
Effects of Freeze–Thaw Cycles on Water Migration, Microstructure and Protein Oxidation in Cuttlefish



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
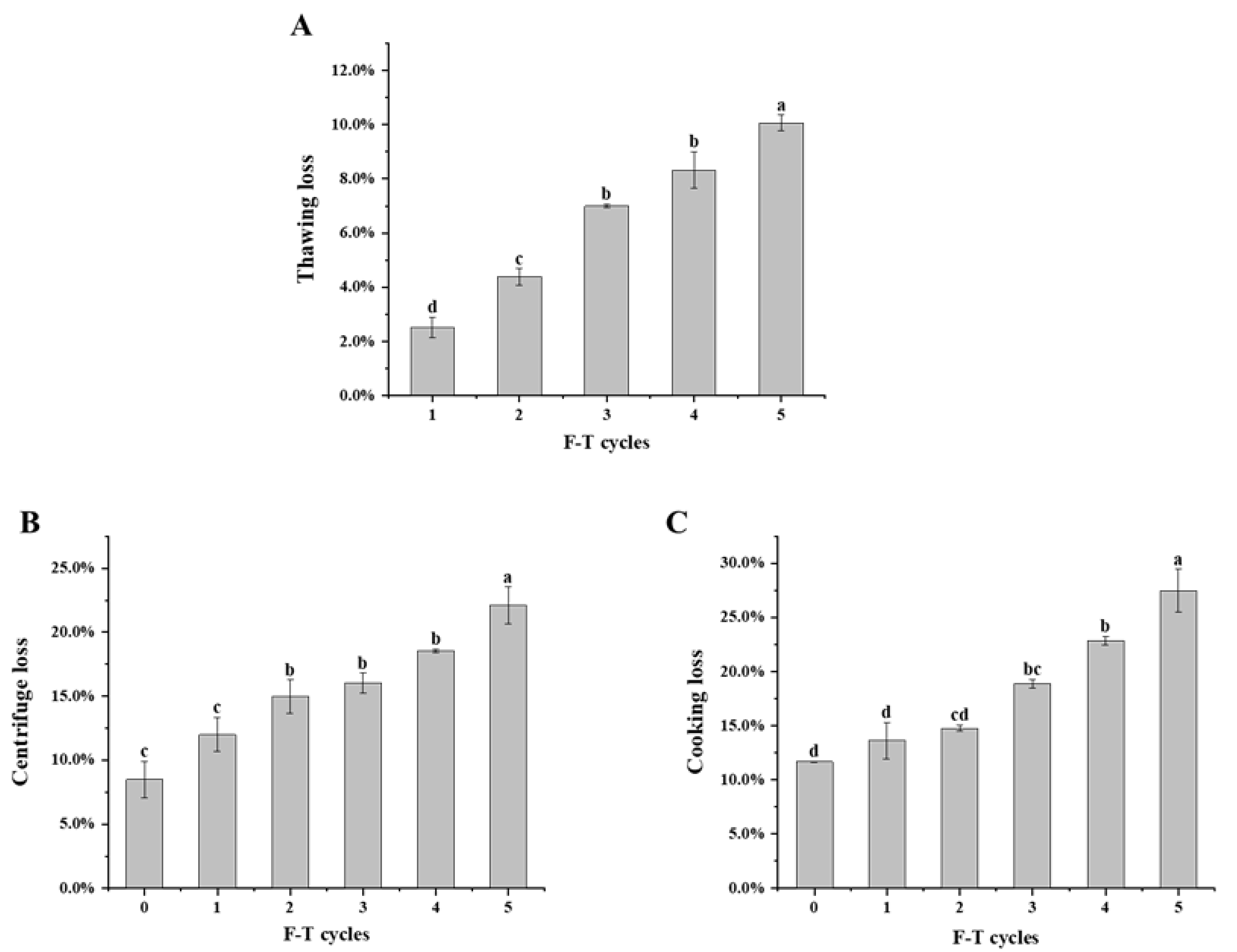
2.2. Thawing Loss
2.3. Cooking Loss
2.4. Centrifugal Loss
2.5. Low-Field Nuclear Magnetic Resonance(LF-NMR) and Proton Magnetic Resonance Imaging (MRI)
2.6. Color Properties Analysis
2.7. pH
2.8. Texture Profile Analysis (TPA)
2.9. Myofibrillar Proteins (MP) Extraction
2.10. Total Sulfhydryl (SH) Group Content and Ca2+-ATPase activity
2.11. Free Amino Acids (FAA)
2.12. Fourier Infrared Spectroscopy
2.13. Intrinsic Fluorescence Spectra
2.14. Total Volatile Base Nitrogen (TVB-N)
2.15. Determination of Microstructure by SEM
2.16. Statistical Analysis
3. Results and Discussion
3.1. Water Holding Capacity
3.2. Moisture Distribution
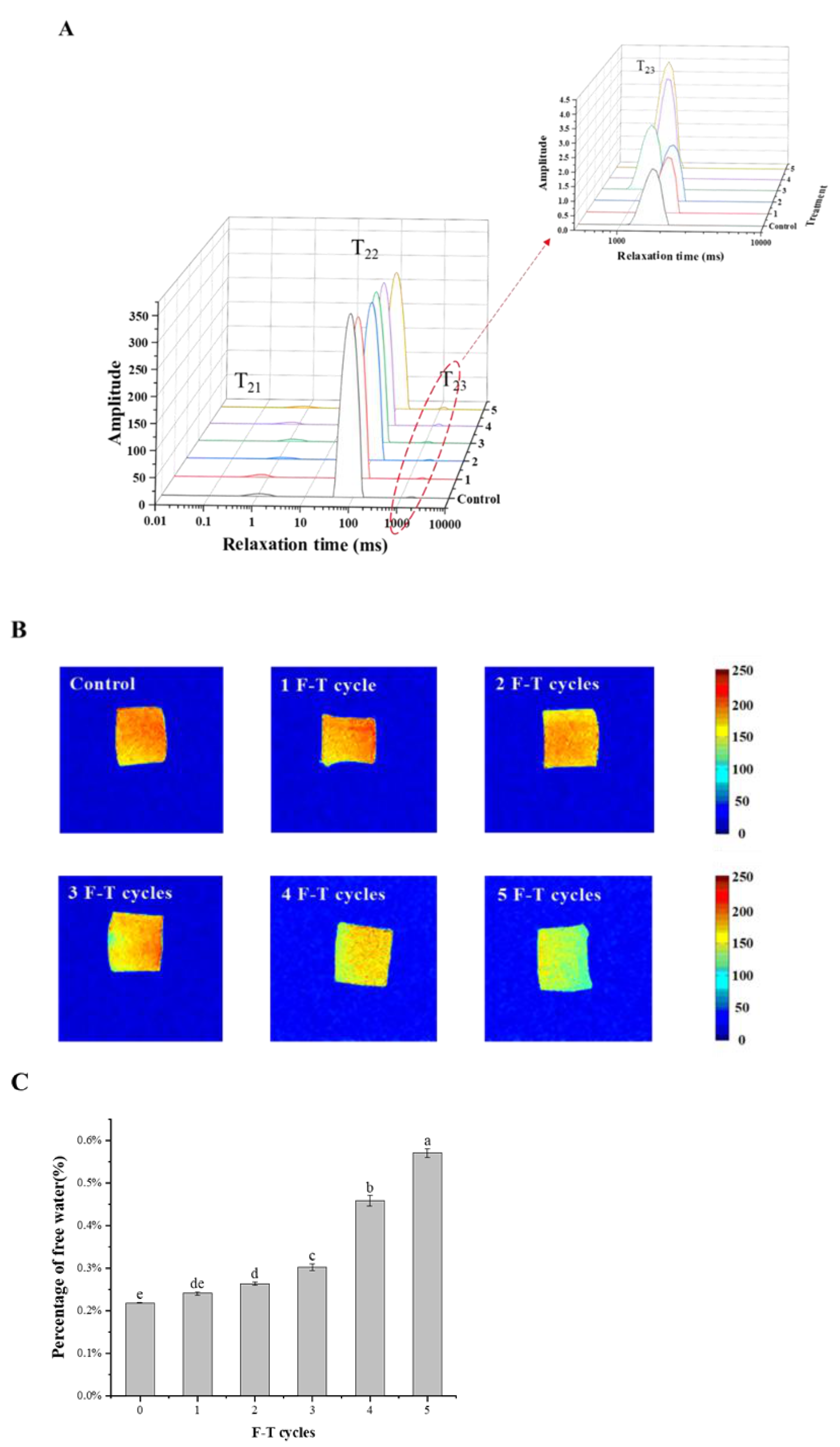
3.3. Colour
3.4. pH
3.5. TPA
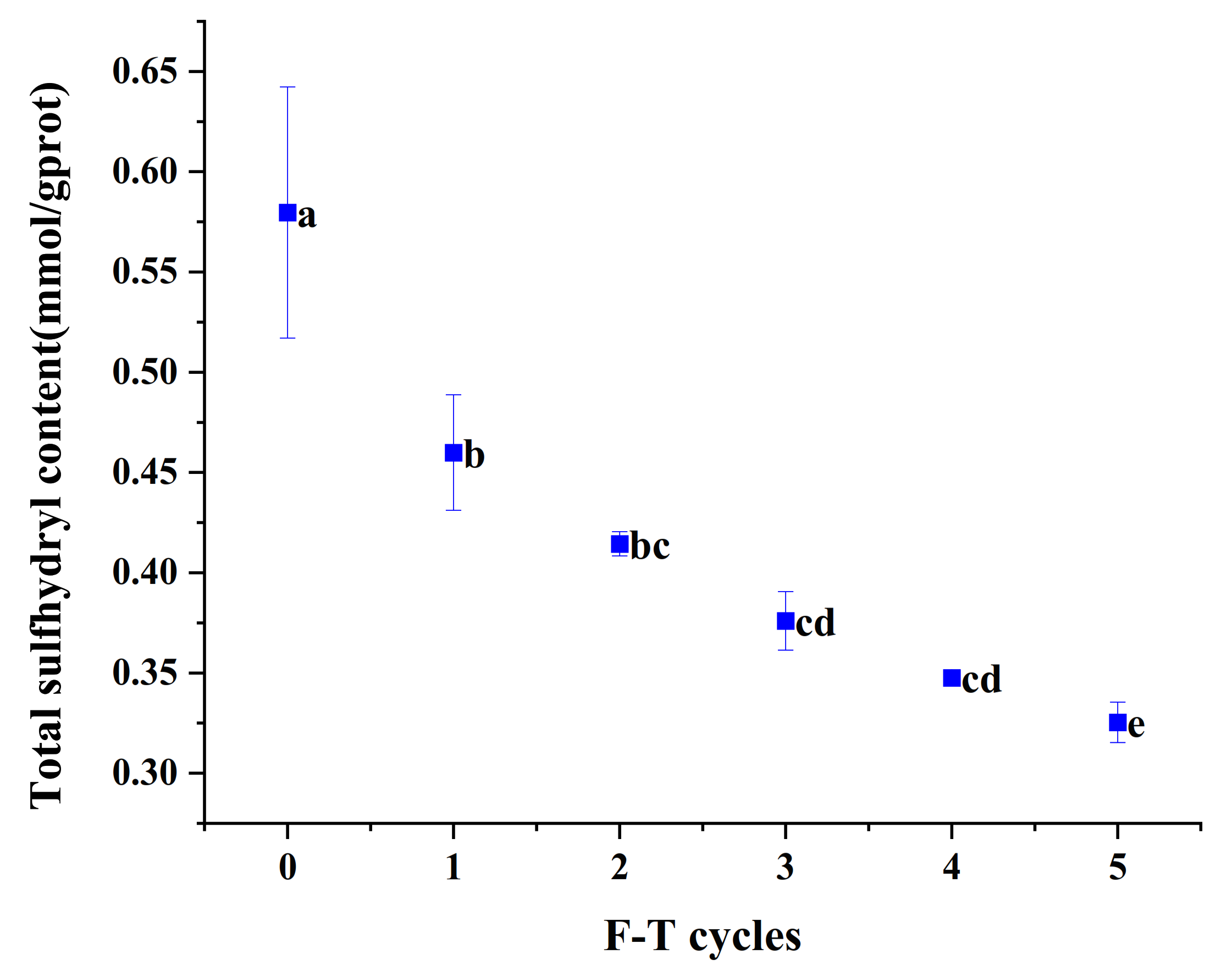
3.6. Total Sulfhydryl Content
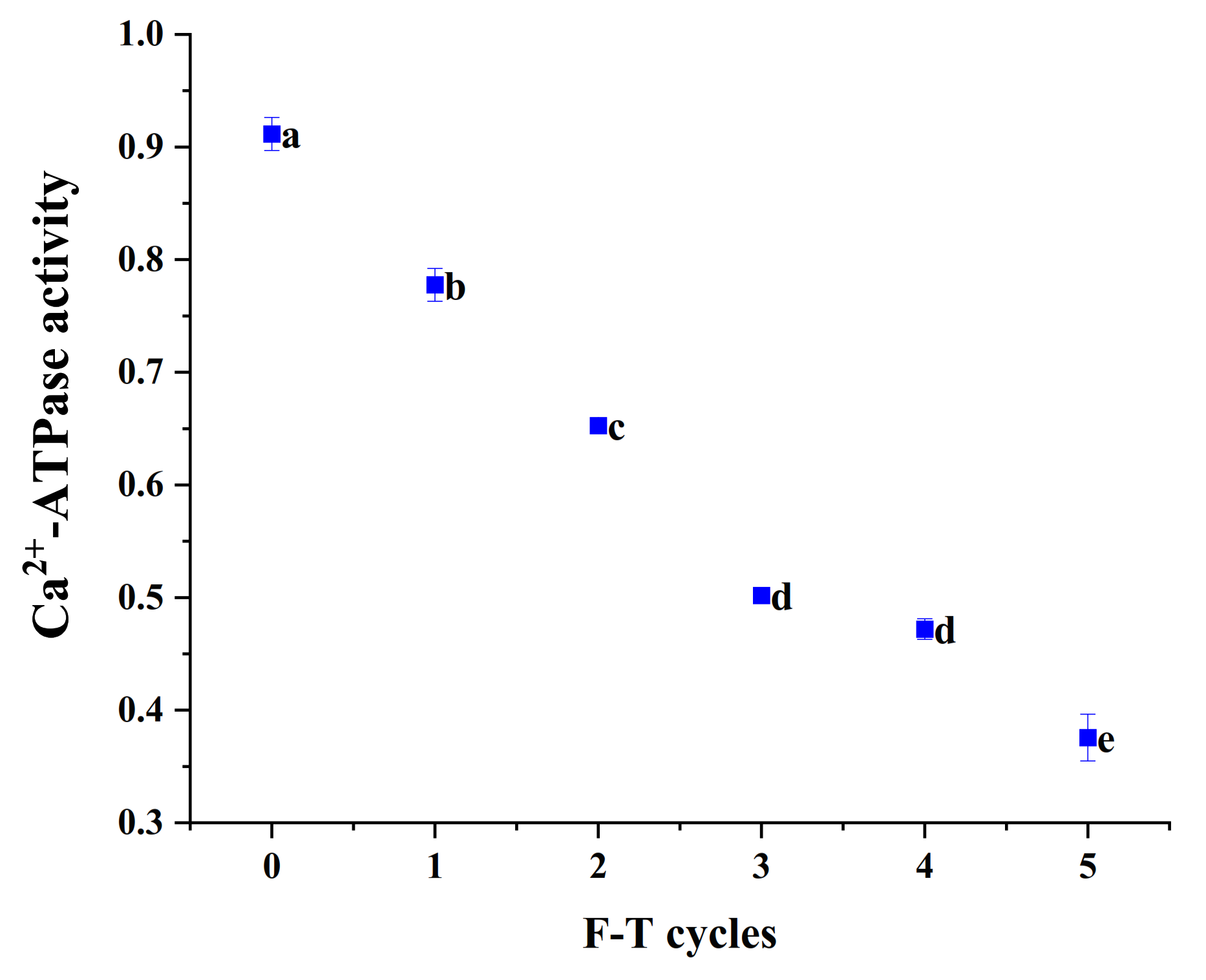
3.7. Ca2+-ATPase Activity
3.8. FAA
3.9. Protein Secondary Structure
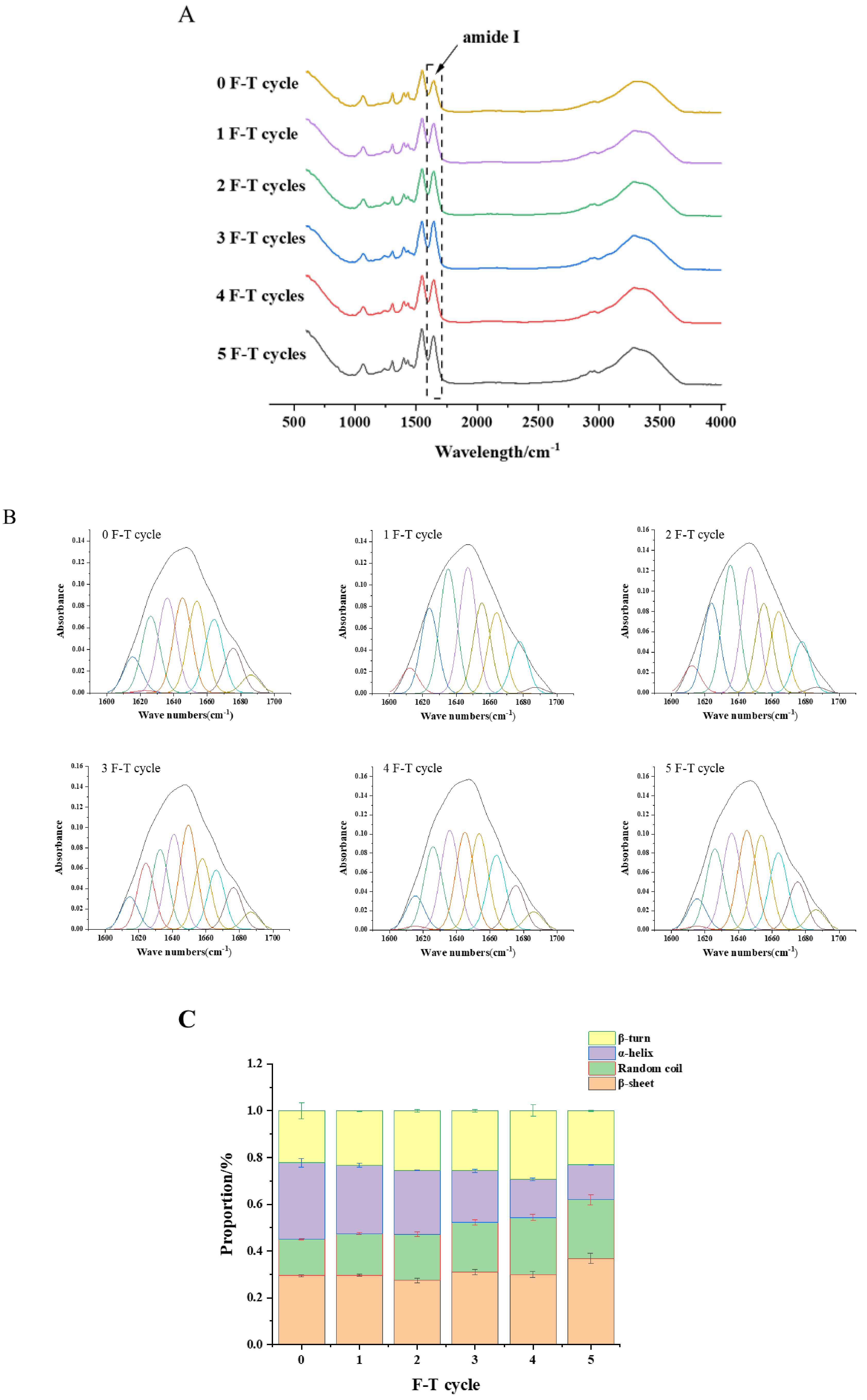
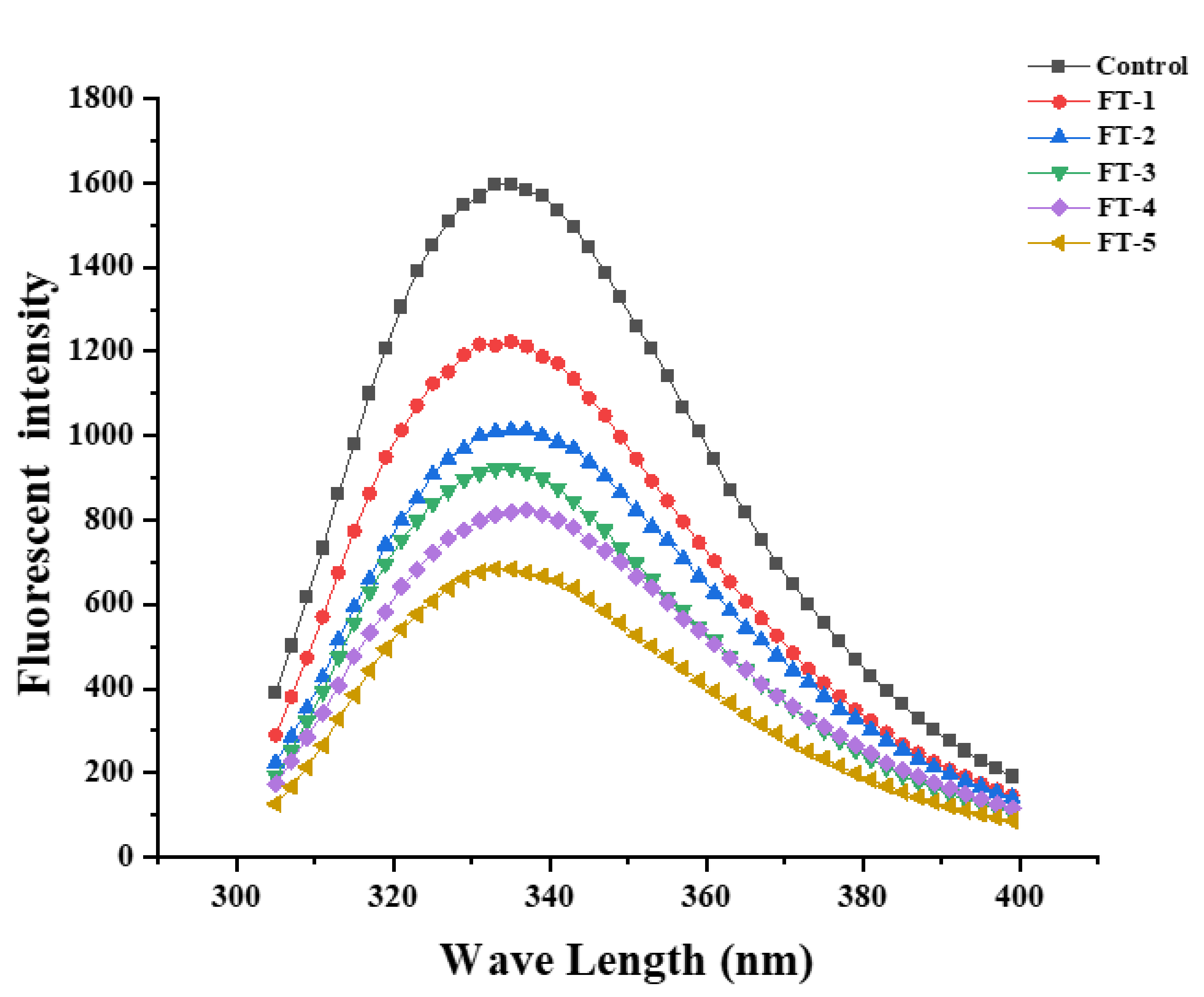
3.10. Intrinsic Fluorescence Spectroscopy Analysis
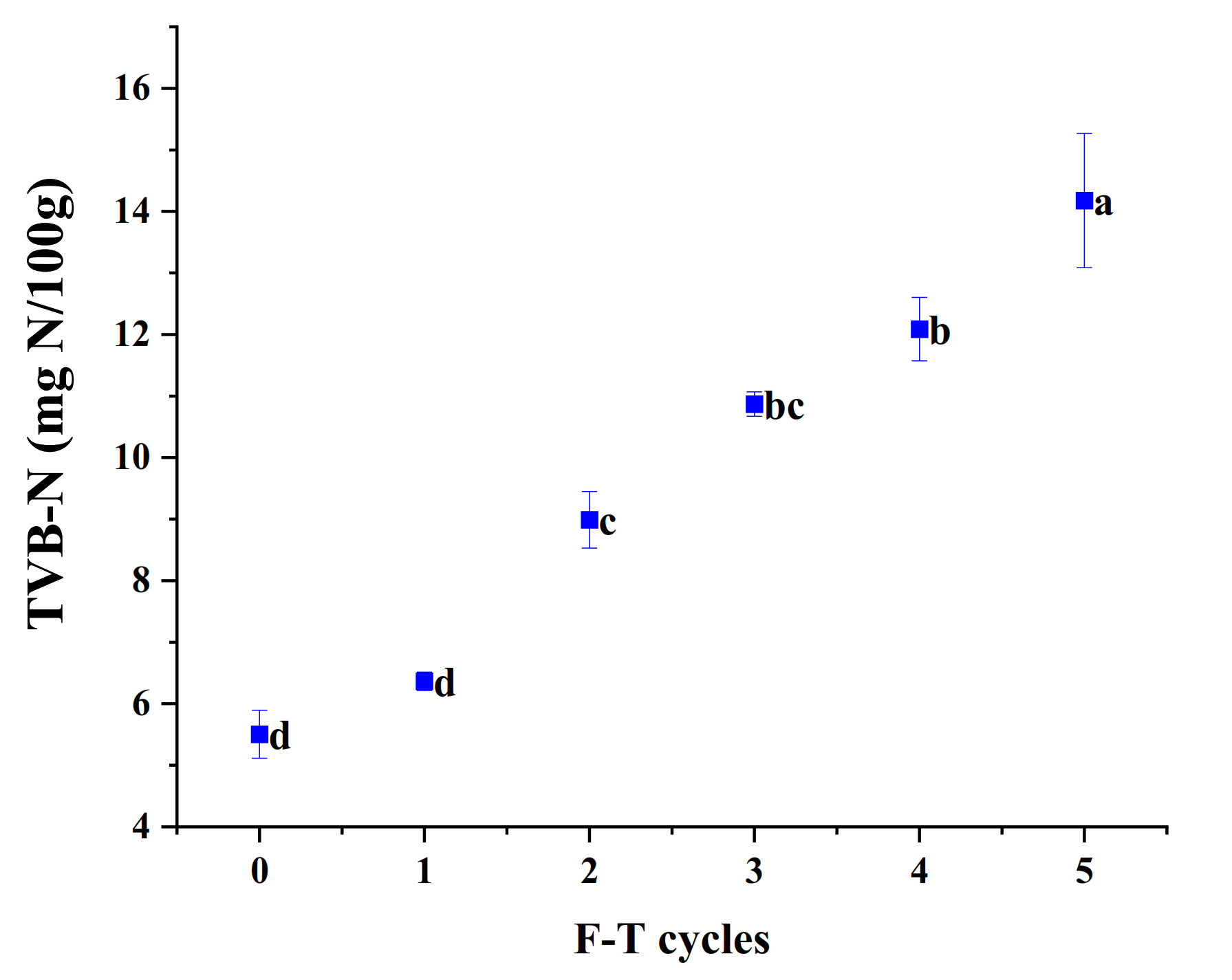
3.11. TVB-N
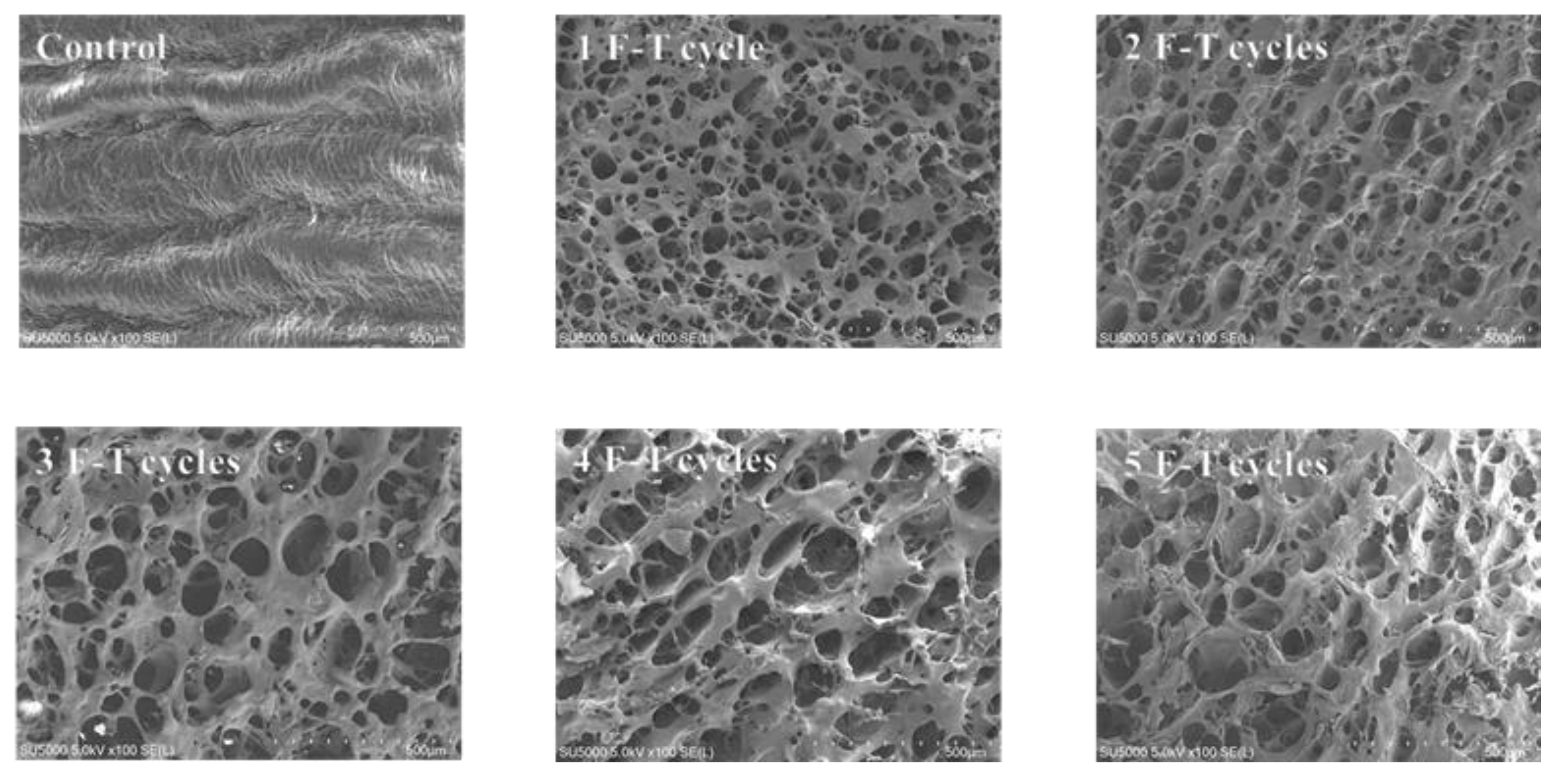
3.12. Microstructure
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bouletis, A.D.; Arvanitoyannis, I.S.; Hadjichristodoulou, C.; Neofitou, C.; Parlapani, F.F.; Gkagtzis, D.C. Quality changes of cuttlefish stored under various atmosphere modifications and vacuum packaging. J. Sci. Food Agric. 2016, 96, 2882–2888. [Google Scholar] [CrossRef] [PubMed]
- Vaz-Pires, P.; Seixas, P.; Mota, M.; Lapa-Guimarães, J.; Pickova, J.; Lindo, A.; Silva, T. Sensory, microbiological, physical and chemical properties of cuttlefish (Sepia officinalis) and broadtail shortfin squid (Illex coindetii) stored in ice. LWT Food Sci. Technol. 2008, 41, 1655–1664. [Google Scholar] [CrossRef] [Green Version]
