Two Faces of Milk Proteins Peptides with Both Allergenic and Multidimensional Health Beneficial Impact—Integrated In Vitro/In Silico Approach



Abstract
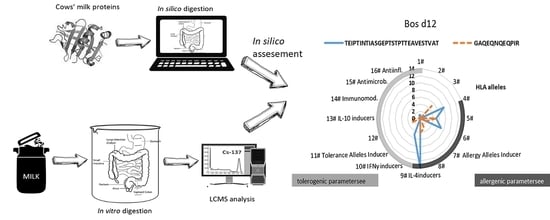
1. Introduction
2. Materials and Methods
2.1. Selection of Proteins for In Silico Analysis
2.2. In Silico Digestion
2.3. In Vitro Milk Digestion
2.4. Spectral Analyses of Milk Peptides
2.5. In Silico Analysis of Immunoreactive and Bioactive Peptides
2.5.1. The Evaluation of Binding Affinities to MHC II and T- and B-Cells Inducers
2.5.2. The Determination of IL4 Inducing MHC II Binders
2.5.3. The Determination of IFNγ Epitopes
2.5.4. The Determination of IL10 Inducing Binders
2.5.5. The Determination of Immune-Bioactive Peptides
2.6. Statistical Analysis and Data Calculation
2.7. In Silico Models Training and Validation
3. Results
3.1. In Silico Digestion
3.2. In Silico Antigenicity Assessment of In Silico Digested Proteins
3.3. In Vitro Digestion and Peptides Determination
3.4. In Silico Antigenicity Assessment of In Vitro Digested Proteins
3.5. Comparative In Silico Affinity Of Peptides to Chosen MHC II
3.6. Immunoreactive Activities Of Peptides
3.7. Integrated Evaluation of Peptides Potential
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- WHO Report. As More Go Hungry and Malnutrition Persists, Achieving Zero Hunger by 2030 in Doubt, UN Report Warns. Available online: https://www.who.int/news/item/13-07-2020-as-more-go-hungry-and-malnutrition-persists-achieving-zero-hunger-by-2030-in-doubt-un-report-warns (accessed on 13 July 2020).
- Mazzucchelli, G.; Holzhauser, T.; Velickovic, T.C.; Diaz-perales, A.; Molina, E.; Roncada, P.; Rodrigues, P.; Verhoeckx, K.; Hoffmann-sommergruber, K. Current (Food) Allergenic Risk Assessment: Is It Fit for Novel Foods? Status Quo and Identification of Gaps. Mol. Nutr. Food Res. 2018, 1700278. [Google Scholar] [CrossRef] [PubMed]
- Amigo, L.; Martinez-Maqueda, D.; Hernandez-Ledessma, B. In silico and in vitro analysis of multifunctionality. Foods 2020, 9, 991. [Google Scholar] [CrossRef] [PubMed]
- Brossard, C.; Pineau, F.; Triballeau, S.; Pietri, M. Allergy to deamidated gluten in patients tolerant to wheat: Specific epitopes linked to deamidation. J. Allergy Clin. Immunol. 2012, 67, 1023–1032. [Google Scholar] [CrossRef]
- Verhoeckx, K.; Bøgh, K.L.; Constable, A.; Epstein, M.M.; Sommergruber, K.H.; Holzhauser, T.; Houben, G.; Kuehn, A.; Roggen, E.; Mahony, L.O.; et al. COST Action ‘ImpARAS’: What have we learnt to improve food allergy risk assessment. A summary of a 4 year networking consortium. Clin. Transl. Allergy 2020, 1–12. [Google Scholar] [CrossRef]
- Sicherer, S.H.; Sampson, H.A. Food allergy: Epidemiology, pathogenesis, diagnosis, and treatment. J. Allergy Clin. Immunol. 2014, 133, 291–307. [Google Scholar] [CrossRef]
- Paparo, L.; Nocerino, R.; Bruno, C.; Di Scala, C.; Cosenza, L.; Bedogni, G.; Di Costanzo, M.; Mennini, M.; Argenio, V.D.; Salvatore, F.; et al. Randomized controlled trial on the influence of dietary intervention on epigenetic mechanisms in children with cow’s milk allergy: The EPICMA study. Nat. Sci. Rev. 2019, 1–10. [Google Scholar] [CrossRef]
- Bøgh, K.L.; Van Bilsen, J.; Głogowski, R.; Expósito, I.L.; Bouchaud, G.; Blanchard, C.; Bodinier, M.; Smit, J.; Pieters, R.; Net, S.B.; et al. Current challenges facing the assessment of the allergenic capacity of food allergens in animal models. Clin. Transl. Allergy 2016, 1–13. [Google Scholar] [CrossRef]
