Anti-Cancer Properties of Coix Seed Oil against HT-29 Colon Cells through Regulation of the PI3K/AKT Signaling Pathway


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Characterization of Coix Seed Oil
2.2.1. Fatty Acid Determination
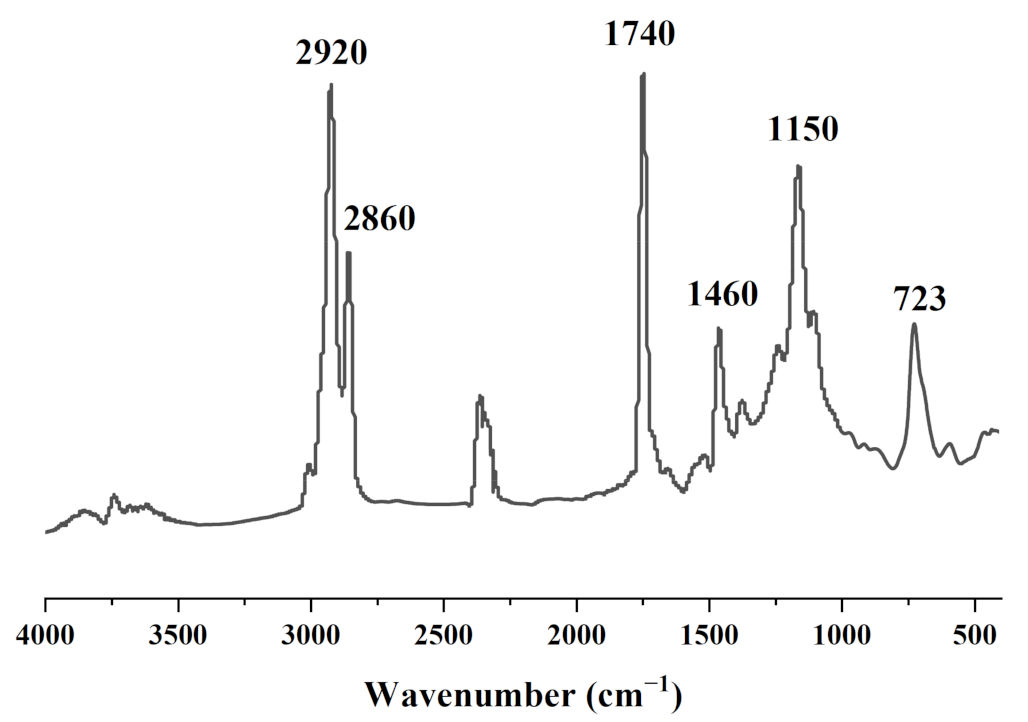
2.2.2. Fourier Transform Infrared (FT-IR) Analysis
2.3. Cell Culture
2.4. Cell Viability
2.5. Cell Morphological Assessment
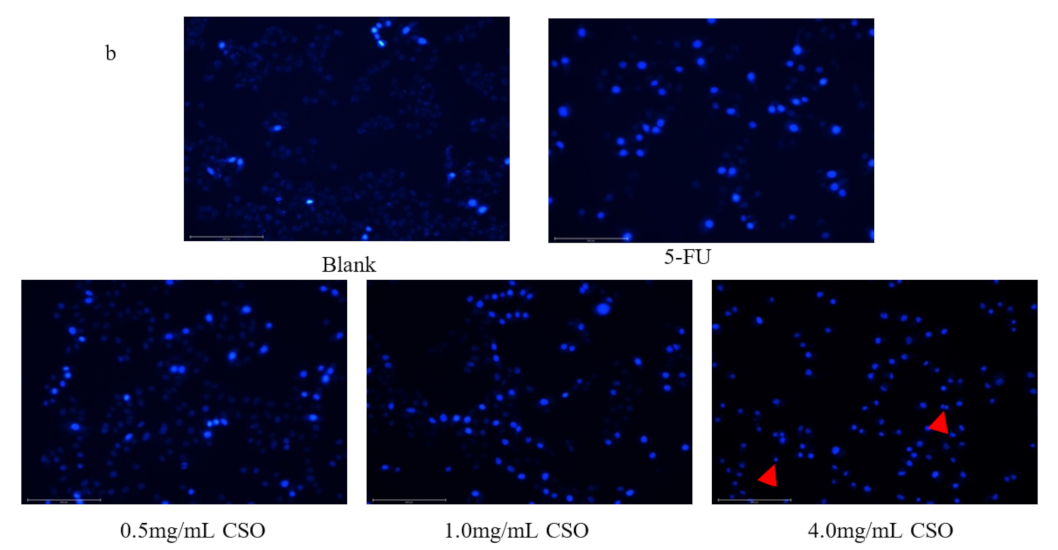
2.6. Hoechst 33258 Staining
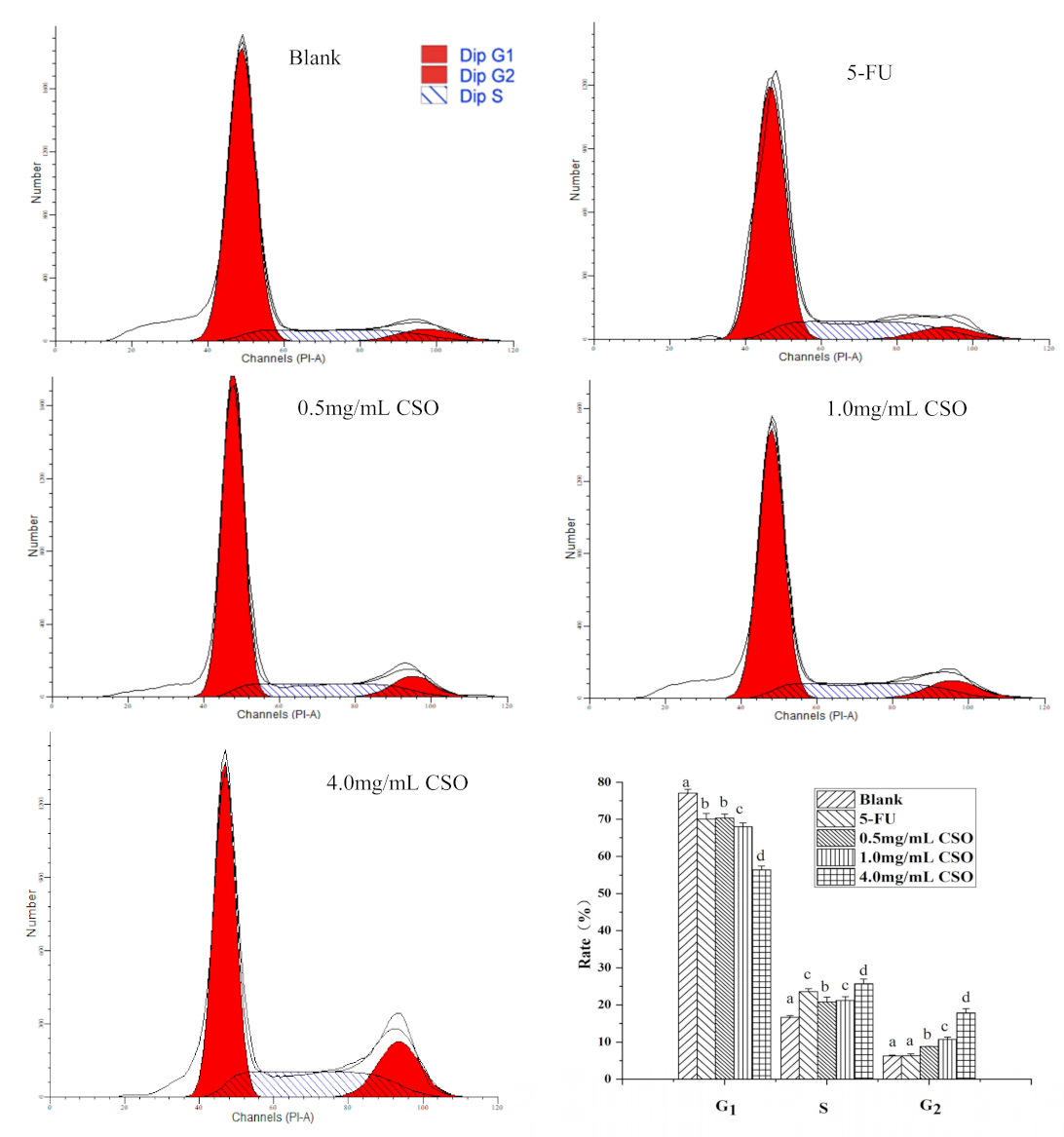
2.7. Cell Cycle Analysis
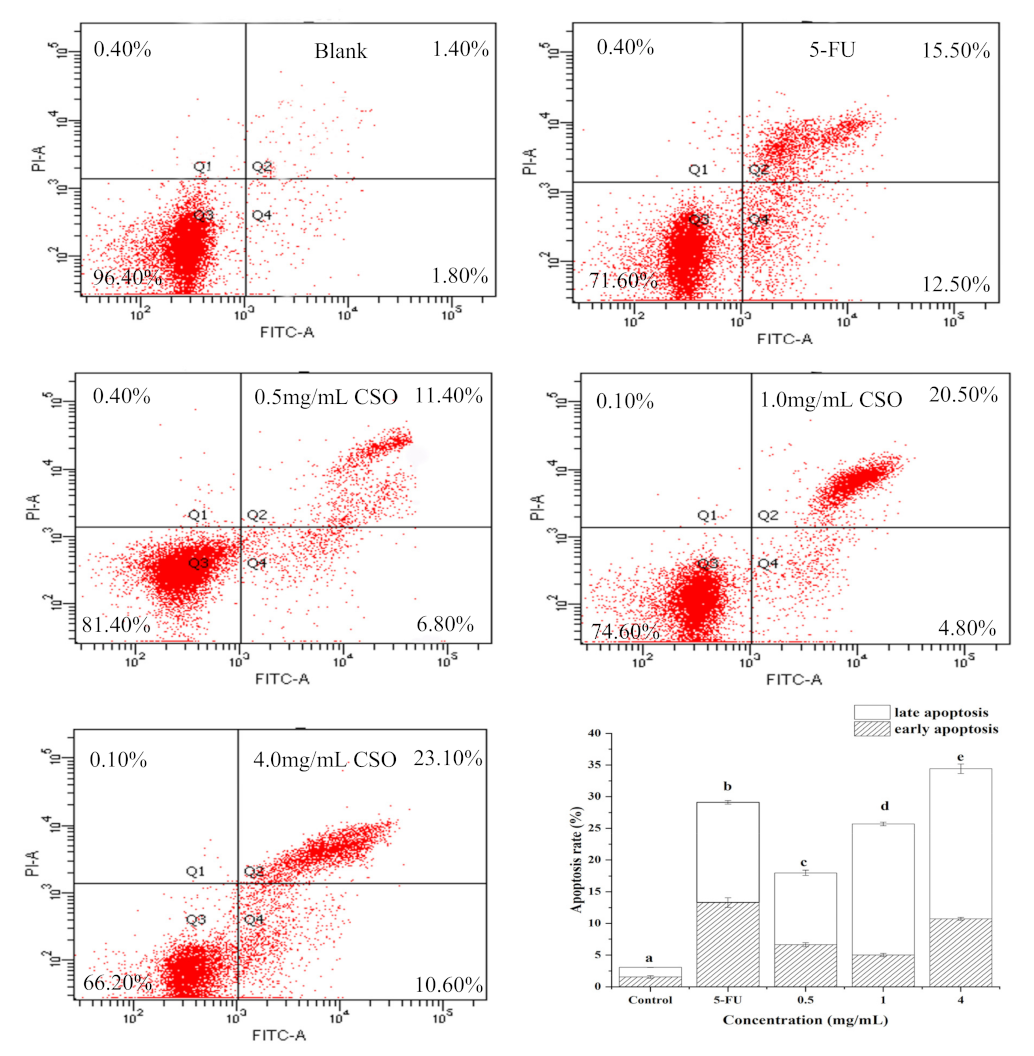
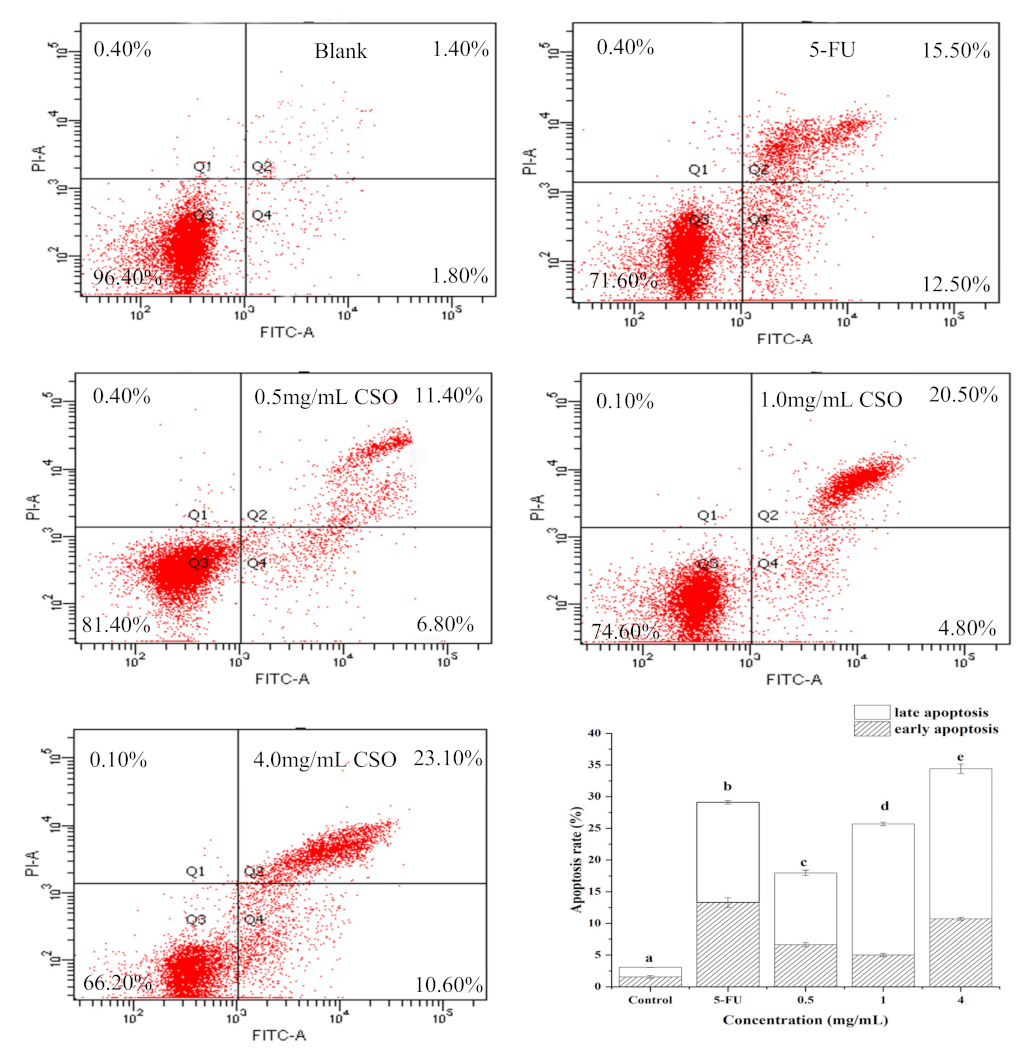
2.8. Cell Apoptosis Analysis
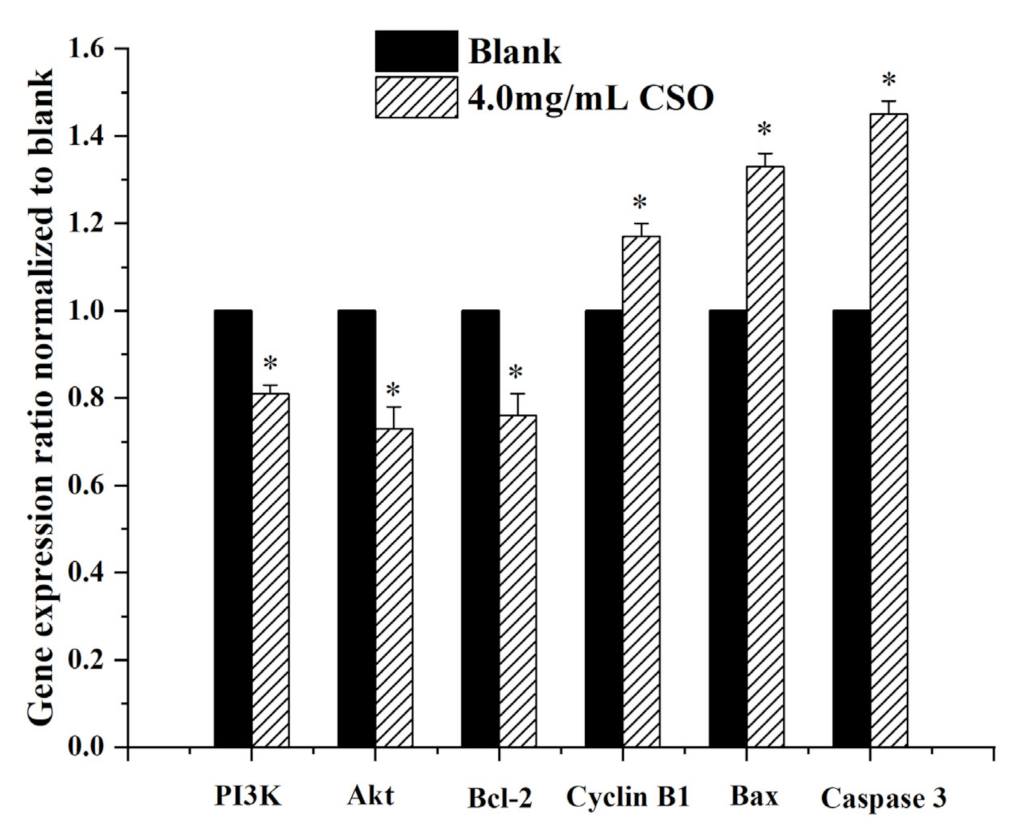
2.9. Real-Time PCR (RT-PCR)
2.10. Western Blot
2.11. Statistical Analysis
3. Results
3.1. Characterization of Coix Seed Oil
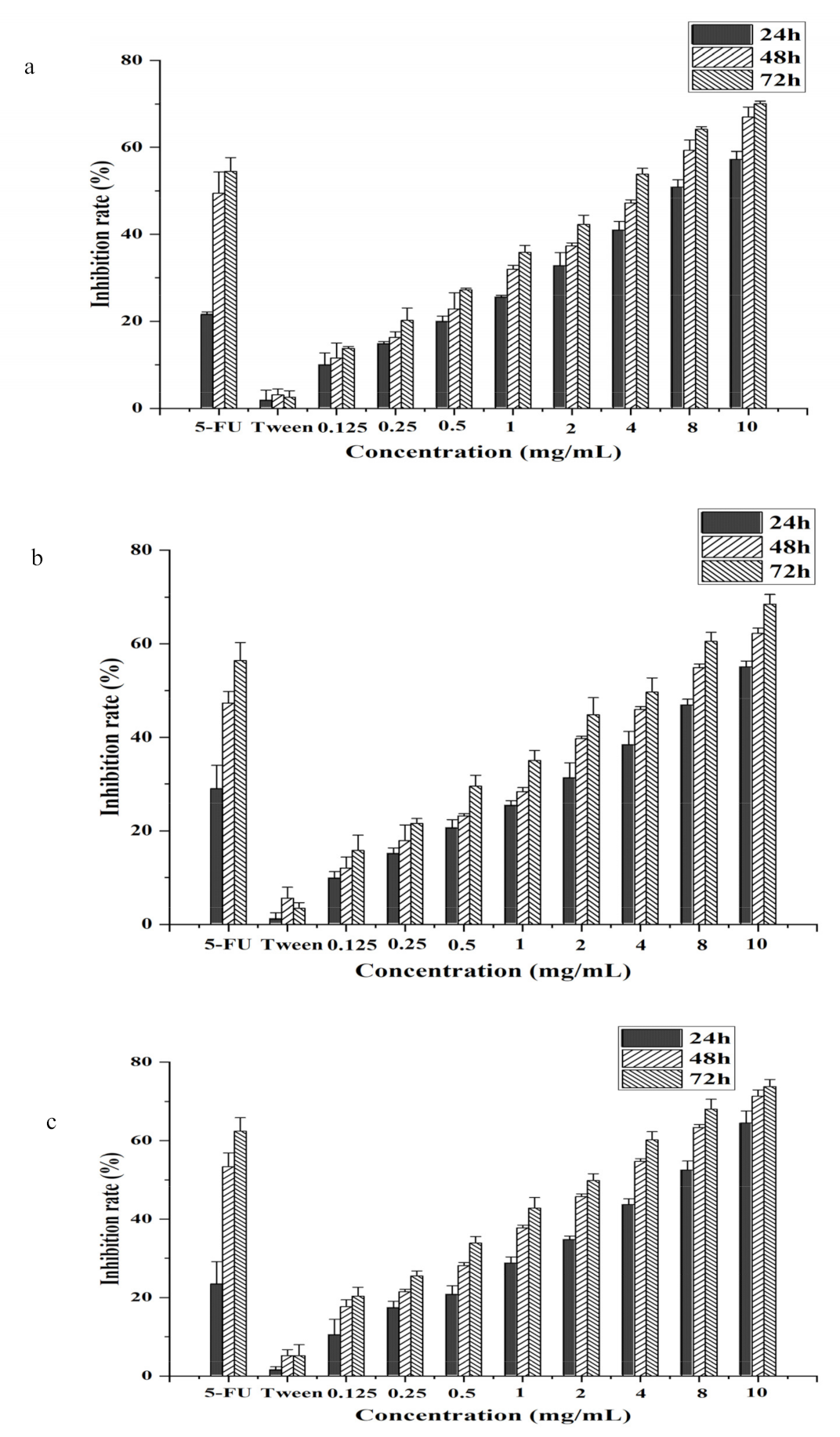
3.2. CSO Inhibited Cell Growth in HT-29, Caco-2, and HCT-116 Human Colon Cancer Cells
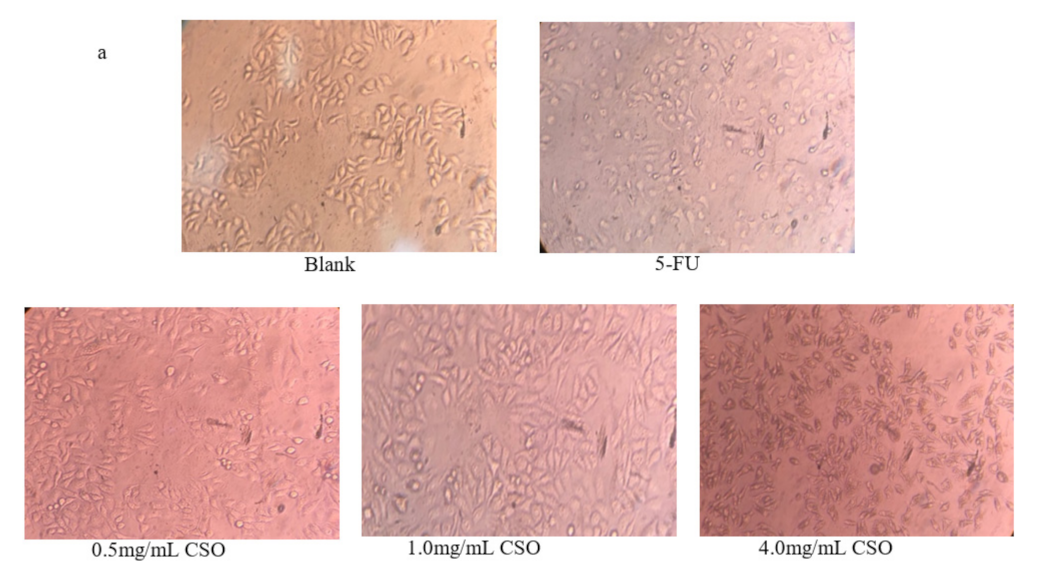
3.3. Morphology Observations
3.4. Apoptotic Cell Observations by Hoechst 33258 Staining
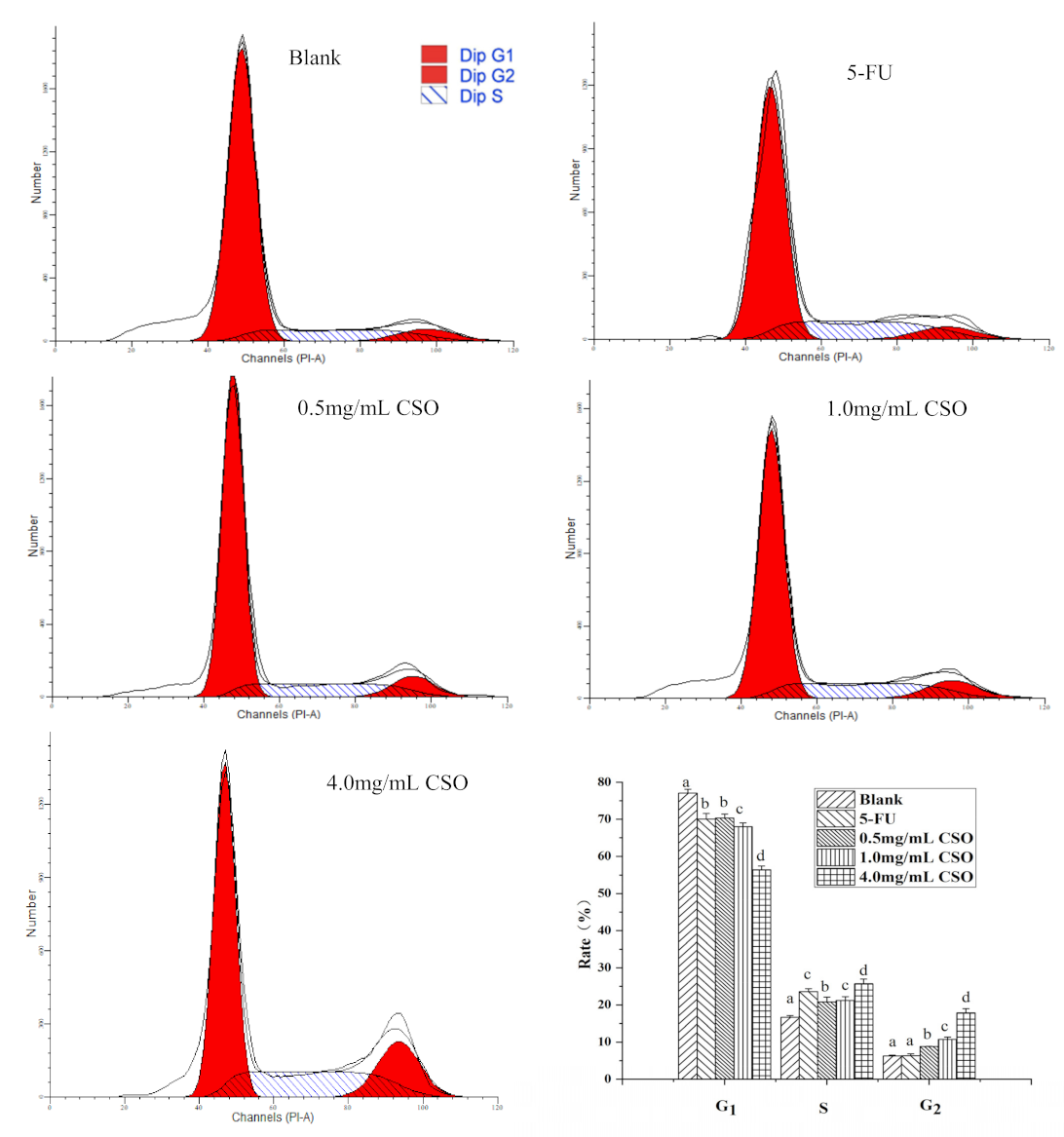
3.5. The Cell Cycle Distribution of HT-29 Cells
3.6. CSO Induced Apoptosis in HT-29 Cells
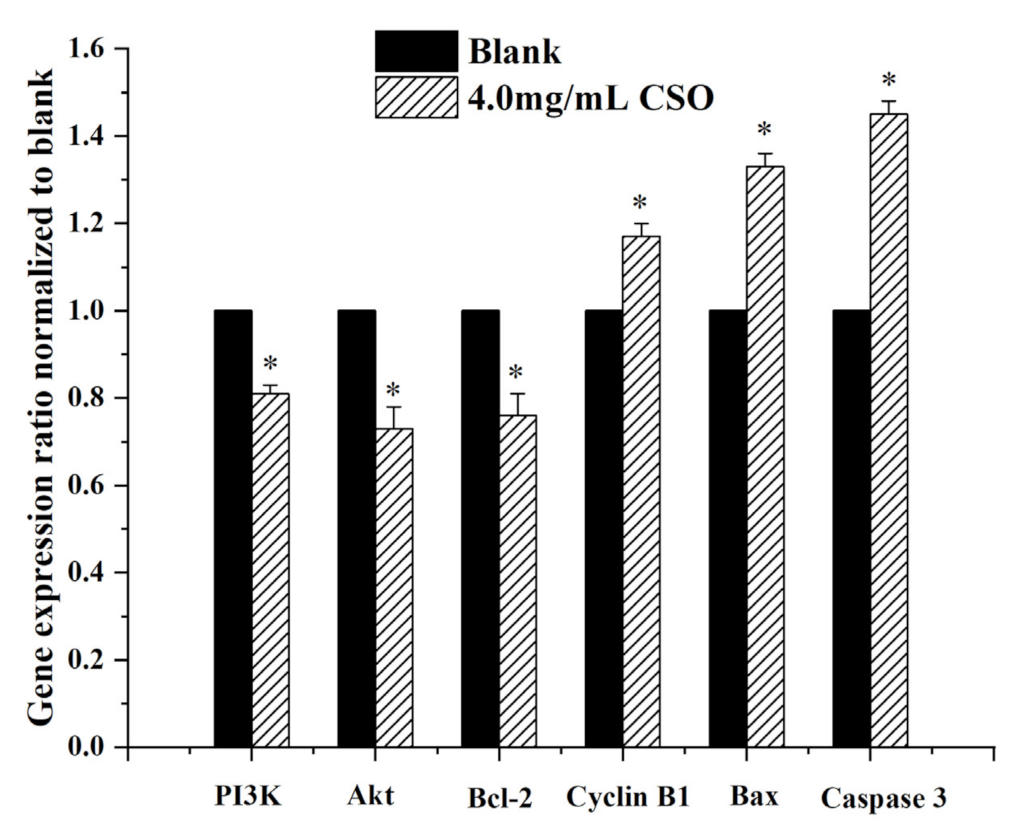
3.7. Gene Expression Levels
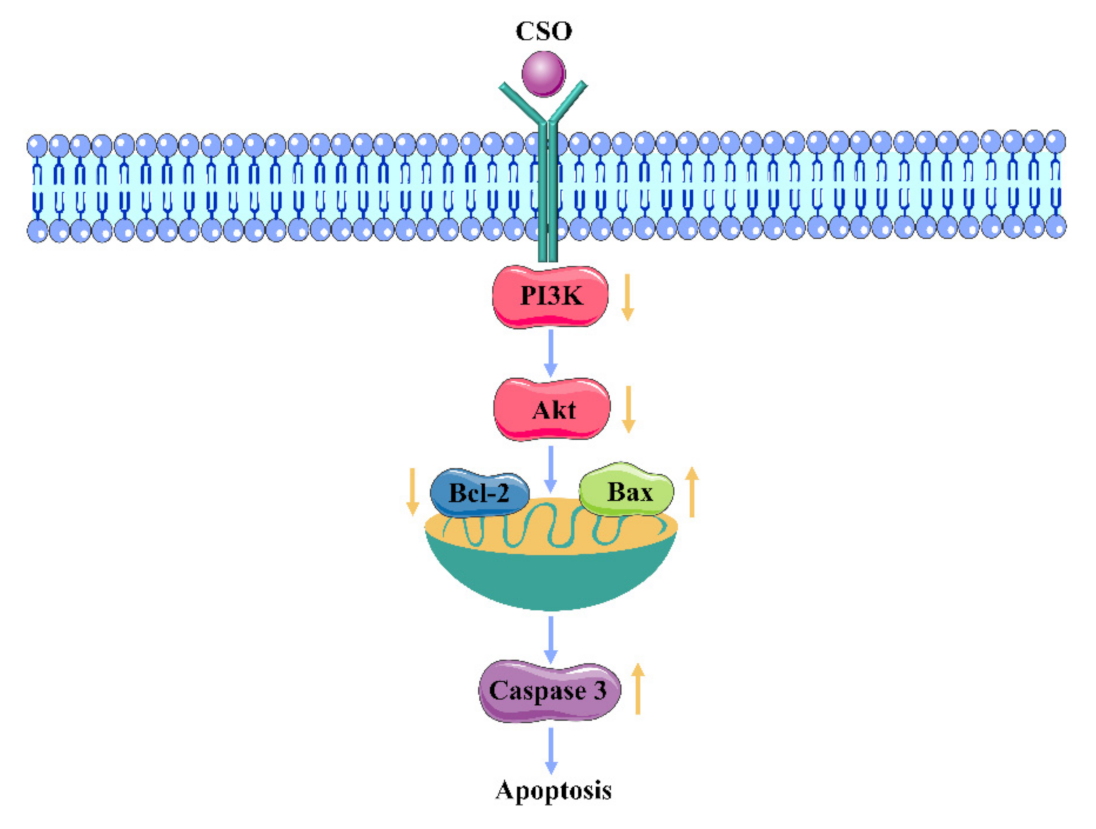
3.8. Western Blot Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Miller, K.D.; Nogueira, L.; Mariotto, A.B.; Rowland, J.H.; Siegel, R.L. Cancer treatment and survivorship statistics, 2019. CA Cancer J. Clin. 2019, 69, 363–385. [Google Scholar] [CrossRef] [Green Version]
- Keum, N.N.; Giovannucci, E. Global burden of colorectal cancer: Emerging trends, risk factors and prevention strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef]
- Xiang, L.-P.; Wang, A.; Ye, J.-H.; Zheng, X.-Q.; Polito, C.A.; Lu, J.-L.; Li, Q.-S.; Liang, Y.-R. Suppressive Effects of Tea Catechins on Breast Cancer. Nutrients 2016, 8, 458. [Google Scholar] [CrossRef]
- Kim, M.K.; Choo, H.; Chong, Y. Water-soluble and cleavable quercetin-amino acid conjugates as safe modulators for P-glycoprotein-based multidrug resistance. J. Med. Chem. 2014, 57, 7216. [Google Scholar] [CrossRef]
