The Cone Optoretinogram as a Function of Retinal Eccentricity †


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. AOSLO System
2.3. AOSLO Optoretinography Imaging Protocol
2.4. Image Registration
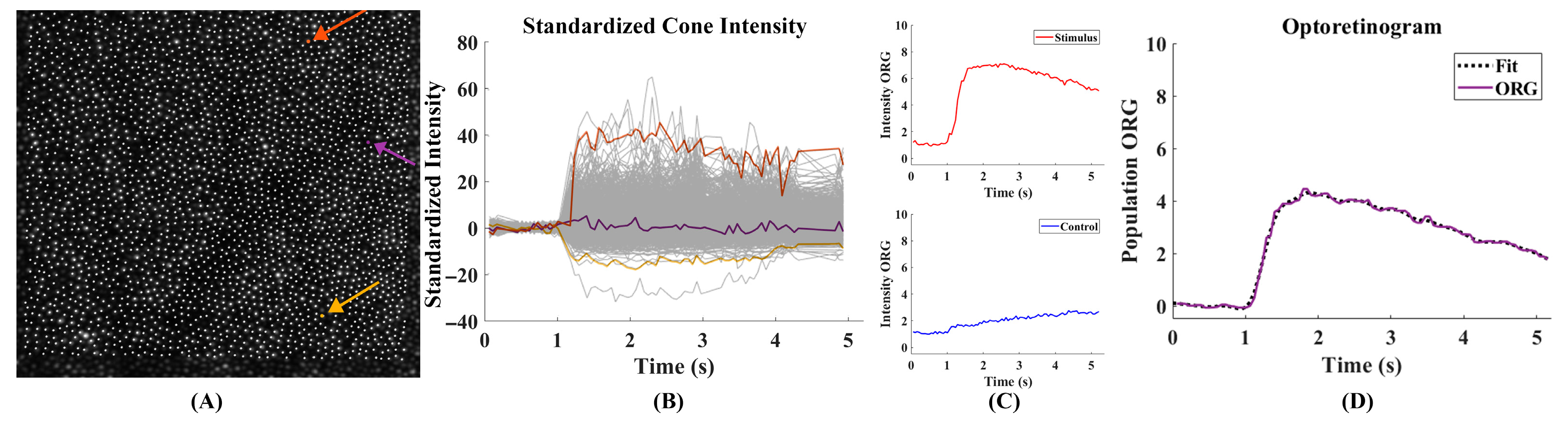
2.5. Intensity ORG Extraction
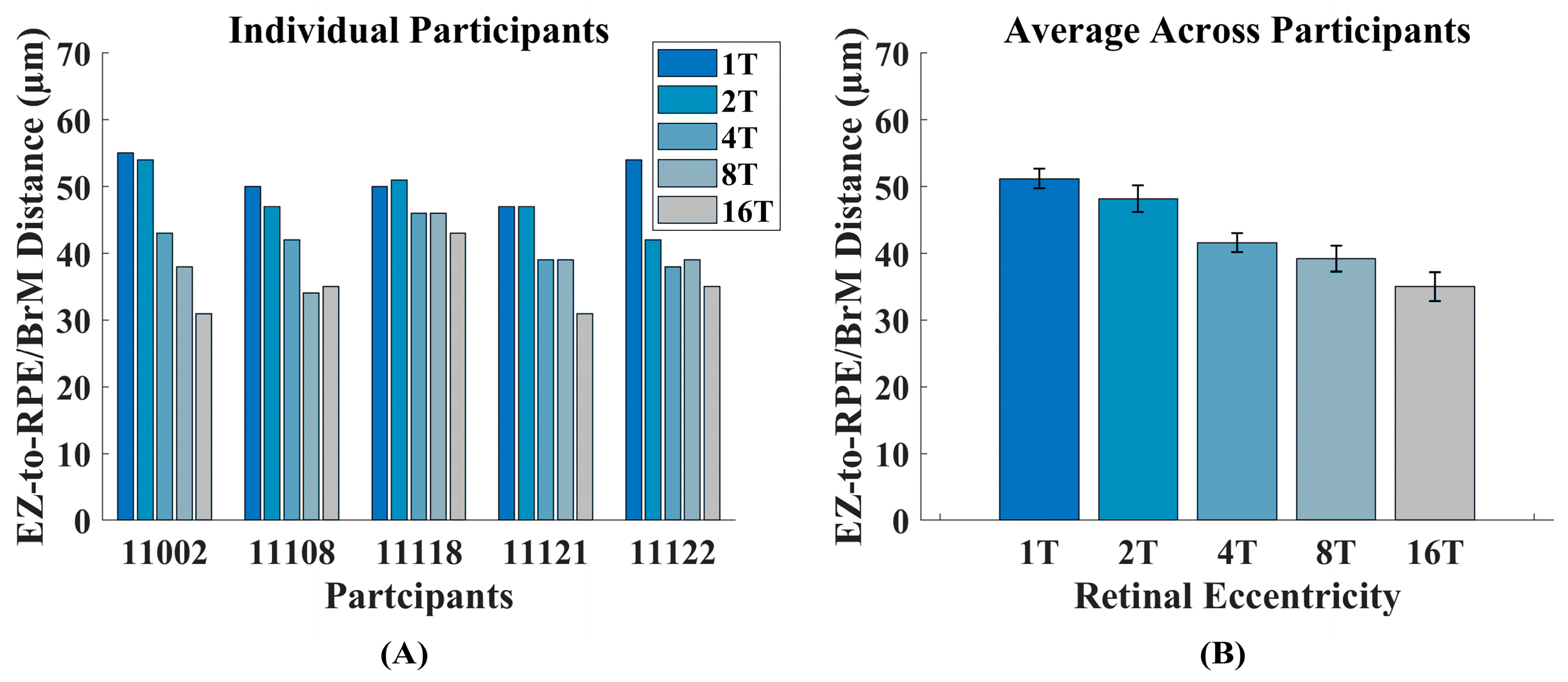
2.6. Measurement of Outer Retinal Thickness
2.7. Statistical Analysis
2.8. Pre-Registration Documentation
3. Results
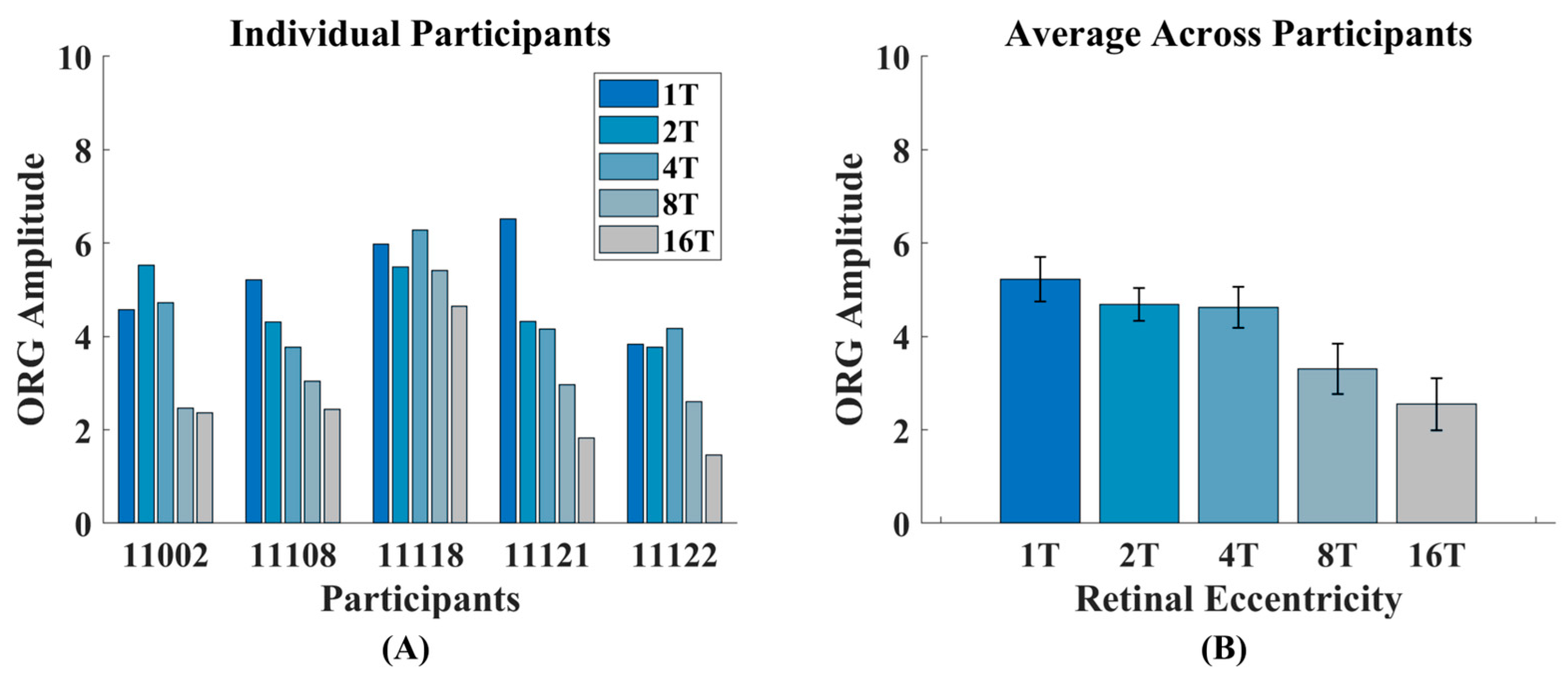
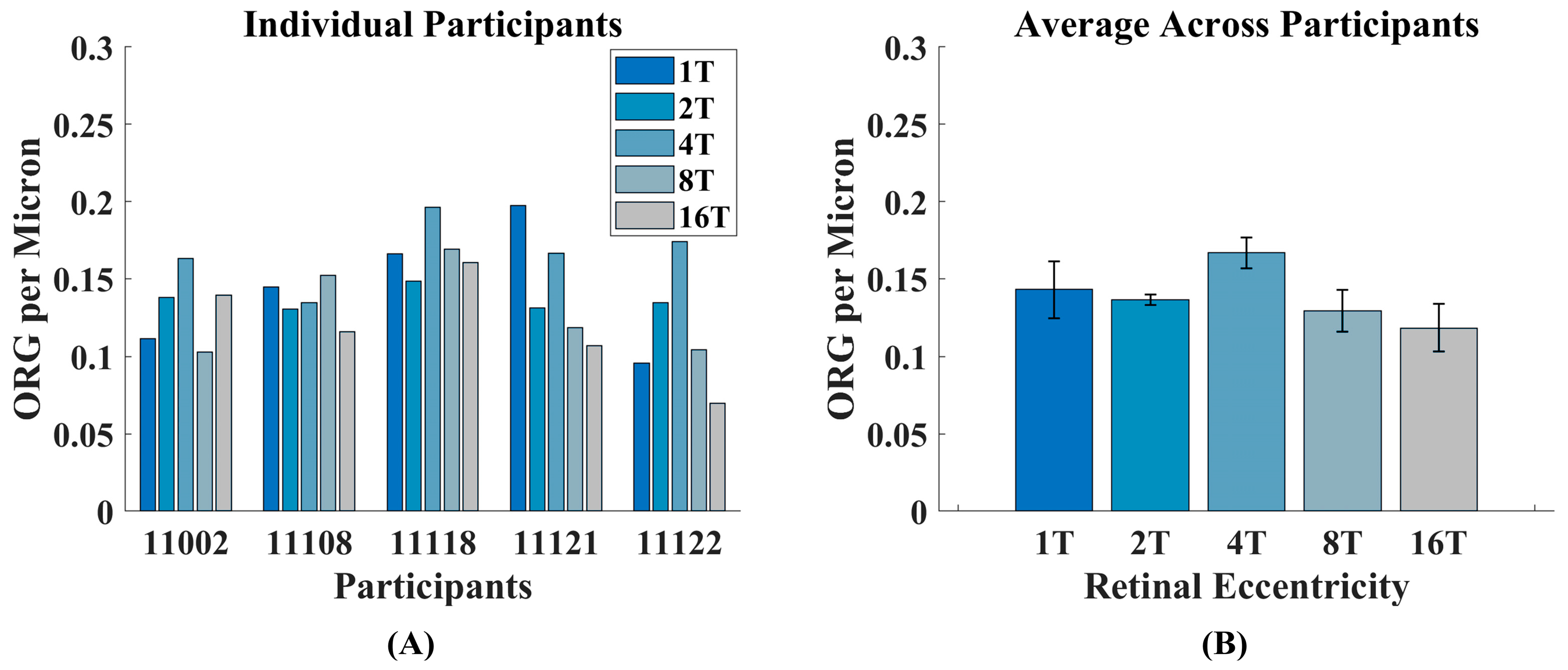
3.1. Intensity ORG Decreases Across Retinal Eccentricity
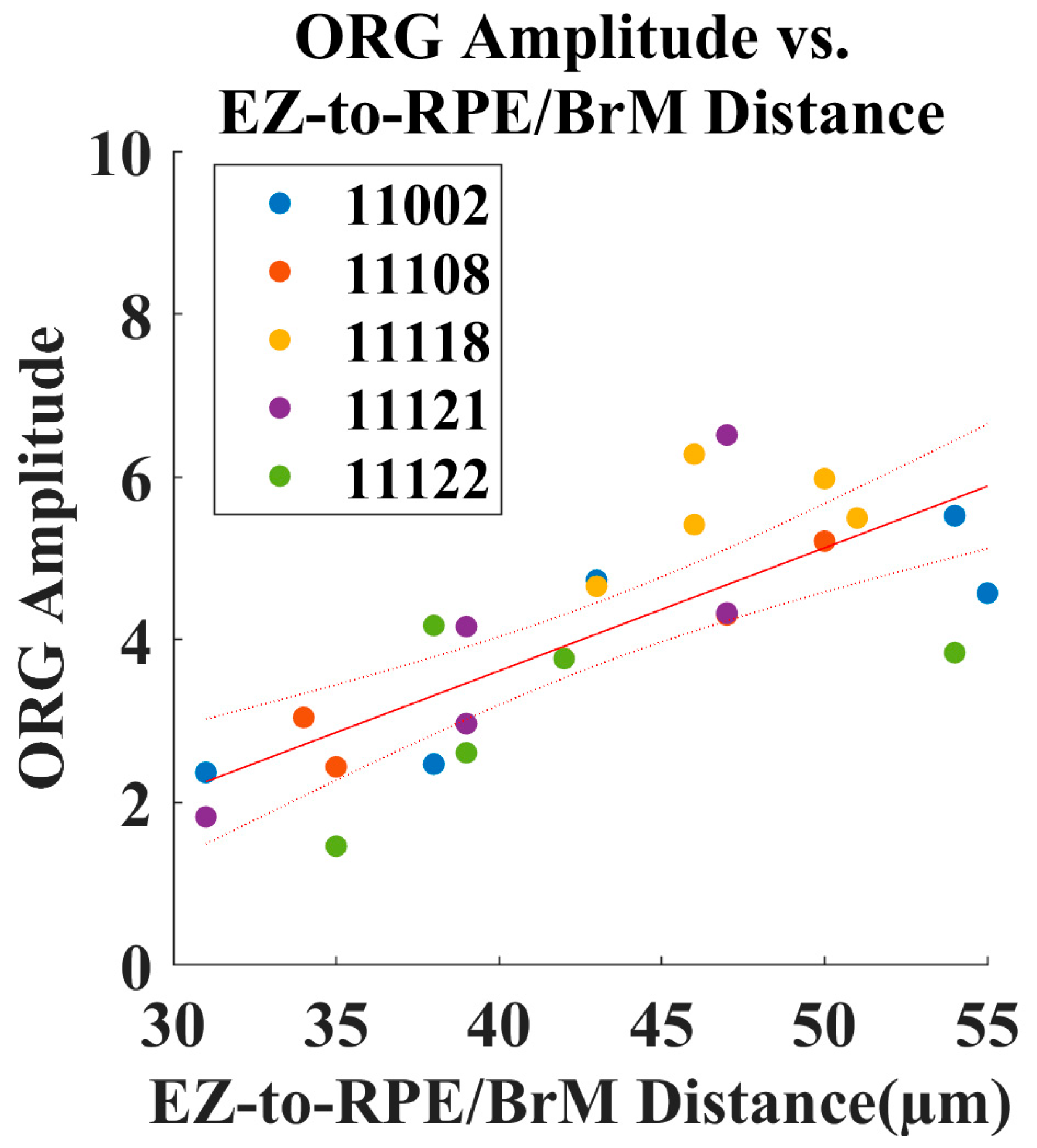
3.2. Increased Outer Retinal Thickness Correlates with ORG Amplitude
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roorda, A.; Duncan, J.L. Adaptive optics ophthalmoscopy. Annu. Rev. Vis. Sci. 2015, 1, 19–50. [Google Scholar] [CrossRef] [PubMed]
- Burns, S.A.; Elsner, A.E.; Sapoznik, K.A.; Warner, R.L.; Gast, T.J. Adaptive optics imaging of the human retina. Prog. Retin. Eye Res. 2019, 68, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.I.W.; Chui, T.Y.P.; Grieve, K. Twenty-five years of clinical applications using adaptive optics ophthalmoscopy [Invited]. Biomed. Opt. Express 2023, 14, 387–428. [Google Scholar] [CrossRef] [PubMed]
- Godara, P.; Dubis, A.M.; Roorda, A.; Duncan, J.L.; Carroll, J. Adaptive optics retinal imaging: Emerging clinical applications. Optom. Vis. Sci. 2010, 87, 930–941. [Google Scholar] [CrossRef]
- Williams, D.R. Imaging single cells in the living retina. Vision. Res. 2011, 51, 1379–1396. [Google Scholar] [CrossRef]