- Remington, B.; Broekman, H.C.H.; Blom, W.M.; Capt, A.; Crevel, R.W.R.; Dimitrov, I.; Faeste, C.K.; Fernandez-canton, R.; Giavi, S.; Houben, G.F.; et al. Approaches to assess IgE mediated allergy risks (sensitization and cross- reactivity) from new or modi fi ed dietary proteins. Food Chem. Toxicol. 2018, 112, 97–107. [Google Scholar] [CrossRef]
- Chatchatee, P.; Järvinen, K.M.; Bardina, L.; Beyer, K.; Sampson, H.A. Identification of IgE- and IgG-binding epitopes on αs1-casein: Differences patients with persistent and transient cow’s milk allergy. J. Allergy Clin. Immunol. 2001, 107, 379–383. [Google Scholar] [CrossRef]
- Yordanov, V.; Dimitrov, I.; Doytchinova, I. Proteochemometrics-Based Prediction of Peptide Binding to HLA-DP Proteins. J. Chem. Inf. Model. 2017. [Google Scholar] [CrossRef]
- Dimitrov, I.; Doytchinova, I. Associations between Milk and Egg Allergens and the HLA-DRB1/DQ Polymorphism: A Bioinformatics Approach. Int. Arch. Allergy Immunol. 2016, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Dhanda, S.K.; Gupta, S.; Vir, P.; Raghava, G.P.S. Prediction of IL4 Inducing Peptides. Clin. Dev. Immunol. 2013, 263952. [Google Scholar] [CrossRef] [PubMed]
- Dhanda, S.K.; Vir, P.; Raghava, G.P.S. Designing of interferon-gamma inducing MHC class-II binders. Biol. Direct 2013, 1–15. Available online: http://www.biology-direct.com/content/8/1/30 (accessed on 13 July 2020).
- Goodman, R.E. AllergenOnline Vesion 20; University of Nebraska: Lincoln, NV, USA, 2020; ISBN 1018736824. [Google Scholar]
- Mari, A.; Rasi, C.; Palazzo, P.; Scala, E. Allergen databases: Current status and perspectives. Curr. Allergy Asthma Rep. 2009, 9, 376–383. [Google Scholar] [CrossRef] [PubMed]
- Uniprot Database. Available online: https://www.uniprot.org (accessed on 13 July 2020).
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server; Springer Protocols Handbooks; Humana Press: Geneva, Switzerland, 2005; pp. 571–607. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutro, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food–An international consensus. Food Funct. 2014, 1113–1124. [Google Scholar] [CrossRef]
- Dimitrov, I.; Garnev, P.; Flower, D.R.; Doytchinova, I. EpiTOP-a proteochemometric tool for MHC class II binding prediction. Bioinformatics 2010, 26, 2066–2068. [Google Scholar] [CrossRef]
- Kadiyska, T.; Mladenova, M.; Dimitrov, I.; Doytchinova, I. Milk allergy in Hla-Drb1* 14:19/14: 21 paediatric patients: A bioinformatics approach. Pharmacia 2018, 65, 23–27. [Google Scholar]
- Jensen, K.K.; Andreatta, M.; Marcatili, P.; Buus, S.; Greenbaum, J.A.; Yan, Z.; Sette, A.; Peters, B.N. Improved methods for predicting peptide binding affinity to MHC class II molecules. Immunology 2018, 154, 394–406. [Google Scholar] [CrossRef]
- Kringelum, J.V.; Lundegaard, C.; Lund, O.; Nielsen, M.; Reliable, B. Cell epitope predictions: Impacts of method development and improved benchmarking. PLoS Comput. Biol. 2012, 8. [Google Scholar] [CrossRef]
- Saha, S.; Raghava, G.P.S. AlgPred: Prediction of allergenic proteins and mapping of IgE epitopes. Nucleic Acids Res. 2006, 34, 202–209. [Google Scholar] [CrossRef]
- Nagpal, G.; Usmani, S.S.; Dhanda, S.K.; Kaur, H. Computer-aided designing of immunosuppressive peptides based on IL-10 inducing potential. Nat. Publ. Gr. 2017, 1–10. [Google Scholar] [CrossRef]
- Wang, P.; Sidney, J.; Kim, Y.; Sette, A.; Lund, O.; Nielsen, M.; Peters, B. Peptide binding predictions for HLA DR, DP and DQ molecules. BMC Bioinform. 2010, 11, 568. Available online: http://www.biomedcentral.com/1471-2105/11/568 (accessed on 13 July 2020). [CrossRef] [PubMed]
