
Green Pea (Pisum sativum L.) Hull Polyphenol Extracts Ameliorate DSS-Induced Colitis through Keap1/Nrf2 Pathway and Gut Microbiota Modulation

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Preparation of GPH Extracts
2.3. Quantitative Analysis by UHPLC-LTQ-Orbitrap-MS/MS
2.3.1. Liquid Chromatographic Conditions
2.3.2. Mass Spectrometric Conditions
2.3.3. Calibration and Quantification
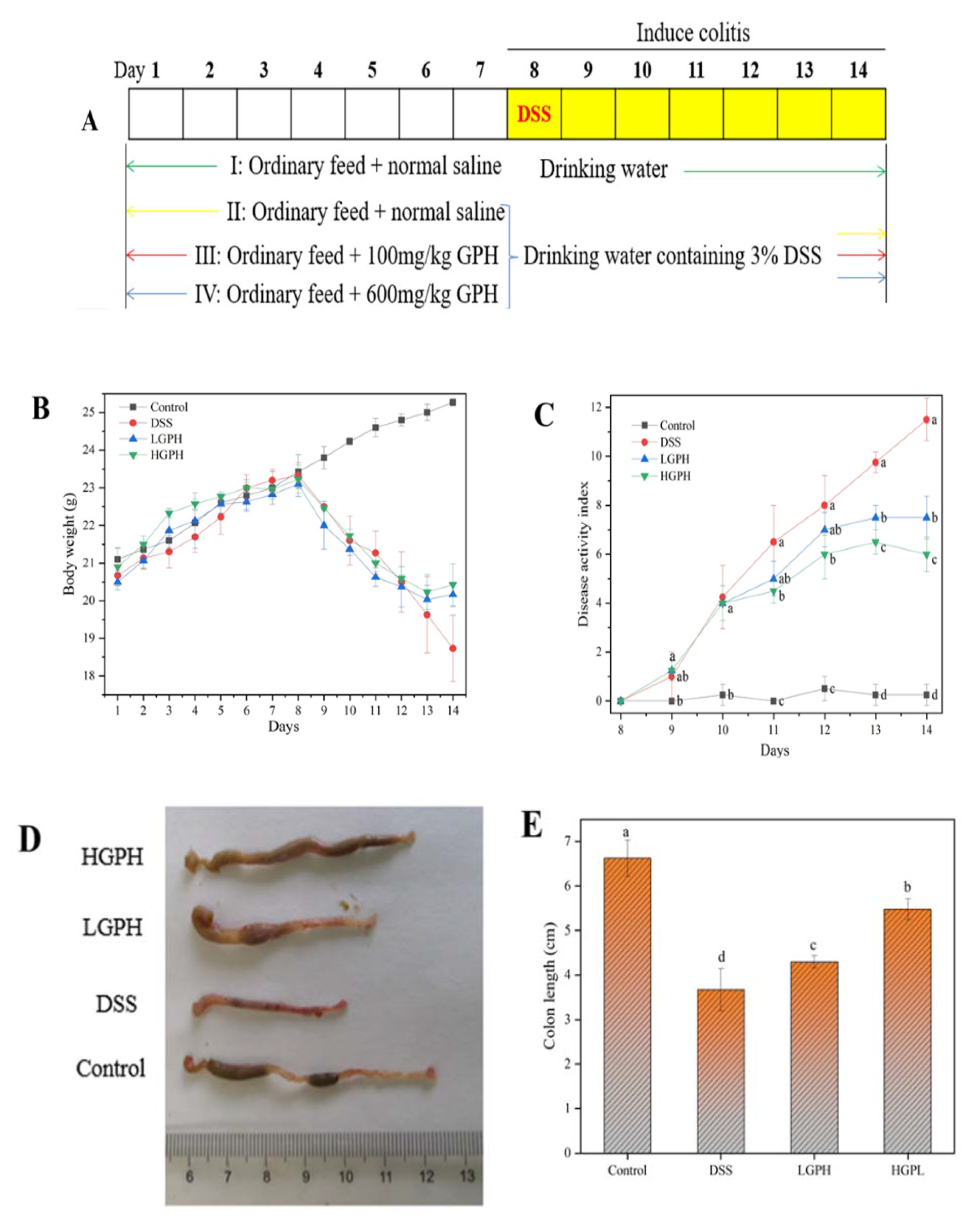
2.4. Experimental Animals
2.4.1. Disease Activity Index (DAI) Assessment
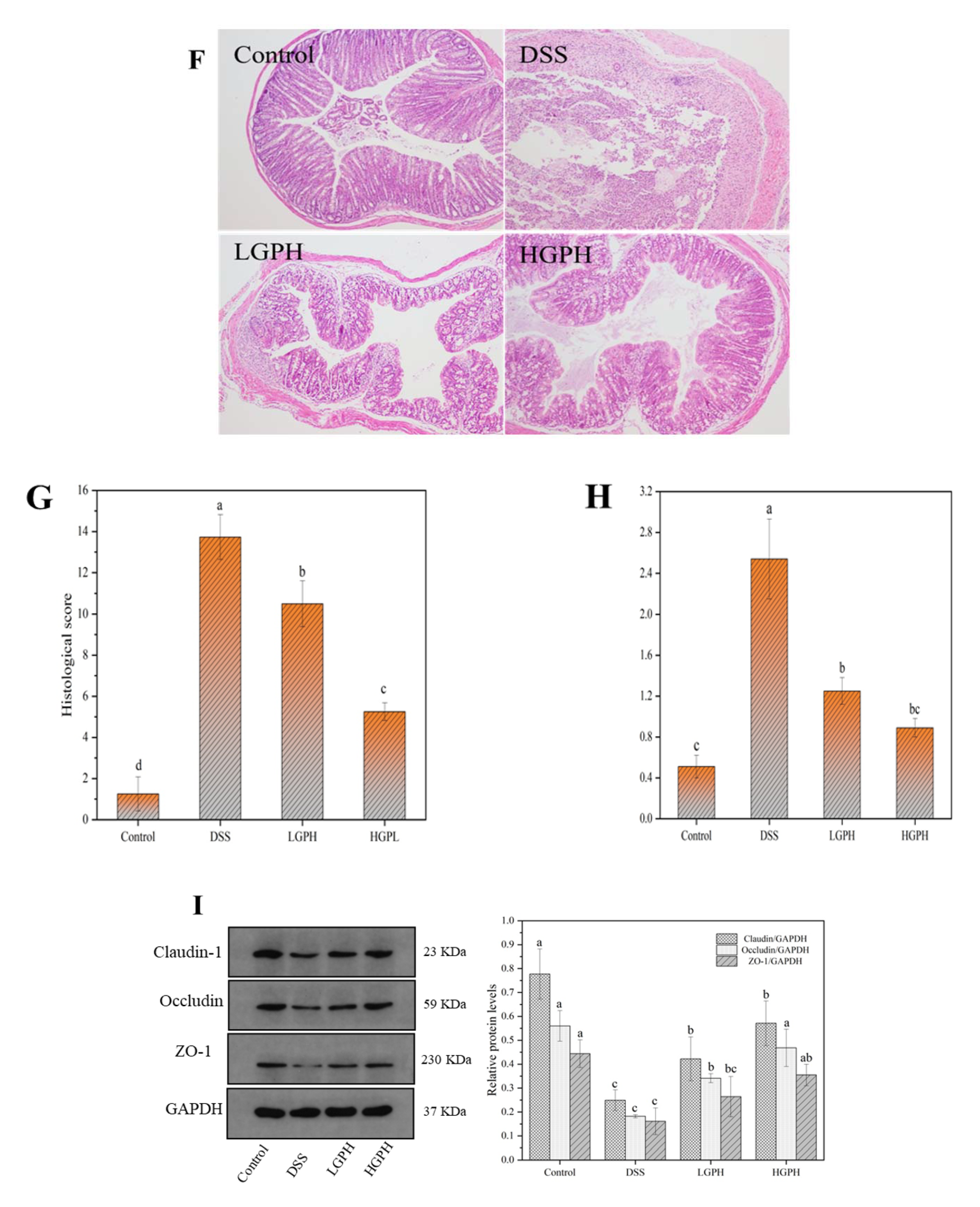
2.4.2. Evaluation of Histological Changes
2.4.3. MPO Assessment
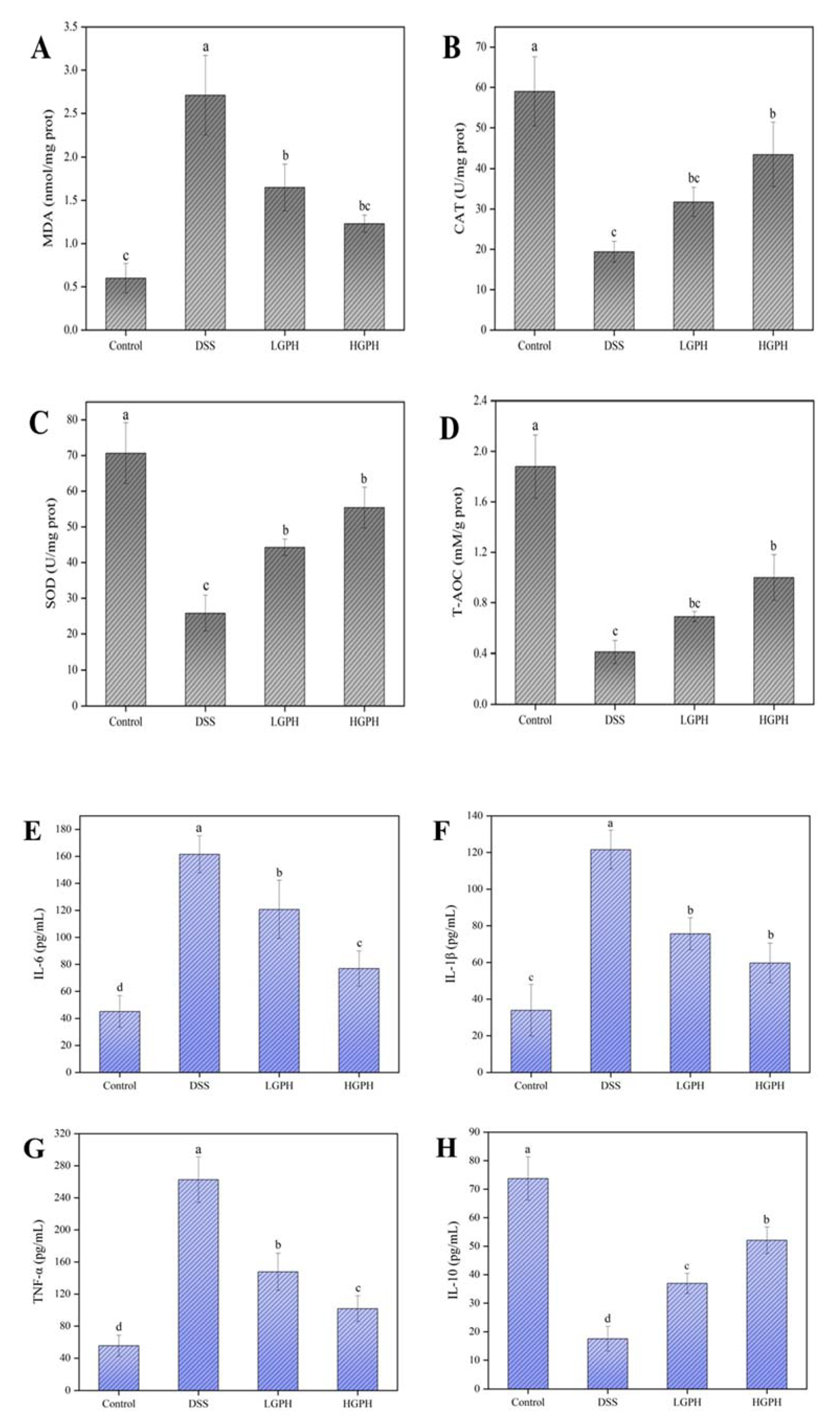
2.4.4. Oxidative Stress Markers
2.4.5. Inflammatory Cytokines
2.4.6. Western Blot Analysis
2.4.7. Quantitative Real-Time Polymerase Chain Reaction (PCR) Analysis
2.4.8. SCFAs Analysis
2.4.9. Gut Microbiota Analysis
2.5. Statistical Analysis
3. Results
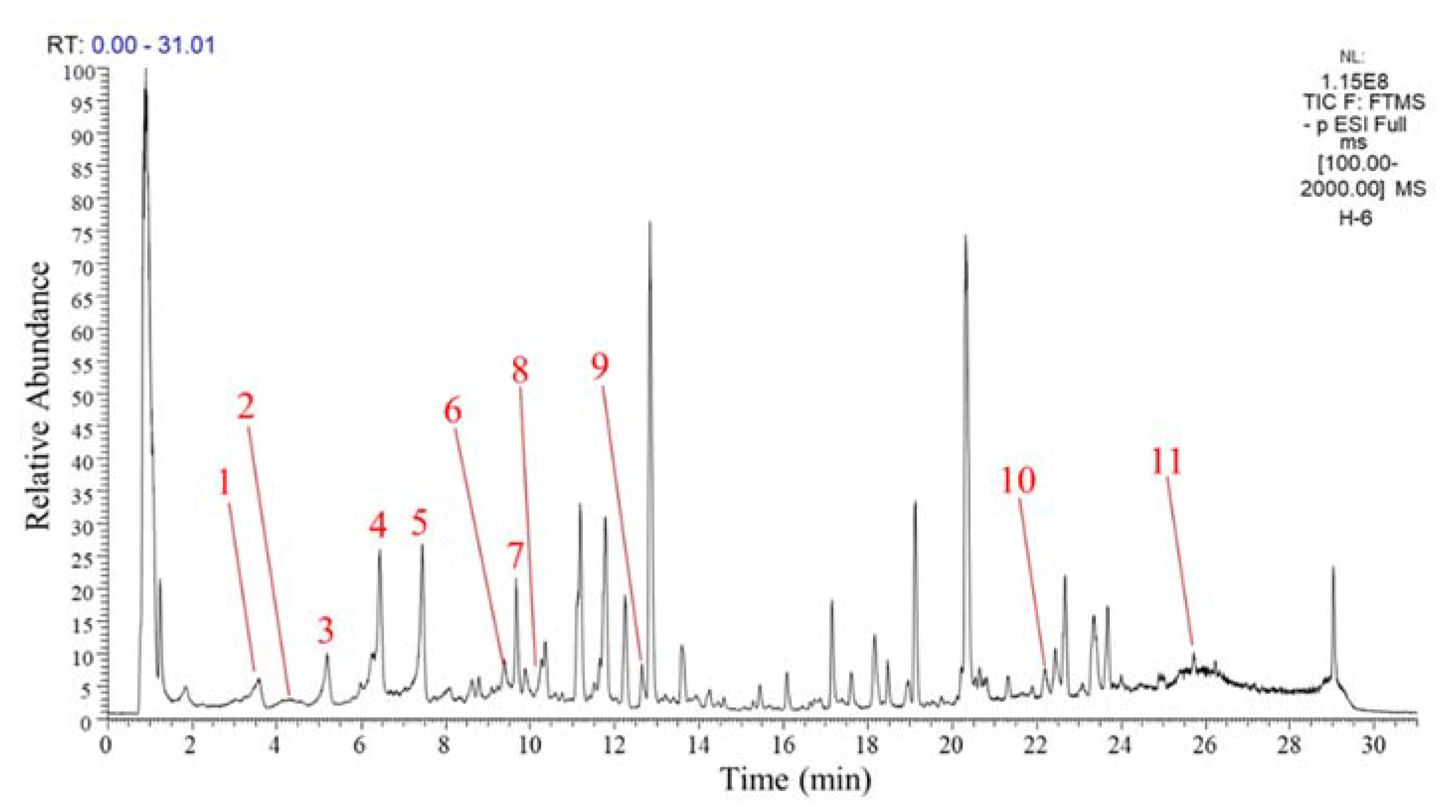
3.1. Quantification of Polyphenols in GPH Extracts
3.2. Anti-Inflammatory Effects of GPH Extracts in Mice with DSS-Induced Colitis
3.2.1. GPH Extracts Mitigated the Symptoms of DSS-Induced UC Mice
3.2.2. GPH Extracts Relieved the Colonic Injury and Inflammatory Infiltration in DSS-Induced UC Mice
3.2.3. GPH Extracts Protected Colon from Oxidative Stress in DSS-Induced UC Mice
3.2.4. GPH Extracts Improved the Inflammatory Status in DSS-Induced UC Mice
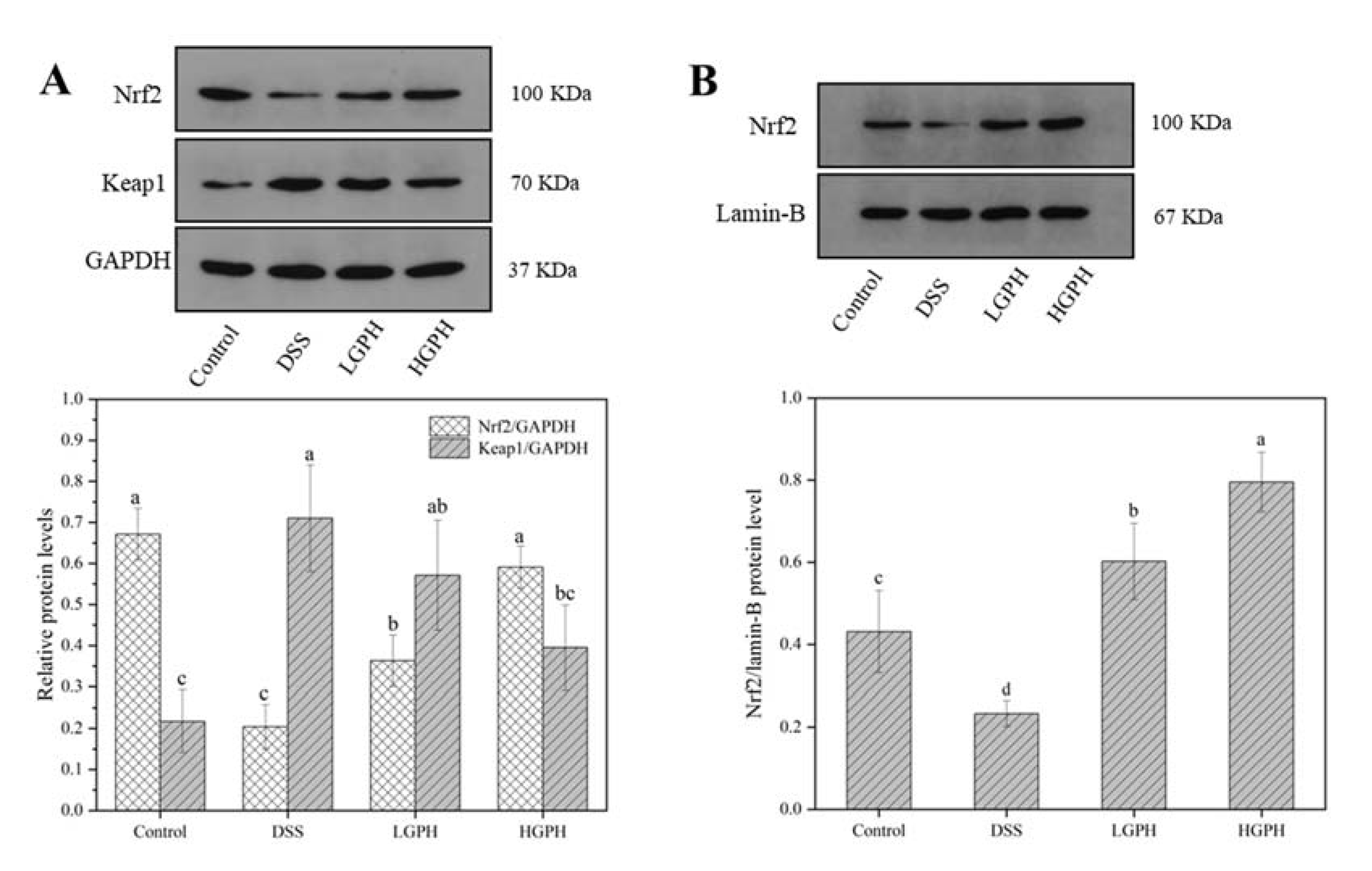
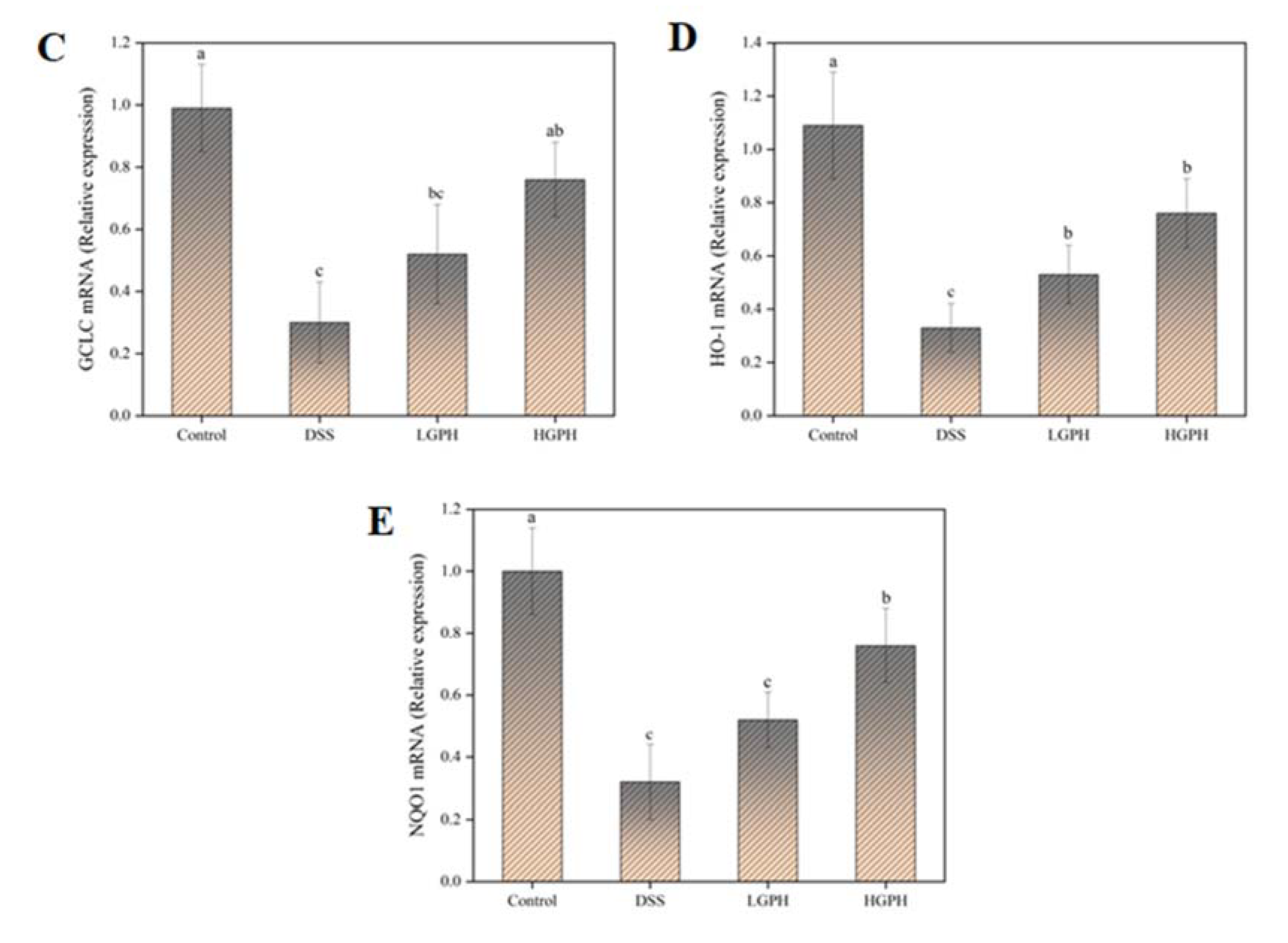
3.2.5. GPH Extracts Activated the Keap1/Nrf2 Signaling Pathway and Promoted the mRNA Expression of Nrf2 Downstream Genes
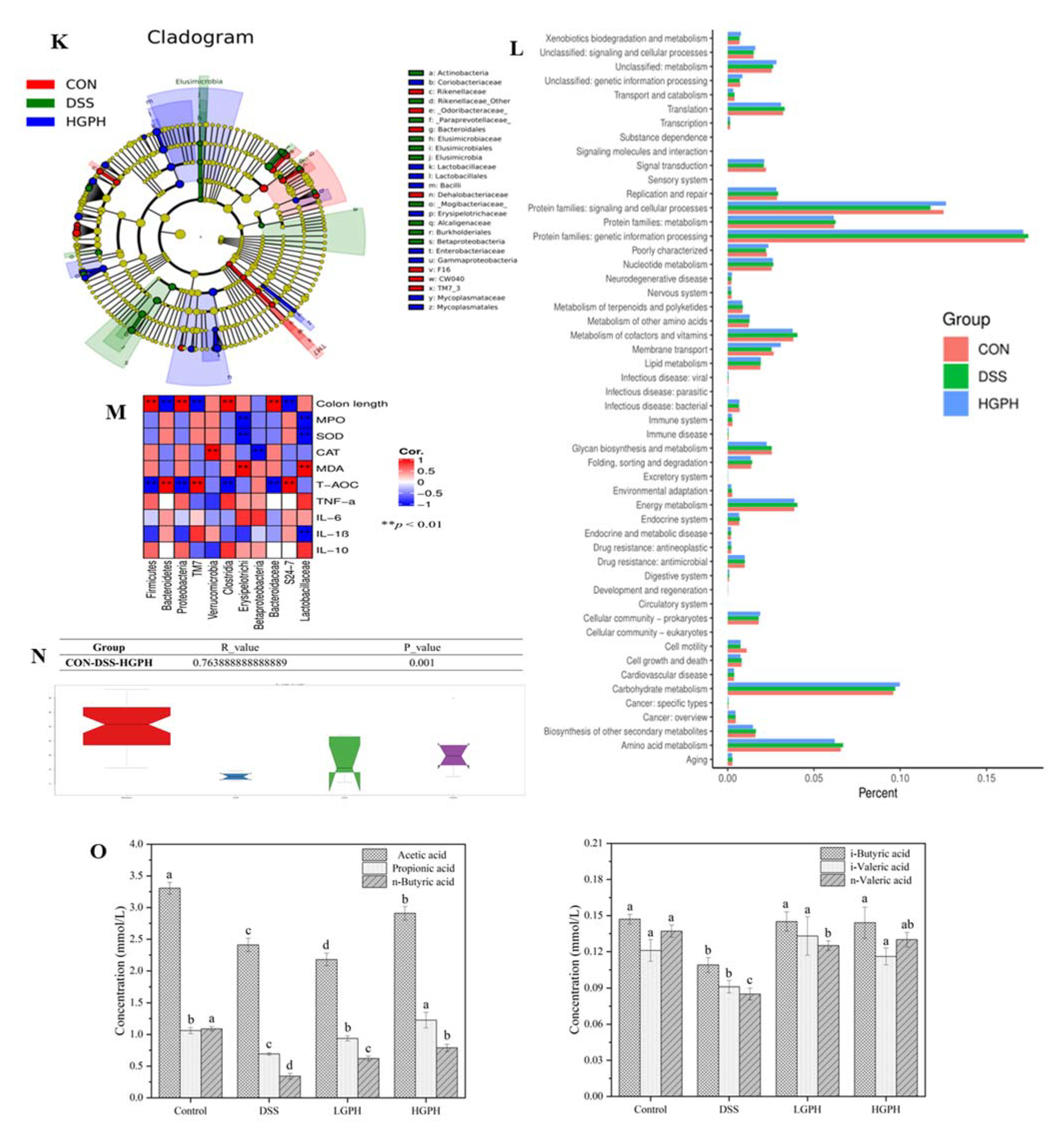
3.2.6. GPH Extracts Increased SCFAs Content in DSS-Induced UC Mice
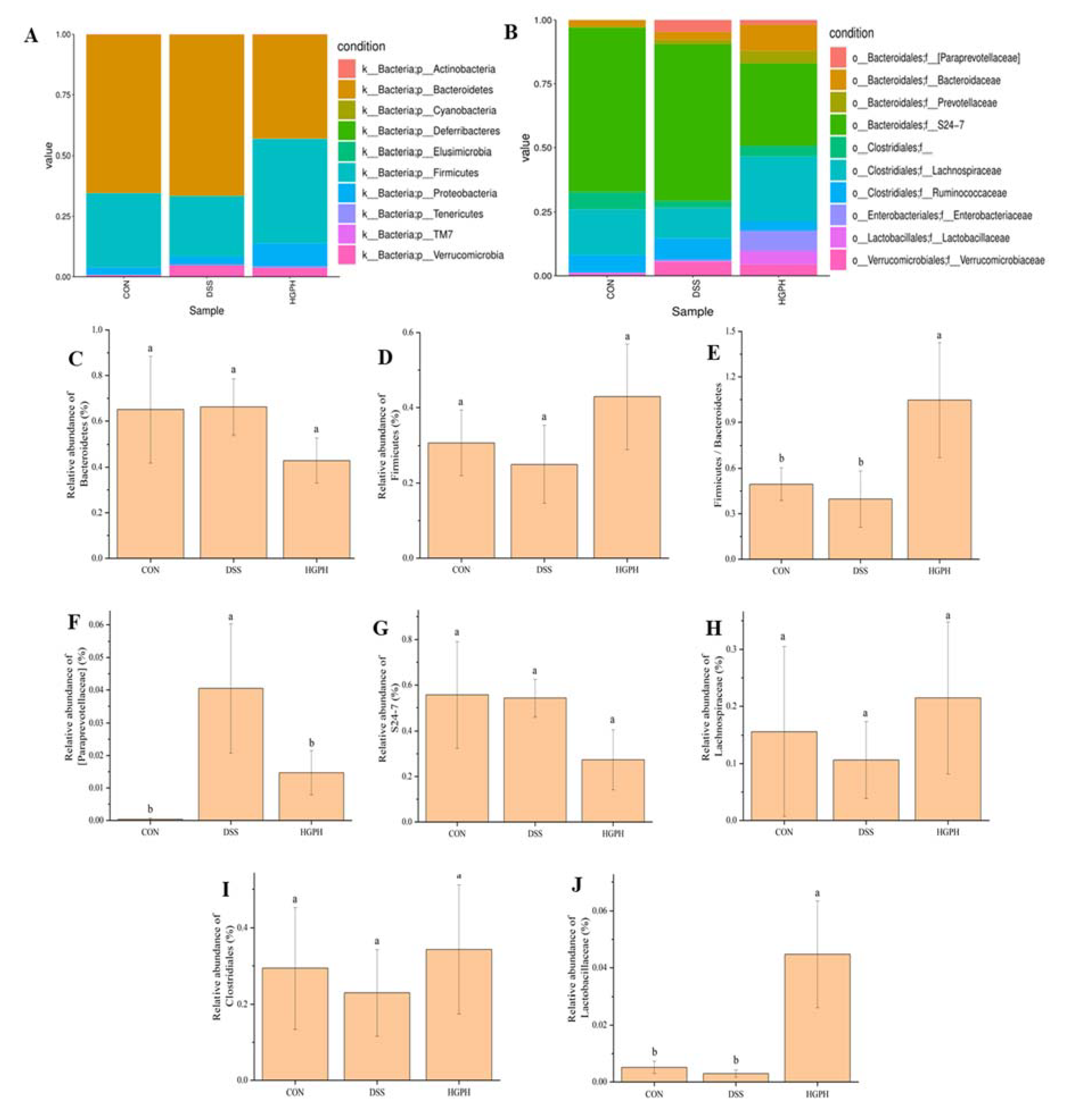
3.3. GPH Extracts Altered Gut Microbiota in Mice with DSS-Induced Colitis
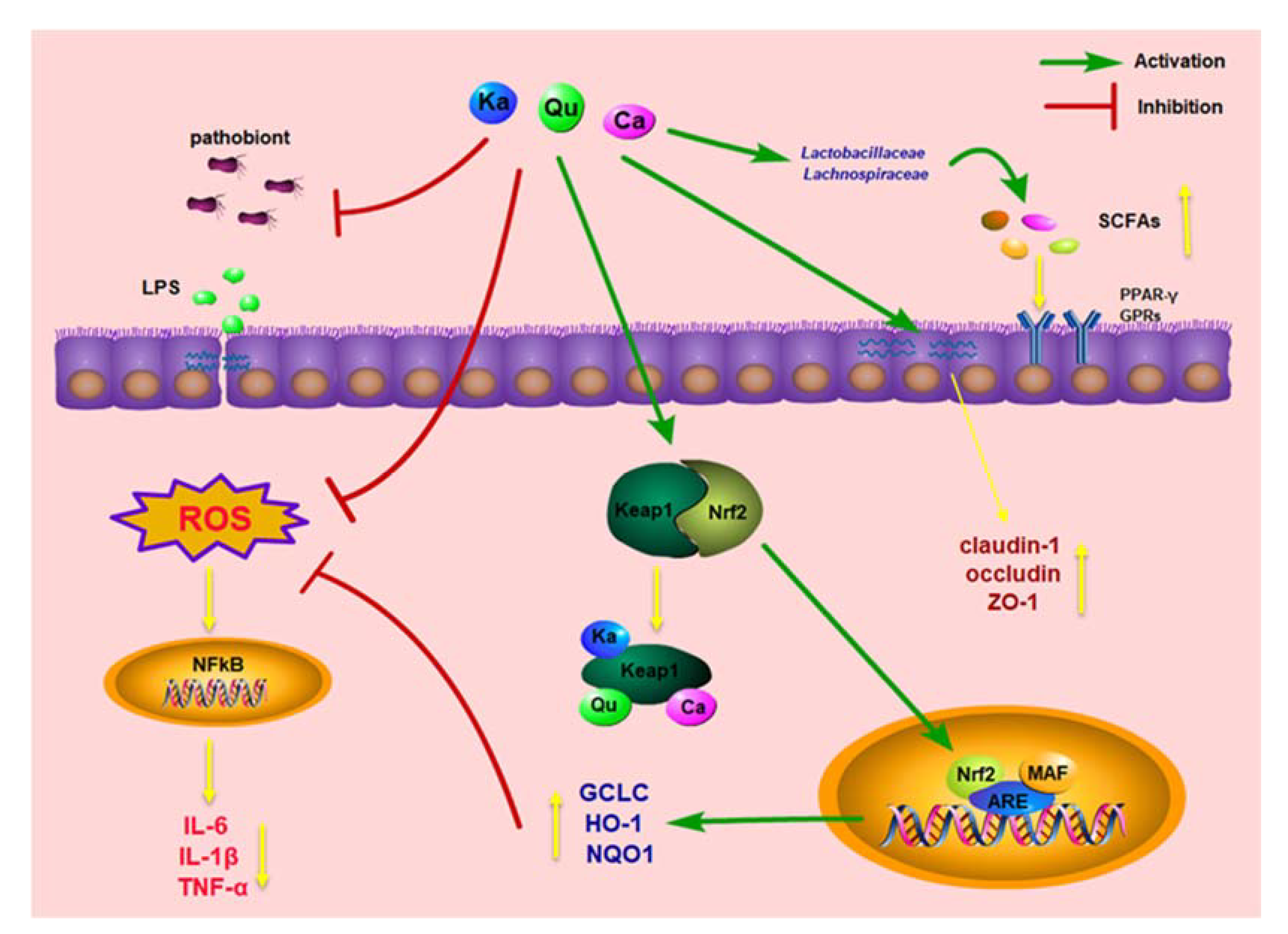