- Badiani, A.; Bonaldo, A.; Testi, S.; Rotolo, M.; Serratore, P.; Giulini, G.; Pagliuca, G.; Gatta, P.P. Good handling practices of the catch: The effect of early icing on the freshness quality of cuttlefish (Sepia officinalis L.). Food Control 2013, 32, 327–333. [Google Scholar] [CrossRef]
- Vaz-Pires, P.; Seixas, P. Development of new quality index method (QIM) schemes for cuttlefish (Sepia officinalis) and broadtail shortfin squid (Illex coindetii). Food Control 2006, 17, 942–949. [Google Scholar] [CrossRef]
- Leygonie, C.; Britz, T.J.; Hoffman, L.C. Impact of freezing and thawing on the quality of meat: Review. Meat Sci. 2012, 91, 93–98. [Google Scholar] [CrossRef]
- Wang, B.; Du, X.; Kong, B.; Liu, Q.; Li, F.; Pan, N.; Xia, X.; Zhang, D. Effect of ultrasound thawing, vacuum thawing, and microwave thawing on gelling properties of protein from porcine longissimus dorsi. Ultrason. Sonochem. 2020, 64, 104860. [Google Scholar] [CrossRef]
- Alonso, V.; Muela, E.; Tenas, J.; Calanche, J.B.; Roncalés, P.; Beltrán, J.A. Changes in physicochemical properties and fatty acid composition of pork following long-term frozen storage. Eur. Food Res. Technol. 2016, 12, 2119–2127. [Google Scholar] [CrossRef]
- Yang, H.; Meng, P.P.; Wang, R.; Li, P.R.; Li, P.; Wang, C.L.; Ma, L.Z. Effect of oxidized myofibrils protein subjected to mutiple freeze-thaw cycles on N-nitrosamine formation in in vitro model system (Conference Paper). Adv. Mater. Res. 2012, 550, 1590–1594. [Google Scholar] [CrossRef]
- Cheng, S.; Wang, X.; Li, R.; Yang, H.; Wang, H.; Wang, H.; Tan, M. Influence of multiple freeze-thaw cycles on quality characteristics of beef semimembranous muscle: With emphasis on water status and distribution by LF-NMR and MRI. Meat Sci. 2019, 147, 44–52. [Google Scholar] [CrossRef]