- Meulenbroek, L.A.P.M.; Den Hartog Jager, C.F.; Lebens, A.F.M.; Knulst, A.C.; Bruijnzeel-Koomen, C.A.F.M.; Garssen, J.; Knippels, L.M.J.; Van Hoffen, E. Characterization of T cell epitopes in bovine α-lactalbumin. Int. Arch. Allergy Immunol. 2014, 163, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Maynard, F.; Jost, R.; Wal, J.M. Human IgE binding capacity of tryptic peptides from bovine alpha-lactalbumin. Int. Arch Allergy Immunol. 1997, 113, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Hochwallner, H.; Schulmeister, U.; Swoboda, I.; Focke-Tejkl, M.; Civaj, V.; Balic, N.; Nystrand, M.; Härlin, A.; Thalhamer, J.; Scheiblhofer, S.; et al. Visualization of clustered IgE epitopes on α-lactalbumin. J. Allergy Clin. Immunol. 2010, 125. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, H.; Inoue, R.; Kaneko, H.; Watanabe, M.; Suzuki, K.; Kato, Z.; Matsushita, S.; Kondo, N. Interaction among human leucocyte antigen-peptide-T cell receptor complexes in cow’s milk allergy: The significance of human leucocyte antigen and T cell receptor-complementarity determining region 3 loops. Clin. Exp. Allergy 2002, 32, 762–770. [Google Scholar] [CrossRef]
- Inoue, R.; Matsushita, S.; Kaneko, H.; Shinoda, S.; Sakaguchi, H.; Nishimura, Y.; Kondo, N. Identification of β-lactoglobulin-derived peptides and class II HLA molecules recognized by T cells from patients with milk allergy. Clin. Exp. Allergy 2001, 31, 1126–1134. [Google Scholar] [CrossRef]
- Tanabe, S.; Kobayashi, Y.; Takahata, Y.; Morimatsu, F.; Shibata, R.; Nishimura, T. Some human B and T cell epitopes of bovine serum albumin, the major beef allergen. Biochem. Biophys. Res. Commun. 2002, 293, 1348–1353. [Google Scholar] [CrossRef]
- Ruiter, B.; Trégoat, V.; M’Rabet, L.; Garssen, J.; Bruijnzeel-Koomen, C.A.F.M.; Knol, E.F.; Van Hoffen, E. Characterization of T cell epitopes in αs1-casein in cow’s milk allergic, atopic and non-atopic children. Clin. Exp. Allergy 2006, 36, 303–310. [Google Scholar] [CrossRef]
- Lin, J.; Bardina, L.; Shreffler, W.G.; Andreae, D.A.; Ge, Y.; Wang, J.; Bruni, F.M.; Fu, Z.; Han, Y.; Sampson, H.A. Development of a novel peptide microarray for large-scale epitope mapping of food allergens. J. Allergy Clin. Immunol. 2009, 124, 315–322. [Google Scholar] [CrossRef]
- Cerecedo, I.; Zamora, J.; Shreffler, W.G.; Lin, J.; Bardina, L.; Dieguez, M.C.; Wang, J.; Muriel, A.; de la Hoz, B.; Sampson, H.A. Mapping of the IgE and IgG4 sequential epitopes of milk allergens with a peptide microarray-based immunoassay. J. Allergy Clin. Immunol. 2008, 122, 589–594. [Google Scholar] [CrossRef]
- Lisson, M.; Novak, N.; Erhardt, G. Immunoglobulin E epitope mapping by microarray immunoassay reveals differences in immune response to genetic variants of caseins from different ruminant species. J. Dairy Sci. 2014, 97, 1939–1954. [Google Scholar] [CrossRef] [PubMed]
- Wildner, G.; Diedrichs-Möhring, M. Autoimmune uveitis induced by molecular mimicry of peptides from rotavirus, bovine casein and retinal S-antigen. Eur. J. Immunol. 2003, 33, 2577–2587. [Google Scholar] [CrossRef] [PubMed]
- Food Allergy Diagnostics & Therapeutics Market Size, Share, & Trends Analysis Report By Product Type (Therapeutic), By Allergen Source (Peanut), By End Use (Hospitals & Clinics), By Region, And Segment Forecasts, 2019–2026. Available online: https://www.factmr.com/report/2456/stationary-fuel-cell-systems-market (accessed on 3 October 2019).