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giang, J.; Lan, X.; Crichton, M.; Marx, W.; Marshall, S. Efficacy and safety of biophenol-rich nutraceuticals in adults with inflammatory gastrointestinal diseases or irritable bowel syndrome: A systematic literature review and meta-analysis. Nutr. Diet. 2021, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zheng, X.; Chen, J.; Luo, D.; Xie, J.; Su, Z.; Huang, X.; Yi, X.; Wei, L.; Cai, J.; et al. Protective Effect of Bruguiera gymnorrhiza (L.) Lam. Fruit on Dextran Sulfate Sodium-Induced Ulcerative Colitis in Mice: Role of Keap1/Nrf2 Pathway and Gut Microbiota. Front. Pharmacol. 2020, 10, 1602. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Mytton, J.; Evison, F.; Kamarajah, S.K.; Reece, J.; Iqbal, T.; Cooney, R.; Thompson, F.; Walmsley, M.; Ferguson, J. A nationwide population-based evaluation of mortality and cancer-risk in patients with ulcerative colitis/primary scle-rosing cholangitis—Young age at diagnosis and the unmet need to reduce mortality. J. Hepatol. 2018, 68, S220–S221. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, T.; Sun, D.; Xie, C.; Wang, T.; Liu, X.; Wang, J.; Wang, Q.; Luo, Y.; Wang, P.; et al. Rutaecarpine inhibits KEAP1-NRF2 interaction to activate NRF2 and ameliorate dextran sulfate sodium-induced colitis. Free. Radic. Biol. Med. 2020, 148, 33–41. [Google Scholar] [CrossRef]
- Wang, X.; Yu, N.; Peng, H.; Hu, Z.; Sun, Y.; Zhu, X.; Jiang, L.; Xiong, H. The profiling of bioactives in Akebia trifoliata pericarp and metabolites, bioavailability and in vivo anti-inflammatory activities in DSS-induced colitis mice. Food Funct. 2019, 10, 3977–3991. [Google Scholar] [CrossRef] [PubMed]
- Russo, L.; Schneider, G.; Gardiner, M.H.; Lanes, S.; Streck, P.; Rosen, S. Role of pharmacoepidemiology studies in ad-dressing pharmacovigilance questions: A case example of pancreatitis risk among ulcerative colitis patients using mesala-zine. Eur. J. Clin. Pharmacol. 2014, 70, 709–717. [Google Scholar] [CrossRef][Green Version]