- Jonnal, R.S. Toward a clinical optoretinogram: A review of noninvasive, optical tests of retinal neural function. Ann. Transl. Med. 2021, 9, 1270. [Google Scholar] [CrossRef]
- Karpe, G. The basis of clinical electroretinography. Acta Ophthalmol. Suppl. 1945, 24, 1–118. [Google Scholar]
- Mulligan, J.B.; MacLeod, D.I.A. In search of an optoretinogram. In Vision Science and Its Applications (Technical Digest Series); Optical Society of America: Washington, DC, USA, 1994; Volume 2. [Google Scholar]
- Warner, R.L.; Brainard, D.H.; Morgan, J.I.W. Repeatability and reciprocity of the cone optoretinogram. Biomed. Opt. Express 2022, 13, 6561–6573. [Google Scholar] [CrossRef]
- Pandiyan, V.P.; Maloney-Bertelli, A.; Kuchenbecker, J.A.; Boyle, K.C.; Ling, T.; Chen, Z.C.; Park, B.H.; Roorda, A.; Palanker, D.; Sabesan, R. The optoretinogram reveals the primary steps of phototransduction in the living human eye. Sci. Adv. 2020, 6, eabc1124. [Google Scholar] [CrossRef]
- Zhang, F.; Kurokawa, K.; Lassoued, A.; Crowell, J.A.; Miller, D.T. Cone photoreceptor classification in the living human eye from photostimulation-induced phase dynamics. Proc. Natl. Acad. Sci. USA 2019, 116, 7951–7956. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, T.; Pandiyan, V.P.; Slezak, E.; Sabesan, R. Coarse-scale optoretinography (CoORG) with extended field-of-view for normative characterization. Biomed. Opt. Express 2022, 13, 5989–6002. [Google Scholar] [CrossRef] [PubMed]
- Tomczewski, S.; Curatolo, A.; Foik, A.; Węgrzyn, P.; Bałamut, B.; Wielgo, M.; Kulesza, W.; Galińska, A.; Kordecka, K.; Gulati, S.; et al. Photopic flicker optoretinography captures the light-driven length modulation of photoreceptors during phototransduction. Proc. Natl. Acad. Sci. USA 2025, 122, e2421722122. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Son, T.; Kim, T.-H.; Yao, X. Functional optoretinography: Concurrent OCT monitoring of intrinsic signal amplitude and phase dynamics in human photoreceptors. Biomed. Opt. Express 2021, 12, 2661–2669. [Google Scholar] [CrossRef]
- Azimipour, M.; Valente, D.; Vienola, K.V.; Werner, J.S.; Zawadzki, R.J.; Jonnal, R.S. Optoretinogram: Optical measurement of human cone and rod photoreceptor responses to light. Opt. Lett. 2020, 45, 4658–4661. [Google Scholar] [CrossRef]
- Pandiyan, V.P.; Jiang, X.; Kuchenbecker, J.A.; Sabesan, R. Reflective mirror-based line-scan adaptive optics OCT for imaging retinal structure and function. Biomed. Opt. Express 2021, 12, 5865–5880. [Google Scholar] [CrossRef]
- Pandiyan, V.P.; Schleufer, S.; Slezak, E.; Fong, J.; Upadhyay, R.; Roorda, A.; Ng, R.; Sabesan, R. Characterizing cone spectral classification by optoretinography. Biomed. Opt. Express 2022, 13, 6574–6594. [Google Scholar] [CrossRef] [PubMed]
- Azimipour, M.; Migacz, J.V.; Zawadzki, R.J.; Werner, J.S.; Jonnal, R.S. Functional retinal imaging using adaptive optics swept-source OCT at 1.6 MHz. Optica 2019, 6, 300–303. [Google Scholar] [CrossRef]
- Pandiyan, V.P.; Jiang, X.; Maloney-Bertelli, A.; Kuchenbecker, J.A.; Sharma, U.; Sabesan, R. High-speed adaptive optics line-scan OCT for cellular-resolution optoretinography. Biomed. Opt. Express 2020, 11, 5274–5296. [Google Scholar] [CrossRef]
- Cooper, R.F.; Tuten, W.S.; Dubra, A.; Brainard, D.H.; Morgan, J.I.W. Non-invasive assessment of human cone photoreceptor function. Biomed. Opt. Express 2017, 8, 5098–5112. [Google Scholar] [CrossRef]