- Wróblewska, B.; Szyc, A.M.; Markiewicz, L.H.; Zakrzewska, M.; Romaszko, E. Increased prevalence of eating disorders as a biopsychosocial implication of food allergy. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Kreiner, E.; Waage, J.; Standl, M.; Brix, S.; Pers, T.H.; Couto Alves, A.; Warrington, N.M.; Tiesler, C.M.T.; Fuertes, E.; Franke, L.; et al. Shared genetic variants suggest common pathways in allergy and autoimmune diseases. J. Allergy Clin. Immunol. 2017, 140, 771–781. [Google Scholar] [CrossRef]
- Ogrodowczyk, A.M.; Zakrzewska, M.; Romaszko, E.; Wroblewska, B. Gestational Dysfunction-Driven Diets and Probiotic Supplementation Correlate with the Profile of Allergen-Specific Antibodies in the Serum of Allergy Sufferers. Nutrients 2020, 12, 2381. [Google Scholar] [CrossRef]
- Albenzio, M.; Santillo, A.; Caroprese, M.; Malva, A. Bioactive Peptides in Animal Food Products. Foods 2017, 6, 35. [Google Scholar] [CrossRef]
- Bottani, M.; Cattaneo, S.; Pica, V.; Stuknyt, M.; Gomarasca, M.; Banfi, G.; De Noni, I.; Ferraretto, A.; Lombardi, G. Gastrointestinal In Vitro Digests of Infant Biscuits Formulated with Bovine Milk Proteins Positively. Foods 2020, 9, 1510. [Google Scholar] [CrossRef]
- Janeway, C.A., Jr.; Travers, P.; Walport, M.; Shlomchik, M.J. Immunobiology: The Immune System in Health and Disease, 5th ed.; Garland Science: New York, NY, USA, 2001. [Google Scholar]
- Sidney, J.; Assarsson, E.; Moore, C.; Ngo, S.; Pinilla, C.; Sette, A.; Peters, B. Quantitative peptide binding motifs for 19 human and mouse MHC class I molecules derived using positional scanning combinatorial peptide libraries. Immunome Res. 2008, 14, 1–14. [Google Scholar] [CrossRef]
- Ivashkiv, L.B.; Program, T.D.; Rosensweig, D.Z.; Program, M.P.; Medicine, W.C. HHS Public Access. Nat. Rev. Immunol. 2019, 18, 545–558. [Google Scholar] [CrossRef]
- Macierzanka, A.; Sancho, A.I.; Mills, E.N.C.; Rigby, N.M.; Mackie, A.R. Emulsification alters simulated gastrointestinal proteolysis of b -casein and b -lactoglobulin. Soft Matter 2009, 538–550. [Google Scholar] [CrossRef]
- Bøgh, K.L.; Madsen, C.B. Critical reviews in food science and nutrition food allergens: Is there a correlation between stability to digestion and allergenicity? Food Sci. Nutr. 2015, 37–41. [Google Scholar] [CrossRef]
- Verhoeckx, K.; Lindholm, K.; Dupont, D.; Egger, L.; Gadermaier, G.; Larré, C.; Mackie, A.; Menard, O.; Adel-patient, K.; Picariello, G.; et al. The relevance of a digestibility evaluation in the allergenicity risk assessment of novel proteins. Opinion of a joint initiative of COST action ImpARAS and COST action INFOGEST. Food Chem. Toxicol. 2019, 129, 405–423. [Google Scholar] [CrossRef] [PubMed]
