
The Bactericidal Efficacy and the Mechanism of Action of Slightly Acidic Electrolyzed Water on Listeria monocytogenes’ Survival


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of SAEW
2.2. Preparation of Bacterial Strains and Culture
2.3. Determination of Antibacterial Effect of SAEW
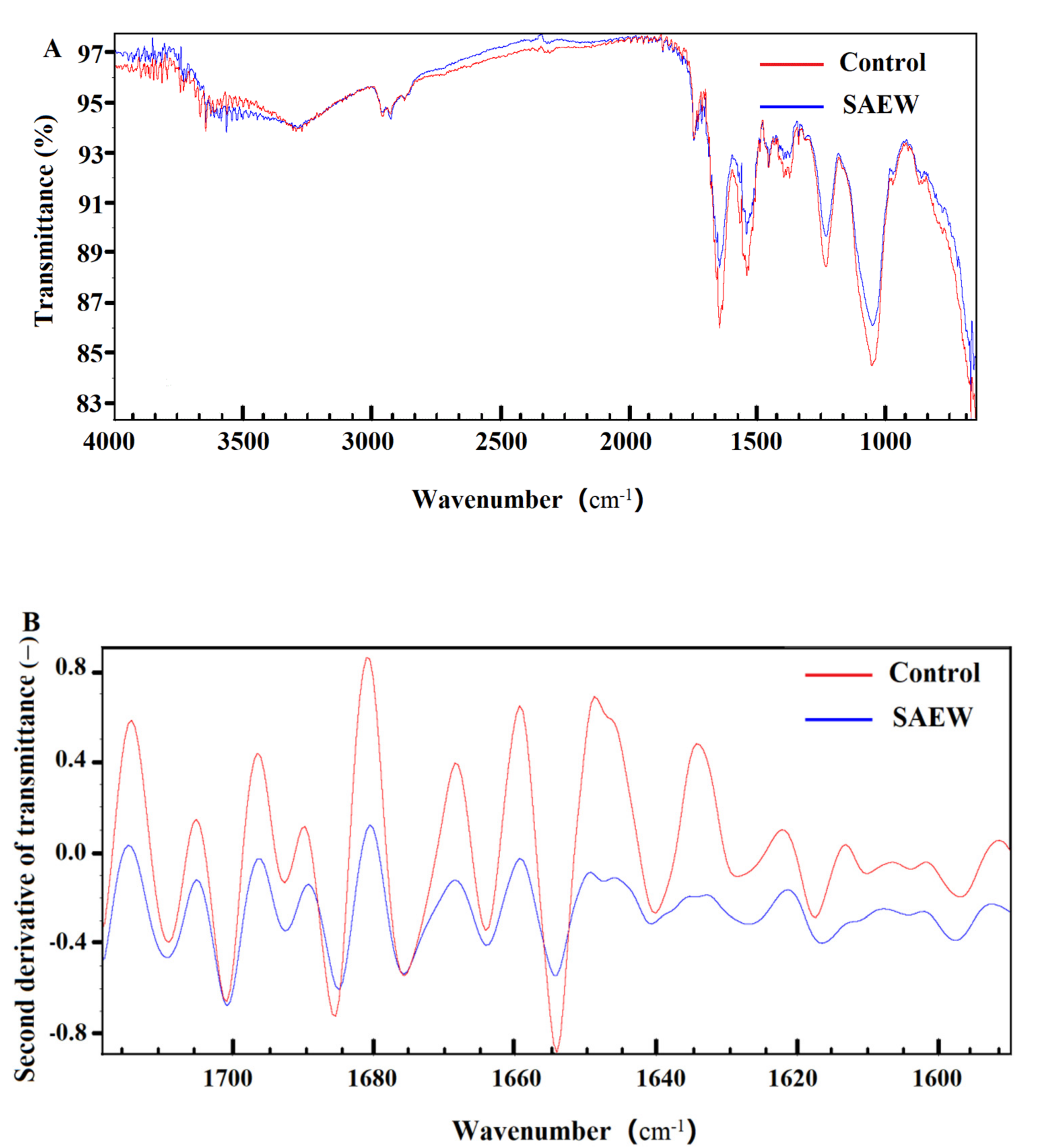
2.4. Observation of Cell Membrane by SEM and FT-IR
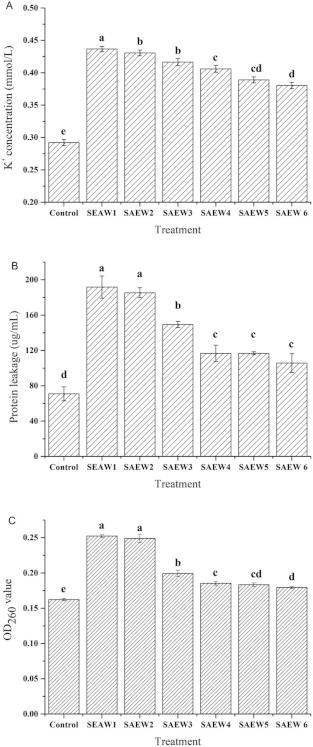
2.5. Measurement of Leakage of Bacterial Intracellular Materials
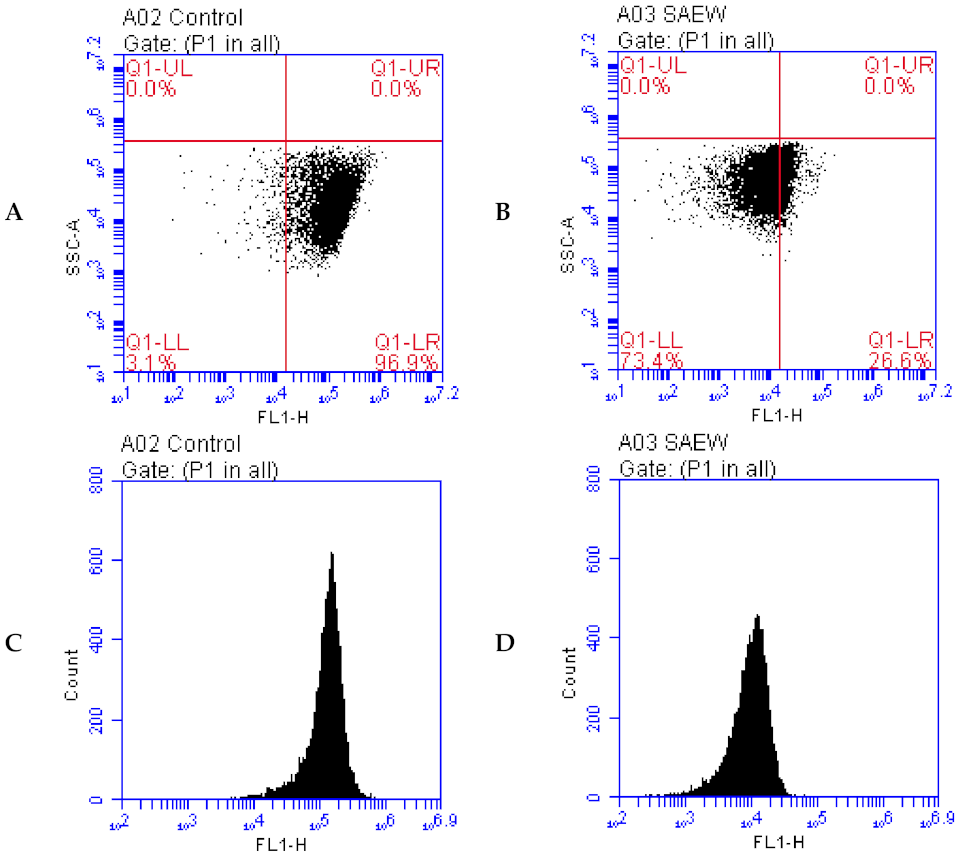
2.6. Evaluation of DNA Damage by Flow Cytometry (FCM)
2.7. ROS Analysis with Laser Scanning Confocal Microscopic (LSCM)
2.8. Measurement of Intracellular Antioxidant Enzyme Activity
2.9. Statistical Analysis
3. Results and Discussions
3.1. Bactericidal Efficacy of SAEW on Strains of L. monocytogenes
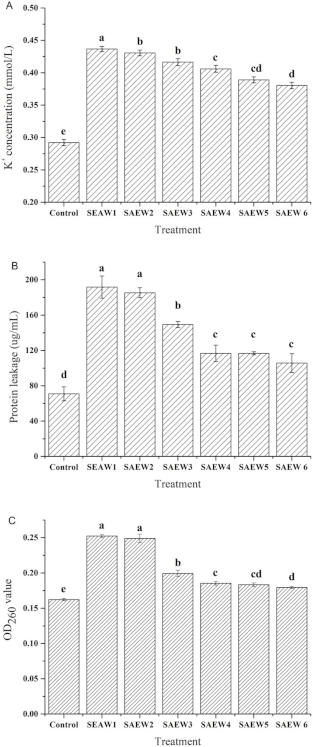
3.2. Effect of SAEW on Intracellular Material Leakages of L. monocytogenes
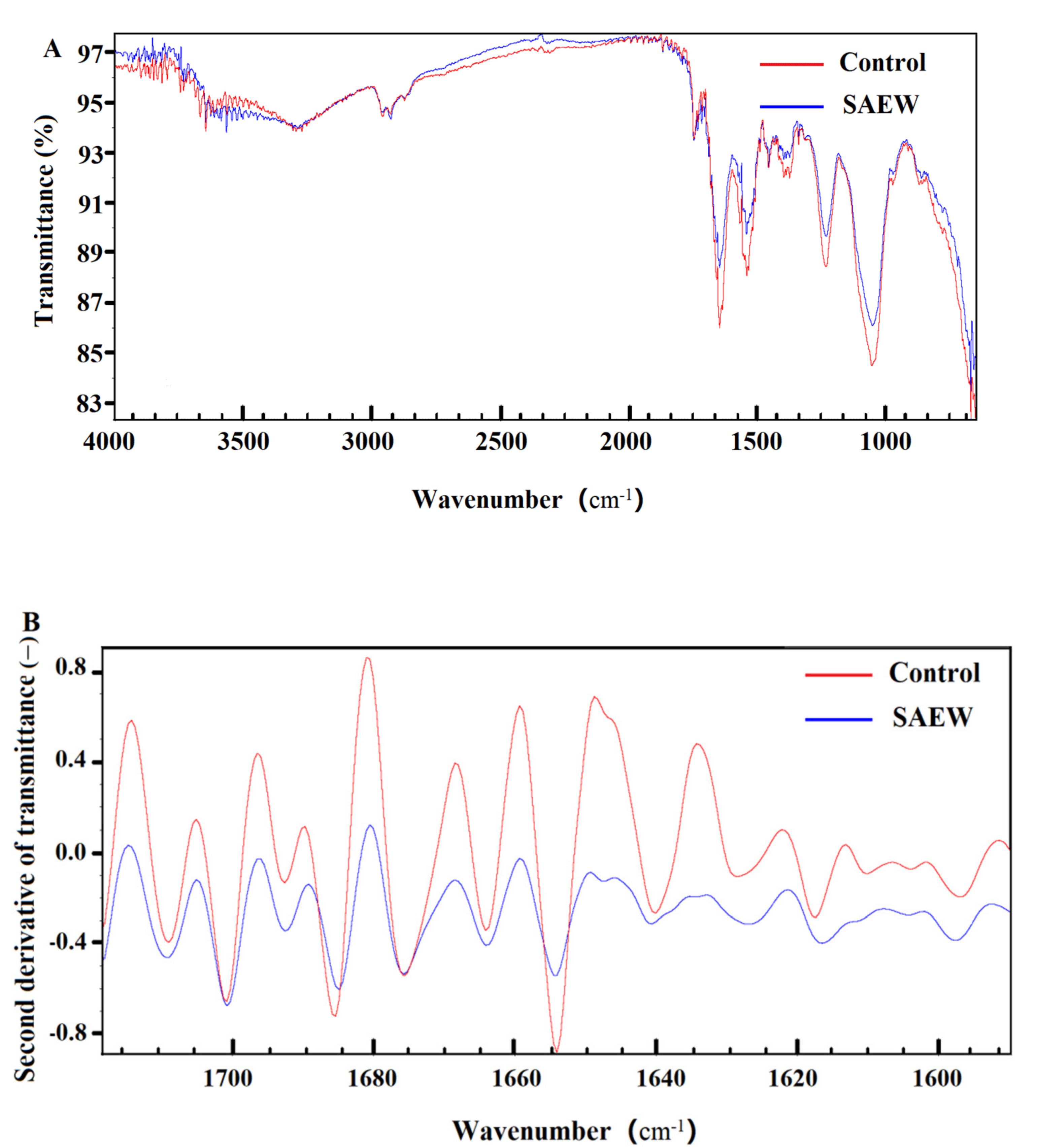
3.3. Effect of SAEW on Cell Membrane of L. monocytogenes
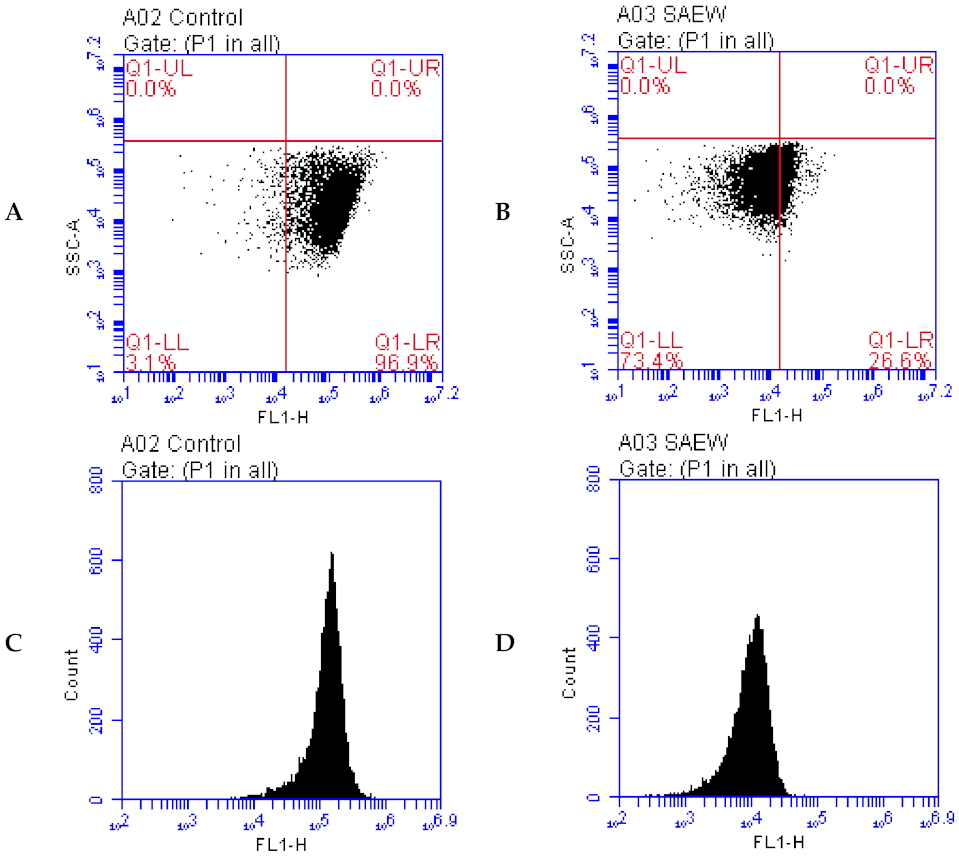
3.4. Effect of SAEW on DNA Damage of L. monocytogenes
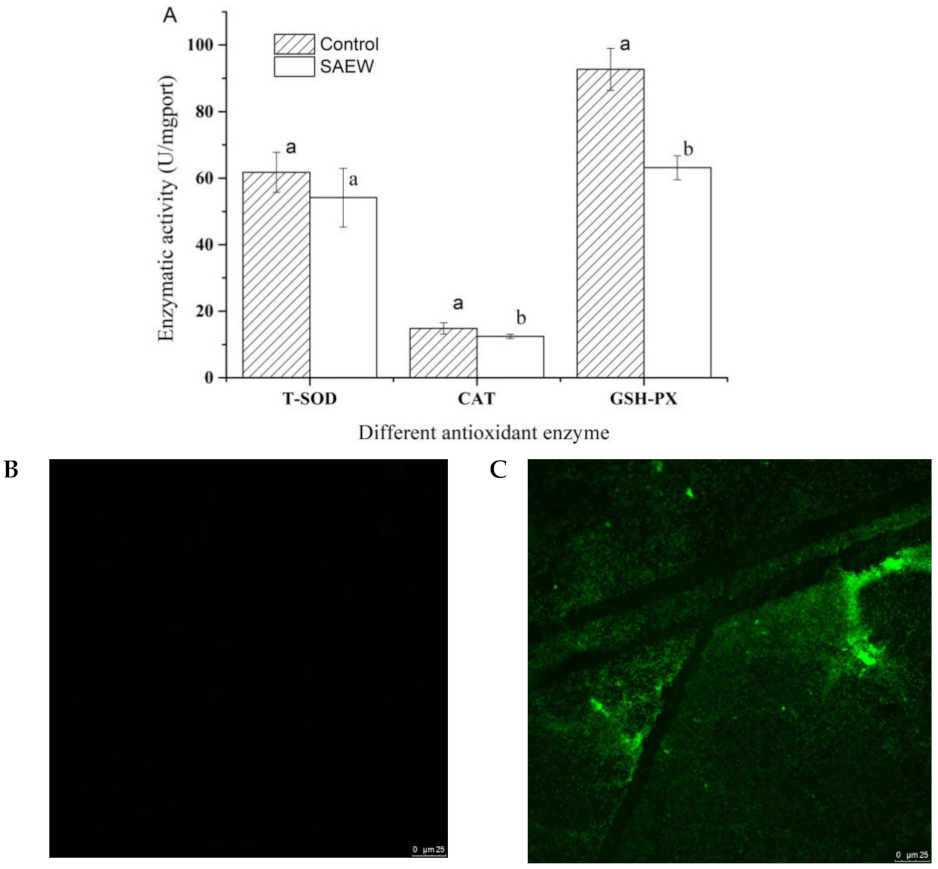
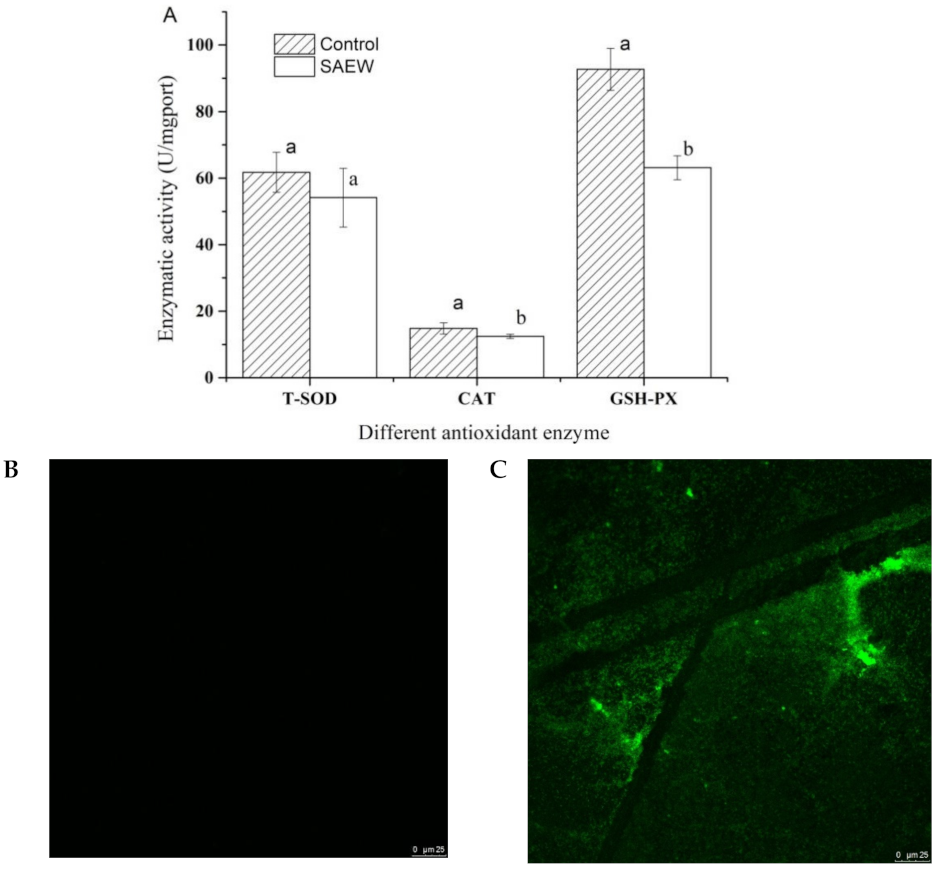
3.5. Effect of SAEW on Antioxidant Enzyme Activity and ROS of L. monocytogenes
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Jadhav, S.; Bhave, M.; Palombo, E. Methods used for the detection and subtyping of Listeria monocytogenes. J. Microbiol. Methods 2012, 88, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Sadekuzzaman, M.; Yang, S.; Mizan, F.R.; Kim, H.-S.; Ha, S.-D. Effectiveness of a phage cocktail as a biocontrol agent against L. monocytogenes biofilms. Food Control 2017, 78, 256–263. [Google Scholar] [CrossRef]