- Lage, N.N.; Layosa, M.A.A.; Arbizu, S.; Chew, B.P.; Pedrosa, M.L.; Mertens-Talcotta, S.; Talcotta, S.; Noratto, G.D. Dark sweet cherry (Prunus avium) phenolics enriched in anthocyanins exhibit enhanced activity against the most aggressive breast cancer subtypes without toxicity to normal breast cells. J. Funct. Foods 2020, 64, 103710. [Google Scholar] [CrossRef]
- Mark, R.; Lyu, X.; Lee, J.; Parra-Saldívar, R.; Chen, W.N. Sustainable production of natural phenolics for functional food applications. J. Funct. Foods 2019, 57, 233–254. [Google Scholar] [CrossRef]
- Kim, T.H.; Shin, Y.J.; Won, A.J.; Lee, B.M.; Choi, W.S.; Jung, J.H.; Chung, H.Y.; Kim, H.S. Resveratrol enhances chemosensitivity of doxorubicin in multidrug-resistant human breast cancer cells via increased cellular influx of doxorubicin. Biochim. Biophys. Acta 2014, 1840, 615–625. [Google Scholar] [CrossRef]
- Lin, L.C.; Wu, C.H.; Shieh, T.M.; Chen, H.Y.; Huang, T.C.; Hsia, S.M. The licorice dietary component isoliquiritigenin chemosensitizes human uterine sarcoma cells to doxorubicin and inhibits cell growth by inducing apoptosis and autophagy via inhibition of m-TOR signaling. J. Funct. Foods 2017, 33, 332–344. [Google Scholar] [CrossRef]
- Wang, X.; Pan, J.; Wang, P.; Yang, C.S.; Wang, X.; Feng, Q.; Goli, S. EGCG Enhances Cisplatin Sensitivity by Regulating Expression of the Copper and Cisplatin Influx Transporter CTR1 in Ovary Cancer. PLoS ONE 2015, 10, e0125402. [Google Scholar]
- Clark, A.M.; Magawa, C.; Pliego-Zamora, A.; Low, P.; Reynolds, M.; Ralph, S.J. Tea tree oil extract causes mitochondrial superoxide production and apoptosis as an anticancer agent, promoting tumor infiltrating neutrophils cytotoxic for breast cancer to induce tumor regression. Biomed. Pharmacother. 2021, 140, 111790. [Google Scholar] [CrossRef] [PubMed]
- Jaradat, N.; Al-Maharik, N.; Abdallah, S.; Shawahna, R.; Mousa, A.; Qtishat, A. Nepeta curviflora essential oil: Phytochemical composition, antioxidant, anti-proliferative and anti-migratory efficacy against cervical cancer cells, and α-glucosidase, α-amylase and porcine pancreatic lipase inhibitory activities. Ind. Crop. Prod. 2020, 158, 112946. [Google Scholar] [CrossRef]