- Pang, K.S.; Peng, H.B.; Noh, K. The Segregated Intestinal Flow Model (SFM) for drug absorption and drug metabolism: Implications on intestinal and liver metabolism and drug–drug interactions. Pharmaceutics 2020, 12, 312. [Google Scholar] [CrossRef] [PubMed]
- Fotschki, J.; Wróblewska, B.; Fotschki, B.; Kalicki, B.; Rigby, N.; Mackie, A. Microbial transglutaminase alters the immunogenic potential and cross-reactivity of horse and cow milk proteins. J. Dairy Sci. 2020, 103, 3. [Google Scholar] [CrossRef] [PubMed]
- Kalčáková, L.; Tremlová, B.; Pospiech, M.; Hostovský, M.; Dordević, D.; Javůrková, Z.; Běhalová, H.; Bartlová, M. Use of IHF-QD microscopic analysis for the detection of food allergenic components: Peanuts and wheat protein. Foods 2020, 9, 239. [Google Scholar] [CrossRef]
- Elsayed, S.; Hill, D.J.; Do, T.V. Evaluation of the allergenicity and antigenicity of bovine-milk αs1-casein using extensively purified synthetic peptides. Scand. J. Immunol. 2004, 60, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Ueno, H.M.; Kato, T.; Ohnishi, H.; Kawamoto, N.; Kato, Z.; Kaneko, H.; Kondo, N.; Nakano, T. T-cell epitope-containing hypoallergenic β-lactoglobulin for oral immunotherapy in milk allergy. Pediatr. Allergy Immunol. 2016, 27, 818–824. [Google Scholar] [CrossRef]
- Liu, J.; Chen, W.M.; Shao, Y.H.; Zhang, J.L.; Tu, Z.C. The mechanism of the reduction in allergenic reactivity of bovine α-lactalbumin induced by glycation, phosphorylation and acetylation. Food Chem. 2020, 310, 125853. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Protein | Peptides Surviving after In Silico Digestion | Start-End Position | Number of HLA Alleles with Weak Binders * | Number of HLA Alleles with Strong Binders * | T Cell Epitopes ** Start-End Position | IgE Epitopes ** | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DRB1 | DQ | DP | DRB1 | DQ | DP | Start-End Position | Ref. | Start-End Position | Ref. | |||
| Bos d4 | DTQAIVQN-NDSTE | 56–68 | 7 | 1 | 5 | 1 | 4 | 2 | 58–75 | [27] | 36–77 | [28] |
| SSNICNISCDK | 88–98 | 0 | 1 | 0 | 0 | 4 | 0 | 74–91 | [27] | 79–98 | [29] | |
| Bos d5 | ENSAEPEQS | 124–132 | 0 | 5 | 3 | 0 | 0 | 0 | 113–133 | [23,24] | [30,31] | |
| Bos d6 | QECCQAEDK | 189–197 | 0 | 3 | 0 | 0 | 0 | 0 | - | |||
| ICDNQDTI-SK | 287–297 | 2 | 3 | 3 | 9 | 2 | 3 | - | ||||
| DAIPENLPPL | 319–328 | 0 | 0 | 0 | 0 | 5 | 7 | 317–336 | [32] | |||
| VPQVSTPTL | 438–446 | 6 | 0 | 3 | 3 | 5 | 4 | - | ||||
| CCTKPESER | 460–468 | 0 | 4 | 1 | 0 | 1 | 0 | - | ||||
| Bos d9 | DIGSESTED-QAM | 58–69 | 6 | 0 | 5 | 2 | 5 | 1 | 58–81 | [33] | 54–63, 64–78 | [10,34] |
