
Detection of Species Substitution in the Meat Value Chain by High-Resolution Melting Analysis of Mitochondrial PCR Products

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Meat Samples
2.2. Identification of Vertebrate Sources of Meat by PCR-HRM
2.3. Analysis of Various Physicochemical Treatments of Meat on PCR-HRM
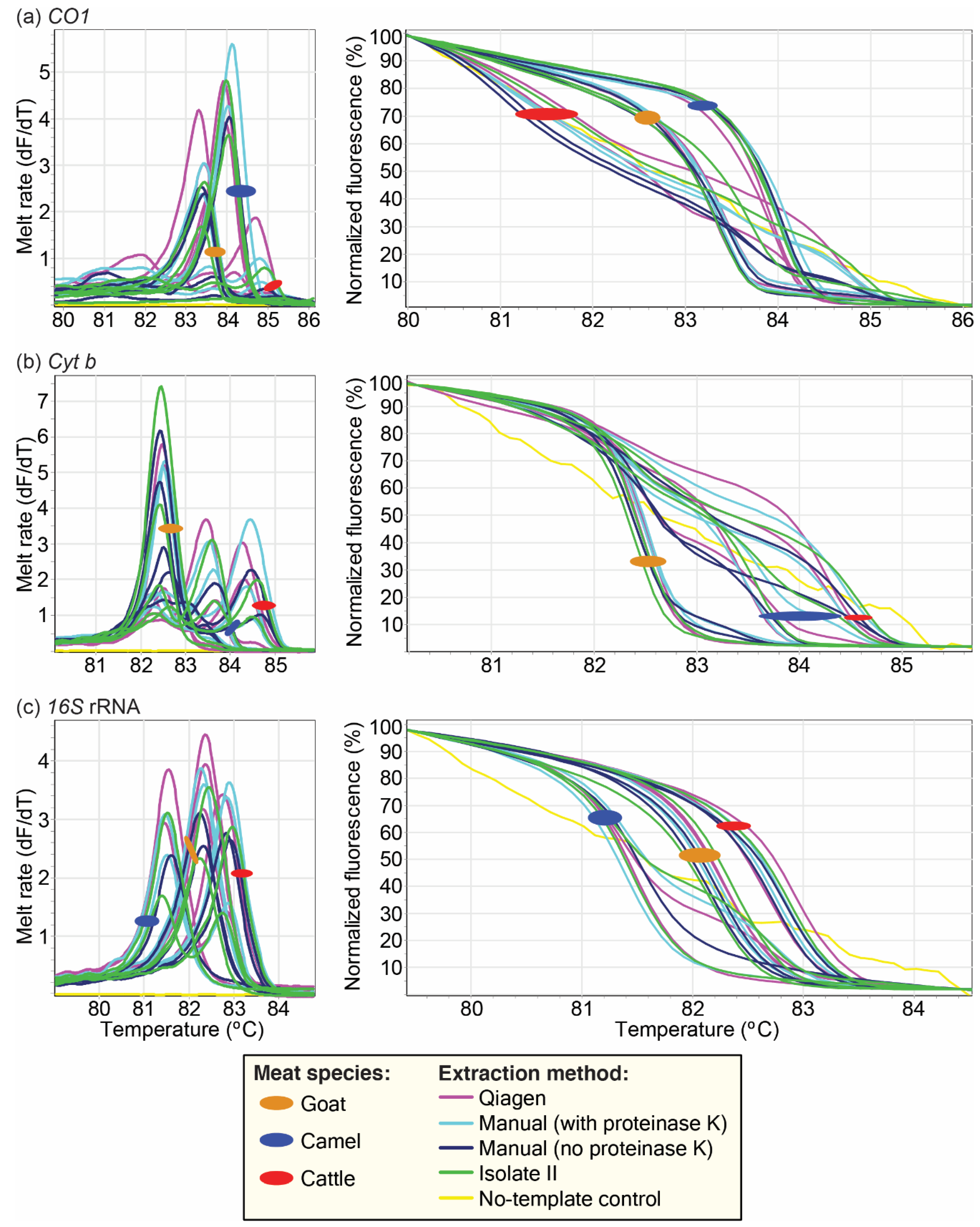
2.4. Analysis of Effect of Different Extraction Protocols
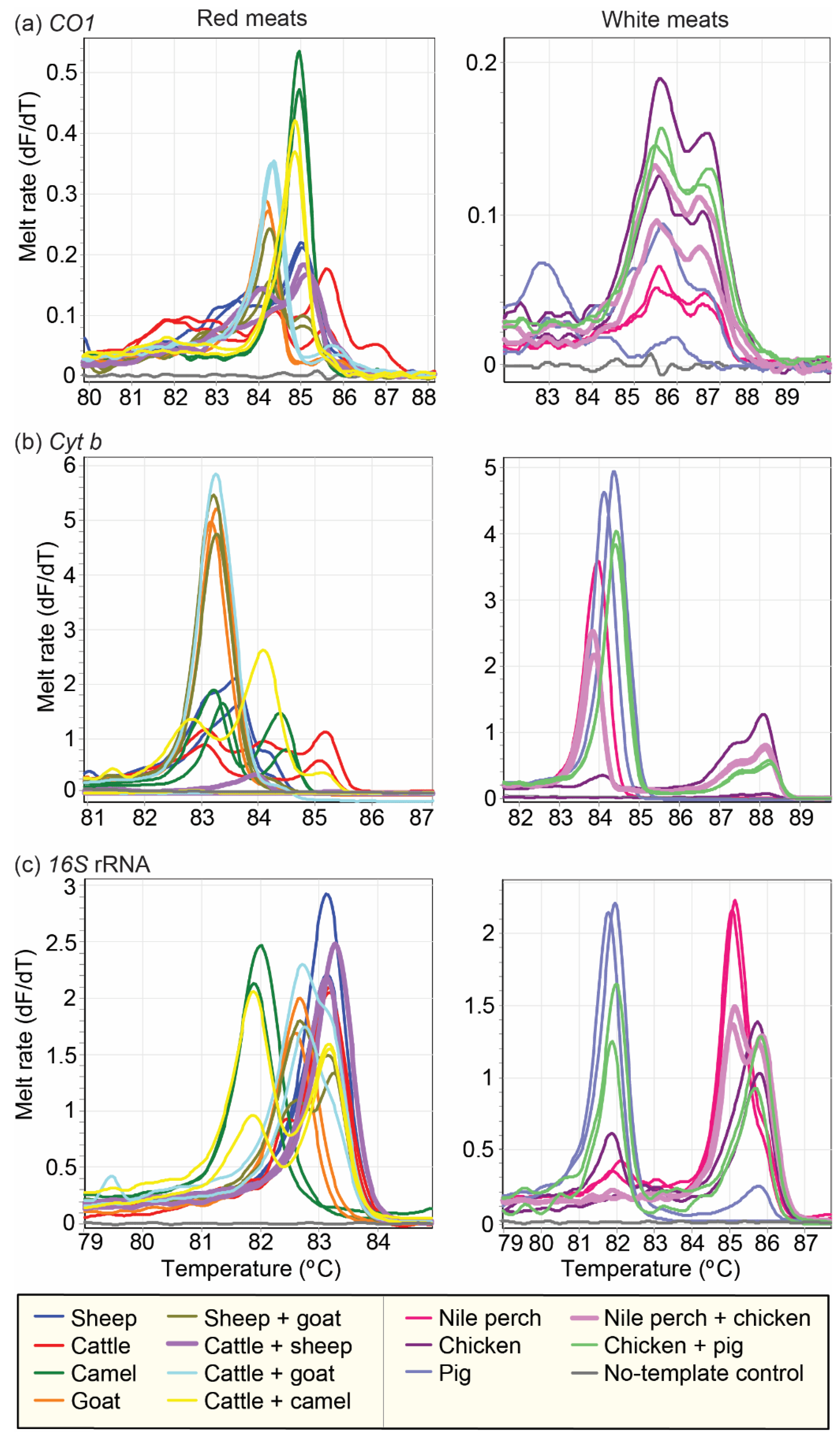
2.5. Analysis of Species Admixtures in Meat by PCR-HRM
2.6. DNA Sequencing for Species Confirmation and Statistical Analysis
3. Results
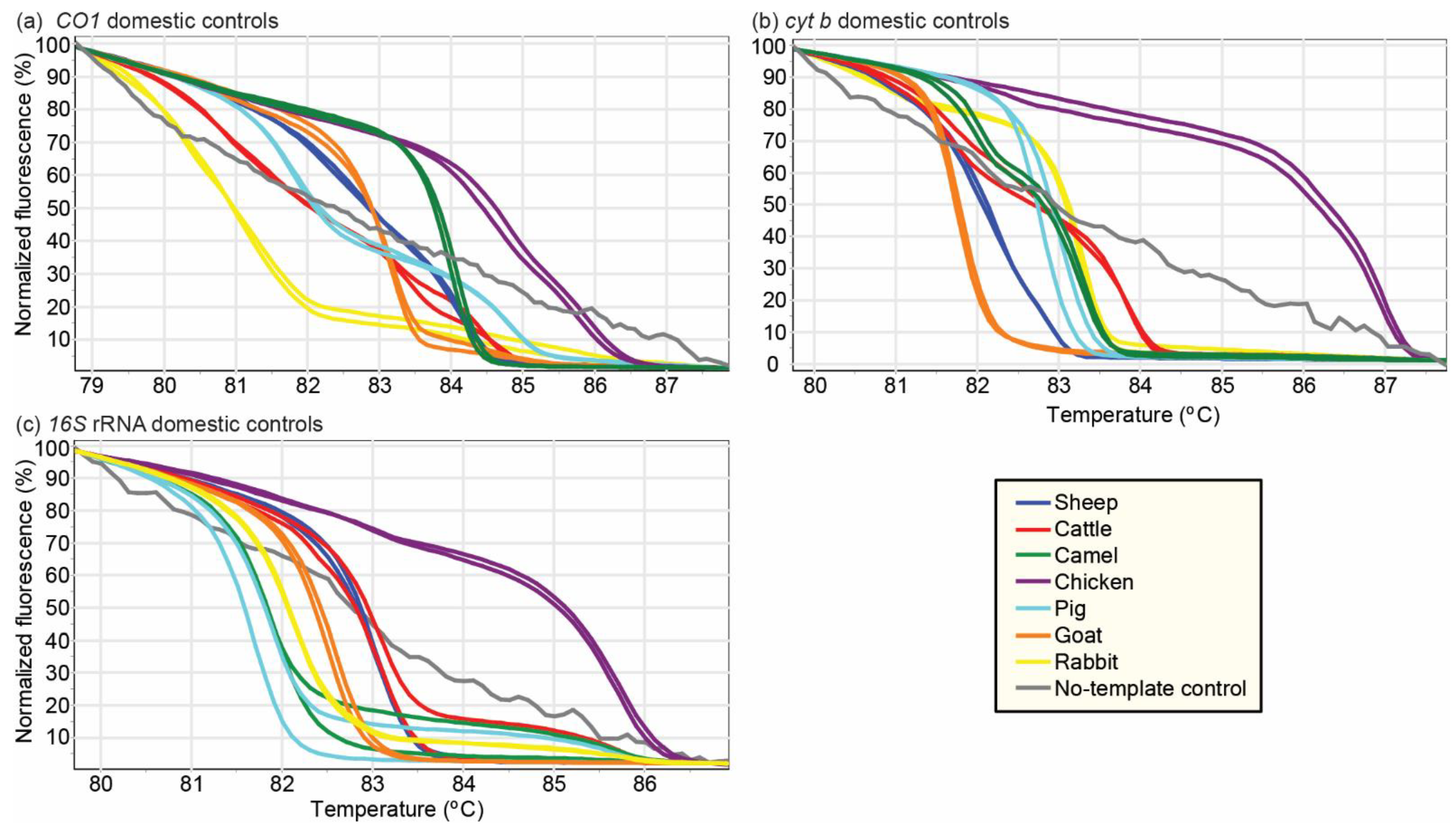
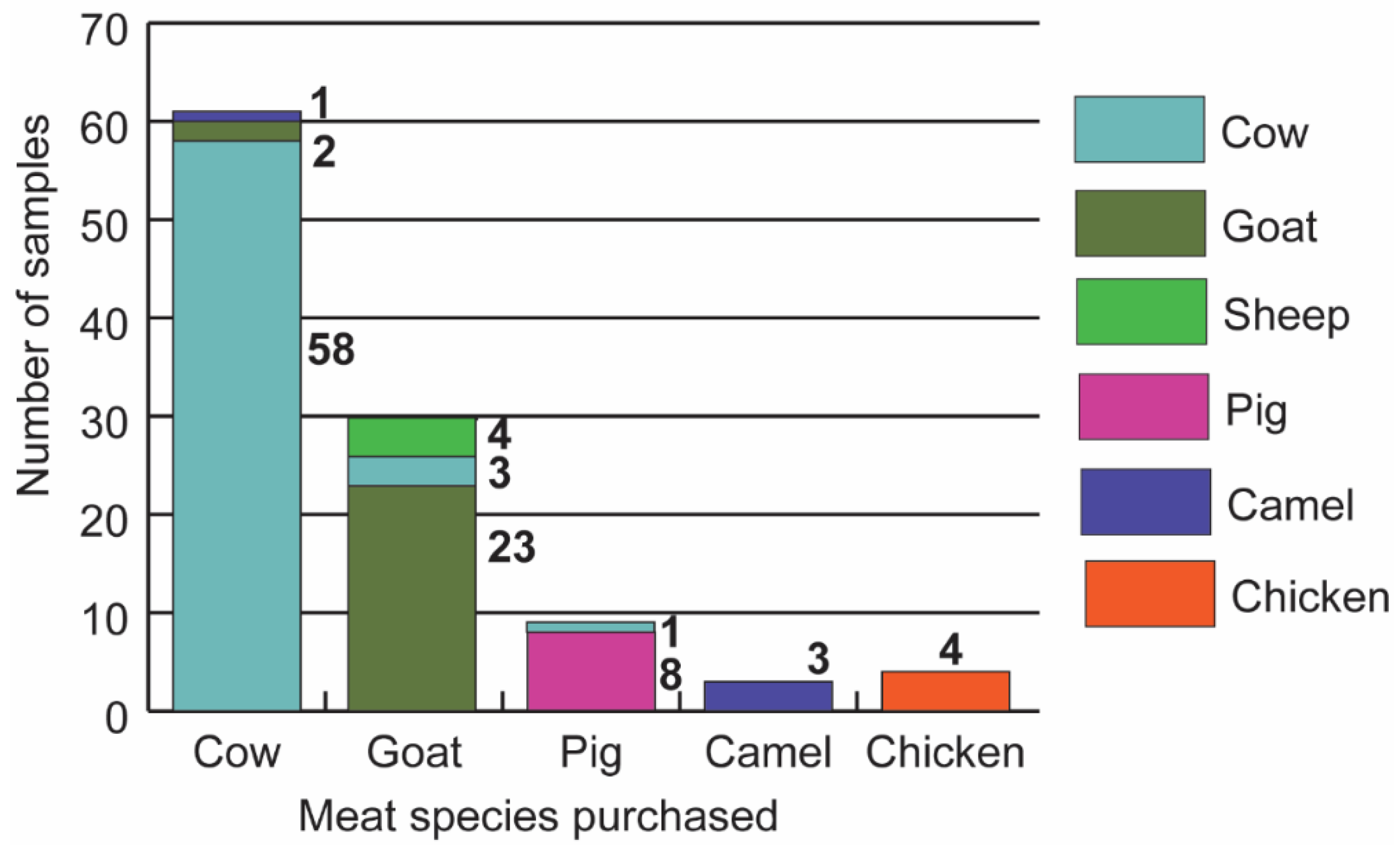
3.1. Vertebrate Sources of Meat Sold in Butcheries in Nairobi
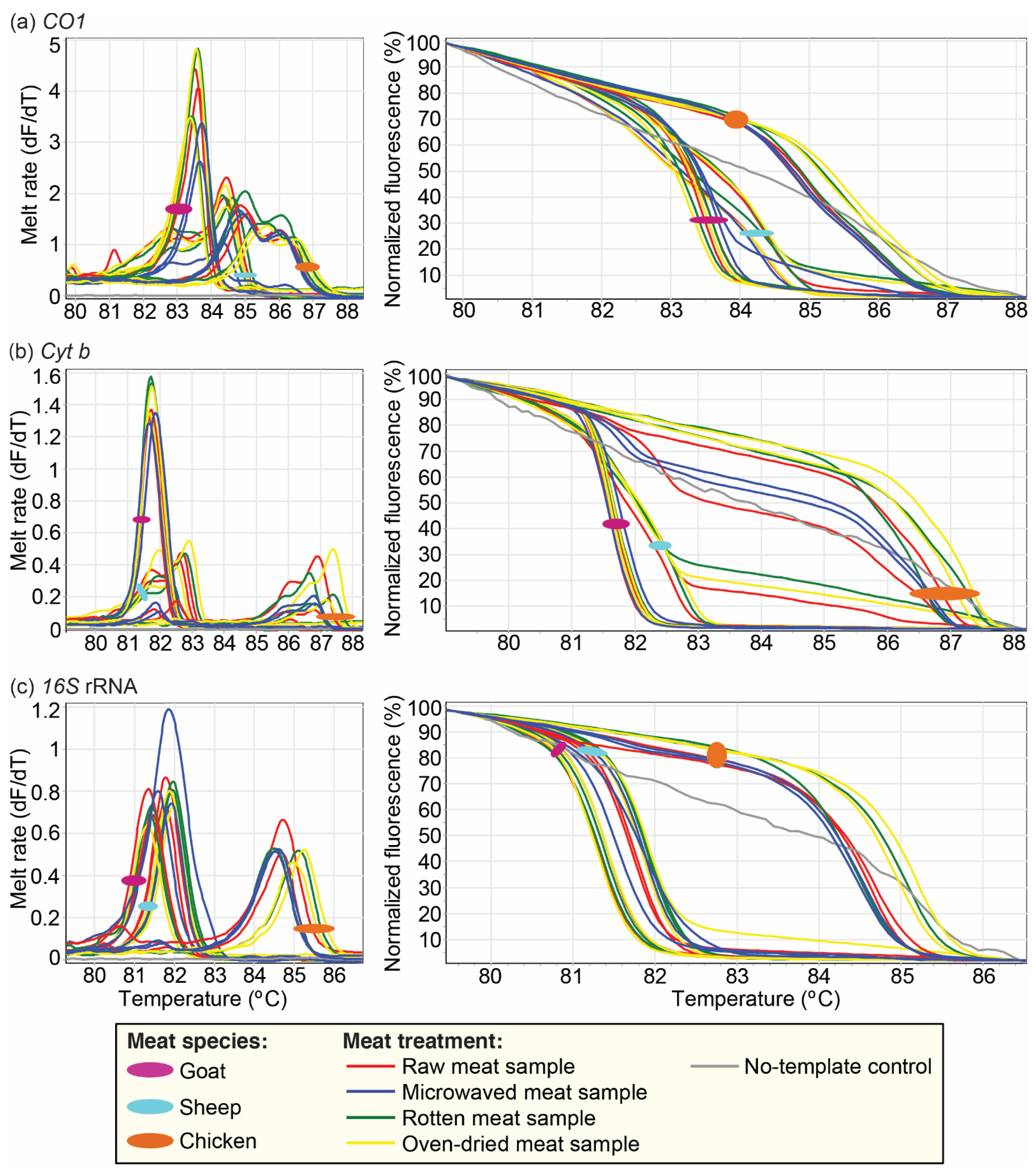
3.2. Effect of Physicochemical Condition of Meat Samples on Vertebrate Species Identification by PCR-HRM
