. Effects of fermentation time and pH on soursop (Annona muricata) vinegar production towards its chemical compositions. Sains Malays. 2017, 46, 1505–1512. [Google Scholar] [CrossRef]
- Ali, Z.; Wang, Z.; Amir, R.M.; Younas, S.; Wali, A.; Adowa, N.; Ayim, I. Potential uses of vinegar as a medicine and related in vivo mechanisms. Int. J. Vitam. Nutr. Res. 2016, 86, 127–151. [Google Scholar] [CrossRef]
- Bouarab-Chibane, L.; Forquet, V.; Lantéri, P.; Clément, Y.; Léonard-Akkari, L.; Oulahal, N.; Degraeve, P.; Bordes, C. Antibacterial properties of polyphenols: Characterization and QSAR (Quantitative Structure–Activity Relationship) models. Front. Microbiol. 2019, 10, 829. [Google Scholar] [CrossRef] [PubMed]
- Peh, E.; Kittler, S.; Reich, F.; Kehrenberg, C. Antimicrobial activity of organic acids against Campylobacter spp. and development of combinations—A synergistic effect? PLoS ONE 2020, 15, e0239312. [Google Scholar] [CrossRef] [PubMed]
- Kadiroğlu, P. FTIR spectroscopy for prediction of quality parameters and antimicrobial activity of commercial vinegars with chemometrics. J. Sci. Food Agric. 2018, 98, 4121–4127. [Google Scholar] [CrossRef]
- Carpenter, C.E.; Broadbent, J.R. External concentration of organic acid anions and pH: Key independent variables for studying how organic acids inhibit growth of bacteria in mildly acidic foods. J. Food Sci. 2009, 74, R12–R15. [Google Scholar] [CrossRef]
- Mani-López, E.; García, H.S.; López-Malo, A. Organic acids as antimicrobials to control Salmonella in meat and poultry products. Food Res. Int. 2012, 45, 713–721. [Google Scholar] [CrossRef]
- Pinto, L.; Caputo, L.; Quintieri, L.; de Candia, S.; Baruzzi, F. Efficacy of gaseous ozone to counteract postharvest table grape sour rot. Food Microbiol. 2017, 66, 190–198. [Google Scholar] [CrossRef]
- Pinto, L.; Malfeito-Ferreira, M.; Quintieri, L.; Silva, A.C.; Baruzzi, F. Growth and metabolite production of a grape sour rot yeast-bacterium consortium on different carbon sources. J. Food Microbiol. 2019, 296, 65–74. [Google Scholar] [CrossRef]
- Chai, L.J.; Shen, M.N.; Sun, J.; Deng, Y.J.; Lu, Z.M.; Zhang, X.J.; Shi, J.S.; Xu, Z.H. Deciphering the D-/l-lactate-producing microbiota and manipulating their accumulation during solid-state fermentation of cereal vinegar. Food Microbiol. 2020, 92, 103559. [Google Scholar] [CrossRef]
- Wang, Q.; Buchanan, R.L.; Tikekar, R.V. Evaluation of adaptive response in E. coli O157:H7 to UV light and gallic acid based antimicrobial treatments. Food Control 2019, 106, 106723. [Google Scholar] [CrossRef]
- Sinsinwar, S.; Vadivel, V. Catechin isolated from cashew nut shell exhibits antibacterial activity against clinical isolates of MRSA through ROS-mediated oxidative stress. Appl. Microbiol. Biot. 2020, 104, 8279–8297. [Google Scholar] [CrossRef] [PubMed]
- Lekbach, Y.; Dong, Y.; Li, Z.; Xu, D.; El Abed, S.; Yi, Y.; Li, L.; Ibnsouda Koraichi, S.; Sun, T.; Wang, F. Catechin hydrate as an eco-friendly biocorrosion inhibitor for 304 L stainless steel with dual-action antibacterial properties against Pseudomonas aeruginosa biofilm. Corros. Sci. 2019, 157, 98–108. [Google Scholar] [CrossRef]
- Bernal-Mercado, A.T.; Vazquez-Armenta, F.J.; Tapia-Rodriguez, M.R.; Islas-Osuna, M.A.; Mata-Haro, V.; Gonzalez-Aguilar, G.A.; Lopez-Zavala, A.A.; Ayala-Zavala, J.F. Comparison of single and combined use of catechin, protocatechuic, and vanillic acids as antioxidant and antibacterial agents against uropathogenic Escherichia coli at planktonic and biofilm levels. Molecules 2018, 23, 2813. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Fu, Y.; Liu, M.; Wang, T.; Zhang, J.; Yang, M.; Sun, Z.; Li, X.; Li, Y. In vitro antibacterial activity and mechanism of vanillic acid against carbapenem-resistant Enterobacter cloacae. Antibiotics 2019, 8, 220. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Yang, M.; Wang, T.; Sun, Z.; Liu, M.; Zhang, J.; Zeng, Q.; Cai, C.; Li, Y. Antibacterial mechanism of vanillic acid on physiological, morphological, and biofilm properties of carbapenem-resistant Enterobacter hormaechei. J. Food Prot. 2020, 83, 576–583. [Google Scholar] [CrossRef]
- Heidtmann-Bemvenuti, R.; Tralamazza, S.M.; Jorge Ferreira, C.F.; Corrêa, B.; Badiale-Furlong, E. Effect of natural compounds on Fusarium graminearum complex. J. Sci. Food Agric. 2016, 96, 3998–4008. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.M.; Fang, T.J. Survival of Escherichia coli O157:H7 and Salmonella enterica serovars Typhimurium in iceberg lettuce and the antimicrobial effect of rice vinegar against E. coli O157:H7. Food Microbiol. 2007, 24, 745–751. [Google Scholar] [CrossRef]
- Maqbul, M.S.; Alshabi, A.M.; Khan, A.A.; Iqubal, S.S.; Mohammed, T.; Shaikh, I.A.; Dawoud, A.; Muddapur, U.M.; Hussain, M.S.; Singh, S.K. Comparison of e-test values for standard antibiotics and conventional antimicrobial assay values for ethanoic acids against nosocomial multidrug-resistant Pseudomonas aeruginosa. J. Pure Appl. Microbiol. 2020, 14, 255–260. [Google Scholar] [CrossRef]
- Krusong, W.; Sriphochanart, W.; Suwapanich, R.; Mekkerdchoo, O.; Sriprom, P.; Wipatanawin, A.; Massa, S. Healthy dried baby corn silk vinegar production and determination of its main organic volatiles containing antimicrobial activity. LWT 2020, 117, 108620. [Google Scholar] [CrossRef]
- Liu, Y.; Hannig, M. Vinegar inhibits the formation of oral biofilm in situ. BMC Oral Health 2020, 20, 167. [Google Scholar] [CrossRef]