| EAESISSSEE-IVPNSVEQK | 76–94 | 0 | 0 | 1 | 8 | 5 | 6 | 88–111 | [33] | 84–93 | [10] | |
| EIVPNSAEER | 125–134 | 3 | 0 | 6 | 2 | 5 | 1 | 124–135 | [33] | 124–135 | [10] | |
| SDIPNPIGSE-NSEK | 195–208 | 8 | 0 | 6 | 0 | 5 | 1 | 188–207, 190–209, 194–213 | [10,34] | |||
| Bos d10 | VSSSEESIISQ-ET | 22–34 | 2 | 0 | 2 | 1 | 5 | 5 | - | 16–34, 28–47 | [35,36] | |
| SIGSSSEESA-EVATEEVK | 68–85 | 5 | 0 | 4 | 3 | 5 | 3 | 73–84 | [37] | 57–71, 82–101 | [34,35] | |
| NAVPITPTL | 130–138 | 0 | 1 | 1 | 0 | 4 | 0 | - | 132–143 | [36] | ||
| Bos d11 | NVPGEIVES | 22–30 | 0 | 3 | 1 | 0 | 2 | 0 | - | 16–31 | [10] | |
| SSSEESITR | 32–40 | 0 | 1 | 1 | 0 | 0 | 0 | - | 31–50 | [34] | ||
| QSEEQQQTE-DE | 49–59 | 0 | 5 | 1 | 0 | 0 | 0 | - | 49–65 | [35] | ||
| PFPGPIPNSL | 76–85 | 6 | 1 | 0 | 3 | 4 | 7 | - | 70–85 | [10] | ||
| TQTPVVVPPF | 93–102 | 0 | 0 | 7 | 0 | 5 | 0 | - | 98–107 | [10] | ||
| Bos d12 | GAQEQNQE-QPIR | 20–31 | 0 | 0 | 5 | 0 | 5 | 2 | - | |||
| SCQAQPTTM | 108–116 | 0 | 0 | 7 | 0 | 5 | 0 | - | ||||
| TEIPTINTIAS-GEPTSTPTTE-AVESTVAT | 138–166 | 1 | 0 | 0 | 8 | 5 | 7 | - | 132–147 | [10] | ||
| EDSPEVIESP-PEINTVQVT-STAV | 168–190 | 4 | 0 | 0 | 7 | 5 | 7 | - | 170–189 | [10] | ||
| Protein | Accession a | Score b | Coverage c [%] | Mass d | PAI e | T Cell Inductor f | Identified Peptides and Its Alergenic Potential/IgE Allergen g | B Cell Inductor h |
|---|---|---|---|---|---|---|---|---|
| Bos d 9 | P02662 | 32,934 | 43 | 24,570 | 195.07 | 25 | GYLEQLLRLKKYKV-PQLEIVPNSA; | 12 |
| SMKEGIHAQQKEP-MIGVNQELAYF; | ||||||||
| QKHIQKEDVPSERY-LGYLEQLLRL; #SDIPNPIGSENSEK | ||||||||
| Bos d 5 | P02754 | 18,489 | 33 | 20,269 | 186.95 | 21 | LVRTPEVDDEALEK-FDKALKALPM | 7 |
| Bos d 11 | P02666 | 3930 | 33 | 25,148 | 4.25 | 25 | - | 13 |
| Bos d 4 | P00711 | 1387 | 26 | 16,236 | 4.99 | 18 | # NNKIWCKDDQNP-SSNICNISCDK | 6 |
| Bos d 10 | P02663 | 688 | 19 | 26,173 | 1.61 | 26 | - | 12 |
| Osteopontin | P31096 | 393 | 33 | 31,000 | 1.95 | 31 | - | 8 |
| Butyrophilin subfamily 1 | P18892 | 171 | 5 | 59,923 | 0.15 | 57 | - | 22 |
| Glycosylation-dependent cell adhesion molecule | P80195 | 81 | 23 | 17,198 | 1.05 | 21 | - | 6 |
| α-1-glycoprotein Bos d2 | Q5GN72 | 74 | 5 | 23,158 | 0.11 | 40 | - | 15 |
| Collagen α-1 (III) chain | P04258 | 71 | 1 | 93,708 | 0.05 | 109 | GPVGNPGPAGPAG-PRGEVGLPGLS | 3 |
| Protein | Peptides Surviving After In Silico Digestion | Start-End Position | Number of HLA Alleles with Weak Binders * | Number of HLA Alleles with Strong Binders * | ||||
|---|---|---|---|---|---|---|---|---|
| DRB1 | DQ | DP | DRB1 | DQ | DP | |||