- Shen, Q.; Pandare, P.; Soni, K.A.; Nannapaneni, R.; Mahmoud, B.S.; Sharma, C.S. Influence of temperature on alkali stress adaptation in Listeria monocytogenes. Food Control 2016, 62, 74–80. [Google Scholar] [CrossRef]
- Cossart, P.; Toledo-Arana, A. Listeria monocytogenes, a unique model in infection biology: An overview. Microbes Infect. 2008, 10, 1041–1050. [Google Scholar] [CrossRef]
- Drolia, R.; Bhunia, A.K. Crossing the intestinal barrier via Listeria adhesion protein and internalin A. Trends Microbiol. 2019, 27, 408–425. [Google Scholar] [CrossRef]
- Dinner, S.; Kaltschmidt, J.; Stump-Guthier, C.; Hetjens, S.; Ishikawa, H.; Tenenbaum, T.; Schroten, H.; Schwerk, C. Mitogen-activated protein kinases are required for effective infection of human choroid plexus epithelial cells by Listeria monocytogenes. Microbes Infect. 2017, 19, 18–33. [Google Scholar] [CrossRef] [PubMed]
- Drolia, R.; Tenguria, S.; Durkes, A.C.; Turner, J.R.; Bhunia, A.K. Listeria adhesion protein induces intestinal epithelial barrier dysfunction for bacterial translocation. Cell Host Microbe 2018, 23, 470–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, S.; Akgul, A.; Karsi, A.; Abdelhamed, H.; Wills, R.W.; Lawrence, M.L. The role of Listeria monocytogenes cell wall surface anchor protein LapB in virulence, adherence, and intracellular replication. Microb. Pathog. 2016, 92, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnupf, P.; Portnoy, D.A. Listeriolysin O: A phagosome-specific lysin. Microbes Infect. 2007, 9, 1176–1187. [Google Scholar] [CrossRef]
- Thakur, M.; Asrani, R.K.; Patial, V. Chapter 6—Listeria monocytogenes: A food-borne pathogen. In Foodborne Diseases; Holban, A.M., Grumezescu, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 157–192. [Google Scholar]