- Paarup, T.; Sanchez, J.A.; Peláez, C.; Moral, A. Sensory, chemical and bacteriological changes in vacuum-packed pressurised squid mantle (Todaropsis eblanae) stored at 4 °C. Int. J. Food Microbiol. 2002, 74, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; Li, C.; Chen, Y.; Gao, F.; Xu, X.; Zhou, G. Changes in meat quality of ovine longissimus dorsi muscle in response to repeated freeze and thaw. Meat Sci. 2012, 92, 619–926. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Xie, J. The Effect of Multiple Freeze–Thaw Cycles on the Microstructure and Quality of Trachurus murphyi. Foods 2021, 10, 1350. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Wang, J.; Li, P.; Xie, J. Storage time prediction of glazed frozen squids during frozen storage at different temperatures based on neural network. Int. J. Food Prop. 2020, 23, 1663–1677. [Google Scholar] [CrossRef]
- Lan, W.; Sun, Y.; Chen, M.; Li, H.; Ren, Z.; Lu, Z.; Xie, J. Effects of pectin combined with plant essential oils on water migration, myofibrillar proteins and muscle tissue enzyme activity of vacuum packaged large yellow croaker (Pseudosciaena crocea) during ice storage. Food Packag. Shelf Life 2021, 30, 100699. [Google Scholar] [CrossRef]
- Tan, M.; Ye, J.; Chu, Y.; Xie, J. The effects of ice crystal on water properties and protein stability of large yellow croaker (Pseudosciaena crocea). Int. J. Refrig. 2021, 130, 242–252. [Google Scholar] [CrossRef]
- Lan, W.; Hu, X.; Sun, X.; Zhang, X.; Xie, J. Effect of the number of freeze-thaw cycles number on the quality of Pacific white shrimp (Litopenaeus vannamei): An emphasis on moisture migration and microstructure by LF-NMR and SEM. Aquac. Fish. 2020, 5, 193–200. [Google Scholar] [CrossRef]