3.3. Effect of Different DNA Extraction Protocols on PCR-HRM
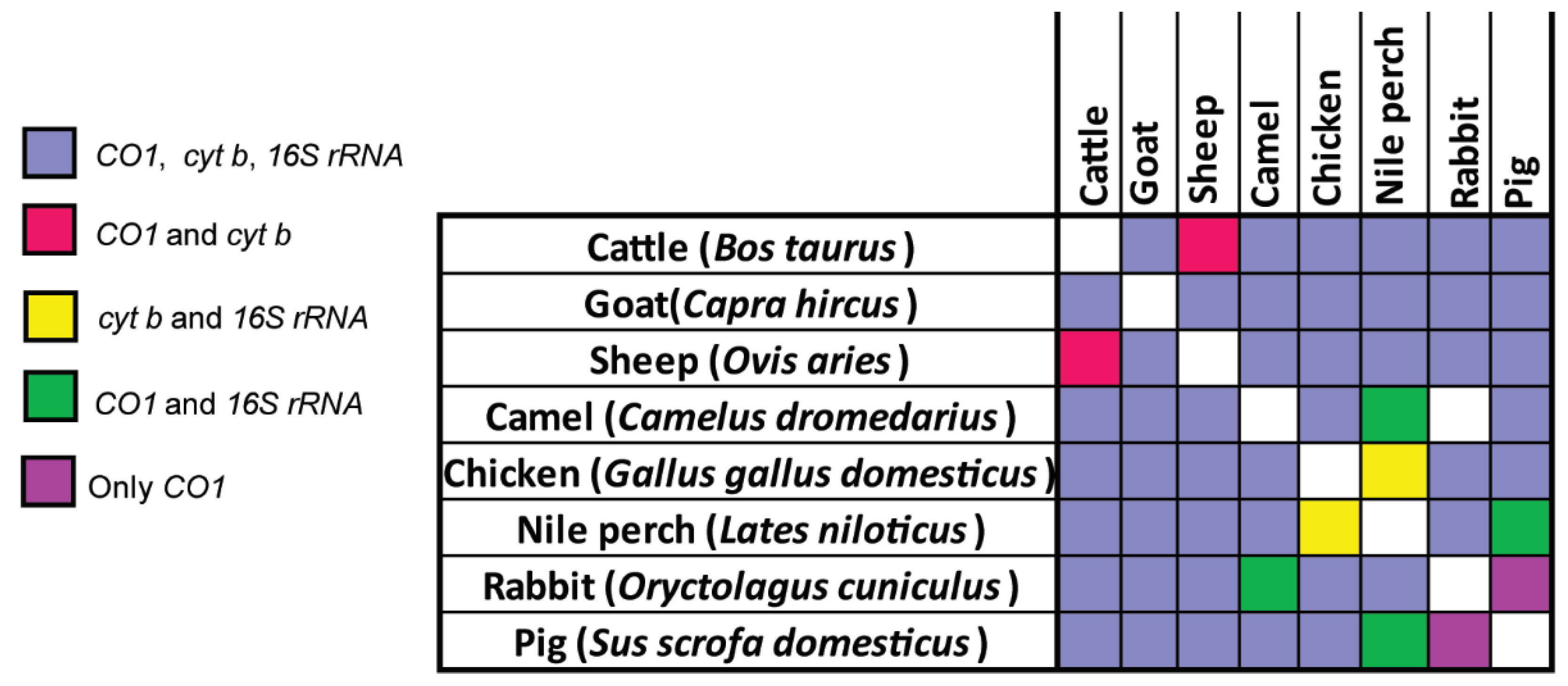
3.4. Distinction of Species in Mixed Meat Samples Using PCR-HRM
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spink, J.; Moyer, D.C. Defining the Public Health Threat of Food Fraud. J. Food Sci. 2011, 76, R157–R163. [Google Scholar] [CrossRef] [PubMed]
- Silvis, I.; van Ruth, S.; van der Fels-Klerx, H.; Luning, P. Assessment of food fraud vulnerability in the spices chain: An explorative study. Food Control. 2017, 81, 80–87. [Google Scholar] [CrossRef]
- Villa, C.; Costa, J.; Meira, L.; Oliveira, M.B.P.; Mafra, I. Exploiting DNA mini-barcodes as molecular markers to authenticate saffron (Crocus sativus L.). Food Control. 2016, 65, 21–31. [Google Scholar] [CrossRef]
- Handford, C.E.; Campbell, K.L.; Elliott, C.T. Impacts of milk fraud on food safety and nutrition with special emphasis on developing countries. Compr. Rev. Food Sci. Food Saf. 2016, 15, 130–142. [Google Scholar] [CrossRef]