- Valente, D.; Vienola, K.V.; Zawadzki, R.J.; Jonnal, R.S. Insight into human photoreceptor function: Modeling optoretinographic responses to diverse stimuli. Sci. Adv. 2025, 11, eadq7332. [Google Scholar] [CrossRef]
- Li, H.; Weiss, C.E.; Pandiyan, V.P.; Nanni, D.; Liu, T.; Kung, P.W.; Tan, B.; Barathi, V.A.; Schmetterer, L.; Sabesan, R.; et al. Optoretinography reveals rapid rod photoreceptor movement upon photoisomerization. bioRxiv 2025. [Google Scholar] [CrossRef]
- Ni, S.; Khan, S.; Jiménez-Villar, A.; Pennesi, M.E.; Huang, D.; Jian, Y.; Chen, S. Optical Assessment of Photoreceptor Function Over the Macula. Transl. Vis. Sci. Technol. 2024, 13, 41. [Google Scholar] [CrossRef]
- Xu, P.; Cooper, R.F.; Jiang, Y.Y.; Morgan, J.I.W. Parafoveal cone function in choroideremia assessed with adaptive optics optoretinography. Sci. Rep. 2024, 14, 8339. [Google Scholar] [CrossRef]
- Gaffney, M.; Connor, T.B.; Cooper, R.F. Intensity-based optoretinography reveals sub-clinical deficits in cone function in retinitis pigmentosa. Front Ophthalmol 2024, 4, 1373549. [Google Scholar] [CrossRef]
- Lassoued, A.; Zhang, F.; Kurokawa, K.; Liu, Y.; Bernucci, M.T.; Crowell, J.A.; Miller, D.T. Cone photoreceptor dysfunction in retinitis pigmentosa revealed by optoretinography. Proc. Natl. Acad. Sci. USA 2021, 118, e2107444118. [Google Scholar] [CrossRef]