- Lv, Y.; Chu, Y.; Zhou, P.; Mei, J.; Xie, J. Effects of Different Freezing Methods on Water Distribution, Microstructure and Protein Properties of Cuttlefish during the Frozen Storage. Appl. Sci. 2021, 11, 6866. [Google Scholar] [CrossRef]
- Song, Y.; Liu, L.; Shen, H.; You, J.; Luo, Y. Effect of sodium alginate-based edible coating containing different anti-oxidants on quality and shelf life of refrigerated bream (Megalobrama amblycephala). Food Control 2011, 22, 608–615. [Google Scholar] [CrossRef]
- Tan, M.; Lin, Z.; Zu, Y.; Zhu, B.; Cheng, S. Effect of multiple freeze-thaw cycles on the quality of instant sea cucumber: Emphatically on water status of by LF-NMR and MRI. Food Res. Int. 2018, 109, 65–71. [Google Scholar] [CrossRef]
- Wang, X.-Y.; Xie, J. Evaluation of water dynamics and protein changes in bigeye tuna (Thunnus obesus) during cold storage. LWT 2019, 108, 289–296. [Google Scholar] [CrossRef]
- Wang, J.; Yu, W.; Xie, J. Effect of Glazing with Different Materials on the Quality of Tuna during Frozen Storage. Foods 2020, 9, 231. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.; Cheng, H.; Yu, H.; Mei, J.; Xie, J. Quality enhancement of large yellow croaker (Pseudosciaena crocea) during frozen (−18 °C) storage by spiral freezing. CyTA J. Food 2021, 19, 710–720. [Google Scholar] [CrossRef]
- Liu, W.; Mei, J.; Xie, J. Effect of locust bean gum-sodium alginate coatings incorporated with daphnetin emulsions on the quality of Scophthalmus maximus at refrigerated condition. Int. J. Biol. Macromol. 2021, 170, 129–139. [Google Scholar] [CrossRef]
- Zhang, M.; Li, F.; Diao, X.; Kong, B.; Xia, X. Moisture migration, microstructure damage and protein structure changes in porcine longissimus muscle as influenced by multiple freeze-thaw cycles. Meat Sci. 2017, 133, 10–18. [Google Scholar] [CrossRef]