- Sakaridis, I.; Ganopoulos, I.; Argiriou, A.; Tsaftaris, A. High resolution melting analysis for quantitative detection of bovine milk in pure water buffalo mozzarella and other buffalo dairy products. Int. Dairy J. 2013, 28, 32–35. [Google Scholar] [CrossRef]
- Yadav, S. Edible oil adulterations: Current issues, detection techniques, and health hazards. Int. J. Chem. Stud. 2018, 6, 1393–1397. [Google Scholar]
- Soares, S.; Grazina, L.; Costa, J.; Amaral, J.S.; Oliveira, M.B.; Mafra, I. Botanical authentication of lavender (Lavandula spp.) honey by a novel DNA-barcoding approach coupled to high resolution melting analysis. Food Control. 2018, 86, 367–373. [Google Scholar] [CrossRef]
- Cawthorn, D.-M.; Duncan, J.; Kastern, C.; Francis, J.; Hoffman, L. Fish species substitution and misnaming in South Africa: An economic, safety and sustainability conundrum revisited. Food Chem. 2015, 185, 165–181. [Google Scholar] [CrossRef]
- Fernandes, T.J.; Costa, J.; Oliveira, M.B.P.; Mafra, I. DNA barcoding coupled to HRM analysis as a new and simple tool for the authentication of Gadidae fish species. Food Chem. 2017, 230, 49–57. [Google Scholar] [CrossRef]
- Fernandes, T.J.; Silva, C.R.; Costa, J.; Oliveira, M.B.P.; Mafra, I. High resolution melting analysis of a COI mini-barcode as a new approach for Penaeidae shrimp species discrimination. Food Control. 2017, 82, 8–17. [Google Scholar] [CrossRef]
- Nasreen, S.; Ahmed, T. Food adulteration and consumer awareness in Dhaka City, 1995-2011. J. Health Popul. Nutr. 2014, 32, 452–464. [Google Scholar] [PubMed]
- Martín-Fernández, B.; Costa, J.; De-Los-Santos-Álvarez, N.; López-Ruiz, B.; Oliveira, M.B.P.; Mafra, I. High resolution melting analysis as a new approach to discriminate gluten-containing cereals. Food Chem. 2016, 211, 383–391. [Google Scholar] [CrossRef]