- Li, X.; Wang, P.; Zhu, J.; Yi, J.; Ji, Z.; Kang, Q.; Hao, L.; Huang, J.; Lu, J. Comparative study on the bioactive components and in vitro biological activities of three green seedlings. Food Chem. 2020, 321, 126716. [Google Scholar] [CrossRef]
- Xu, Y.; Zhu, X.; Ma, X.; Xiong, H.; Zeng, Z.; Peng, H.; Hu, J. Enzymatic production of trans-free shortening from coix seed oil, fully hydrogenated palm oil and Cinnamomum camphora seed oil. Food Biosci. 2018, 22, 1–8. [Google Scholar] [CrossRef]
- Zhu, F. Coix: Chemical composition and health effects. Trends Food Sci. Technol. 2017, 61, 160–175. [Google Scholar] [CrossRef]
- Chung, C.P.; Hsu, C.Y.; Lin, J.H.; Kuo, Y.H.; Chiang, W.; Lin, Y.L. Antiproliferative lactams and spiroenone from adlay bran in human breast cancer cell lines. J. Agric. Food Chem. 2011, 59, 1185–1194. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.; Peng, H.; Xiong, H.; Liu, Y.; Zhao, L.; Xiao, X. Carboxymethylchitosan-coated proliposomes containing coix seed oil: Characterisation, stability and in vitro release evaluation. Food Chem. 2011, 129, 1695–1702. [Google Scholar] [CrossRef]
- He, W.; Yin, M.; Yang, R.; Zhao, W. Optimization of adlay (Coix lacryma-jobi) bran oil extraction: Variability in fatty acids profile and fatty acid synthase inhibitory activities. Biocatal. Agric. Biotechnol. 2020, 28, 101740. [Google Scholar] [CrossRef]
- Yu, F.; Zhang, J.; Li, Y.; Zhao, Z.; Liu, C. Research and application of adlay in medicinal field. Chin. Herb. Med. 2017, 9, 126–133. [Google Scholar] [CrossRef]
- Huang, X.; Qin, J.; Lu, S. Kanglaite stimulates anticancer immune responses and inhibits HepG2 cell transplantation-induced tumor growth. Mol. Med. Rep. 2014, 10, 2153–2159. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Gao, J.; Zeng, Y.; Liu, C.-X. Inhibition of Coix seed extract on fatty acid synthase, a novel target for anticancer activity. J. Ethnopharmacol. 2008, 119, 252–258. [Google Scholar] [CrossRef]
- Xi, X.J.; Zhu, Y.G.; Tong, Y.P.; Yang, X.L.; Cheng, Z. Assessment of the Genetic Diversity of Different Job’s Tears (Coix lacryma-jobi L.) Accessions and the Active Composition and Anticancer Effect of Its Seed Oil. PLoS ONE 2016, 11, e0153269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, D.; Liu, M.; Huang, M.; Wang, L.; Yan, C.; Liu, C.; Liu, Y. Octanoyl galactose ester-modified microemulsion system self-assembled by coix seed components to enhance tumor targeting and hepatoma therapy. Int. J. Nanomed. 