- Algieri, F.; Zorrilla, P.; Rodriguez-Nogales, A.; Mesa, N.G.; Bañuelos, Ó.; González-Tejero, R.; Casares-Porcel, M.; Mesa, J.M.; Zarzuelo, A.; Utrilla, M.P.; et al. Intestinal anti-inflammatory activity of hydroalcoholic extracts of Phlomis purpurea L. and Phlomis lychnitis L. in the trinitrobenzenesulphonic acid model of rat colitis. J. Ethnopharmacol. 2013, 146, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Nikkhah-Bodaghi, M.; Maleki, I.; Agah, S.; Hekmatdoost, A. Zingiber officinale and oxidative stress in patients with ulcerative colitis: A randomized, placebo-controlled, clinical trial. Complement. Ther. Med. 2019, 43, 1–6. [Google Scholar] [CrossRef]
- Lu, M.-C.; Ji, J.-A.; Jiang, Y.-L.; Chen, Z.-Y.; Yuan, Z.-W.; You, Q.-D.; Jiang, Z. An inhibitor of the Keap1-Nrf2 protein-protein interaction protects NCM460 colonic cells and alleviates experimental colitis. Sci. Rep. 2016, 6, 26585. [Google Scholar] [CrossRef]
- Kitakaze, T.; Makiyama, A.; Samukawa, Y.; Jiang, S.; Yamashita, Y.; Ashida, H. A physiological concentration of luteolin induces phase II drug-metabolizing enzymes through the ERK1/2 signaling pathway in HepG2 cells. Arch. Biochem. Biophys. 2019, 663, 151–159. [Google Scholar] [CrossRef]
- Zhou, Y.; Jiang, Z.; Lu, H.; Xu, Z.; Tong, R.; Shi, J.; Jia, G. Recent Advances of Natural Polyphenols Activators for Keap1-Nrf2 Signaling Pathway. Chem. Biodivers. 2019, 16, e1900400. [Google Scholar] [CrossRef]
- de Paula do Nascimento, R.; da Fonseca Machado, A.P.; Galvez, J.; Cazarin, C.B.B.; Junior, M.R.M. Ulcerative colitis: Gut microbiota, immunopathogenesis and application of natural products in animal models. Life Sci. 2020, 258, 118129. [Google Scholar] [CrossRef]
- LeBlanc, J.-F.; Segal, J.; Braz, L.D.C.; Hart, A. The Microbiome as a Therapy in Pouchitis and Ulcerative Colitis. Nutrients 2021, 13, 1780. [Google Scholar] [CrossRef] [PubMed]
- Miao, R.-R.; Zhan, S.; Hu, X.-T.; Yuan, W.-M.; Wu, L.-J.; Cui, S.-X.; Qu, X.-J. Myricetin and M10, a myricetin-3-O-β-d-lactose sodium salt, modify composition of gut microbiota in mice with ulcerative colitis. Toxicol. Lett. 2021, 346, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Yu, N.X.; Wang, Z.L.; Qiu, T.T.; Jiang, L.; Zhu, X.M.; Sun, Y.; Xiong, H. Akebia trifoliata pericarp extract ameliorates inflammation through NF-kappa B/MAPK signaling pathways and modifies gut microbiota. Food Funct. 2020, 11, 4682–4696. [Google Scholar] [CrossRef] [PubMed]