| Bos d 4 | NNKIWCKDDQNPHS- SNICNISCDK | 75–98 | 1 | 1 | 4 | 7 | 4 | 3 |
| Bos d 5 | LVRTPEVDDEALEK-FDKALKALPM | 139–163 | 3 | 0 | 0 | 9 | 5 | 7 |
| Bos d 9 | GYLEQLLRLKKYKV-PQLEIVPNSA | 108–131 | 0 | 0 | 0 | 12 | 5 | 7 |
| SMKEGIHAQQKEP-MIGVNQELAYF | 137–160 | 3 | 0 | 0 | 8 | 5 | 7 | |
| SDIPNPIGSENSK | 195–208 | 8 | 0 | 6 | 0 | 5 | 1 | |
| QKHIQKEDVPSERYL-GYLEQLLRL | 94–116 | 1 | 0 | 0 | 8 | 5 | 7 | |
| Collagen α-1 | GPVGNPGPAGPAGP-RGEVGLPGLS | 266–289 | 6 | 0 | 4 | 2 | 5 | 3 |
| Affinity IC50values [nM] Peptides Surviving after Digestion | Involved in Milk Allergy | Involved in Tolerance | ||||||
|---|---|---|---|---|---|---|---|---|
| DRB1 *01:01 | DQ7 (DQA1 *05:01/DQB1 *03:01) | DQ8 (DQA1 *03:01/ DQB1 *03:02) | DRB1 *03:01 | DRB1 *14:19 | DRB1 *14:21 | |||
| (A) IS | Bos d 4 | DTQAIVQNNDSTE | 121.63 | 101.36 | 76.41 | 93.56 | 96.68 | 134.10 |
| #SSNICNISCDK | 502.78 | 308.52 | 291.38 | 365.66 | 445.64 | 462.78 | ||
| Bos d 6 | ICDNQDTISSK | 462.78 | 514.2 | 354.23 | 234.25 | 359.94 | 468.5 | |
| Bos d 9 | DIGSESTEDQAM | 266.88 | 240.19 | 68.2 | 169.02 | 287.63 | 287.63 | |
| EAESISSSEEIVPNSVEQK | 462.53 | 107.92 | 16.96 | 174.73 | 277.52 | 426.55 | ||
| # SDIPNPIGSENSEK | 96.91 | 67.84 | 48.46 | 99.07 | 105.53 | 106.6 | ||
| Bos d 10 | VSSSEESIISQET | 124.75 | 46.78 | 1.72 | 102.92 | 145.02 | 146.58 | |
| SIGSSSEESAEVATEEVK | 272.29 | 47.35 | 0.53 | 260.45 | 293.01 | 295.97 | ||
| Bos d 11 | QSEEQQQTEDE | 531.34 | 508.49 | 165.69 | 565.62 | 571.34 | 571.34 | |
| Bos d 12 | GAQEQNQEQPIR | 278.74 | 281.70 | 204.61 | 275.77 | 278.74 | 287.63 | |
| TEIPTINTIASGEPTSTPTTEAVESTVAT | >5000 | 4253.87 | >5000 | >5000 | >5000 | >5000 | ||
| EDSPEVIESPPEINTVQVTSTAV | 973.05 | 858.58 | 629.62 | 2146.44 | 2432.64 | 2690.21 | ||
| (B) IV | Bos d 4 | #NNKIWCKDDQNPHSSNICNISCDK | 4244.21 | 3480.25 | 3862.23 | 3352.93 | 3650.02 | 4116.89 |
| Bos d 5 | LVRTPEVDDEALEKFDKALKALPM | 1952.34 | 3268.04 | 1273.26 | 2588.97 | 1527.92 | 1315.71 | |
| Bos d 9 | GYLEQLLRLKKYKVPQLEIVPNSA | 1145.94 | 3013.39 | 3310.49 | 2970.95 | 1612.8 | 373.49 | |
| SMKEGIHAQQKEPMIGVNQELAYF | 3437.81 | 3310.49 | 1230.82 | 1527.92 | 2376.76 | 1527.82 | ||
| # SDIPNPIGSENSEK | 96.91 | 67.84 | 48.46 | 99.07 | 105.53 | 106.6 | ||
| QKHIQKEDVPSERYLGYLEQLLRL | 1909.9 | 3055.83 | 1867.45 | 2037.22 | 2970.95 | 2291.87 | ||
| Collagen α-1 | GPVGNPGPAGPAG-PRGEVGLPGLS | 2776.07 | 343.43 | 2747.45 | 2833.31 | 2861.93 | 2861.93 | |
| Peptides Survived after Digestion | IL-4 Inducers a | IFN γ Inducers b | Bioactive Peptides c | ||||
|---|---|---|---|---|---|---|---|
| Inductors | SVM Score | Inductors | SVM Score | Function (Number of Sequences) | |||