- Van Troys, M.; Lambrechts, A.; David, V.; Demol, H.; Puype, M.; Pizarro-Cerda, J.; Gevaert, K.; Cossart, P.; Vandekerckhove, J. The actin propulsive machinery: The proteome of Listeria monocytogenes tails. Biochem. Biophys. Res. Commun. 2008, 375, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-R.; Hung, Y.-C.; Hsu, S.-Y.; Huang, Y.-W.; Hwang, D.-F. Application of electrolyzed water in the food industry. Food Control 2008, 19, 329–345. [Google Scholar] [CrossRef]
- Koide, S.; Takeda, J.-I.; Shi, J.; Shono, H.; Atungulu, G.G. Disinfection efficacy of slightly acidic electrolyzed water on fresh cut cabbage. Food Control 2009, 20, 294–297. [Google Scholar] [CrossRef]
- Waters, B.W.; Hung, Y.-C. Evaluation of different methods for determination of properties of chlorine-based sanitizers. Food Control 2013, 30, 41–47. [Google Scholar] [CrossRef]
- Jeon, H.R.; Kwon, M.J.; Yoon, K.S. Control of Listeria innocua Biofilms on Food Contact Surfaces with Slightly Acidic Electrolyzed Water and the Risk of Biofilm Cells Transfer to Duck Meat. J. Food Prot. 2018, 81, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Qiu, S.; Li, H.; Chen, T.; Liu, H.; Li, L. Roles of hydroxyl radicals in electrolyzed oxidizing water (EOW) for the inactivation of Escherichia coli. Int. J. Food Microbiol. 2012, 155, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Xiong, K.; Liu, H.-J.; Liu, R.; Li, L.-T. Differences in fungicidal efficiency against Aspergillus flavus for neutralized and acidic electrolyzed oxidizing waters. Int. J. Food Microbiol. 2010, 137, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Guentzel, J.L.; Lam, K.L.; Callan, M.A.; Emmons, S.A.; Dunham, V.L. Reduction of bacteria on spinach, lettuce, and surfaces in food service areas using neutral electrolyzed oxidizing water. Food Microbiol. 