
GDF15 Contributes to the Regulation of the Mechanosensitive Responses of PdL Fibroblasts through the Modulation of IL-37

Abstract
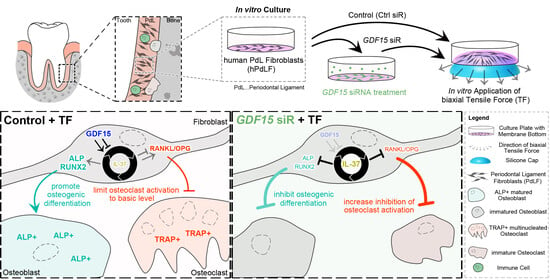
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Tensile Strain Application
2.3. siRNA-Mediated Down-regulations of GDF15 and IL37
2.4. RNA Extraction and Quantitative Polymerase Chain Reaction (PCR)
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. THP1 Activation Assay
2.7. Osteoclast Activation Assay and TRAP Staining
2.8. ALP Activity Assay
2.9. Immunofluorescent Staining
2.10. Microscopy and Image Analysis
2.11. Statistics
3. Results
3.1. GDF15 Limits IL-37 in Stretched PdL Fibroblasts without Impacting Their Anti-Inflammatory Mechanoresponse
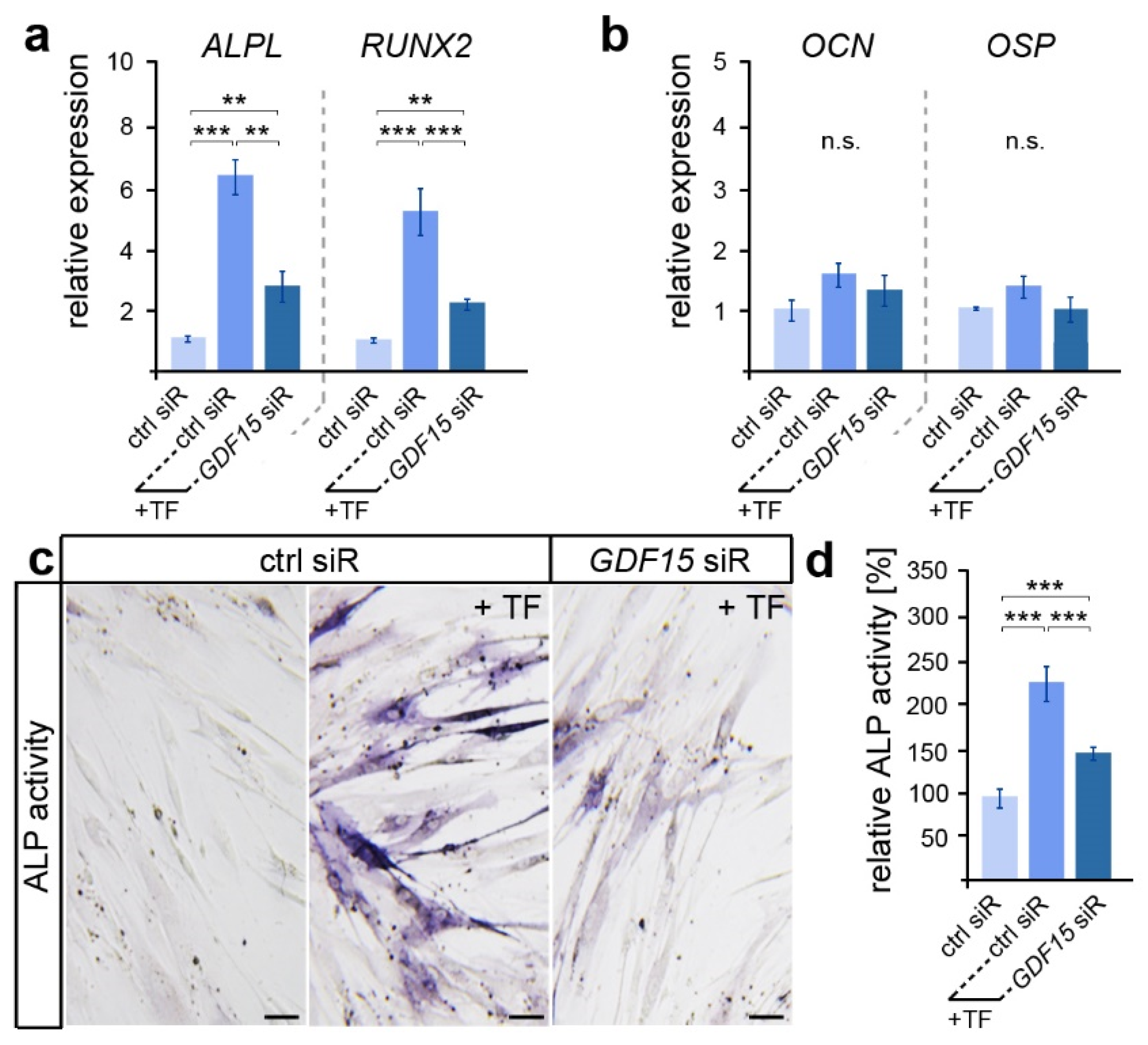
3.2. GDF15 Promotes the Differentiation of Stretched PdL Fibroblasts
3.3. GDF15 Balances the Activity of Osteoclasts in Elongated PdL Fibroblasts Potentially through an IL-37-Dependent Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Navabi, N.; Farnudi, H.; Rafiei, H.; Arashlow, M.T. Orthodontic treatment and the oral health-related quality of life of patients. J. Dent. 2012, 9, 247–254. [Google Scholar]
- Reitan, K. Clinical and histologic observations on tooth movement during and after orthodontic treatment. Am. J. Orthod. 1967, 53, 721–745. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jacox, L.A.; Little, S.H.; Ko, C.C. Orthodontic tooth movement: The biology and clinical implications. Kaohsiung J. Med. Sci. 2018, 34, 207–214. [Google Scholar] [CrossRef]
- Asiry, M.A. Biological aspects of orthodontic tooth movement: A review of literature. Saudi J. Biol. Sci. 2018, 25, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Maltha, J.C.; Kuijpers-Jagtman, A.M. Mechanobiology of orthodontic tooth movement: An update. J. World Fed. Orthod. 2023, 12, 156–160. [Google Scholar] [CrossRef]
- Garlet, T.P.; Coelho, U.; Silva, J.S.; Garlet, G.P. Cytokine expression pattern in compression and tension sides of the periodontal ligament during orthodontic tooth movement in humans. Eur. J. Oral Sci. 2007, 115, 355–362. [Google Scholar] [CrossRef]
- Pinkerton, M.N.; Wescott, D.C.; Gaffey, B.J.; Beggs, K.T.; Milne, T.J.; Meikle, M.C. Cultured human periodontal ligament cells constitutively express multiple osteotropic cytokines and growth factors, several of which are responsive to mechanical deformation. J. Periodontal. Res. 2008, 43, 343–351. [Google Scholar] [CrossRef]
- Toyama, N.; Ono, T.; Ono, T.; Nakashima, T. The interleukin-6 signal regulates orthodontic tooth movement and pain. Biochem. Biophys. Res. Commun. 2023, 684, 149068. [Google Scholar] [CrossRef]
- Meikle, M.C. The tissue, cellular, and molecular regulation of orthodontic tooth movement: 100 years after carl sandstedt. Eur. J. Orthod. 2006, 28, 221–240. [Google Scholar] [CrossRef]
- Li, M.; Zhang, C.; Yang, Y. Effects of mechanical forces on osteogenesis and osteoclastogenesis in human periodontal ligament fibroblasts: A systematic review of in vitro studies. Bone Jt. Res. 2019, 8, 19–31. [Google Scholar] [CrossRef]