- Ben-Avraham, S.; Harman-Boehm, I.; Schwarzfuchs, D.; Shai, I. Dietary strategies for patients with type 2 diabetes in the era of multi-approaches: Review and results from the Dietary Intervention Randomized Controlled Trial (DIRECT). Diabetes Res. Clin. Pract. 2009, 86, S41–S48. [Google Scholar] [CrossRef]
- Johnston, C.S.; Buller, A.J. Vinegar and peanut products as complementary foods to reduce postprandial glycemia. J. Am. Diet. Assoc. 2005, 105, 1939–1942. [Google Scholar] [CrossRef] [PubMed]
- Johnston, C.S.; Gaas, C.A. Vinegar: Medicinal uses and antiglycemic effect. Med. Gen. Med. 2006, 8, 61. [Google Scholar]
- Santos, H.O.; de Moraes, W.M.; da Silva, G.A.; Prestes, J.; Schoenfeld, B.J. Vinegar (acetic acid) intake on glucose metabolism: A narrative review. Clin. Nutr. ESPEN 2019, 32, 1–7. [Google Scholar] [CrossRef]
- Lim, J.; Henry, C.J.; Haldar, S. Vinegar as a functional ingredient to improve postprandial glycemic control-human intervention findings and molecular mechanisms. Mol. Nutr. Food Res. 2016, 60, 1837–1849. [Google Scholar] [CrossRef] [PubMed]
- Fushimi, T.; Suruga, K.; Oshima, Y.; Fukiharu, M.; Tsukamoto, Y.; Goda, T. Dietary acetic acid reduces serum cholesterol and triacylglycerols in rats fed a cholesterol-rich diet. Br. J. Nutr. 2006, 95, 916–924. [Google Scholar] [CrossRef] [PubMed]
- Fushimi, T.; Sato, Y. Effect of acetic acid feeding on the circadian changes in glycogen and metabolites of glucose and lipid in liver and skeletal muscle of rats. Br. J. Nutr. 2005, 94, 714–719. [Google Scholar] [CrossRef]
- Mitrou, P.; Petsiou, E.; Papakonstantinou, E.; Maratou, E.; Lambadiari, V.; Dimitriadis, P.; Spanoudi, F.; Raptis, S.A.; Dimitriadis, G. Vinegar consumption increases insulin-stimulated glucose uptake by the forearm muscle in humans with type 2 diabetes. J. Diabetes Res. 2015, 2015, 175204. [Google Scholar] [CrossRef] [PubMed]
- Mitrou, P.; Petsiou, E.; Papakonstantinou, E.; Maratou, E.; Lambadiari, V.; Dimitriadis, P.; Spanoudi, F.; Raptis, S.A.; Dimitriadis, G. The role of acetic acid on glucose uptake and blood flow rates in the skeletal muscle in humans with impaired glucose tolerance. Eur. J. Clin. Nutr. 2015, 69, 734. [Google Scholar] [CrossRef]
- Pan, J.H.; Kim, J.H.; Kim, H.M.; Lee, E.S.; Shin, D.H.; Kim, S.; Shin, M.; Kim, S.H.; Lee, J.H.; Kim, Y.J. Acetic acid enhances endurance capacity of exercise-trained mice by increasing skeletal muscle oxidative properties. Biosci. Biotechnol. Biochem. 2015, 79, 1535–1541. [Google Scholar] [CrossRef]
- Ogawa, N.; Satsu, H.; Watanabe, H.; Fukaya, M.; Tsukamoto, Y.; Miyamoto, Y.; Shimizu, M. Acetic acid suppresses the increase in disaccharidase activity that occurs during culture of caco-2 cells. J. Nutr. 2000, 130, 507–513. [Google Scholar] [CrossRef]
- Liljeberg, H.; Björck, I. Delayed gastric emptying rate may explain improved glycaemia in healthy subjects to a starchy meal with added vinegar. Eur. J. Clin. Nutr. 1998, 52, 368–371. [Google Scholar] [CrossRef] [PubMed]
- White, A.M.; Johnston, C.S. Vinegar ingestion at bedtime moderates waking glucose concentrations in adults with well-controlled type 2 diabetes. Diab. Care 2007, 30, 2814–2815. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Papoutsis, K.; Zhang, J.; Bowyer, M.C.; Brunton, N.; Gibney, E.R.; Lyng, J. Fruit, vegetables, and mushrooms for the preparation of extracts with α-amylase and α-glucosidase inhibition properties: A review. Food Chem. 2021, 338, 128119. [Google Scholar] [CrossRef]
- Hua, F.; Zhou, P.; Wu, H.Y.; Chu, G.X.; Xie, Z.W.; Bao, G.H. Inhibition of α-glucosidase and α-amylase by flavonoid glycosides from Lu’an GuaPian tea: Molecular docking and interaction mechanism. Food Funct. 2018, 9, 4173–4183. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Tian, J.; Yang, W.; Chen, S.; Liu, D.; Fang, H.; Zhang, H.; Ye, X. Inhibition mechanism of ferulic acid against α-amylase and α-glucosidase. Food Chem. 2020, 317, 126346. [Google Scholar] [CrossRef]
- Gu, X.; Zhao, H.L.; Sui, Y.; Guan, J.; Chan, J.C.; Tong, P.C. White rice vinegar improves pancreatic beta-cell function and fatty liver in streptozotocin-induced diabetic rats. Acta Diabetol. 2012, 49, 185–191. [Google Scholar] [CrossRef]
- Ma, T.; Shan, F.; Changxi, J.I.A. Effects of Tartary buckwheat vinegar on blood glucose of diabetic mice. J. Chin. Cereals Oils Assoc. 2010, 25, 42–48. [Google Scholar]
- Cheng, L.J.; Jiang, Y.; Wu, V.X.; Wang, W. A systematic review and meta-analysis: Vinegar consumption on glycaemic control in adults with type 2 diabetes mellitus. J. Adv. Nurs. 2020, 76, 459–474. [Google Scholar] [CrossRef]
- Petsiou, E.I.; Mitrou, P.I.; Raptis, S.A.; Dimitriadis, G.D. Effect and mechanisms of action of vinegar on glucose metabolism, lipid profile, and body weight. Nutr. Rev. 2014, 72, 651–661. [Google Scholar] [CrossRef]
- Beheshti, Z.; Chan, Y.H.; Nia, H.S.; Hajihosseini, F.; Nazari, R.; Shaabani, M. Influence of apple cider vinegar on blood lipids. Life Sci. J. 2012, 9, 2431–2440. [Google Scholar]
- Yamashita, H.; Fujisawa, K.; Ito, E.; Idei, S.; Kawaguchi, N.; Kimoto, M.; Hiemori, M.; Tsuji, H. Improvement of obesity and glucose tolerance by acetate in Type 2 diabetic Otsuka Long-Evans Tokushima Fatty (OLETF) rats. Biosci. Biotechnol. Biochem. 2007, 71, 1236–1243. [Google Scholar] [CrossRef] [PubMed]
- Launholt, T.L.; Kristiansen, C.B.; Hjorth, P. Safety and side effects of apple vinegar intake and its effect on metabolic parameters and body weight: A systematic review. Eur. J. Nutr. 2020, 59, 2273–2289. [Google Scholar] [CrossRef]