2008, 25, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.B.; Chen, W.M.; Xiao, X.M. The generation and inactivation mechanism of oxidation–reduction potential of electrolyzed oxidizing water. J. Food Eng. 2007, 78, 1326–1332. [Google Scholar] [CrossRef]
- Ding, T.; Xuan, X.-T.; Li, J.; Chen, S.-G.; Liu, D.-H.; Ye, X.-Q.; Shi, J.; Xue, S.J. Disinfection efficacy and mechanism of slightly acidic electrolyzed water on Staphylococcus aureus in pure culture. Food Control 2016, 60, 505–510. [Google Scholar] [CrossRef]
- Issa-Zacharia, A.; Kamitani, Y.; Morita, K.; Iwasaki, K. Sanitization potency of slightly acidic electrolyzed water against pure cultures of Escherichia coli and Staphylococcus aureus, in comparison with that of other food sanitizers. Food Control 2010, 21, 740–745. [Google Scholar] [CrossRef]
- Liao, X.; Xuan, X.; Li, J.; Suo, Y.; Liu, D.; Ye, X.; Chen, S.; Ding, T. Bactericidal action of slightly acidic electrolyzed water against Escherichia coli and Staphylococcus aureus via multiple cell targets. Food Control 2017, 79, 380–385. [Google Scholar] [CrossRef]
- Klintham, P.; Tongchitpakdee, S.; Chinsirikul, W.; Mahakarnchanakul, W. Combination of microbubbles with oxidizing sanitizers to eliminate Escherichia coli and Salmonella Typhimurium on thai leafy vegetables. Food Control 2017, 77, 260–269. [Google Scholar] [CrossRef]
- Liang, D.; Wang, Q.; Zhao, D.; Han, X.; Hao, J. Systematic application of slightly acidic electrolyzed water (SAEW) for natural microbial reduction of buckwheat sprouts. LWT 2019, 108, 14–20. [Google Scholar] [CrossRef]
- Mikš-Krajnik, M.; James Feng, L.X.; Bang, W.S.; Yuk, H.-G. Inactivation of Listeria monocytogenes and natural microbiota on raw salmon fillets using acidic electrolyzed water, ultraviolet light or/and ultrasounds. Food Control 2017, 74, 54–60. [Google Scholar] [CrossRef]