- Wang, Z.; He, Z.; Zhang, D.; Chen, X.; Li, H. Effect of multiple freeze-thaw cycles on protein and lipid oxidation in rabbit meat. Int. J. Food Sci. Technol. 2021, 56, 3004–3015. [Google Scholar] [CrossRef]
- Nian, L.; Cao, A.; Cai, L.; Ji, H.; Liu, S. Effect of vacuum impregnation of red sea bream (Pagrosomus major) with herring AFP combined with CS@Fe3O4 nanoparticles during freeze-thaw cycles. Food Chem. 2019, 291, 139–148. [Google Scholar] [CrossRef]
- Qiu, H.; Guo, X.; Deng, X.; Guo, X.; Mao, X.; Xu, C.; Zhang, J. The influence of endogenous cathepsin in different subcellular fractions on the quality deterioration of Northern pike (Esox lucius) fillets during refrigeration and partial freezing storage. Food Sci. Biotechnol. 2020, 10, 1331–1341. [Google Scholar] [CrossRef]
- Wang, Y.; Miyazaki, R.; Saitou, S.; Hirasaka, K.; Takeshita, S.; Tachibana, K.; Taniyama, S. The effect of ice crystals formations on the flesh quality of frozen horse mackerel (Trachurus japonicus). J. Texture Stud. 2018, 5, 485–491. [Google Scholar] [CrossRef]
- Cai, L.; Nian, L.; Cao, A.; Zhang, Y.; Li, X. Effect of Carboxymethyl Chitosan Magnetic Nanoparticles Plus Herring Antifreeze Protein on Conformation and Oxidation of Myofibrillar Protein from Red Sea Bream (Pagrosomus major) after Freeze-Thaw Treatment. Food Bioprocess Technol. 2020, 2, 355–366. [Google Scholar] [CrossRef]
- Benjakul, S.; Bauer, F. Physicochemical and enzymatic changes of cod muscle proteins subjected to different freeze-thaw cycles. J. Sci. Food Agric. 2000, 8, 1143–1150. [Google Scholar] [CrossRef]
- Kong, C.; Wang, H.; Li, D.; Zhang, Y.; Pan, J.; Zhu, B.; Luo, Y. Quality changes and predictive models of radial basis function neural networks for brined common carp (Cyprinus carpio) fillets during frozen storage. Food Chem. 2016, 201, 327–333. [Google Scholar] [CrossRef]