- Baloul, S.S. Osteoclastogenesis and osteogenesis during tooth movement. Front. Oral Biol. 2016, 18, 75–79. [Google Scholar] [PubMed]
- Yamaguchi, M. Rank/rankl/opg during orthodontic tooth movement. Orthod. Craniofacial Res. 2009, 12, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Marcadet, L.; Bouredji, Z.; Argaw, A.; Frenette, J. The roles of rank/rankl/opg in cardiac, skeletal, and smooth muscles in health and disease. Front. Cell Dev. Biol. 2022, 10, 903657. [Google Scholar] [CrossRef] [PubMed]
- Udagawa, N.; Koide, M.; Nakamura, M.; Nakamichi, Y.; Yamashita, T.; Uehara, S.; Kobayashi, Y.; Furuya, Y.; Yasuda, H.; Fukuda, C.; et al. Osteoclast differentiation by rankl and opg signaling pathways. J. Bone Miner. Metab. 2021, 39, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Kanzaki, H.; Chiba, M.; Shimizu, Y.; Mitani, H. Periodontal ligament cells under mechanical stress induce osteoclastogenesis by receptor activator of nuclear factor kappab ligand up-regulation via prostaglandin e2 synthesis. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2002, 17, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Nishijima, Y.; Yamaguchi, M.; Kojima, T.; Aihara, N.; Nakajima, R.; Kasai, K. Levels of rankl and opg in gingival crevicular fluid during orthodontic tooth movement and effect of compression force on releases from periodontal ligament cells in vitro. Orthod. Craniofacial Res. 2006, 9, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Aihara, N.; Kojima, T.; Kasai, K. Rankl increase in compressed periodontal ligament cells from root resorption. J. Dent. Res. 2006, 85, 751–756. [Google Scholar] [CrossRef]
- Tan, L.; Ren, Y.; Wang, J.; Jiang, L.; Cheng, H.; Sandham, A.; Zhao, Z. Osteoprotegerin and ligand of receptor activator of nuclear factor kappab expression in ovariectomized rats during tooth movement. Angle Orthod. 2009, 79, 292–298. [Google Scholar] [CrossRef]
- Boyce, B.F.; Xing, L. The rankl/rank/opg pathway. Curr. Osteoporos. Rep. 2007, 5, 98–104. [Google Scholar] [CrossRef]
- Kanzaki, H.; Chiba, M.; Sato, A.; Miyagawa, A.; Arai, K.; Nukatsuka, S.; Mitani, H. Cyclical tensile force on periodontal ligament cells inhibits osteoclastogenesis through opg induction. J. Dent. Res. 2006, 85, 457–462. [Google Scholar] [CrossRef]
- Holland, R.; Bain, C.; Utreja, A. Osteoblast differentiation during orthodontic tooth movement. Orthod. Craniofacial Res. 2019, 22, 177–182. [Google Scholar] [CrossRef]
- Basdra, E.K.; Komposch, G. Osteoblast-like properties of human periodontal ligament cells: An in vitro analysis. Eur. J. Orthod. 1997, 19, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Kawarizadeh, A.; Bourauel, C.; Gotz, W.; Jager, A. Early responses of periodontal ligament cells to mechanical stimulus in vivo. J. Dent. Res. 2005, 84, 902–906. [Google Scholar] [CrossRef]