- Shang, L.; Liu, H.; Yu, H.; Chen, M.; Yang, T.; Zeng, X.; Qiao, S. Core Altered Microorganisms in Colitis Mouse Model: A Comprehensive Time-Point and Fecal Microbiota Transplantation Analysis. Antibiotics 2021, 10, 643. [Google Scholar] [CrossRef]
- Liu, F.; Wang, X.; Li, D.; Cui, Y.; Li, X. Apple polyphenols extract alleviated dextran sulfate sodium-induced ulcerative colitis in C57BL /6 male mice by restoring bile acid metabolism disorder and gut microbiota dysbiosis. Phytother. Res. 2021, 35, 1468–1485. [Google Scholar] [CrossRef]
- Zhao, H.; Cheng, N.; Zhou, W.; Chen, S.; Wang, Q.; Gao, H.; Xue, X.; Wu, L.; Cao, W. Honey Polyphenols Ameliorate DSS-Induced Ulcerative Colitis via Modulating Gut Microbiota in Rats. Mol. Nutr. Food Res. 2019, 63, e1900638. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wu, X.; Cao, S.; Cromie, M.; Shen, Y.; Feng, Y.; Yang, H.; Li, L. Chlorogenic Acid Ameliorates Experimental Colitis by Promoting Growth of Akkermansia in Mice. Nutrients 2017, 9, 677. [Google Scholar] [CrossRef]
- Zhang, Z.; Wu, X.; Cao, S.; Wang, L.; Wang, D.; Yang, H.; Feng, Y.; Wang, S.; Shoulin, W. Caffeic acid ameliorates colitis in association with increased Akkermansia population in the gut microbiota of mice. Oncotarget 2016, 7, 31790–31799. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, Q.; Mi, X.; Qiu, L.; Tao, X.; Zhang, Z.; Xia, J.; Wu, Q.; Wei, H. Ripened Pu-erh Tea Extract Promotes Gut Microbiota Resilience against Dextran Sulfate Sodium Induced Colitis. J. Agric. Food Chem. 2021, 69, 2190–2203. [Google Scholar] [CrossRef]
- Guo, F.; Tsao, R.; Wang, X.; Jiang, L.; Sun, Y.; Xiong, H. Phenolics of Yellow Pea (Pisum sativum L.) Hulls, Their Plasma and Urinary Metabolites, Organ Distribution, and In Vivo Antioxidant Activities. J. Agric. Food Chem. 2021, 69, 5013–5025. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Xiong, H.; Wang, X.; Jiang, L.; Yu, N.; Hu, Z.; Sun, Y.; Tsao, R. Phenolics of Green Pea (Pisum sativum L.) Hulls, Their Plasma and Urinary Metabolites, Bioavailability, and in Vivo Antioxidant Activities in a Rat Model. J. Agric. Food Chem. 2019, 67, 11955–11968. [Google Scholar] [CrossRef]