- Wendel, B.J.; Pandiyan, V.P.; Liu, T.; Jiang, X.; Lassoued, A.; Slezak, E.; Schleufer, S.; Bharadwaj, P.; Tuten, W.S.; Mustafi, D.; et al. Multimodal High-Resolution Imaging in Retinitis Pigmentosa: A Comparison Between Optoretinography, Cone Density, and Visual Sensitivity. Invest. Ophthalmol. Vis. Sci. 2024, 65, 45. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wendel, B.; Huey, J.; Pandiyan, V.P.; Mustafi, D.; Chao, J.R.; Sabesan, R. Longitudinal changes in optoretinography provide an early and sensitive biomarker of outer retinal disease. medRxiv 2025. [Google Scholar] [CrossRef] [PubMed]
- Curcio, C.A.; Sloan, K.R.; Kalina, R.E.; Hendrickson, A.E. Human photoreceptor topography. J. Comp. Neurol. 1990, 292, 497–523. [Google Scholar] [CrossRef] [PubMed]
- Hoang, Q.V.; Linsenmeier, R.A.; Chung, C.K.; Curcio, C.A. Photoreceptor inner segments in monkey and human retina: Mitochondrial density, optics, and regional variation. Vis. Neurosci. 2002, 19, 395–407. [Google Scholar] [CrossRef]
- Cooper, R.F.; Wilk, M.A.; Tarima, S.; Carroll, J. Evaluating Descriptive Metrics of the Human Cone Mosaic. Invest. Ophthalmol. Vis. Sci. 2016, 57, 2992–3001. [Google Scholar] [CrossRef]
- Warner, R.L.; Xu, P.; Brainard, D.H.; Morgan, J.I.W. Optoretinography as a function of retinal eccentricity. Investig. Ophthalmol. Vis. Sci. 2024, 65, 3405. [Google Scholar]
- Dubra, A.; Sulai, Y. Reflective afocal broadband adaptive optics scanning ophthalmoscope. Biomed. Opt. Express 2011, 2, 1757–1768. [Google Scholar] [CrossRef]
- Scoles, D.; Sulai, Y.N.; Langlo, C.S.; Fishman, G.A.; Curcio, C.A.; Carroll, J.; Dubra, A. In vivo imaging of human cone photoreceptor inner segments. Invest. Ophthalmol. Vis. Sci. 2014, 55, 4244–4251. [Google Scholar] [CrossRef]
- Dubra, A.; Harvey, Z. Registration of 2D Images from Fast Scanning Ophthalmic Instruments. Lect. Notes Comput. Sci. 2010, 6204, 60–71. [Google Scholar]