- Tang, N.; Zhao, Z.; Zhang, L.; Yu, Q.; Li, J.; Xu, Z.; Li, X. Up-regulated osteogenic transcription factors during early response of human periodontal ligament stem cells to cyclic tensile strain. Arch. Med. Sci. 2012, 8, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Losch, L.; Stemmler, A.; Fischer, A.; Steinmetz, J.; Schuldt, L.; Hennig, C.L.; Symmank, J.; Jacobs, C. Gdf15 promotes the osteogenic cell fate of periodontal ligament fibroblasts, thus affecting their mechanobiological response. Int. J. Mol. Sci. 2023, 24, 10011. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Qiu, L.; Chang, H.; Yang, Y.; Jian, C.; Xiong, J.; Zhou, J.; Dong, S. Cyclic tension promotes osteogenic differentiation in human periodontal ligament stem cells. Int. J. Clin. Exp. Pathol. 2014, 7, 7872–7880. [Google Scholar] [PubMed]
- Symmank, J.; Zimmermann, S.; Goldschmitt, J.; Schiegnitz, E.; Wolf, M.; Wehrbein, H.; Jacobs, C. Mechanically-induced gdf15 secretion by periodontal ligament fibroblasts regulates osteogenic transcription. Sci. Rep. 2019, 9, 11516. [Google Scholar] [CrossRef] [PubMed]
- Stemmler, A.; Symmank, J.; Steinmetz, J.; von Brandenstein, K.; Hennig, C.L.; Jacobs, C. Gdf15 supports the inflammatory response of pdl fibroblasts stimulated by p. Gingivalis lps and concurrent compression. Int. J. Mol. Sci. 2021, 22, 13608. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, Q.; Zhu, Y.; Hu, W. Gdf15 induced by compressive force contributes to osteoclast differentiation in human periodontal ligament cells. Exp. Cell Res. 2020, 387, 111745. [Google Scholar] [CrossRef]
- Bootcov, M.R.; Bauskin, A.R.; Valenzuela, S.M.; Moore, A.G.; Bansal, M.; He, X.Y.; Zhang, H.P.; Donnellan, M.; Mahler, S.; Pryor, K.; et al. Mic-1, a novel macrophage inhibitory cytokine, is a divergent member of the tgf-beta superfamily. Proc. Natl. Acad. Sci. USA 1997, 94, 11514–11519. [Google Scholar] [CrossRef]
- Sarkar, S.; Legere, S.; Haidl, I.; Marshall, J.; MacLeod, J.B.; Aguiar, C.; Lutchmedial, S.; Hassan, A.; Brunt, K.R.; Kienesberger, P.; et al. Serum gdf15, a promising biomarker in obese patients undergoing heart surgery. Front. Cardiovasc. Med. 2020, 7, 103. [Google Scholar] [CrossRef]
- Conte, M.; Giuliani, C.; Chiariello, A.; Iannuzzi, V.; Franceschi, C.; Salvioli, S. Gdf15, an emerging key player in human aging. Ageing Res. Rev. 2022, 75, 101569. [Google Scholar] [CrossRef] [PubMed]
- Rochette, L.; Zeller, M.; Cottin, Y.; Vergely, C. Insights into mechanisms of gdf15 and receptor gfral: Therapeutic targets. Trends Endocrinol. Metab. TEM 2020, 31, 939–951. [Google Scholar] [CrossRef] [PubMed]
- Mullican, S.E.; Lin-Schmidt, X.; Chin, C.N.; Chavez, J.A.; Furman, J.L.; Armstrong, A.A.; Beck, S.C.; South, V.J.; Dinh, T.Q.; Cash-Mason, T.D.; et al. Gfral is the receptor for gdf15 and the ligand promotes weight loss in mice and nonhuman primates. Nat. Med. 2017, 23, 1150–1157. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Jiang, D.; Matsuda, J.L.; Ternyak, K.; Zhang, B.; Chu, H.W. Cigarette smoke induces human airway epithelial senescence via growth differentiation factor 15 production. Am. J. Respir. Cell Mol. Biol. 2016, 55, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Artz, A.; Butz, S.; Vestweber, D. Gdf-15 inhibits integrin activation and mouse neutrophil recruitment through the alk-5/tgf-betarii heterodimer. Blood 2016, 128, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Min, K.W.; Liggett, J.L.; Silva, G.; Wu, W.W.; Wang, R.; Shen, R.F.; Eling, T.E.; Baek, S.J. Nag-1/gdf15 accumulates in the nucleus and modulates transcriptional regulation of the smad pathway. Oncogene 2016, 35, 377–388. [Google Scholar] [CrossRef]
- Lee, J.; Kim, I.; Yoo, E.; Baek, S.J. Competitive inhibition by nag-1/gdf-15 nls peptide enhances its anti-cancer activity. Biochem. Biophys. Res. Commun. 2019, 519, 29–34. [Google Scholar] [CrossRef]
- Nazet, U.; Schroder, A.; Spanier, G.; Wolf, M.; Proff, P.; Kirschneck, C. Simplified method for applying static isotropic tensile strain in cell culture experiments with identification of valid rt-qpcr reference genes for pdl fibroblasts. Eur. J. Orthod. 2020, 42, 359–370. [Google Scholar] [CrossRef]
- Symmank, J.; Appel, S.; Bastian, J.A.; Knaup, I.; Marciniak, J.; Hennig, C.L.; Doding, A.; Schulze-Spate, U.; Jacobs, C.; Wolf, M. Hyperlipidemic conditions impact force-induced inflammatory response of human periodontal ligament fibroblasts concomitantly challenged with p. Gingivalis-lps. Int. J. Mol. Sci. 2021, 22, 6069. [Google Scholar] [CrossRef]
- Schuldt, L.; Reimann, M.; von Brandenstein, K.; Steinmetz, J.; Doding, A.; Schulze-Spate, U.; Jacobs, C.; Symmank, J. Palmitate-triggered cox2/pge2-related hyperinflammation in dual-stressed pdl fibroblasts is mediated by repressive h3k27 trimethylation. Cells 2022, 11, 955. [Google Scholar] [CrossRef] [PubMed]
- Schuldt, L.; von Brandenstein, K.; Jacobs, C.; Symmank, J. Oleic acid-related anti-inflammatory effects in force-stressed pdl fibroblasts are mediated by h3 lysine acetylation associated with altered il10 expression. Epigenetics 2022, 17, 1892–1904. [Google Scholar] [CrossRef]
- Abulizi, P.; Loganathan, N.; Zhao, D.; Mele, T.; Zhang, Y.; Zwiep, T.; Liu, K.; Zheng, X. Growth differentiation factor-15 deficiency augments inflammatory response and exacerbates septic heart and renal injury induced by lipopolysaccharide. Sci. Rep. 2017, 7, 1037. [Google Scholar] [CrossRef]
- Wu, M.; Chen, G.; Li, Y.P. Tgf-beta and bmp signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone research 2016, 4, 16009. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Tao, X. Current understanding of il-37 in human health and disease. Front. Immunol. 2021, 12, 696605. [Google Scholar] [CrossRef]
- Talic, N.F. Adverse effects of orthodontic treatment: A clinical perspective. Saudi Dent. J. 2011, 23, 55–59. [Google Scholar] [CrossRef]
- Wischhusen, J.; Melero, I.; Fridman, W.H. Growth/differentiation factor-15 (gdf-15): From biomarker to novel targetable immune checkpoint. Front. Immunol. 2020, 11, 951. [Google Scholar] [CrossRef] [PubMed]
- Pence, B.D. Growth differentiation factor-15 in immunity and aging. Front. Aging 2022, 3, 837575. [Google Scholar] [CrossRef]