2017, 12, 2045–2059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fei, F.; Wan, Y.; Wu, T. Kanglaite injection combined with hepatic arterial intervention for unresectable hepatocellular carcinoma: A meta-analysis. J. Cancer Res. Ther. 2014, 1 (Suppl. S1), 38–41. [Google Scholar]
- Wang, Y.; Zhang, C.; Zhang, S.; Zhao, Z.; Wang, J.; Song, J.; Wang, Y.; Liu, J.; Hou, S. Author Correction: Kanglaite sensitizes colorectal cancer cells to Taxol via NF-κΒ inhibition and connexin 43 upregulation. Sci. Rep. 2018, 8, 6141. [Google Scholar] [CrossRef] [Green Version]
- Qu, D.; He, J.; Liu, C.; Zhou, J.; Yan, C. Triterpene-loaded microemulsion using Coix lacryma-jobi seed extract as oil phase for enhanced antitumor efficacy: Preparation and in vivo evaluation. Int. J. Nanomed. 2014, 9, 109–119. [Google Scholar]
- Porta, C.; Paglino, C.; Mosca, A. Targeting PI3K/Akt/mTOR Signaling in Cancer. Front. Oncol. 2014, 4, 64. [Google Scholar] [CrossRef] [Green Version]
- Jafari, M.; Ghadami, E.; Dadkhah, T.; Akhavan-Niaki, H. PI3k/AKT signaling pathway: Erythropoiesis and beyond. J. Cell. Physiol. 2018, 234, 2373–2385. [Google Scholar] [CrossRef]
- Lee, H.J.; Saralamma, V.V.G.; Kim, S.M. Pectolinarigenin Induced Cell Cycle Arrest, Autophagy, and Apoptosis in Gastric Cancer Cell via PI3K/AKT/mTOR Signaling Pathway. Nutrients 2018, 10, 1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, A.; Zhang, Z.; Zheng, J.; Wang, Y.; Chen, Q.; Liu, R.; Liu, X.; Zhang, S. Optimizations and comparison of two supercritical extractions of adlay oil. Innov. Food Sci. Emerg. Technol. 2012, 13, 128–133. [Google Scholar] [CrossRef]
- Ratnayake, W.M.N.; Hansen, S.L.; Kennedy, M.P. Evaluation of the CP-Sil 88 and SP-2560 GC columns used in the recently approved AOCS Official Method Ce 1h-05: Determination of c/s-, trans-, saturated, monounsaturated, and polyunsaturated fatty acids in vegetable or non-ruminant animal oils and fats by. J. Am. Oil Chem. Soc. 2006, 83, 475–488. [Google Scholar] [CrossRef]