- Olaimat, A.N.; Holley, R.A. Factors influencing the microbial safety of fresh produce: A review. Food Microbiol. 2012, 32, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ding, T.; Liao, X.; Chen, S.; Ye, X.; Liu, D. Synergetic effects of ultrasound and slightly acidic electrolyzed water against Staphylococcus aureus evaluated by flow cytometry and electron microscopy. Ultrason. Sonochem. 2017, 38, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Wu, T.; Li, H.; Liu, H. Differences of Bactericidal Efficacy on Escherichia coli, Staphylococcus aureus, and Bacillus subtilis of Slightly and Strongly Acidic Electrolyzed Water. Food Bioprocess Technol. 2017, 10, 155–164. [Google Scholar] [CrossRef]
- Koide, S.; Shitanda, D.; Note, M.; Cao, W. Effects of mildly heated, slightly acidic electrolyzed water on the disinfection and physicochemical properties of sliced carrot. Food Control 2011, 22, 452–456. [Google Scholar] [CrossRef] [Green Version]
- Quan, Y.; Choi, K.-D.; Chung, D.; Shin, I.-S. Evaluation of bactericidal activity of weakly acidic electrolyzed water (WAEW) against Vibrio vulnificus and Vibrio parahaemolyticus. Int. J. Food Microbiol. 2010, 136, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Tango, C.N.; Khan, I.; Kounkeu, P.-F.N.; Momna, R.; Hussain, M.; Oh, D.-H. Slightly acidic electrolyzed water combined with chemical and physical treatments to decontaminate bacteria on fresh fruits. Food Microbiol. 2017, 67, 97–105. [Google Scholar] [CrossRef]
- Izumi, H.; Inoue, A. Viability of sublethally injured coliform bacteria on fresh-cut cabbage stored in high CO2 at-mospheres following rinsing with electrolyzed water. Int. J. Food Microbiol. 2018, 266, 207–212. [Google Scholar] [CrossRef]