- Li, P.; Mei, J.; Xie, J. Chitosan-sodium alginate bioactive coatings containing ε-polylysine combined with high CO2 modified atmosphere packaging inhibit myofibril oxidation and degradation of farmed pufferfish (Takifugu obscurus) during cold storage. LWT 2021, 140, 110652. [Google Scholar] [CrossRef]
- Özden, Ö. Changes in amino acid and fatty acid composition during shelf-life of marinated fish. J. Sci. Food Agric. 2005, 12, 2015–2020. [Google Scholar] [CrossRef]
- Baranenko, V.K.D.; Broyko, Y. Effect of cold treatment on the amino acid composition of veal. Agron. Res. 2014, 12, 705–716. [Google Scholar]
- Gokoglu, N.; Topuz, O.K.; Yerlikaya, P.; Yatmaz, H.A.; Ucak, I. Effects of Freezing and Frozen Storage on Protein Functionality and Texture of Some Cephalopod Muscles. J. Aquat. Food Prod. Technol. 2018, 2, 211–218. [Google Scholar] [CrossRef]
- Shi, J.; Wang, Q.; Pan, D.; Liu, T.; Jiang, M. Characterization of interactions of simvastatin, pravastatin, fluvastatin, and pitavastatin with bovine serum albumin: Multiple spectroscopic and molecular docking. J. Biomol. Struct. Dyn. 2017, 7, 1529–1546. [Google Scholar] [CrossRef]
- Cai, L.; Nian, L.; Zhao, G.; Zhang, Y.; Sha, L.; Li, J. Effect of Herring Antifreeze Protein Combined with Chitosan Magnetic Nanoparticles on Quality Attributes in Red Sea Bream (Pagrosomus major). Food Bioprocess Technol. 2019, 3, 409–421. [Google Scholar] [CrossRef]
- Xia, W.; Ma, L.; Chen, X.; Li, X.; Zhang, Y. Physicochemical and structural properties of composite gels prepared with myofibrillar protein and lecithin at various ionic strengths. Food Hydrocoll. 2018, 82, 135–143. [Google Scholar] [CrossRef]
- Cao, Y.; Xiong, Y.L. Chlorogenic acid-mediated gel formation of oxidatively stressed myofibrillar protein. Food Chem. 2015, 180, 235–243. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F-T Cycles | L* | a* | b* |
|---|---|---|---|
| 0 | 65.49 ± 1.13 a | −2.84 ± 0.10 a | −5.64 ± 0.40 a |