- Ciro, Y.; Rojas, J.; di Virgilio, A.L.; Alhajj, M.J.; Carabali, G.A.; Salamanca, C.H. Production, physicochemical characterization, and anticancer activity of methotrexate-loaded phytic acid-chitosan nanoparticles on HT-29 human colon adenocarcinoma cells. Carbohydr. Polym. 2020, 243, 116436. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Ren, Y.; Li, L. The relationship between charge intensity and bioactivities/processing characteristics of exopolysaccharides from lactic acid bacteria. LWT-Food sci. Technol. 2022, 153, 112345. [Google Scholar] [CrossRef]
- Fan, J.; Li, B.-R.; Zhang, Q.; Zhao, X.-H.; Wang, L. Pretreatment of IEC-6 cells with quercetin and myricetin resists the indomethacin-induced barrier dysfunction via attenuating the calcium-mediated JNK/Src activation. Food Chem. Toxicol. 2021, 147, 111896. [Google Scholar] [CrossRef]
- Khan, S.; Ansari, A.A.; Malik, A.; Chaudhary, A.A.; Syed, J.B.; Khan, A.A. Preparation, characterizations and in vitro cytotoxic activity of nickel oxide nanoparticles on HT-29 and SW620 colon cancer cell lines. J. Trace Elem. Med. Biol. 2019, 52, 12–17. [Google Scholar] [CrossRef]
- Yang, C.; Hou, A.; Yu, C.; Dai, L.; Wang, W.; Zhang, K.; Shao, H.; Ma, J.; Xu, W. Kanglaite reverses multidrug resistance of HCC by inducing apoptosis and cell cycle arrest via PI3K/AKT pathway. Oncotargets Ther. 2018, 11, 983–996. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Hou, Y.; Yin, Y.; Wang, F.; Yang, M.; Zhang, Y.; Fan, C.; Sun, B. CCT128930 induces G1-phase arrest and apoptosis and synergistically enhances the anticancer efficiency of VS5584 in human osteosarcoma cells. Biomed. Pharmacother. 2020, 130, 110544. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Wang, H.; Xu, F.; Zhang, Y.; Li, Z.; Ju, X.; Wang, L. Insoluble-bound polyphenols of adlay seed ameliorate H2O2-induced oxidative stress in HepG2 cells via Nrf2 signalling. Food Chem. 2020, 325, 126865. [Google Scholar] [CrossRef]
- Hamdan, N.T.; Jwad, B.A.A.A.A.; Jasim, S.A. Synergistic anticancer effects of phycocyanin and Citrullus colocynthis extract against WiDr, HCT-15 and HCT-116 colon cancer cell lines. Gene Rep. 2021, 22, 100972. [Google Scholar] [CrossRef]
- Bie, N.; Han, L.; Wang, Y.; Wang, X.; Wang, C. A polysaccharide from Grifola frondosa fruit body induces HT-29 cells apoptosis by PI3K/AKT-MAPKs and NF-κB-pathway. Int. J. Biol. Macromol. 2020, 147, 79–88. [Google Scholar] [CrossRef]