- Östman, E.; Granfeldt, Y.; Persson, L.; Björck, I. Vinegar supplementation lowers glucose and insulin responses and increases satiety after a bread meal in healthy subjects. Eur. J. Clin. Nutr. 2005, 59, 983–988. [Google Scholar] [CrossRef]
- Chen, J.C.; Zheng, B.D.; Zhao, Y.Y.; Xu, J.; Wu, J.J.; Luo, C.; Pang, J.; He, G.Q. Hypolipidemic effects on lipid metabolism and lipase inhibition by black vinegar powder. J. Biobased Mater. Bioenergy 2012, 6, 634–639. [Google Scholar] [CrossRef]
- Chen, H.; Chen, T.; Giudici, P.; Chen, F. Vinegar functions on health: Constituents, sources, and formation mechanisms. Compr. Rev. Food Sci. Food Saf. 2016, 15, 1124–1138. [Google Scholar] [CrossRef] [PubMed]
- Samad, A.; Azlan, A.; Ismail, A. Therapeutic effects of vinegar: A review. Curr. Opin. Food Sci. 2016, 8, 56–61. [Google Scholar] [CrossRef]
- Hamadate, N.; Nakamura, K.I.; Hirai, M.; Yamamoto, T.; Yamaguchi, H.; Iizuka, M.; Yamamoto, E.; Iwama, Y.; Yazawa, K. Effect of a dietary supplement containing Kurozu (a Japanese traditional health drink) concentrate on several obesity-related parameters in obese Japanese adults: A randomized, double-blind, placebo-controlled trial. Funct. Foods Health Dis. 2013, 3, 310–322. [Google Scholar] [CrossRef]
- Hamadate, N.; Seto, K.; Yazawa, K. Effects of a Dietary Supplement Containing Kurozu Concentrate on Body Fat and Energy Metabolism. Jpn. J. Complem. Altern. Med. 2014, 11, 67–74. [Google Scholar] [CrossRef]
- Nakasone, Y.; Miura, H. Effect of a traditional Japanese health food made from Kurozu (unrefined black rice vinegar) and garlic combination on serum cholesterol in subjects with prehypercholesterolemia or mild-to-moderate hypercholesterolemia-A randomized, double-blind, placebo-controlled, intervention study. Jpn. Pharmacol. Ther. 2017, 45, 51–60. [Google Scholar]
- Abe, S.; Hasegawa, M.; Tsuruoka, J.; Matsumoto, Y.; Koyanagi, S. Effect of dietary supplement containing Kurozu concentrate on visceral fat accumulation. Jpn. J. Complem. Altern. Med. 2019, 16, 13–19. [Google Scholar] [CrossRef]
- Li, B.; Li, Z.; Wei, Y.; Zhang, X.; Wu, R.; Fan, Y.; Bu, L. Study on the effects of brans and Aspergillus niger about corn vinegar on reducing obesity and blood lipids in rat. J. Northwest A F Univ. Nat. Sci. Ed. 2009, 37, 194–198. [Google Scholar]
- Tong, L.T.; Katakura, Y.; Kawamura, S.; Baba, S.; Tanaka, Y.; Udono, M.; Kondo, Y.; Nakamura, K.; Imaizumi, K.; Sato, M. Effects of Kurozu concentrated liquid on adipocyte size in rats. Lipids Health Dis. 2010, 9, 134. [Google Scholar] [CrossRef]
- Liu, L.; Yang, X. Hypolipidemic and antioxidant effects of freeze-dried powder of Shanxi mature vinegar in hyperlipidaemic mice. Food Sci. 2015, 36, 141–145. [Google Scholar]
- Shibayama, Y.; Nagano, M.; Hashiguchi, K.; Fujii, A.; Iseki, K. Supplementation of concentrated Kurozu, a Japanese black vinegar, reduces the onset of hepatic steatosis in mice fed with a high-fat diet. Funct. Foods Health Dis. 2019, 9, 276–296. [Google Scholar] [CrossRef]
- Zhao, Y.; He, Z.; Hao, W.; Zhu, H.; Liang, N.; Liu, J.; Zhang, C.; Ma, K.Y.; He, W.S.; Yang, Y.; et al. Vinegars but not acetic acid are effective in reducing plasma cholesterol in hamsters fed a high-cholesterol diet. Food Funct. 2020, 11, 2163. [Google Scholar] [CrossRef]
- Ruppin, H.; Bar-Meir, S.; Soergel, K.H.; Wood, C.M.; Schmitt, M.G., Jr. Absorption of short-chain fatty acids by the colon. Gastroenterology 1980, 78, 1500–1507. [Google Scholar] [CrossRef]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. SCFA: Mechanisms and functional importance in the gut. Proc. Nutr. Soc. 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Tasdemir, S.S.; Sanlier, N. An insight into the anticancer effects of fermented foods: A review. J. Funct. Foods 2020, 75, 104281. [Google Scholar] [CrossRef]
- Moloney, J.N.; Cotter, T.G. ROS signalling in the biology of cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Gullo, M. Acetic acid bacteria. In Proceedings of the 4th International Conference on Acetic Acid Bacteria—Vinegar and Other Products (AAB 2015), Taiyuan, China, 15–19 September 2015. [Google Scholar] [CrossRef]
- Baba, N.; Higashi, Y.; Kanekura, T. Japanese black vinegar “Izumi” inhibits the proliferation of human squamous cell carcinoma cells via necroptosis. Nutr. Cancer 2013, 65, 1093–1097. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Morimura, S.; Shigematsu, T.; Maeda, H.; Kida, K. Antitumor activity of rice-shochu post-distillation slurry and vinegar produced from the post-distillation slurry via oral administration in a mouse model. Biofactors 2004, 22, 103–105. [Google Scholar] [CrossRef]
- Fukuyama, N.; Jujo, S.; Ito, I.; Shizuma, T.; Myojin, K.; Ishiwata, K.; Nagano, M.; Nakazawa, H.; Mori, H. Kurozu moromimatsu inhibits tumor growth of Lovo cells in a mouse model in vivo. Nutrition 2007, 23, 81–86. [Google Scholar] [CrossRef]
- Shizuma, T.; Ishiwata, K.; Nagano, M.; Mori, H.; Fukuyama, N. Protective effects of fermented rice vinegar sediment (Kurozu moromimatsu) in a diethylnitrosamine-induced hepatocellular carcinoma animal model. J. Clin. Biochem. Nutr. 2011, 49, 31–35. [Google Scholar] [CrossRef]
- Xibin, S.; Meilan, H.; Moller, H.; Evans, H.S.; Dixin, D.; Wenjie, D.; Jianbang, L. Risk factors for oesophageal cancer in Linzhou, China: A case-control study. Asian Pac. J. Cancer Prev. 2003, 4, 119–124. [Google Scholar]
- Huang, L.; Chen, L.; Gui, Z.X.; Liu, S.; Wei, Z.J.; Xu, A.M. Preventable lifestyle and eating habits associated with gastric adenocarcinoma: A case-control study. J. Cancer 2020, 11, 1231. [Google Scholar] [CrossRef]