| (A) IS | Bos d 4 | DTQAIVQNNDSTE | 0 | - | 2 | 0.33–0.39 | - |
| # SSNICNISCDK | 4 | 0.99–1.58 | 0 | - | - | ||
| Bos d 5 | ENSAEPEQS | 2 | 1.15–2.04 | 0 | - | - | |
| Bos d 6 | ICDNQDTISSK | 3 | 0.52–1.05 | 3 | 0.34–0.39 | - | |
| Bos d 9 | DIGSESTEDQAM | 2 | 0.21–0.38 | 0 | - | - | |
| EAESISSSEEIVPNSVEQK | 2 | 0.25–0.99 | 1 | 0.75 | Immunomod. (3) | ||
| # SDIPNPIGSENSEK | 7 | 0.56–1.66 | 2 | 0.07–0.35 | Antimicrobial (1) Immunomod. (1) | ||
| Bos d 10 | VSSSEESIISQET | 1 | 0.26 | 2 | 0.01–0.39 | - | |
| SIGSSSEESAEVATEEVK | 5 | 1.12–1.92 | 8 | 0.05–0.72 | - | ||
| Bos d 11 | QSEEQQQTEDE | 0 | - | 0 | - | - | |
| Bos d 12 | GAQEQNQEQPIR | 4 | 0.47–0.8 | 0 | - | Antiinflam. (1) | |
| TEIPTINTIASGEPTSTPTTEAVESTVAT | 14 | 0.29–1.14 | 5 | 0.11–0.31 | Antimicrobial (1) | ||
| EDSPEVIESPPEINTVQVTSTAV | 0 | - | 0 | - | Antimicrobial (1) | ||
| (B) IV | Bos d 4 | # NNKIWCKDDQNPHSSNICNISCDK | 1 | 0.27 | 0 | - | Antiinflam. (2) Antimicrobial (1) Immunomod. (1) |
| Bos d 5 | LVRTPEVDDEALEKFDKALKALPM | 0 | - | 5 | 0.11–0.31 | Antiinflam. (2) IL-10 Inducer (2) d | |
| Bos d 9 | GYLEQLLRLKKYKVPQLEIVPNSA | 1 | 0.89 | 1 | 0.04 | Antiinflam. (3) Immunomod. (1) IL-10 Inducer (2) d | |
| SMKEGIHAQQKEPMIGVNQELAYF | 2 | 0.32–0.47 | 0 | - | Antiinflam. (3) | ||
| # SDIPNPIGSENSEK | 7 | 0.56–1.66 | 0 | - | Antimicrobial (1) Immunomod. (1) | ||
| QKHIQKEDVPSERYLGYLEQLLRL | 1 | 0.51 | 2 | 0.21–0.63 | Antiinflam. (3) Immunomod. (1) IL-10 Inducer (1) d | ||
| Collagen α-1 | GPVGNPGPAGPAGPRGEVGLPGLS | 1 | 0.54 | 2 | 0.04–0.11 | Antiinflam. (2) Immunomod. (1) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogrodowczyk, A.M.; Dimitrov, I.; Wróblewska, B. Two Faces of Milk Proteins Peptides with Both Allergenic and Multidimensional Health Beneficial Impact—Integrated In Vitro/In Silico Approach. Foods 2021, 10, 163. https://doi.org/10.3390/foods10010163
Ogrodowczyk AM, Dimitrov I, Wróblewska B. Two Faces of Milk Proteins Peptides with Both Allergenic and Multidimensional Health Beneficial Impact—Integrated In Vitro/In Silico Approach. Foods. 2021; 10(1):163. https://doi.org/10.3390/foods10010163
Chicago/Turabian StyleOgrodowczyk, Anna Maria, Ivan Dimitrov, and Barbara Wróblewska. 2021. "Two Faces of Milk Proteins Peptides with Both Allergenic and Multidimensional Health Beneficial Impact—Integrated In Vitro/In Silico Approach" Foods 10, no. 1: 163. https://doi.org/10.3390/foods10010163
APA StyleOgrodowczyk, A. M., Dimitrov, I., & Wróblewska, B. (2021). Two Faces of Milk Proteins Peptides with Both Allergenic and Multidimensional Health Beneficial Impact—Integrated In Vitro/In Silico Approach. Foods, 10(1), 163. https://doi.org/10.3390/foods10010163