- Kai, W.; Jin, X.; Li, Q.; Franklan, S.; Le, L.; Conlon, M.A.; Wu, L.; Hu, F. Propolis from Different Geographic Origins Suppress Intestinal Inflammation in a Model of DSS-Induced Colitis is Associated with Decreased Bacteroides spp. in the Gut. Mol. Nutr. Food Res. 2018, 62, 1800080. [Google Scholar]
- Wang, J.; Chen, H.; Yang, B.; Gu, Z.; Zhang, H.; Chen, W.; Chen, Y.Q. Lactobacillus plantarum ZS2058 produces CLA to ameliorate DSS-induced acute colitis in mice. RSC Adv. 2016, 6, 14457–14464. [Google Scholar] [CrossRef]
- Li, S.Y.; Wang, T.M.; Wu, B.N.; Fu, W.Y.; Xu, B.J.; Pamuru, R.R.; Kennett, M.J.; Vanamala, J.K.P.; Reddivari, L. An-thocyanin-containing purple potatoes ameliorate DSS-induced colitis in mice. J. Nutr. Biochem. 2021, 93, 108616. [Google Scholar] [CrossRef]
- Bai, Y.; Jia, X.; Huang, F.; Zhang, R.; Dong, L.; Liu, L.; Zhang, M. Structural elucidation, anti-inflammatory activity and intestinal barrier protection of longan pulp polysaccharide LPIIa. Carbohydr. Polym. 2020, 246, 116532. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; He, L.; Wang, W.; Wang, T.; Hua, W.; Li, T.; Wang, L.; Gao, T.; Chen, F.; Tang, L. Polyphenols from Penthorum chinense Pursh. Attenuates high glucose-induced vascular inflammation through directly interacting with Keap1 protein. J. Ethnopharmacol. 2021, 268, 113617. [Google Scholar] [CrossRef] [PubMed]
- Rached, W.; Zeghada, F.Z.; Bennaceur, M.; Barros, L.; Calhelha, R.C.; Heleno, S.; Alves, M.J.; Carvalho, A.M.; Marouf, A.; Ferreira, I.C. Phytochemical analysis and assessment of antioxidant, antimicrobial, anti-inflammatory and cytotoxic properties of Tetraclinis articulata (Vahl) Masters leaves. Ind. Crop. Prod. 2018, 112, 460–466. [Google Scholar] [CrossRef]