| 1 | 65.65 ± 0.26 a | −2.80 ± 0.27 a | −5.65 ± 1.10 a |
| 2 | 59.14 ±1.11 b | −3.46 ± 0.24 b | −5.66 ± 0.45 a |
| 3 | 51.89 ± 1.61 c | −2.35 ± 0.27 a | −7.49 ± 0.36 b |
| 4 | 57.90 ± 1.01 b | −3.01 ± 0.22 a | −8.10 ± 0.37 b |
| 5 | 54.02 ± 0.74 c | −3.21 ± 0.41 b | −5.79 ± 1.14 a |
| F-T Cycles | 0 | 1 | 2 | 3 | 4 | 5 |
| Hardness (g) | 42,438.83 ± 1628.43 a | 38,855.80 ± 591.46 b | 35,806.38 ± 237.32 b | 28,311.67 ± 612.78 c | 17,880.11 ± 51.20 d | 12,404.90 ± 1193.63 e |
| Springiness | 0.958 ± 0.00 a | 0.922 ± 0.00 a | 0.917 ± 0.01 a | 0.916 ± 0.03 a | 0.794 ± 0.17 bc | 0.526 ± 0.03 c |
| Cohesiveness | 0.740 ± 0.01 a | 0.738 ± 0.01 a | 0.572 ± 0.23 ab | 0.399 ± 0.02 ab | 0.246 ± 0.06 b | 0.25 ± 0.04 b |
| Chewiness | 30,086.68 ± 1682.64 a | 27,658.23 ± 3295.74 a | 26,425.71 ± 103.67 a | 22,171.80 ± 3120.66 ab | 15,435.80 ± 3105.86 bc | 10,326.34 ± 161.10 c |
| 0 | 1 | 2 | 3 | 4 | 5 | |
| ASP | 3.84 ± 0.15 e | 3.86 ± 0.01 e | 7.03 ± 0.11 d | 9.69 ± 0.03 c | 12.02 ± 0.00 a | 11.57 ± 0.07 b |
| Thr | 18.71 ± 1.81 d | 26.87 ± 0.09 c | 36.74 ± 0.29 b | 43.21 ± 0.14 a | 35.80 ± 0.08 b | 41.94 ± 0.10 a |
| Ser | 19.92 ± 2.14 e | 26.82 ± 0.06 d | 36.33 ± 0.04 b | 30.50 ± 0.30 c | 38.10 ± 0.08 b | 45.06 ± 0.25 a |
| Glu | 24.21 ± 1.79 e | 19.42 ± 0.02 f | 37.94 ± 0.35 d | 77.81 ± 0.10 a | 50.55 ± 0.04 c | 61.54 ± 0.11 b |
| Gly | 13.58 ± 1.25 e | 14.99 ± 0.05 de | 18.94 ± 0.11 c | 22.76 ± 0.01 b | 16.83 ± 0.01 d | 26.24 ± 0.04 a |
| Ala | 34.00 ± 3.84 bc | 30.85 ± 0.05 c | 45.27 ± 0.28 a | 46.98 ± 0.02 a | 38.72 ± 0.06 b | 49.61 ± 0.13 a |
| Cys | 2.24 ± 0.25 d | 3.73 ± 0.03 c | 6.05 ± 0.05 a | 4.88 ± 0.04 b | 2.27 ± 0.03 d | 3.44 ± 0.11 c |
| Val | 17.23 ± 1.59 d | 16.22 ± 0.03 d | 22.60 ± 0.14 c | 30.23 ± 0.03 b | 30.71 ± 0.02 b | 36.66 ± 0.24 a |
| Met | 15.36 ± 1.21 e | 23.55 ± 0.05 d | 26.45 ± 0.18 c | 40.49 ± 0.08 a | 29.30 ± 0.12 b | 39.90 ± 0.96 a |