- Woolfe, M.; Primrose, S. Food forensics: Using DNA technology to combat misdescription and fraud. Trends Biotechnol. 2004, 22, 222–226. [Google Scholar] [CrossRef]
- Chuah, L.-O.; Bin He, X.; Effarizah, M.E.; Syahariza, Z.A.; Shamila-Syuhada, A.K.; Rusul, G. Mislabelling of beef and poultry products sold in Malaysia. Food Control. 2016, 62, 157–164. [Google Scholar] [CrossRef]
- Cawthorn, D.-M.; Steinman, H.A.; Hoffman, L. A high incidence of species substitution and mislabelling detected in meat products sold in South Africa. Food Control. 2013, 32, 440–449. [Google Scholar] [CrossRef]
- Farag, M.R. Identification of different animal species in meat and meat products: Trends and advances. Adv. Anim. Veter- Sci. 2015, 3, 334–346. [Google Scholar] [CrossRef]
- Di Pinto, A.; Bottaro, M.; Bonerba, E.; Bozzo, G.; Ceci, E.; Marchetti, P.; Mottola, A.; Tantillo, G.M. Occurrence of mislabeling in meat products using DNA-based assay. J. Food Sci. Technol. 2015, 52, 2479–2484. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Zhang, C. Detection of adulterated murine components in meat products by TaqMan© real-time PCR. Food Chem. 2016, 192, 485–490. [Google Scholar] [CrossRef]
- Ouso, D.O.; Otiende, M.Y.; Jeneby, M.M.; Oundo, J.W.; Bargul, J.L.; Miller, S.E.; Wambua, L.; Villinger, J. Three-gene PCR and high-resolution melting analysis for differentiating vertebrate species mitochondrial DNA for biodiversity research and complementing forensic surveillance. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Kenya Markets Trust. A Study on Meat End-Market Trends in Kenya. 2019. Available online: https://www.kenyamarkets.org/wp-content/uploads/2019/11/Meat-End-Market-Trends-in-Kenya.pdf (accessed on 14 August 2020).