- Chen, M.; Cooper, R.F.; Gee, J.C.; Brainard, D.H.; Morgan, J.I.W. Automatic longitudinal montaging of adaptive optics retinal images using constellation matching. Biomed. Opt. Express 2019, 10, 6476–6496. [Google Scholar] [CrossRef] [PubMed]
- Garrioch, R.; Langlo, C.; Dubis, A.M.; Cooper, R.F.; Dubra, A.; Carroll, J. Repeatability of in vivo parafoveal cone density and spacing measurements. Optom. Vis. Sci. 2012, 89, 632–643. [Google Scholar] [CrossRef]
- Tam, J.; Martin, J.A.; Roorda, A. Noninvasive visualization and analysis of parafoveal capillaries in humans. Invest. Ophthalmol. Vis. Sci. 2010, 51, 1691–1698. [Google Scholar] [CrossRef]
- Aleman, T.S.; Huckfeldt, R.M.; Serrano, L.W.; Pearson, D.J.; Vergilio, G.K.; McCague, S.; Marshall, K.A.; Ashtari, M.; Doan, T.M.; Weigel-DiFranco, C.A.; et al. AAV2-hCHM Subretinal Delivery to the Macula in Choroideremia: Two Year Interim Results of an Ongoing Phase I/II Gene Therapy Trial. Ophthalmology 2022, 129, 1177–1191. [Google Scholar] [CrossRef]
- Boughman, J.A.; Conneally, P.M.; Nance, W.E. Population genetic studies of retinitis pigmentosa. Am. J. Hum. Genet. 1980, 32, 223–235. [Google Scholar]
- Jacobson, S.G.; Cideciyan, A.V.; Sumaroka, A.; Aleman, T.S.; Schwartz, S.B.; Windsor, E.A.; Roman, A.J.; Stone, E.M.; MacDonald, I.M. Remodeling of the human retina in choroideremia: Rab escort protein 1 (REP-1) mutations. Invest. Ophthalmol. Vis. Sci. 2006, 47, 4113–4120. [Google Scholar] [CrossRef]
- Jonnal, R.S.; Rha, J.; Zhang, Y.; Cense, B.; Gao, W.; Miller, D.T. In vivo functional imaging of human cone photoreceptors. Opt. Express 2007, 15, 16141–16160. [Google Scholar] [CrossRef] [PubMed]