- Radwanska, A.; Cottage, C.T.; Piras, A.; Overed-Sayer, C.; Sihlbom, C.; Budida, R.; Wrench, C.; Connor, J.; Monkley, S.; Hazon, P.; et al. Increased expression and accumulation of gdf15 in ipf extracellular matrix contribute to fibrosis. JCI Insight 2022, 7, e153058. [Google Scholar] [CrossRef]
- Lin, W.; Zhang, W.W.; Lyu, N.; Cao, H.; Xu, W.D.; Zhang, Y.Q. Growth differentiation factor-15 produces analgesia by inhibiting tetrodotoxin-resistant nav1.8 sodium channel activity in rat primary sensory neurons. Neurosci. Bull. 2021, 37, 1289–1302. [Google Scholar] [CrossRef]
- Klein, A.B.; Nicolaisen, T.S.; Ortenblad, N.; Gejl, K.D.; Jensen, R.; Fritzen, A.M.; Larsen, E.L.; Karstoft, K.; Poulsen, H.E.; Morville, T.; et al. Pharmacological but not physiological gdf15 suppresses feeding and the motivation to exercise. Nat. Commun. 2021, 12, 1041. [Google Scholar] [CrossRef]
- Lorenz, G.; Ribeiro, A.; von Rauchhaupt, E.; Wurf, V.; Schmaderer, C.; Cohen, C.D.; Vohra, T.; Anders, H.J.; Lindenmeyer, M.; Lech, M. Gdf15 suppresses lymphoproliferation and humoral autoimmunity in a murine model of systemic lupus erythematosus. J. Innate Immun. 2022, 14, 673–689. [Google Scholar] [CrossRef]
- Quinn, J.M.; Itoh, K.; Udagawa, N.; Hausler, K.; Yasuda, H.; Shima, N.; Mizuno, A.; Higashio, K.; Takahashi, N.; Suda, T.; et al. Transforming growth factor beta affects osteoclast differentiation via direct and indirect actions. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2001, 16, 1787–1794. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, T.; Kawabata, H.; Miura, Y.; Yoshioka, S.; Iwasa, M.; Yao, H.; Sakamoto, S.; Fujimoto, M.; Haga, H.; Kadowaki, N.; et al. The role of growth differentiation factor 15 in the pathogenesis of primary myelofibrosis. Cancer Med. 2015, 4, 1558–1572. [Google Scholar] [CrossRef] [PubMed]
- Wakchoure, S.; Swain, T.M.; Hentunen, T.A.; Bauskin, A.R.; Brown, D.A.; Breit, S.N.; Vuopala, K.S.; Harris, K.W.; Selander, K.S. Expression of macrophage inhibitory cytokine-1 in prostate cancer bone metastases induces osteoclast activation and weight loss. Prostate 2009, 69, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, J.A.; Seshacharyulu, P.; Muniyan, S.; Pothuraju, R.; Khan, P.; Vengoji, R.; Chaudhary, S.; Maurya, S.K.; Lele, S.M.; Jain, M.; et al. Gdf15 promotes prostate cancer bone metastasis and colonization through osteoblastic ccl2 and rankl activation. Bone Res. 2022, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Deng, C.; Li, Y.P. Tgf-beta and bmp signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272–288. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.H.; Chen, D.Q.; Wang, Y.N.; Feng, Y.L.; Cao, G.; Vaziri, N.D.; Zhao, Y.Y. New insights into tgf-beta/smad signaling in tissue fibrosis. Chem. Biol. Interact. 2018, 292, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Abulkhir, A.; Samarani, S.; Amre, D.; Duval, M.; Haddad, E.; Sinnett, D.; Leclerc, J.M.; Diorio, C.; Ahmad, A. A protective role of il-37 in cancer: A new hope for cancer patients. J. Leukoc. Biol. 2017, 101, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ma, Y.M.; Zheng, P.S.; Zhang, P. Gdf15 promotes the proliferation of cervical cancer cells by phosphorylating akt1 and erk1/2 through the receptor erbb2. J. Exp. Clin. Cancer Res. 2018, 37, 80. [Google Scholar] [CrossRef]