- Lesjak, M.; Beara, I.; Simin, N.; Pintac, D.; Majkic, T.; Bekvalac, K.; Orcic, D.; Mimica-Dukic, N. Antioxidant and an-ti-inflammatory activities of quercetin and its derivatives. J. Funct. Foods 2018, 40, 68–75. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, Z.; Liu, J.; Zheng, Z.; Li, Q.; Wang, H.; Chen, Z.; Wang, K. Identification of the core active structure of a Dendrobium officinale polysaccharide and its protective effect against dextran sulfate sodium-induced colitis via alleviating gut microbiota dysbiosis. Food Res. Int. 2020, 137, 109641. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Hu, Q.; Wu, X.; Chen, G.; Wu, H.; Ren, J. Alginate and Probiotics Synergistically Reversed Dextran Sulfate Sodium Salt (DSS)-Induced Gut Barrier Damage. Macromol. Res. 2019, 27, 888–894. [Google Scholar] [CrossRef]
- Chen, M.; Liu, Y.; Xiong, S.; Wu, M.; Li, B.; Ruan, Z.; Hu, X. Dietary l-tryptophan alleviated LPS-induced intestinal barrier injury by regulating tight junctions in a Caco-2 cell monolayer model. Food Funct. 2019, 10, 2390–2398. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Tang, H.; Huang, F.; Qiao, Z.; Wang, X.; Yang, C.; Deng, Q. Algal Oil Rich in n-3 PUFA Alleviates DSS-Induced Colitis via Regulation of Gut Microbiota and Restoration of Intestinal Barrier. Front. Microbiol. 2020, 11, 11. [Google Scholar] [CrossRef]
- Chen, X.-M.; Kitts, D.D. Flavonoid composition of orange peel extract ameliorates alcohol-induced tight junction dysfunction in Caco-2 monolayer. Food Chem. Toxicol. 2017, 105, 398–406. [Google Scholar] [CrossRef]
- Maren, A.; Susanne, S.; Salah, A.; Joachim, M.; Martin, Z.; Michael, F.; Schulzke, J.D. Quercetin enhances epithelial barrier function and increases claudin-4 expression in Caco-2 cells. J. Nutr. 2008, 138, 1067–1073. [Google Scholar]
- Huang, G.; Wang, Z.; Wu, G.; Zhang, R.; Dong, L.; Huang, F.; Zhang, M.; Su, D. Lychee (Litchi chinensis Sonn.) Pulp Phenolics Activate the Short-Chain Fatty Acid-Free Fatty Acid Receptor Anti-inflammatory Pathway by Regulating Microbiota and Mitigate Intestinal Barrier Damage in Dextran Sulfate Sodium-Induced Colitis in Mice. J. Agric. Food Chem. 2021, 69, 3326–3339. [Google Scholar] [CrossRef]