- Meadway, A.; Sincich, L.C. Light reflectivity and interference in cone photoreceptors. Biomed. Opt. Express 2019, 10, 6531–6554. [Google Scholar] [CrossRef] [PubMed]
- Forrester, J.V.; Dick, A.D.; McMenamin, P.G.; Roberts, F.; Pearlman, E. Chapter 1-Anatomy of the eye and orbit. In The Eye, 4th ed.; Forrester, J.V., Dick, A.D., McMenamin, P.G., Roberts, F., Pearlman, E., Eds.; W.B. Saunders: Philadelphia, PA, USA, 2016; pp. 1–102.e2. [Google Scholar]
- Xu, P.; Jiang, Y.Y.; Morgan, J.I.W. Cone Photoreceptor Morphology in Choroideremia Assessed Using Non-Confocal Split-Detection Adaptive Optics Scanning Light Ophthalmoscopy. Invest. Ophthalmol. Vis. Sci. 2023, 64, 36. [Google Scholar] [CrossRef]
- Cooper, R.F.; Aguirre, G.K.; Morgan, J.I.W. Fully Automated Estimation of Spacing and Density for Retinal Mosaics. Transl. Vis. Sci. Technol. 2019, 8, 26. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jung, H.; Dubra, A.; Tam, J. Cone Photoreceptor Cell Segmentation and Diameter Measurement on Adaptive Optics Images Using Circularly Constrained Active Contour Model. Invest. Ophthalmol. Vis. Sci. 2018, 59, 4639–4652. [Google Scholar] [CrossRef]
- Song, H.; Chui, T.Y.; Zhong, Z.; Elsner, A.E.; Burns, S.A. Variation of cone photoreceptor packing density with retinal eccentricity and age. Invest. Ophthalmol. Vis. Sci. 2011, 52, 7376–7384. [Google Scholar] [CrossRef]
- Sawides, L.; de Castro, A.; Burns, S.A. The organization of the cone photoreceptor mosaic measured in the living human retina. Vision. Res. 2017, 132, 34–44. [Google Scholar] [CrossRef]