- Timilsena, Y.P.; Vongsvivut, J.; Adhikari, R.; Adhikari, B. Physicochemical and thermal characteristics of Australian chia seed oil. Food Chem. 2017, 228, 394–402. [Google Scholar] [CrossRef]
- Ali, M.; Al-Hattab, T.; Al-Hydary, I. Extraction of date palm seed oil (Phoenix dactylifera) by soxhlet apparatus. Int. J. Adv. Eng.Technol. 2015, 8, 261–271. [Google Scholar]
- Liu, J.; Chen, P.; He, J.; Deng, L.; Wang, L.; Lei, J.; Rong, L. Extraction of oil from Jatropha curcas seeds by subcritical fluid extraction. Ind. Crop. Prod. 2014, 62, 235–241. [Google Scholar] [CrossRef]
- Abdul, R.; Che, M.Y.B. Determination of extra virgin olive oil in quaternary mixture using FTIR spectroscopy and multivariate calibration. Spectroscopy 2011, 26, 203–211. [Google Scholar]
- Lee, S.Y.; Debnath, T.; Kim, S.-K.; Lim, B.O. Anti-cancer effect and apoptosis induction of cordycepin through DR3 pathway in the human colonic cancer cell HT-29. Food Chem. Toxicol. 2013, 60, 439–447. [Google Scholar] [CrossRef]
- Chang, W.L.; Cheng, F.C.; Wang, S.P.; Chou, S.T.; Ying, S. Cinnamomum cassia essential oil and its major constituent cinnamaldehyde induced cell cycle arrest and apoptosis in human oral squamous cell carcinoma HSC-3 cells. Environ. Toxicol. 2017, 32, 456–468. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.L.; Wei, Y.Q.; Wang, X.J.; Fei, X.; Li, S.F.; Jie, Z. Growth inhibition and apoptosis induction of tanshinone II-A on human hepatocellular carcinoma cells. World J. Gastroenterol. 2004, 10, 2024–2028. [Google Scholar] [CrossRef] [PubMed]
- Hoxhaj, G.; Manning, B.D. The PI3K–AKT network at the interface of oncogenic signalling and cancer metabolism. Nat. Rev. Cancer 2020, 20, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Deng, F.; Zhao, L.; Du, H.; Qing, Z. Characterization of extractable components of fresh and fermented Huarong large-leaf mustard and their inhibitory effects on human colon cancer cells. Food Biosci. 2021, 43, 101280. [Google Scholar] [CrossRef]