- Ballin, N.Z.; Vogensen, F.; Karlsson, A.H. Species determination—Can we detect and quantify meat adulteration? Meat Sci. 2009, 83, 165–174. [Google Scholar] [CrossRef]
- Al-Kahtani, H.A.; Ismail, E.; Ahmed, M.A. Pork detection in binary meat mixtures and some commercial food products using conventional and real-time PCR techniques. Food Chem. 2017, 219, 54–60. [Google Scholar] [CrossRef]
- Alarcon, P.; Fèvre, E.M.; Muinde, P.; Murungi, M.K.; Kiambi, S.; Akoko, J.; Rushton, J. Urban livestock keeping in the city of Nairobi: Diversity of production systems, supply chains, and their disease management and risks. Front. Veter-Sci. 2017, 4, 171. [Google Scholar] [CrossRef]
- Abbas, O.; Zadravec, M.; Baeten, V.; Mikuš, T.; Lešić, T.; Vulić, A.; Prpić, J.; Jemeršić, L.; Pleadin, J. Analytical methods used for the authentication of food of animal origin. Food Chem. 2018, 246, 6–17. [Google Scholar] [CrossRef]
- Bourguiba-Hachemi, S.; Fathallah, M.D. DNA Testing of meat foods raises issues beyond adulteration. Sky J. Food Sci. 2016, 8, 1–7. [Google Scholar]
- Mbugua, D.; Kimwele, C.N.; Jung’a, J.O.; Gatebe, E.; Gakuya, F.; Kinuthia, M.J.; Kwamboka, E.S. Use of cytochrome oxidase 1 gene region: A molecular tool for the domestic and wildlife industry in Kenya. Sci. J. Anim. Sci. 2014, 3, 87–94. [Google Scholar] [CrossRef]
- Reed, G.H.; Kent, J.O.; Wittwer, C.T. High-resolution DNA melting analysis for simple and efficient molecular diagnostics. Pharmacogenomics 2007, 8, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Druml, B.; Cichna-Markl, M. High resolution melting (HRM) analysis of DNA–Its role and potential in food analysis. Food Chem. 2014, 158, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Sakaridis, I.; Ganopoulos, I.; Argiriou, A.; Tsaftaris, A. A fast and accurate method for controlling the correct labeling of products containing buffalo meat using High Resolution Melting (HRM) analysis. Meat Sci. 2013, 94, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Ogola, E.; Villinger, J.; Mabuka, D.; Omondi, D.; Orindi, B.; Mutunga, J.; Owino, V.; Masiga, D.K. Composition of Anopheles mosquitoes, their blood-meal hosts, and Plasmodium falciparum infection rates in three islands with disparate bed net coverage in Lake Victoria, Kenya. Malar. J. 2017, 16, 1–12. [Google Scholar] [CrossRef]
- Lee, P.-S.; Sing, K.-W.; Wilson, J.-J. Reading mammal diversity from flies: The persistence period of amplifiable mammal mtdna in blowfly guts (Chrysomya megacephala) and a new DNA Mini-Barcode target. PLoS ONE 2015, 10, e0123871. [Google Scholar] [CrossRef]
- Meusnier, I.; Singer, G.A.; Landry, J.-F.; Hickey, D.A.; Hebert, P.D.; Hajibabaei, M. A universal DNA mini-barcode for biodiversity analysis. BMC Genom. 2008, 9, 214. [Google Scholar] [CrossRef] [PubMed]
- Omondi, D.; Masiga, D.K.; Ajamma, Y.U.; Fielding, B.C.; Njoroge, L.; Villinger, J. Unraveling Host-Vector-Arbovirus Interactions by Two-Gene High Resolution Melting Mosquito Bloodmeal Analysis in a Kenyan Wildlife-Livestock Interface. PLoS ONE 2015, 10, e0134375. [Google Scholar] [CrossRef] [PubMed]
- Boakye, D.A.; Tang, J.; Truc, P.; Merriweather, A.; Unnasch, T.R. Identification of bloodmeals in haematophagous Diptera by cytochrome B heteroduplex analysis. Med. Veter-Èntomol. 1999, 13, 282–287. [Google Scholar] [CrossRef]
- Peña-García, V.H.; Fernandez, G.J.; Gómez-Palacio, A.; Mejía-Jaramillo, A.; Cantillo, O.; Triana-Chávez, O. High-Resolution Melting (HRM) of the Cytochrome B Gene: A Powerful Approach to Identify Blood-Meal Sources in Chagas Disease Vectors. PLOS Neglected Trop. Dis. 2012, 6, e1530. [Google Scholar] [CrossRef]
- Kipanga, P.N.; Omondi, D.; Mireji, P.O.; Sawa, P.; Masiga, D.K.; Villinger, J. High-resolution melting analysis reveals low Plasmodium parasitaemia infections among microscopically negative febrile patients in western Kenya. Malar. J. 2014, 13, 429. [Google Scholar] [CrossRef][Green Version]
- Ivanova, N.V.; Clare, E.L.; Borisenko, A.V. DNA barcoding in mammals. In Methods in Molecular Biology; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2012; Volume 858, pp. 153–182. [Google Scholar]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Lee, H.C.; Lai, K.; Lorenc, M.T.; Imelfort, M.; Duran, C.; Edwards, D. Bioinformatics tools and databases for analysis of next-generation sequence data. Briefings Funct. Genom. 2011, 11, 12–24. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System (http://www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef]