- Radosavljević, V.; Janković, S.; Marinković, J.; Dokić, M. Non-occupational risk factors for bladder cancer a case-control study. Tumori J. 2004, 90, 175–180. [Google Scholar] [CrossRef]
- Yousefian, M.; Shakour, N.; Hosseinzadeh, H.; Hayes, A.W.; Hadizadeh, F.; Karimi, G. The natural phenolic compounds as modulators of NADPH oxidases in hypertension. Phytomedicine 2019, 55, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Na, L.; Chu, X.; Jiang, S.; Li, C.; Li, G.; He, Y.; Liu, Y.; Li, Y.; Sun, C. Vinegar decreases blood pressure by down-regulating AT1R expression via the AMPK/PGC-1α/PPARγ pathway in spontaneously hypertensive rats. Eur. J. Nutr. 2016, 55, 1245–1253. [Google Scholar] [CrossRef]
- Kondo, S.; Tayama, K.; Tsukamoto, Y.; Ikeda, K.; Yamori, Y. Antihypertensive effects of acetic acid and vinegar on spontaneously hypertensive rats. Biosci. Biotechnol. Biochem. 2001, 65, 2690–2694. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, Y.; Takata, Y.; Nagai, Y.; Mori, T.; Kawada, T.; Ishihara, N. Antihypertensive effects of Kurosu extract, a traditional vinegar produced from unpolished rice, in the SHR rats. J. Jpn. Soc. Food Sci. 2001, 48, 73–75. [Google Scholar] [CrossRef][Green Version]
- Saito, N.; Segawa, Y.; Maruyama, S.; Yamaoka, A.; Hashimoto, H.; Osera, T.; Kurihara, N. Abstract P196: Effect of combined oral intake of ginger extract and rice vinegar on blood pressure with 2-kidney, 1-clip renovascular hypertensive rats. Hypertension 2017, 70 (Suppl. 1), AP196. [Google Scholar]
- Maruyama, S.; Segawa, Y.; Saito, N.; Yamaoka, A.; Hashimoto, H.; Osera, T.; Kurihara, N. Abstract P198: Combination of Undaria pinnatifida sporophyll and vinegar remarkably decreases blood pressure in 2-kidney, 1-clip renovascular hypertensive rats. Hypertension 2017, 70 (Suppl. 1), AP198. [Google Scholar]
- Tanaka, H.; Watanabe, K.; Ma, M.; Hirayama, M.; Kobayashi, T.; Oyama, H.; Sakaguchi, Y.; Kanda, M.; Kodama, M.; Aizawa, Y. The effects of γ-aminobutyric acid, vinegar, and dried bonito on blood pressure in normotensive and mildly or moderately hypertensive volunteers. J. Clin. Biochem. Nutr. 2009, 45, 93–100. [Google Scholar] [CrossRef]
- Kajimoto, S.; Ooshima, Y.; Tayama, K. Effect of vinegar-added beverages on hypotensive action in healthy or mild hypertensive subjects (Japanese). J. Nutr. Food 2003, 6, 51–68. [Google Scholar]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-aminobutyric acid as a bioactive compound in foods: A review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Hayakawa, K.; Kimura, M.; Kamata, K. Mechanism underlying γ-aminobutyric acid-induced antihypertensive effect in spontaneously hypertensive rats. Eur. J. Pharmacol. 2002, 438, 107–113. [Google Scholar] [CrossRef]
- Matsubara, F.; Ueno, H.; Tadano, K.; Suyama, T.; Imaizumi, K.; Suzuki, T.; Magata, K.; Kikuchi, N.; Muneyuki, K.; Nakamichi, N.; et al. Effects of GABA supplementation on blood pressure and safety in adults with mild hypertension. Jpn. Pharmacol. Ther. 2002, 30, 963–972. [Google Scholar]
- Chen, C.; Chen, F. Study on the conditions to brew rice vinegar with high content of γ-amino butyric acid by response surface methodology. Food Bioprod. Process. 2009, 87, 334–340. [Google Scholar] [CrossRef]
- Xia, T.; Yao, J.; Wang, J.; Zhang, J.; Wang, M. Antioxidant activity and hepatoprotective activity of Shanxi aged vinegar in hydrogen peroxide-treated HepG-2 cells. In Advances in Applied Biotechnology; ICAB 2016, Lecture Notes in Electrical Engineering; Liu, H., Song, C., Ram, A., Eds.; Springer: Singapore, 2018; Volume 444, pp. 309–316. [Google Scholar] [CrossRef]
- Mayeux, R.; Stern, Y. Epidemiology of Alzheimer disease. Cold Spring Harb. Perspect. Med. 2012, 2, a006239. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.; Mazumder, P.M. Apple cider vinegar (ACV) and their pharmacological approach towards Alzheimer’s disease (AD): A review. Indian J. Pharm. Educ. Res. 2020, 54, s67–s74. [Google Scholar] [CrossRef]
- Tripathi, S.; Kumari, U.; Mitra Mazumder, P. Ameliorative effects of apple cider vinegar on neurological complications via regulation of oxidative stress markers. J. Food Biochem. 2020, 44, e13504. [Google Scholar] [CrossRef]
- Kanouchi, H.; Kakimoto, T.; Nakano, H.; Suzuki, M.; Nakai, Y.; Shiozaki, K.; Akikoka, K.; Otomaru, K.; Nagano, M.; Matsumoto, M. The brewed rice vinegar Kurozu increases HSPA1A expression and ameliorates cognitive dysfunction in aged P8 mice. PLoS ONE 2016, 11, e0150796. [Google Scholar] [CrossRef]
- Cao, L.; Song, X.; Song, Y.; Bi, J.; Cong, S.; Yu, C.; Tan, M. Fluorescent nanoparticles from mature vinegar: Their properties and interaction with dopamine. Food Funct. 2017, 8, 4744–4751. [Google Scholar] [CrossRef] [PubMed]
- Loke, C.; Lee, J.; Sander, S.; Mei, L.; Farella, M. Factors affecting intra-oral pH—A review. J. Oral Rehabil. 2016, 43, 778–785. [Google Scholar] [CrossRef]
- Willershausen, I.; Weyer, V.; Schulte, D.; Lampe, F.; Buhre, S.; Willershausen, B. In vitro study on dental erosion caused by different vinegar varieties using an electron microprobe. Clin. Lab. 2014, 60, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.W.; Li, D.Z.; Lu, J.Z.; Hu, W.; Chen, D.; Zhou, X.D. Effects of vinegar on tooth bleaching and dental hard tissues in vitro. Yi Xue Ban 2014, 45, 933–936. [Google Scholar]