- Sollazzo, V.; Pezzetti, F.; Massari, L.; Palmieri, A.; Brunelli, G.; Zollino, I.; Lucchese, A.; Caruso, G.; Carinci, F. Evaluation of gene expression in mg63 human osteoblastlike cells exposed to tantalum powder by microarray technology. Int. J. Periodontics Restor. Dent. 2011, 31, e17–e28. [Google Scholar]
- Ye, C.; Zhang, W.; Hang, K.; Chen, M.; Hou, W.; Chen, J.; Chen, X.; Chen, E.; Tang, L.; Lu, J.; et al. Extracellular il-37 promotes osteogenic differentiation of human bone marrow mesenchymal stem cells via activation of the pi3k/akt signaling pathway. Cell Death Dis. 2019, 10, 753. [Google Scholar] [CrossRef] [PubMed]
- Westhrin, M.; Moen, S.H.; Holien, T.; Mylin, A.K.; Heickendorff, L.; Olsen, O.E.; Sundan, A.; Turesson, I.; Gimsing, P.; Waage, A.; et al. Growth differentiation factor 15 (gdf15) promotes osteoclast differentiation and inhibits osteoblast differentiation and high serum gdf15 levels are associated with multiple myeloma bone disease. Haematologica 2015, 100, e511–e514. [Google Scholar] [CrossRef] [PubMed]
- Hinoi, E.; Ochi, H.; Takarada, T.; Nakatani, E.; Iezaki, T.; Nakajima, H.; Fujita, H.; Takahata, Y.; Hidano, S.; Kobayashi, T.; et al. Positive regulation of osteoclastic differentiation by growth differentiation factor 15 upregulated in osteocytic cells under hypoxia. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2012, 27, 938–949. [Google Scholar] [CrossRef]
- Vanhara, P.; Lincova, E.; Kozubik, A.; Jurdic, P.; Soucek, K.; Smarda, J. Growth/differentiation factor-15 inhibits differentiation into osteoclasts--a novel factor involved in control of osteoclast differentiation. Differ. Res. Biol. Divers. 2009, 78, 213–222. [Google Scholar]
- Olsen, O.E.; Skjaervik, A.; Stordal, B.F.; Sundan, A.; Holien, T. Tgf-beta contamination of purified recombinant gdf15. PLoS ONE 2017, 12, e0187349. [Google Scholar] [CrossRef]
- Tang, R.; Yi, J.; Yang, J.; Chen, Y.; Luo, W.; Dong, S.; Fei, J. Interleukin-37 inhibits osteoclastogenesis and alleviates inflammatory bone destruction. J. Cell. Physiol. 2019, 234, 7645–7658. [Google Scholar] [CrossRef]
- Jing, L.; Kim, S.; Sun, L.; Wang, L.; Mildner, E.; Divaris, K.; Jiao, Y.; Offenbacher, S. Il-37- and il-35/il-37-producing plasma cells in chronic periodontitis. J. Dent. Res. 2019, 98, 813–821. [Google Scholar] [CrossRef]
- Saeed, J.; Kitaura, H.; Kimura, K.; Ishida, M.; Sugisawa, H.; Ochi, Y.; Kishikawa, A.; Takano-Yamamoto, T. Il-37 inhibits lipopolysaccharide-induced osteoclast formation and bone resorption in vivo. Immunol. Lett. 2016, 175, 8–15. [Google Scholar] [CrossRef]