- Lan, L.; Zhang, R.; Zhang, X.; Shi, H. Sublethal injury and recovery of Listeria monocytogenes and Escherichia coli O157:H7 after exposure to slightly acidic electrolyzed water. Food Control 2019, 106, 106746. [Google Scholar] [CrossRef]
- Xuan, X.-T.; Ding, T.; Li, J.; Ahn, J.-H.; Zhao, Y.; Chen, S.-G.; Ye, X.-Q.; Liu, D.-H. Estimation of growth parameters of Listeria monocytogenes after sublethal heat and slightly acidic electrolyzed water (SAEW) treatment. Food Control 2017, 71, 17–25. [Google Scholar] [CrossRef]
- Afari, G.K.; Liu, H.; Hung, Y.-C. The effect of produce washing using electrolyzed water on the induction of the viable but non-culturable (VBNC) state in Listeria monocytogenes and Escherichia coli O157:H7. LWT 2019, 110, 275–282. [Google Scholar] [CrossRef]
- Han, D.; Hung, Y.-C.; Wang, L. Evaluation of the antimicrobial efficacy of neutral electrolyzed water on pork products and the formation of viable but nonculturable (VBNC) pathogens. Food Microbiol. 2018, 73, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Churklam, W.; Chaturongakul, S.; Ngamwongsatit, B.; Aunpad, R. The mechanisms of action of carvacrol and its synergism with nisin against Listeria monocytogenes on sliced bologna sausage. Food Control 2020, 108, 106864. [Google Scholar] [CrossRef]
- Xiang, Q.; Wang, W.; Zhao, D.; Niu, L.; Li, K.; Bai, Y. Synergistic inactivation of Escherichia coli O157:H7 by plasma-activated water and mild heat. Food Control 2019, 106, 106741. [Google Scholar] [CrossRef]
- Yonggang, W.; Guangrui, Y.; Xueqing, M.; Minggang, W.; Feifan, L.; Fan, W.; Jianzhong, M.; Jixiang, C. Antibacterial mechanisms of endophytic fungi Alternaria alternata of ethyl acetate extract on Escherichia coli. Chin. Tradit. Herb. Drugs 2018, 49, 374–381. [Google Scholar]