- Anderson, S.; Gonzalez, L.A.; Jasbi, P.; Johnston, C.S. Evidence that daily vinegar ingestion may contribute to erosive tooth wear in adults. J. Med. Food 2020. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.H. Corrosive oesophageal injury following vinegar ingestion. Hong Kong Med. J. 2002, 8, 365–366. [Google Scholar]
- Chang, J.; Han, S.E.; Paik, S.S.; Kim, Y.J. Corrosive esophageal injury due to a commercial vinegar beverage in an adolescent. Clin. Endosc. 2020, 53, 366–369. [Google Scholar] [CrossRef]
- Lhotta, K.; Hofle, G.; Gasser, R.; Finkenstedt, G. Hypokalemia, hyperreninemia and osteoporosis in a patient ingesting large amounts of cider vinegar. Nephron 1998, 80, 242–243. [Google Scholar] [CrossRef]
- Varvarelis, N.; Khallafi, H.; Pappachen, B.; Krishnamurthy, M. Natural therapies—When ignorance is not bliss! J. Am. Geriatr. Soc. 2007, 55, 1892–1893. [Google Scholar] [CrossRef] [PubMed]
- Irkoren, S.; Sivrioglu, N. Unusual burn injury due to application of white vinegar and aspirin mixture. Int. Wound J. 2014, 11, 348–349. [Google Scholar] [CrossRef] [PubMed]
- Feldstein, S.; Afshar, M.; Krakowski, A.C. Chemical burn from vinegar following an internet-based protocol for self-removal of nevi. J. Clin. Aesthet. Dermatol. 2015, 8, 50. [Google Scholar] [PubMed]
- Benmeir, P.; Lusthaus, S.; Weinberg, A.; Neuman, A.; Eldad, A. Facial chemical burn. Burns 1994, 20, 282. [Google Scholar] [CrossRef]


{kind=link}
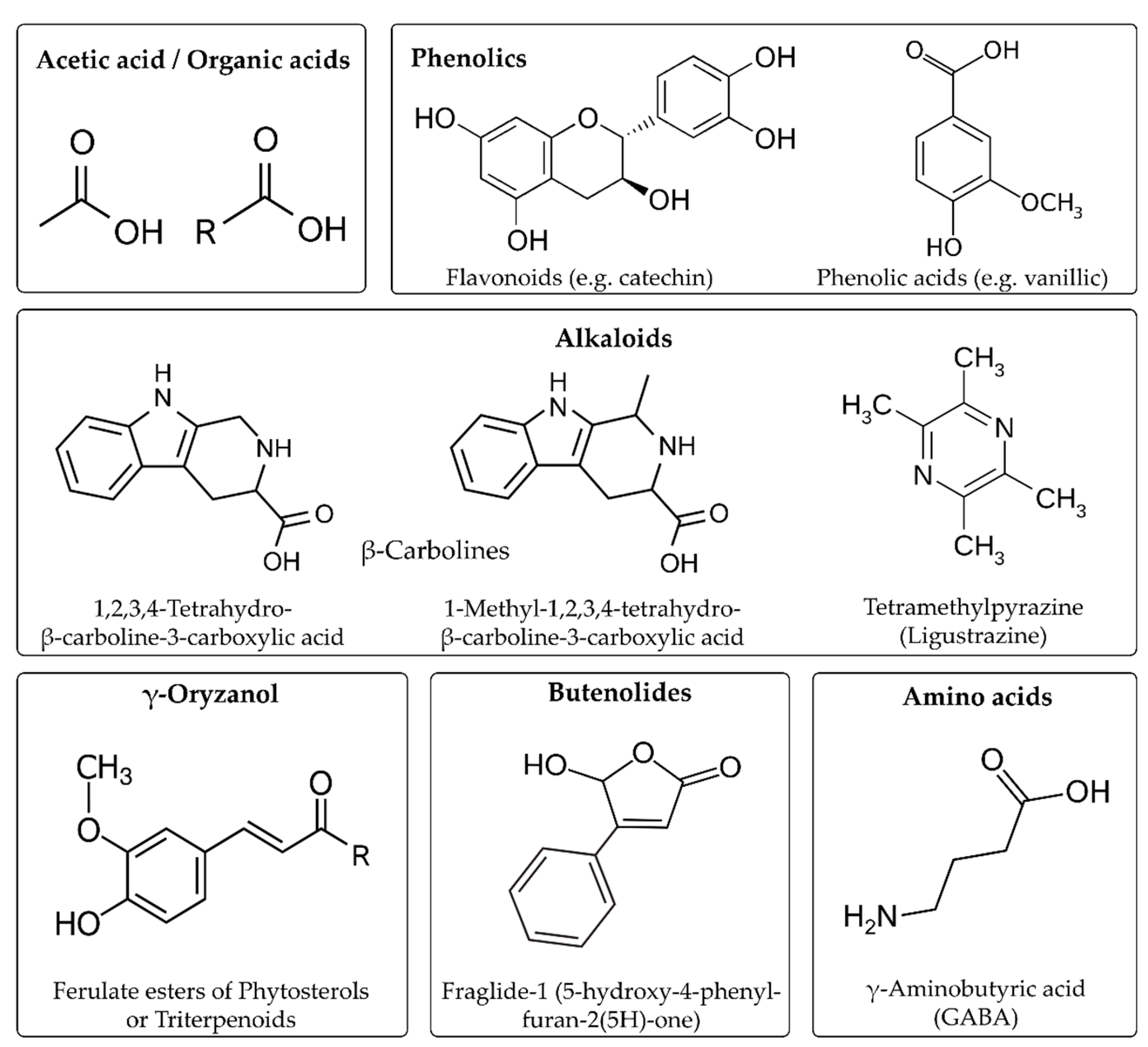{kind=link}
| Compounds | Reported Levels in Cereal Vinegars | Potential Bioactivity/Functionality | Types of Assayed Vinegars |
|---|---|---|---|
| Organic acids | |||
| Acetic acid (AA) | AA and LA prevail in cereal vinegars (>70–80%) [16,17]. Mean AA levels (g/L) in Chinese cereal vinegars 22.5–71.5 g/L (38.8–83.6% of total acids) [16], in SAVs (SBB) 12.6–33.0 [17], in ZAV (glutinous rice/bran) 17.7 [17], in 0–7-year-old traditional ZAVs 46.8 [18], in 0–7-year-old industrial ZAVs 39.1 [18], and in 10-year-old ZAV 66.1 [19]. | Acidity and flavor in vinegars. Antimicrobial properties. Regulation of lipid metabolism. Regulation of postprandial blood sugar levels. | RV, SAV cereal vinegars (SoV, OaV, TBV), ZAV vinegars (ZHV, LRV, traditional, industrial, aged and non-aged), WBrV, TDMV, FMV, SBrV [9,16,17,18,19,20,21,22,23,24]. |
| Lactic acid (LA) | Mean LA levels (g/L) in Chinese cereal vinegars 4.2–50.1 (11.2–40.4% of total acids) [16], in SAVs (SBB) 11.0–16.3 [17], in ZAV (glutinous rice/bran) 15.2 [17], in 0–7-year-old traditional ZAVs 24.1 [18] in 0–7-year-old industrial ZAVs 26.8 [18], in 10-year-old ZAV 32.8 [19], and in traditional Chinese rose vinegar (sticky rice) 1.33 [20]. | ||
| L-pyroglutamic acid (L-P) | Mean L-P content (g/L) in 0–7-year-old traditional ZAVs 16.4 [18], in 0–7-year-old industrial ZAVs 13.7 [18], and in 10-year-old ZAV 13.6 [19]. | ||
| Oxalic acid (OA) | Mean OA content (g/L) in 0–7-year-old traditional ZAVs 1.7 [18], in 0–7-year-old industrial ZAVs 1.4 [18], and in 10-year-old ZAV 1.9 [19]. | ||
| Tartaric acid (TA) | Mean TA content (g/L) in SAVs (SBB) 2.4–3.6 [17], in ZAV (glutinous rice/bran) 3.8 [17], in 0–7-year-old traditional ZAVs 0.9 [18], in 0–7-year-old industrial ZAVs 0.8 [18], and in 10-year-old ZAV 0.9 [19]. | ||
| Succinic acid (SA) | Mean SA content (g/L) in SAVs (SBB) 0.5–0.7 [17], in ZAV (glutinous rice/bran) 0.2 [17], in 0–7-year-old traditional ZAVs 9.2 [18], in 0–7-year-old industrial ZAVs 3.1 [18], and in 10-year-old ZAV 5.3 [19]. | ||
| Malic acid (MA) | Mean MA content (g/L) in SAVs (SBB) 0.2–0.8 [17], in ZAV (glutinous rice/bran) 1.0 [17], in 0–7-year-old traditional ZAVs 2.0 [18], in 0–7-year-old industrial ZAVs 1.8 [18], and in 10-year-old ZAV 2.2 [19]. | ||
| Citric acid (CA) | Mean CA content (g/L) in SAVs (SBB) 0.4–1.0, and in ZAV (glutinous rice/bran) 0.4 [17]. | ||
| Propanedioic acid (PA) | Mean PA content (g/L) in SAVs (SBB) 1.7–2.9, and in ZAV (glutinous rice/bran) 0.6 [17]. | ||
| Quinic acid (QA) | Mean QA content (g/L) in SAVs (SBB) 3.7–5.7, and in ZAV (glutinous rice/bran) 6.0 [17]. | ||
| Other acids at lower levels: butyric, caproic, formic, fumaric, glycolic, D-gluconic, hexanoic, 3-hydroxypropanoic and other hydroxy acids, α-ketoglutaric, 2-methylpropanoic, 3-methylbutanoic, octanoic, pentanoic, propanoic, higher MW fatty acids, etc. | Total mean organic acid content (g/L) 75–176 (including acetic acid) in SAV vinegars [16,23], 85.3 in ZAVs [16], 101.1 in 0–7-year-old traditional ZAVs [18], 49.5–77.3 in 0–10-year-old ZAVs [19], 122.6 in SBrVs [16], 87.3 in FMVs [16], and 55.6 in TDMVs [16]. Total acidity in 1–10 years aged SAVs was in the range 35–80 g/L (generally acidity increases with aging time) [24]. | ||
| Carbohydrates and sugar alcohols | |||
| Glucose (Glu), fructose (Fru), arabinose (Ara), xylose (Xyl) | Glu is the major sugar in SAVs (~38.7%), and Fru in other Chinese vinegars (>38.9%) (SBrVs, TDMVs, ZAVs), followed by Ara and Xyl [16]. Fru and Glu levels in Chinese cereal vinegars are found in the 1.6–13.9 and 2.7–8.7 g/L, respectively [16]. | Fermentable sugars serve as C-sources for the vinegar fermentation microflora. Several polysaccharides exert anticoagulation, antioxidant, immunoregulatory, and prebiotic properties. | RV (brown, sticky rice), SoV, SAV, ZAV, TBV, TDMV, FMV, SBrV [16,20,22,25,26]. |