| lle | 12.78 ± 0.69 d | 9.53 ± 0.03 e | 13.20 ± 0.10 d | 19.63 ± 0.00 c | 26.06 ± 0.15 b | 30.86 ± 1.01 a |
| Leu | 28.54 ± 1.39 f | 38.41 ± 0.00 e | 45.18 ± 0.29 d | 71.34 ± 0.02 b | 54.57 ± 0.07 c | 78.45 ± 0.53 a |
| Tyr | 15.96 ± 0.32 d | 27.65 ± 0.03 c | 27.88 ± 0.24 c | 46.42 ± 0.24 a | 35.12 ± 0.16 b | 46.88 ± 0.60 a |
| Phe | 20.25 ± 1.42 f | 42.59 ± 0.16 d | 37.88 ± 0.26 e | 69.14 ± 0.25 b | 51.63 ± 0.35 c | 80.27 ± 0.84 a |
| Lys | 25.00 ± 0.13 f | 34.58 ± 0.07 e | 46.36 ± 0.33 d | 58.69 ± 0.08 b | 50.35 ± 0.08 c | 66.46 ± 0.24 a |
| His | 7.04 ± 0.25 e | 11.15 ± 0.00 d | 15.01 ± 0.11 c | 17.28 ± 0.02 b | 15.63 ± 0.37 c | 20.70 ± 0.02 a |
| Arg | 40.01 ± 6.33 bc | 20.22 ± 0.05 d | 48.69 ± 0.52 ab | 50.47 ± 0.01 a | 14.33 ± 0.00 d | 36.87 ± 0.01 c |
| Pro | 65.30 ± 10.16 b | 43.57 ± 0.49 b | 54.21 ± 1.76 b | 44.58 ± 1.47 b | 36.36 ± 1.50 b | 136.75 ± 16.00 a |
| Total | 363.95 ± 31.89 d | 394.01 ± 0.18 d | 525.74 ± 5.07 c | 684.09 ± 2.17 b | 538.35 ± 0.43 c | 813.19 ± 20.26 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, Y.; Xie, J. Effects of Freeze–Thaw Cycles on Water Migration, Microstructure and Protein Oxidation in Cuttlefish. Foods 2021, 10, 2576. https://doi.org/10.3390/foods10112576
Lv Y, Xie J. Effects of Freeze–Thaw Cycles on Water Migration, Microstructure and Protein Oxidation in Cuttlefish. Foods. 2021; 10(11):2576. https://doi.org/10.3390/foods10112576
Chicago/Turabian StyleLv, Ying, and Jing Xie. 2021. "Effects of Freeze–Thaw Cycles on Water Migration, Microstructure and Protein Oxidation in Cuttlefish" Foods 10, no. 11: 2576. https://doi.org/10.3390/foods10112576
APA StyleLv, Y., & Xie, J. (2021). Effects of Freeze–Thaw Cycles on Water Migration, Microstructure and Protein Oxidation in Cuttlefish. Foods, 10(11), 2576. https://doi.org/10.3390/foods10112576