- Campana, R.; Baffone, W. Carvacrol efficacy in reducing microbial biofilms on stainless steel and in limiting re-growth of injured cells. Food Control 2018, 90, 10–17. [Google Scholar] [CrossRef]
- Hu, M.; Du, J.; Du, L.; Luo, Q.; Xiong, J. Anti-fatigue activity of purified anthocyanins prepared from purple passion fruit (P. edulis Sim) epicarp in mice. J. Funct. Foods 2020, 65, 103725. [Google Scholar] [CrossRef]
- Sun, H.; Luo, M.; Zhou, X.; Zhou, Q.; Sun, Y.; Ge, W.; Wei, B.; Cheng, S.; Ji, S. Exogenous glycine betaine treatment alleviates low temperature-induced pericarp browning of ‘Nanguo’ pears by regulating antioxidant enzymes and proline metabolism. Food Chem. 2020, 306, 125626. [Google Scholar] [CrossRef] [PubMed]
- Gansukh, E.; Mya, K.K.; Jung, M.; Keum, Y.-S.; Kim, D.H.; Saini, R.K. Lutein derived from marigold (Tagetes erecta) petals triggers ROS generation and activates Bax and caspase-3 mediated apoptosis of human cervical carcinoma (HeLa) cells. Food Chem. Toxicol. 2019, 127, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Selvi, S.K.; Vinoth, A.; Varadharajan, T.; Weng, C.F.; Padma, V.V. Neferine augments therapeutic efficacy of cisplatin through ROS- mediated non-canonical autophagy in human lung adenocarcinoma (A549 cells). Food Chem. Toxicol. 2017, 103, 28–40. [Google Scholar] [CrossRef] [PubMed]



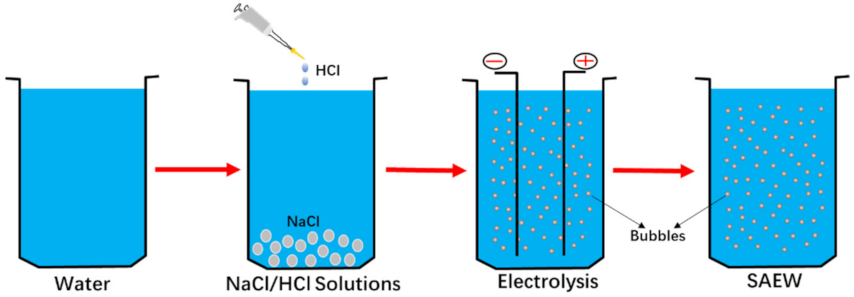{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SAEW Solutions | pH Value | ACC (mg/L) | Treatment Time (s) | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 30 | 60 | 90 | 120 | 150 | |||
| SAEW 1 | 5.73 ± 0.05 A | 24.81 ± 0.18 A | 7.34 ± 0.04 Aa | 0 Ab | 0 Ab | 0 Ab | 0 Ab | 0 Ab |