| Other components at lower levels:allose, arabitol, deoxygalactose, dulcitol, erythritol, fucose, galactose, β-Glucans, glycerol, hemicellulosic arabinoxylans, glucose/mannose-rich polysaccharides, inositol, maltose, mannitol, mannose, microbial extracellular polysaccharides, pinitol, rhamnose, ribitol, ribose, sorbitol, threitol, trehalose, xylitol, etc. | Total sugar content (g/L) is in the range 2.6–28.7 in SAVs, and total mean content is 21.6 in SBrVs, 17.3 in TDMVs, and 17.2 in ZAVs [16]. Pentose levels (g/L) in ZAVs is in the range 1.2–3.8, and in SBrVs 2.2–2.6 [16]. Hexose levels (g/L) in ZAVs is in the range 6.8–23.4, and in SBrVs 11.2–21.0 [16]. | ||
| Minerals | |||
| Potassium (K) | Mean K content in KuV 3067 mg/kg [26], and in SoV (Taiwan) 365 ± 16 ppm [27]. | Essential nutrients. Blood pressure regulation. High bioavailability as vinegar components. Decreased absorption of dietary fat. | RV, KuV, SAV cereal vinegars (SoV, OaV, TBV), Taiwan SoV, ZAV vinegars (ZHV, LRV), KuV [23,27,28]. |
| Sodium (Na) | Mean Na content in SoV (Taiwan) 72.6 ± 0.2 ppm [27]. | ||
| Calcium (Ca) | Mean Ca content (mg/kg) in KuV 3.1 [28], in SAVs 651–1246 (the highest levels in OaV) [23], in ZAVs 31–134 [23], and in SoV (Taiwan) 5.7 ppm [27]. | ||
| Phosphorus (P) | Mean P content (mg/kg) in SAVs 3335–4723 mg/kg (the highest levels in OaV), and in ZAVs 22–1968 [23]. | ||
| Magnesium (Mg) | Mean Mg content in KuV 953.7 mg/kg [28], and in SoV (Taiwan) 12.6 ppm [27]. | ||
| Iron (Fe) | Mean Fe content (mg/kg) in KuV 24.7 [28], in SAVs 140–305 (the highest levels in OaV) [23], in ZAVs 1.5–38 [16], and in SoV (Taiwan) 1.8 ppm [27]. | ||
| Manganese (Mn) | Mean Mn content (mg/kg) in KuV 32.9 [28], in SAVs 40–70 (the highest levels in OaV) [23], in ZAVs 0.04–45 [23], and in SoV (Taiwan) 0.8 ppm [27]. | ||
| Zinc (Zn) | Mean Zn content (mg/kg) in SAVs 18–38 (the highest levels in OaV) [23], in ZAVs 0.2–17 [23], and in SoV (Taiwan) 0.3 ppm [27]. | ||
| Cooper (Cu) | Mean Cu content in SoV (Taiwan) 0.03 ppm [27]. | ||
| Selenium (Se) | Mean Se content (mg/kg) in KuV 0.002 [28], in SAVs 0.01–0.02 (the highest levels in OaV) [23], and in ZAVs 0.001–0.012 [23]. | ||
| Iodine (I) | Mean I content (mg/kg) in SAVs 0.1–0.53 (the highest levels in OaV), and in ZAVs 0.07–0.31 [23]. | ||
| Vitamins | |||
| Niacin, Nicotinamide (B3) | The B3 vitamers prevail in SAV and ZAV vinegars [23]. Total mean content (mg/100 g) in SAVs 29–38 niacin and 13–35 nicotinamide (the highest levels in OaV) [23]. Total mean content (mg/100 g) in ZAVs 2.3–12 niacin and 1.2–14 nicotinamide [23]. | Essential nutrients. Important for energy metabolism. Lowering cholesterol excretion (B3). Good skin health and vision (B1). Brain function (B1, B3). Blood vessel expansion (B3). | SAV cereal vinegars (SoV, OaV, TBV), ZAV vinegars (ZHV, LRV) [22,23]. |
| Riboflavin (B2) | Mean content (mg/100 g) of B2 in SAVs 0.9–1.6 (the highest levels in OaV) [6], and in ZAVs 0–2 [23]. | ||
| Thiamine (B1) | Mean content (μg/100 g) of B1 in SAVs 0.4–0.7 (the highest levels in OaV), and in ZAVs 0–0.13 [23]. | ||
| Amino acids | |||
| Glutamic acid (Glu) | Mean content (mg/100 g) of Glu in SAVs 463–888 [23], in ZAVs 295–659 [23], and in dried KuV 3306 [28]. | Regulation of cellular metabolism. Improvement of immunity. Promotion of brain development. Antioxidant activity. Regulation of glycemia levels. Antiobesity effects. Contribution to the product taste (umami, sweet, or bitter). | RV (glutinous, non-glutinous, black), SAV cereal vinegars (SoV, OaV, TBV), ZAV vinegars (ZHV, LRV, traditional, industrial, aged and non-aged), dried KuV, TDMV, FMV, SBrV [15,16,19,22,23,28] |
| Asparagine (Asn) | Mean content (mg/100 g) of Asn in SAVs 130–227, and in ZAVs 51–422 [23]. | ||
| Alanine (Ala) | Mean content (mg/100 g) of Ala in SAVs 119–251 [23], in ZAVs 153–170 [23], in 10-year-old ZAV 78 [19], and in dried KuV 979 [28]. | ||
| Valine (Val) | Mean content (mg/100 g) of Val in SAVs 107–216 [23], in ZAVs 106–118 [23], in 10-year-old ZAV 89 [19], and in dried KuV 654 [28]. | ||
| Proline (Pro) | Mean content (mg/100 g) of Pro in SAVs 130–294 [23], in ZAVs 96–180 [23], and in dried KuV 1185 [28]. | ||
| Glycine (Gly) | Mean content (mg/100 g) of Gly in SAVs 100–175 [23], in ZAVs 81–131 [23], in 10-year-old ZAV 144 [19], and in dried KuV 754 [28]. | ||
| Serine (Ser) | Mean content (mg/100 g) of Ser in SAVs 62–112 [23], in ZAVs 50–83 [23], and in dried KuV 530 [28]. | ||
| Isoleucine (Ile) | Mean content (mg/100 g) of Ile in SAVs 51–106 [23], in ZAVs 57–81 [23], in 10-year-old ZAV 67 [19], and in dried KuV 443 [28]. | ||
| Leucine (Leu) | Mean content (mg/100 g) of Leu in SAVs 93–175 [23], in ZAVs 96–98 [23], in 10-year-old ZAV 154 [19], and in dried KuV 734 [28]. | ||
| Methionine (Met) | Mean content (mg/100 g) of Met in SAVs 11–21 [23], in ZAVs 21–22 [23], in 10-year-old ZAV 17 [19], and in dried KuV 161 [28]. | ||
| Threonine (Thr) | Mean content (mg/100 g) of Thr in SAVs 50–92 [23], in ZAVs 51–137 [23], in 10-year-old ZAV 141 [19], and in dried KuV 451 [28]. | ||
| Histidine (His) | Mean content (mg/100 g) of His in SAVs 30–52 [23], in ZAVs 10–75 [23], in 10-year-old ZAV 86 [19], and in dried KuV 306 [28]. | ||
| Phenylalanine (Phe) | Mean content (mg/100 g) of Phe in SAVs 54–70 [23], in ZAVs 19–66 [23], in 10-year-old ZAV 50 [19], and in dried KuV 346 [28]. | ||
| Tryptophan (Trp) | Mean content of Trp in dried KuV 17 mg/100 g [28]. | ||
| Cysteine (Cys) | Mean content (mg/100 g) of Cys in SAVs 21–30 [23], in ZAVs 10–53 [23], and in dried KuV 226 [28]. | ||
| Arginine (Arg) | Mean content (mg/100 g) of Arg in SAVs 37–51 [23], in ZAVs 24–85 [23], in 10-year-old ZAV 16 [19], and in dried KuV 600 [28]. | ||