- Rabizadeh, S.; Rhee, K.-J.; Wu, S.; Huso, D.; Gan, C.M.; Golub, J.E.; Wu, X.; Zhang, M.; Sears, C.L. Enterotoxigenic Bacteroides fragilis: A potential instigator of colitis. Inflamm. Bowel Dis. 2007, 13, 1475–1483. [Google Scholar] [CrossRef]
- Geirnaert, A.; Calatayud, M.; Grootaert, C.; Laukens, D.; Devriese, S.; Smagghe, G.; De Vos, M.; Boon, N.; Van De Wiele, T. Butyrate-producing bacteria supplemented in vitro to Crohn’s disease patient microbiota increased butyrate production and enhanced intestinal epithelial barrier integrity. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Pan, P.; Oshima, K.; Huang, Y.-W.; Agle, K.A.; Drobyski, W.R.; Chen, X.; Zhang, J.; Yearsley, M.M.; Yu, J.; Wang, L.-S. Loss of FFAR2 promotes colon cancer by epigenetic dysregulation of inflammation suppressors. Int. J. Cancer 2018, 143, 886–896. [Google Scholar] [CrossRef]
- Zheng, S.; Zhang, H.; Liu, R.; Huang, C.-L.; Li, H.; Deng, Z.-Y.; Tsao, R. Do short chain fatty acids and phenolic metabolites of the gut have synergistic anti-inflammatory effects?—New insights from a TNF-α-induced Caco-2 cell model. Food Res. Int. 2021, 139, 109833. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′-3′) | PCR Products |
|---|---|---|---|
| mus GAPDH | Forward | ATGGGTGTGAACCACGAGA | 229 bp |
| Reverse | CAGGGATGATGTTCTGGGCA | ||
| mus HO-1 | Forward | CCTCACTGGCAGGAAATCA | 217 bp |
| Reverse | TCGGGAAGGTAAAAAAAGC | ||
| mus NQO1 | Forward | GGCTGGTTTGAGAGAGTGCT | 205 bp |
| Reverse | GGAAGCCACAGAAACGCAG | ||
| mus GSTA1 | Forward | TCTCAACTACATCGCCACCA | 198 bp |
| Reverse | TCAAAGGCAGGCAAGTAACG | ||
| Mus GCLC | Forward | AAGCCTCCTCCTCCAAACTC | 187 bp |
| Reverse | GGGCCACTTTCATGTTCTCG |
| No. | tR (min) | Formula | [M − H]− (m/z) | Major Fragment Ions (m/z) | Tentative Identification | μg Quercetin/g GPH Extract |
|---|---|---|---|---|---|---|
| 1 | 3.56 | C25H24O9 | 467.1317 | 289.1351 | (Epi)catechin conjugate 1 | 496.53 ± 30.60 |
| 2 | 4.25 | C30H26O14 | 609.1241 | 305.0539, 423.0485, 441.0551, 483.0265, 515.5356, 591.1233 | (Epi)gallocatechin dimer | 214.68 ± 15.01 |