| SAEW 2 | 5.75 ± 0.04 A | 12.35 ± 0.22 B | 7.28 ± 0.07 Aa | 0 Ab | 0 Ab | 0 Ab | 0 Ab | 0 Ab |
| SAEW 3 | 5.76 ± 0.03 A | 6.03 ± 0.13 C | 7.45 ± 0.13 Aa | 5.34 ± 0.04 Bbc | 5.50 ± 0.20 Bb | 5.12 ± 0.05 Bc | 4.09 ± 0.09 Bd | 0 Ae |
| SAEW 4 | 5.73 ± 0.05 A | 4.25 ± 0.38 D | 7.34 ± 0.04 Aa | 6.88 ± 0.03 Cb | 6.95 ± 0.07 Cb | 6.78 ± 0.10 Cbc | 6.73 ± 0.06 Cbc | 6.60 ± 0.15 Cc |
| SAEW 5 | 5.80 ± 0.06 A | 3.54 ± 0.24 E | 7.37 ± 0.02 Aa | 7.02 ± 0.05 Db | 6.94 ± 0.03 Cbc | 6.85 ± 0.04 Cbc | 6.81 ± 0.10 Cc | 6.57 ± 0.11 Cd |
| SAEW 6 | 5.81 ± 0.05 A | 2.39 ± 0.16 F | 7.39 ± 0.02 Aa | 7.11 ± 0.03 Eb | 6.96 ± 0.06 Cbc | 6.80 ± 0.10 Cc | 6.82 ± 0.05 Cc | 6.62 ± 0.10 Cd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Liang, D.; Huang, J.; Cui, C.; Rao, H.; Zhao, D.; Hao, J. The Bactericidal Efficacy and the Mechanism of Action of Slightly Acidic Electrolyzed Water on Listeria monocytogenes’ Survival. Foods 2021, 10, 2671. https://doi.org/10.3390/foods10112671
Li H, Liang D, Huang J, Cui C, Rao H, Zhao D, Hao J. The Bactericidal Efficacy and the Mechanism of Action of Slightly Acidic Electrolyzed Water on Listeria monocytogenes’ Survival. Foods. 2021; 10(11):2671. https://doi.org/10.3390/foods10112671
Chicago/Turabian StyleLi, Huiying, Duo Liang, Jin Huang, Chaojing Cui, Huan Rao, Dandan Zhao, and Jianxiong Hao. 2021. "The Bactericidal Efficacy and the Mechanism of Action of Slightly Acidic Electrolyzed Water on Listeria monocytogenes’ Survival" Foods 10, no. 11: 2671. https://doi.org/10.3390/foods10112671
APA StyleLi, H., Liang, D., Huang, J., Cui, C., Rao, H., Zhao, D., & Hao, J. (2021). The Bactericidal Efficacy and the Mechanism of Action of Slightly Acidic Electrolyzed Water on Listeria monocytogenes’ Survival. Foods, 10(11), 2671. https://doi.org/10.3390/foods10112671