| Aspartic acid (Asp) | Mean content of Asp in dried KuV 828 mg/100 g [28]. | ||
| Tyrosine (Tyr) | Mean content (mg/100 g) of Tyr in SAVs 31–51 [23], in ZAVs 12–56 [23], in 10-year-old ZAV 29 [19], and in dried KuV 184 [28]. | ||
| Lysine (Lys) | Mean content (mg/100 g) of Lys in SAVs 46–101 [23], in ZAVs 48–72 [23], in 10-year-old ZAV 47 [19], and in dried KuV 223 [28]. | ||
| Other amino acids at lower levels: allo-isoleucine, ornithine, taurine, etc. | Total mean amino acid content in SAVs 15.3–28.8 g/L (the highest being that of aged SoV) [23]. Total mean amino acid content in ZAVs 12.2–24.7 g/L (the highest being that of ZHV) [23]. Total mean amino acid content (g/L) in TDMVs 13.1, in FMVs 6.2, in SBrVs 12.0, in ZAVs 8.8, and in SAVs 7.6 [16]. More than 1.8 g/100 g of bioactive branched amino acids (leucine, isoleucine, and valine) in freeze-dried black vinegar powder [15]. | ||
| γ-aminobutyric acid (GABA) | GABA was one of the main amino acids in SBrV, along with Ala and Leu [16]. GABA in vinegars fermented from germinated pigmented rice was in the range 1.14–1.73 mg/L (the highest content in germinated Hom Nil rice vinegar) [29]. | Inhibitory neurotransmitter in brain. Improvement of cerebral blood flow and cell metabolism. Anti-anxiety and tranquilizing effects. Diouretic. Lipid levels and blood pressure lowering. | ZAV, SAV, TDMV, FMV, SBrV [16,22,29] |
| Phenolic acids | |||
| Vanillic acid (VA) | Mean content (mg/L) of VA in SAVs (SBBs) 18.7–110.8 [17], in ZAV (glutinous rice/bran) 13.3 [17], in 6-year-old ZAV 148.4 [30,31], in 0–7-year-old traditional ZAVs 5.8 [18], in 0–7-year-old industrial ZAVs 7.6 [18], in aged XFV 1.5 [30], in 10-year-old ZAV 15.2 [19], and in 10-year-old ZAV extract 57.0 (prevailing phenolic compound in most cereal vinegars) [30]. | Antioxidant propeties—oxidative stress reduction. Antitumor and hepatoprotective effects. Antidiabetic (regulation of glucose metabolism, and amelioration of high glucose-induced insulin resistance). | RV (unpolished, black), SAVs, SoV, BrV, BaV, SBB, KuV, ZAVs (traditional, industrial, aged and non-aged), XFV, ZAV extract (5:15 in ethanol) [9,15,17,18,19,22,28,30,31,32,33,34,35,36] |
| p-hydroxybenzoic acid (p-HBA) | Mean content (mg/L) of p-HBA in 6-year-old ZAV 27.1 [31], in 0–7-year-old traditional ZAVs 7.5 [18], in 0–7-year-old industrial ZAVs 4.2 [18], in 10-year-old ZAV 24.7 [19], in 10-year-old ZAV extract 30.6 [30], in rice vinegar 5.0 [36], and in dried KuV 1.6 g/100 g [28]. | ||
| Gallic acid (GA) | Mean content of GA (mg/L) in SAVs (SBBs) 89.8–252.4 [17], in ZAV (glutinous rice/bran) 555.3 [17], in 10-year-old ZAV 1.0 [19], in 10-year ZAV extract 15.6 [30], in aged XFV 5.9 [35], in rice vinegar 3.4 [36], and in dried KuV 1.3 g/100 g [28]. | ||
| Chlorogenic acid (ChA) | Mean content of ChA (mg/L) in SAVs (SBBs) 12.7–20.8 [17], in ZAV (glutinous rice/bran) 7.4 [17], in 0–7-year-old traditional ZAVs 7.0 [18], in 0–7-year-old industrial ZAVs 6.6 [18], in 10-year-old ZAV 6.1 [19], in 10-year ZAV extract 12.0 [30], and in dried KuV 2.6 g/100 g [28]. | ||
| Caffeic acid (CA) | Mean content of CA (mg/L) in SAVs (SBBs) 17.5 [17], in 0–7-year-old traditional ZAVs 5.3 [18], in 0–7-year-old industrial ZAVs 5.3 [18], and in 10-year-old ZAV extract 6.6 [30]. | ||
| Syringic acid (SyA) | Mean content of SyA (mg/L) in 0–7-year-old traditional ZAVs 12.8 [18], in 0–7-year-old industrial ZAVs 3.1 [18], in 10-year-old ZAV 2.1 [19], in 10-year-old ZAV extract 13.5 [30], and in rice vinegar 1.5 [36]. | ||
| p-coumaric acid (p-CoA) | Mean content of p-CoA (mg/L) in SAVs (SBBs) 8.3–9.3 [17], in ZAV (glutinous rice/bran) 2.6 [17], in 0–7-year-old traditional ZAVs 0.7 [18], in aged XFV 0.1 [35], in 10-year-old ZAV 1.7 [19], in 10-year-old ZAV extract 4.4 [30], and in dried KuV 1.8 g/100 g [28]. | ||
| Trans-ferulic acid (FA) | Mean content of FA (mg/L) in SAVs (SBBs) 2.5–9.2 [17], in ZAV (glutinous rice/bran) 5.6 [17], in 0–7-year-old traditional ZAVs 6.7 [18], in 0–7-year-old industrial ZAVs 2.8 [18], in 10-year-old ZAV 1.6 [19], in aged XFV 0.2 [35], in 10-year-old ZAV extract 10.2 [30], and in dried KuV 1.5 g/100 g [28]. | ||
| Sinapic acid (SiA) | Mean content of SiA (mg/L) in 0–7-year-old traditional ZAVs 3.6 [18], in 0–7-year-old industrial ZAVs 2.8 [18], in 10-year-old ZAV 1.4 [19], in 10-year-old ZAV extract 4.8 [30], and in dried KuV 1.6 g/100 g [28]. | ||
| Protocatechuic acid (PA) | Mean content of PA 4.1 mg/L in rice vinegar [36]. | ||
| Salicylic acid (SaA) | Mean content of SaA 7.2 mg/L in SAVs (SBBs) [17]. | ||
| Other acids: o-coumaric, dihydroferulic, dihydrosinapic, gentisic, 3-hydroxyphenylacetic, isoferulic, 3-methylsalicylic, 3-phenylacetic, phloretic, phloroglucinic, 2,4,5-trihydroxybenzoic, etc. | Total phenolic acid content in 5-year-old SAV up to 980 ± 11 μg/g as gallic acid [32]. | ||
| Flavonoids and other phenolics | |||
| Catechin | Mean content (mg/L) of catechin (as (±)-catechin hydrate) in SAVs (SBB) 112.7–139.5 and in ZAV (glutinous rice/bran) 34.0 [17], in 6-year-old ZAV 57.5 [31], in 10-year-old ZAV 36.8 [19], in 0–7-year-old traditional ZAVs 41.4 [18], in 0–7-year-old industrial ZAVs 64.2 [18], in 10-year-old ZAV extract 133.2 [30], in rice vinegar 3.6 [36], and in dried KuV 2.3 g/100 g [28]. | Antioxidant properties—oxidative stress reduction. Antitumor properties. Hepatoprotective effects. Anti-inflammatory properties. Improvement of gut microbiota-related disorders. Inhibition of amylolytic enzyme activities and dietary carbohydrate absorption. Antioxidant activities in the food matrix (prevention of lipid oxidation, color, taste, and texture preservation). | SAV cereal vinegars (SoV, OaV, TBV, SBB, etc.), ZAV (ZHV, LRV, traditional, industrial, aged and non-aged), BRV, KuV [17,18,19,23,24,28,30,31,33,34,37,38]. |