- Ma, X.; Wang, Z. Anticancer drug discovery in the future: An evolutionary perspective. Drug Discov. Today 2009, 14, 1136–1142. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.-J.; Koh, W.; Kim, B.; Kim, S.-H. Are there new therapeutic options for treating lung cancer based on herbal medicines and their metabolites? J. Ethnopharmacol. 2011, 138, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Tettamanti, G.; Carata, E.; Montali, A.; Dini, L.; Fimia, G.M. Autophagy in development and regeneration: Role in tissue remodelling and cell survival. Eur. Zool. J. 2019, 86, 113–131. [Google Scholar] [CrossRef] [Green Version]
- Bao, R.K.; Zheng, S.F.; Wang, X.Y. Selenium protects against cadmium-induced kidney apoptosis in chickens by activating the PI3K/AKT/Bcl-2 signaling pathway. Environ. Sci. Pollut. Res. 2017, 24, 20342–20353. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Qin, D.; Yang, H.; Wang, W.; Xiao, J.; Zhou, L.; Fu, H. Neuroprotective Effects of Deuterium-Depleted Water (DDW) Against H 2 O 2 -Induced Oxidative Stress in Differentiated PC12 Cells Through the PI3K/Akt Signaling Pathway. Neurochem. Res. 2020, 45, 1034–1044. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Cui, H.; Xi, P.; Jing, F.; Zuo, Z.; Deng, J.; Wang, X.; Wu, B.; Chen, K.; Jie, D. Modulation of the PI3K/Akt Pathway and Bcl-2 Family Proteins Involved in Chicken’s Tubular Apoptosis Induced by Nickel Chloride (NiCl2). Int. J. Mol. Sci. 2015, 16, 22989–23011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, E.S.; Kim, Y.O.; Park, C.G.; Park, K.H.; Jeong, S.H.; Park, J.W.; Kim, S.H. Coix lacryma-jobi var. ma-yuen Stapf sprout extract has anti-metastatic activity in colon cancer cells in vitro. BMC Complementary Altern. Med. 2017, 17, 486. [Google Scholar] [CrossRef] [Green Version]
- Trejo-Solís, C.; Serrano-Garcia, N.; Escamilla-Ramírez, Á.; Castillo-Rodríguez, R.; Jimenez-Farfan, D.; Palencia, G.; Calvillo, M.; Alvarez-Lemus, M.; Flores-Nájera, A.; Cruz-Salgado, A. Autophagic and Apoptotic Pathways as Targets for Chemotherapy in Glioblastoma. Int. J. Mol. Sci. 2018, 19, 3773. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Zheng, Y.; Wang, R.; Huang, W.; Liu, L.; Hu, X.; Liu, S.; Yue, J.; Tong, T.; Jing, X. Tong Antitumor activity of folate-targeted, paclitaxel-loaded polymeric micelles on a human esophageal EC9706 cancer cell line. Int. J. Nanomed. 2012, 7, 3487–3502. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid Composition | CSO |