- Papadopoulou, K.; Keilig, L.; Eliades, T.; Krause, R.; Jager, A.; Bourauel, C. The time-dependent biomechanical behaviour of the periodontal ligament--an in vitro experimental study in minipig mandibular two-rooted premolars. Eur. J. Orthod. 2014, 36, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.L.; Liu, Y.F.; Peng, W.; Dong, H.Y.; Zhang, J.X. A biomechanical case study on the optimal orthodontic force on the maxillary canine tooth based on finite element analysis. J. Zhejiang Univ. Sci. B 2018, 19, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Suriben, R.; Chen, M.; Higbee, J.; Oeffinger, J.; Ventura, R.; Li, B.; Mondal, K.; Gao, Z.; Ayupova, D.; Taskar, P.; et al. Antibody-mediated inhibition of gdf15-gfral activity reverses cancer cachexia in mice. Nat. Med. 2020, 26, 1264–1270. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Symbol | NCBI Gene ID | Primer Sequence |
|---|---|---|---|
| Alkaline phosphatase | ALPL | 249 | fw ACTGCAGACATTCTCAAA rev GAGTGAGTGAGTGAGCA |
| Bone gamma- carboxyglutamate protein | BGLAP (alias OCN) | 632 | fw GCAGCGAGGTAGTGAAGAGA rev AGCAGAGCGACACCCTAGA |
| Growth differentiation factor 15 | GDF15 | 9518 | fw CCGAAGACTCCAGATTCCGA rev CCCGAGAGATACGCAGGTG |
| Interleukin 10 | IL10 | 3586 | fw AGCCATGAGTGAGTTTGACA rev AGAGCCCCAGATCCGATTTT |
| Interleukin 1 receptor antagonist | IL1RN | 3557 | fw GATGTGCCTGTCCTGTGTCA rev ACTCAAAACTGGTGGTGGGG |
| Interleukin 37 | IL37 | 27178 | fw AGTCCGATTCTCCTGGGG rev TTTATAGGGCTCAGGTGGGC |
| Ribosomal protein L22 | RPL22 | 6146 | fw TGATTGCACCCACCCTGTAG rev GGTTCCCAGCTTTTCCGTTC |
| RUNX family transcription factor 2 | RUNX2 | 860 | fw: CCCACGAATGCACTATCC rev: GGACATACCGAGGGACA |
| Secreted phosphoprotein 1 | SSP1 (alias OSP) | 6696 | fw TGATTTTCCCACGGACCTGC rev TCGCTTTCCATGTGTGAGGT |
| TATA box binding protein | TBP | 6908 | fw CGGCTGTTTAACTTCGCTTCC rev TGGGTTATCTTCACACGCCAAG |
| TNF receptor superfamily member 11b | TNFRSF11B (alias OPG) | 4982 | fw: GAAGGGCGCTACCTTGA rev: GCAAACTGTATTTCGCTC |
| TNF superfamily member 11 | TNFSF11 (alias RANKL) | 8600 | fw: ATCACAGCACATCAGAGCAGA rev: TCACTTTATGGGAACCAGATGGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steinmetz, J.; Stemmler, A.; Hennig, C.-L.; Symmank, J.; Jacobs, C. GDF15 Contributes to the Regulation of the Mechanosensitive Responses of PdL Fibroblasts through the Modulation of IL-37. Dent. J. 2024, 12, 39. https://doi.org/10.3390/dj12020039
Steinmetz J, Stemmler A, Hennig C-L, Symmank J, Jacobs C. GDF15 Contributes to the Regulation of the Mechanosensitive Responses of PdL Fibroblasts through the Modulation of IL-37. Dentistry Journal. 2024; 12(2):39. https://doi.org/10.3390/dj12020039
Chicago/Turabian StyleSteinmetz, Julia, Albert Stemmler, Christoph-Ludwig Hennig, Judit Symmank, and Collin Jacobs. 2024. "GDF15 Contributes to the Regulation of the Mechanosensitive Responses of PdL Fibroblasts through the Modulation of IL-37" Dentistry Journal 12, no. 2: 39. https://doi.org/10.3390/dj12020039
APA StyleSteinmetz, J., Stemmler, A., Hennig, C.-L., Symmank, J., & Jacobs, C. (2024). GDF15 Contributes to the Regulation of the Mechanosensitive Responses of PdL Fibroblasts through the Modulation of IL-37. Dentistry Journal, 12(2), 39. https://doi.org/10.3390/dj12020039