| Rutin | Mean content (mg/L) of rutin in 10-year-old ZAV 1.8 [19], in 0–7-year-old traditional ZAVs 13.1 [18], and in 10-year-old ZAV extract 22.3 [30]. | ||
| L-epicatechin | Mean content of epicatechin 13.3 mg/L in ZAV (glutinous rice/bran) [17]. | ||
| (-)-epicatechin gallate | Mean content of epicatechin gallate 37.9 mg/L in SAVs (SBB) [17]. | ||
| Anthocyanins | Mean anthocyanin content in BRV 10.9 mg/L [37]. | ||
| Other phenolics: o-cresol, syringaldehyde, syringol, sinapaldehyde, vanillin, etc. | |||
| Total phenolic content (TPC), flavonoid content (TFC), and antioxidant capacity (AC) | TPC, TFC, and AC in cereal vinegars generally increase with brewing and aging time [24]. Mean TPC (mg GAE/mL) were found to be in SAVs 2.8 (OaV), 39.7 (SoV) and 45.2 (TBV) [23], in SAVs (SBB) 2.17 and in ZAV (glutinous rice/bran) 1.46 [17], in 1–10 years aged SAVs 1.0–5,8 [24], in 0–7 years aged traditional ZAV 4.3 [18], and in ZAVs 1.6 (ZHV) and 0.38 (LRB) [23]. Mean TFC (mg/L) in SAVs (SBB) was 21.0 [17], in 0–7 years aged traditional ZAV 320 [18], and in 1–10 years aged SAVs 330–4500 mg/kg [24]. | ||
| Non-enzymatic browning reaction products | |||
| Melanoidins | A mean browning index (A420 nm) of 0.26–0.83 for high MW melanoidins in 0–6-year-old ZAV [31]. | Antioxidant properties. Antiobesity properties. Hepatoprotective effects (melanoidins). Color, texture and flavor in vinegars. Possible negative health effects at high levels (mainly HMF and 2-F). | KuV, ZAV, SAV, FMV, TDMV [16,18,31,39,40,41] |
| 5-hydroxymethylfurfural (HMF) | HMF content (mg/L) in ZAVs 61.8–455.1, in SAVs 35.8–205.7, in FMVs 26.8–59.5, and mean content in TDMVs 92.4 [16]. | ||
| 2-furfural (2-F) | 2-F content (mg/L) in ZAVs 110.3–272.2, in SAVs 6.3–290.6, in FMVs 4.4–5.7, and in TDMVs (av.) 114.3 [16]. | ||
| Alkaloids | |||
| Saponin/total alkaloids | Mean levels in cereal SAVs (mg/mL): saponin 0.59 (in SoV), 0.66 (in OaV), 0.60 (in TBV), and total alkaloids 0.82 (in SoV), 1.21 (in OaV), 0.65 (in TBV) [23]. Mean levels in ZAVs (mg/mL): saponin 0.29 (in ZHV), 0.18 (in LRV), and total alkaloids 0.98 (in ZHV), 0.09 (in LRV) [23]. | Antioxidant properties. Anti-inflammatory properties. Antimicrobial and antimalarial properties. Hypouricemic effects. Anticarcinogenic properties. Neuromodulatory effects (inhibition of the antidepressant monoamine oxidase and benzodiazepine receptor). Negative health effects include neurotoxicity and genotoxicity. | RV, SAV cereal vinegars (SoV, OaV, TBV), ZAV (ZHV, LRV), Taiwan SoV [22,23,27,35,42,43,44,45,46]. |
| β-carbolines [1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid (THCA) and 1-methyl-1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid (MTCA)] | Mean β-carbolines content (μg/L) THCA 50, MTCA-SS 90, and MTCA-RS 1380 in SoV (Taiwan) [27]. | ||
| 2,3,5,6-tetramethylpyrazine (ligustrazine) | Above 30 mg/L in SAV according to national standards [42]. Mean content (mg/L) 6.4 in non-aged XFV, 3.3 in aged XFV [35], and 697 in 6-year-old ZAV [43]. | Inhibition of reactive species. Increase in antioxidant enzymes. Inhibition of platelet aggregation. Blood vessel vasodilation. Reduction in blood lipids. Hepatoprotective effects. Aroma contributor in vinegar (peanut, nutty). | TBV, SAV, ZAV, BrV, FV [22,35,42,43,44,45,46]. |
| Butenolides (4-C heterocyclic ring lactones) | |||
| Fraglide-1 (5-Hydroxy-4-phenyl-butenolide) | Content (μg/L) in brown rice vinegar (KuV) 19.99 ± 3.11, in 6 months aged Kozu 164–1336, in 8 years aged Kozu 239–1987, and in Chinese sticky rice husk 122.2 ± 0.7 [47]. | Anticancer properties. Antimicrobial and antiviral properties. Anti-inflammatory properties. Regulation of fatty acid storage and glucose metabolism (agonist for PPARγ). Antiobesity effects. | ZAV (Kozu), KuV, Chinese sticky rice husk [47]. |
| γ-Oryzanol | |||
| Mixture of triterpene alcohol and phytosterol ferulates | Not reported in cereal vinegars. Total content in Chinese rice varieties was 10.9–33.5 mg/100 g [48]. Cycloartenol ferulate, 24-methylenecycloartanyl, campesterol ferulate and β-sitosterol ferulate were the most abundant consituents [48]. Mean content (mg/100 g) of γ-oryzanol in brown rice from Wuchang was 21.5 ± 5.0, and 16.6 ± 6.3 in brown rice from Xiangshui [48]. | Antioxidant, anticarginogenic, anti-inflammatory, antihyperlipidemic, and neuroprotective properties. Inhibition of lipid peroxidation in food. | Rice [48]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kandylis, P.; Bekatorou, A.; Dimitrellou, D.; Plioni, I.; Giannopoulou, K. Health Promoting Properties of Cereal Vinegars. Foods 2021, 10, 344. https://doi.org/10.3390/foods10020344
Kandylis P, Bekatorou A, Dimitrellou D, Plioni I, Giannopoulou K. Health Promoting Properties of Cereal Vinegars. Foods. 2021; 10(2):344. https://doi.org/10.3390/foods10020344
Chicago/Turabian StyleKandylis, Panagiotis, Argyro Bekatorou, Dimitra Dimitrellou, Iris Plioni, and Kanella Giannopoulou. 2021. "Health Promoting Properties of Cereal Vinegars" Foods 10, no. 2: 344. https://doi.org/10.3390/foods10020344
APA StyleKandylis, P., Bekatorou, A., Dimitrellou, D., Plioni, I., & Giannopoulou, K. (2021). Health Promoting Properties of Cereal Vinegars. Foods, 10(2), 344. https://doi.org/10.3390/foods10020344