| 3 | 5.18 | C25H24O9 | 467.1312 | 289.1352, 305.0813, 351.1628 | (Epi)catechin conjugate 2 | 628.70 ± 40.06 |
| 4 | 6.44 | C20H22O15 | 501.0922 | / | Quercetin derivative 1 | 1409.24 ± 60.01 |
| 5 | 7.43 | C20H22O15 | 501.0922 | 264.9600, 294.9618, 338.9902, 380.9662, 411.0221, 483.0662 | Quercetin derivative 2 | 1381.57 ± 91.18 |
| 6 | 9.38 | C33H40O21 | 771.1999 | 609.1423, 285.0310 | Kaempferol trihexoside 1 | 492.17 ± 36.28 |
| 7 | 9.66 | C33H40O21 | 771.2006 | 609.1426, 285.0313 | Kaempferol trihexoside 2 | 989.83 ± 53.77 |
| 8 | 10.11 | C21H20O12 | 463.0887 | 301.0334 | Quercetin 3-glucoside | 45.76 ± 5.02 |
| 9 | 12.64 | C52H39O25 | 1062.7570 | 1016.7891 | Theaflavin derivative | 23.34 ± 2.52 |
| 10 | 22.18 | C27H32O15 | 595.2894 | 222.9419, 240.9657, 279.1984, 315.0365 | Isorhamnetin glycoside | 294.18 ± 25.20 |
| 11 | 25.71 | C15H12O5 | 271.2285 | 225.1866 | Naringenin | 239.15 ± 13.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, F.; Tsao, R.; Li, C.; Wang, X.; Zhang, H.; Jiang, L.; Sun, Y.; Xiong, H. Green Pea (Pisum sativum L.) Hull Polyphenol Extracts Ameliorate DSS-Induced Colitis through Keap1/Nrf2 Pathway and Gut Microbiota Modulation. Foods 2021, 10, 2765. https://doi.org/10.3390/foods10112765
Guo F, Tsao R, Li C, Wang X, Zhang H, Jiang L, Sun Y, Xiong H. Green Pea (Pisum sativum L.) Hull Polyphenol Extracts Ameliorate DSS-Induced Colitis through Keap1/Nrf2 Pathway and Gut Microbiota Modulation. Foods. 2021; 10(11):2765. https://doi.org/10.3390/foods10112765
Chicago/Turabian StyleGuo, Fanghua, Rong Tsao, Chuyao Li, Xiaoya Wang, Hua Zhang, Li Jiang, Yong Sun, and Hua Xiong. 2021. "Green Pea (Pisum sativum L.) Hull Polyphenol Extracts Ameliorate DSS-Induced Colitis through Keap1/Nrf2 Pathway and Gut Microbiota Modulation" Foods 10, no. 11: 2765. https://doi.org/10.3390/foods10112765
APA StyleGuo, F., Tsao, R., Li, C., Wang, X., Zhang, H., Jiang, L., Sun, Y., & Xiong, H. (2021). Green Pea (Pisum sativum L.) Hull Polyphenol Extracts Ameliorate DSS-Induced Colitis through Keap1/Nrf2 Pathway and Gut Microbiota Modulation. Foods, 10(11), 2765. https://doi.org/10.3390/foods10112765