|---|---|
| Saturated fatty acid | 14.46 ± 0.48 |
| Caprylic acid (C8:0) | 0.07 ± 0.01 |
| Myristic acid (C14:0) | 0.17 ± 0.01 |
| Palmitic acid (C16:0) | 11.74 ± 0.5 |
| Stearic acid (C18:0) | 2.45 ± 0.06 |
| Arachidic acid (C20:0) | 0.03 ± 0.01 |
| Monounsaturated fatty acid | 51.15 ± 0.73 |
| Palmitoleic acid (C16:1 cis9) | 0.16 ± 0.04 |
| Oleic acid (C18:1 cis9) | 50.54 ± 0.63 |
| Arachidonic acid (C20:1 cis11) | 0.45 ± 0.06 |
| Polyunsaturated fatty acid | 33.95 ± 0.07 |
| Linoleic acid (C18:2 cis9,12) (ω-6) | 33.76 ± 0.21 |
| α-linolenic acid(C18:3 cis9,12,15)(ω-3) | 0.19 ± 0.01 |
| Odd carbon fatty acid | 0.44 ± 0.11 |
| Undecanoic acid (C11:0) | 0.02 ± 0.00 |
| 10-Heptadecenoicacid (C17:1 cis10) | 0.04 ± 0.01 |
| Heptadecanoic acid (C17:0) | 0.09 ± 0.03 |
| Tricosanoicacid(C23:0) | 0.29 ± 0.05 |
| (C16:0 + C18:2)/C18:1 | 0.90 |
| Unsaturated fatty acid/Saturated fatty acid | 5.88 |
| IC50(mg/mL) | |||
|---|---|---|---|
| 24 h | 48 h | 72 h | |
| Caco-2 | 7.00 ± 0.34 a | 4.00 ± 0.21 a | 2.84 ± 0.21 a |
| HCT-116 | 8.74 ± 0.17 b | 4.80 ± 0.22 b | 3.00 ± 025 a |
| HT-29 | 5.30 ± 0.21 c | 2.56 ± 0.30 c | 1.78 ± 0.11 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ni, C.; Li, B.; Ding, Y.; Wu, Y.; Wang, Q.; Wang, J.; Cheng, J. Anti-Cancer Properties of Coix Seed Oil against HT-29 Colon Cells through Regulation of the PI3K/AKT Signaling Pathway. Foods 2021, 10, 2833. https://doi.org/10.3390/foods10112833
Ni C, Li B, Ding Y, Wu Y, Wang Q, Wang J, Cheng J. Anti-Cancer Properties of Coix Seed Oil against HT-29 Colon Cells through Regulation of the PI3K/AKT Signaling Pathway. Foods. 2021; 10(11):2833. https://doi.org/10.3390/foods10112833
Chicago/Turabian StyleNi, Chunlei, Bailiang Li, Yangyue Ding, Yue Wu, Qiuye Wang, Jiarong Wang, and Jianjun Cheng. 2021. "Anti-Cancer Properties of Coix Seed Oil against HT-29 Colon Cells through Regulation of the PI3K/AKT Signaling Pathway" Foods 10, no. 11: 2833. https://doi.org/10.3390/foods10112833
APA StyleNi, C., Li, B., Ding, Y., Wu, Y., Wang, Q., Wang, J., & Cheng, J. (2021). Anti-Cancer Properties of Coix Seed Oil against HT-29 Colon Cells through Regulation of the PI3K/AKT Signaling Pathway. Foods, 10(11), 2833. https://doi.org/10.3390/foods10112833