- Zhang, T.; Godara, P.; Blanco, E.R.; Griffin, R.L.; Wang, X.; Curcio, C.A.; Zhang, Y. Variability in Human Cone Topography Assessed by Adaptive Optics Scanning Laser Ophthalmoscopy. Am. J. Ophthalmol. 2015, 160, 290–300 e291. [Google Scholar] [CrossRef]
- Domdei, N.; Reiniger, J.L.; Holz, F.G.; Harmening, W.M. The Relationship Between Visual Sensitivity and Eccentricity, Cone Density and Outer Segment Length in the Human Foveola. Invest. Ophthalmol. Vis. Sci. 2021, 62, 31. [Google Scholar] [CrossRef]
- Reumueller, A.; Wassermann, L.; Salas, M.; Schranz, M.; Hacker, V.; Mylonas, G.; Sacu, S.; Drexler, W.; Pircher, M.; Schmidt-Erfurth, U.; et al. Three-dimensional composition of the photoreceptor cone layers in healthy eyes using adaptive-optics optical coherence tomography (AO-OCT). PLoS ONE 2021, 16, e0245293. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Warner, R.L.; Xu, P.; Brainard, D.H.; Morgan, J.I.W. The Cone Optoretinogram as a Function of Retinal Eccentricity. Photonics 2025, 12, 676. https://doi.org/10.3390/photonics12070676
Warner RL, Xu P, Brainard DH, Morgan JIW. The Cone Optoretinogram as a Function of Retinal Eccentricity. Photonics. 2025; 12(7):676. https://doi.org/10.3390/photonics12070676
Chicago/Turabian StyleWarner, Raymond L., Peiluo Xu, David H. Brainard, and Jessica I. W. Morgan. 2025. "The Cone Optoretinogram as a Function of Retinal Eccentricity" Photonics 12, no. 7: 676. https://doi.org/10.3390/photonics12070676
APA StyleWarner, R. L., Xu, P., Brainard, D. H., & Morgan, J. I. W. (2025). The Cone Optoretinogram as a Function of Retinal Eccentricity. Photonics, 12(7), 676. https://doi.org/10.3390/photonics12070676