- Alarcon, P.; Fevre, E.; Murungi, M.K.; Muinde, P.; Akoko, J.; Dominguez-Salas, P.; Kiambi, S.; Ahmed, S.; Haesler, B.; Rushton, J. Mapping of beef, sheep and goat food systems in Nairobi—A framework for policy making and the identification of structural vulnerabilities and deficiencies. Agric. Syst. 2017, 152, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Gorski, I.; Chung, W.-C.; Herr, K.; Mehta, K. Nyama Choma culture: Implications of increased red meat and alcohol consumption in East Africa. J. Sustain. Dev. 2016, 9, 96. [Google Scholar] [CrossRef]
- Gray, C.; van Zyl, A.; Strauss, L. ‘Midnight Anaphylaxis’ to Red Meat in Patients with Alpha-Gal Sensitisation: A Recent Discovery in the Food Allergy World and a Case Report from South Africa. Curr. Allergy Clin. Immunol. 2016, 29, 102–104. [Google Scholar]
- Ali, E.; Razzak, A.; Hamid, S.B.A.; Rahman, M.; Amin, A.; Rashid, N.R. Asing Multiplex PCR assay for the detection of five meat species forbidden in Islamic foods. Food Chem. 2015, 177, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Izadpanah, M.; Mohebali, N.; Gorji, Z.E.; Farzaneh, P.; Vakhshiteh, F.; Fazeli, S.A.S. Simple and fast multiplex PCR method for detection of species origin in meat products. J. Food Sci. Technol. 2018, 55, 698–703. [Google Scholar] [CrossRef] [PubMed]
- Kitpipit, T.; Sittichan, K.; Thanakiatkrai, P. Direct-multiplex PCR assay for meat species identification in food products. Food Chem. 2014, 163, 77–82. [Google Scholar] [CrossRef] [PubMed]
- O’Mahony, P.J. Finding horse meat in beef products—A global problem. Qjm Int. J. Med. 2013, 106, 595–597. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, X.; Zhang, Q.; Liu, Z.; Zhou, X.; Liu, B. A multiplex PCR method for detection of five animal species in processed meat products using novel species-specific nuclear DNA sequences. Eur. Food Res. Technol. 2020, 246, 1351–1360. [Google Scholar] [CrossRef]
- López-Oceja, A.; Nuñez, C.; Baeta, M.; Gamarra, D.; de Pancorbo, M. Species identification in meat products: A new screening method based on high resolution melting analysis of cyt b gene. Food Chem. 2017, 237, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Ramanan, S.S.; Kerala Agricultural University; Khapugin, A.A.; Reserve, M.S.N. Mordovia State University Book Review: Problematic Wildlife: A Cross-Disciplinary Approach. Nat. Conserv. Res. 2017, 2. [Google Scholar] [CrossRef]
- Arslan, A.; Ilhak, O.I.; Calicioglu, M. Effect of method of cooking on identification of heat processed beef using polymerase chain reaction (PCR) technique. Meat Sci. 2006, 72, 326–330. [Google Scholar] [CrossRef]
- Tan, Z.-J.; Chen, S.-J. Nucleic acid helix stability: Effects of salt concentration, cation valence and size, and chain length. Biophys. J. 2006, 90, 1175–1190. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; He, Y.; Lv, R.; Chen, H.; Wang, Q.; Pan, L. Detection and quantification of beef and pork materials in meat products by duplex droplet digital PCR. PLoS ONE 2017, 12, e0181949. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Name | Primer Sequence (5′-3′) | Amplicon Size (bp) | Citation |
|---|---|---|---|---|
| CO1 | Uni-Minibar-F1 | TCCACTAATCACAARGATATTGGTAC | 205 | [19,31,32] |
| Ronping_R | TATCAGGGGCTCCGATTAT | |||
| 16S rRNA | Vert16S For | GAGAAGACCCTRTGGARCTT | 200 | [33] |
| Vert16S Rev | CGCTGTTATCCCTAGGGTA | |||
| cyt b | Cyt b For | CCCCTCAGAATGATATTTGTCCTCA | 383 | [34,35] |
| Cyt b Rev | CATCCAACATCTCAGCATGATGAAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Njaramba, J.K.; Wambua, L.; Mukiama, T.; Amugune, N.O.; Villinger, J. Detection of Species Substitution in the Meat Value Chain by High-Resolution Melting Analysis of Mitochondrial PCR Products. Foods 2021, 10, 3090. https://doi.org/10.3390/foods10123090
Njaramba JK, Wambua L, Mukiama T, Amugune NO, Villinger J. Detection of Species Substitution in the Meat Value Chain by High-Resolution Melting Analysis of Mitochondrial PCR Products. Foods. 2021; 10(12):3090. https://doi.org/10.3390/foods10123090
Chicago/Turabian StyleNjaramba, Jane Kagure, Lillian Wambua, Titus Mukiama, Nelson Onzere Amugune, and Jandouwe Villinger. 2021. "Detection of Species Substitution in the Meat Value Chain by High-Resolution Melting Analysis of Mitochondrial PCR Products" Foods 10, no. 12: 3090. https://doi.org/10.3390/foods10123090
APA StyleNjaramba, J. K., Wambua, L., Mukiama, T., Amugune, N. O., & Villinger, J. (2021). Detection of Species Substitution in the Meat Value Chain by High-Resolution Melting Analysis of Mitochondrial PCR Products. Foods, 10(12), 3090. https://doi.org/10.3390/foods10123090