
HPLC-MS, GC and NMR Profiling of Bioactive Lipids of Human Milk and Milk of Dairy Animals (Cow, Sheep, Goat, Buffalo, Camel, Red Deer)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples and Reagents
2.2. Extraction of Lipids
2.3. Analysis of Fatty Acids by Gas Chromatography (GC)
2.4. Analysis of Phospholipids by Nuclear Magnetic Resonance (NMR)
2.5. Analysis of Neutral Glycosphingolipids (Glucosyl and Lactosylceramides, Cerebrosides)
2.6. Analysis of Gangliosides GM3 and GD3 by HPLC-MS
2.7. Statistical Analysis
3. Results and Discussion
3.1. Total Lipids
3.2. Fatty Acids by GC
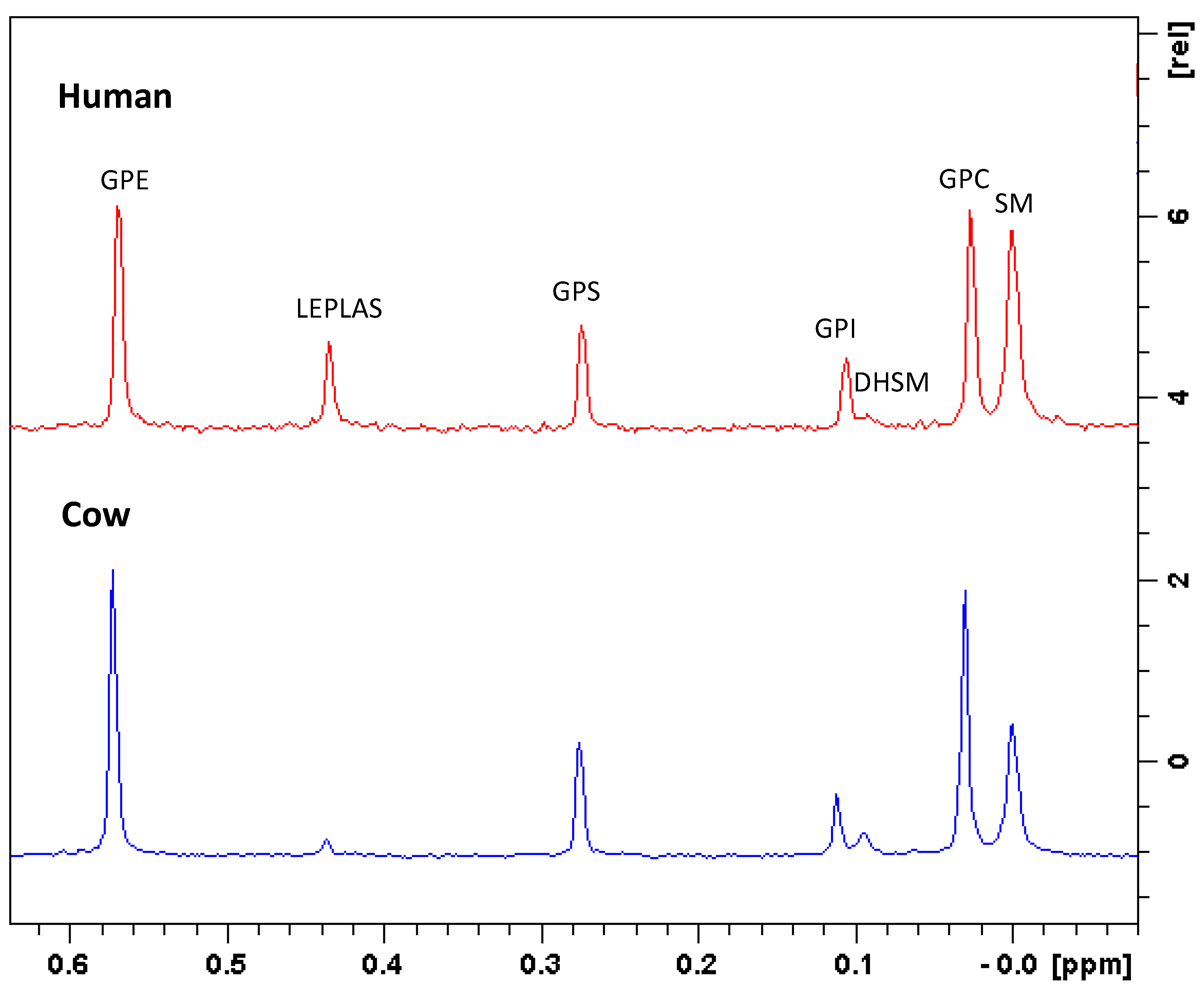
3.3. Phospholipids by 31P-NMR
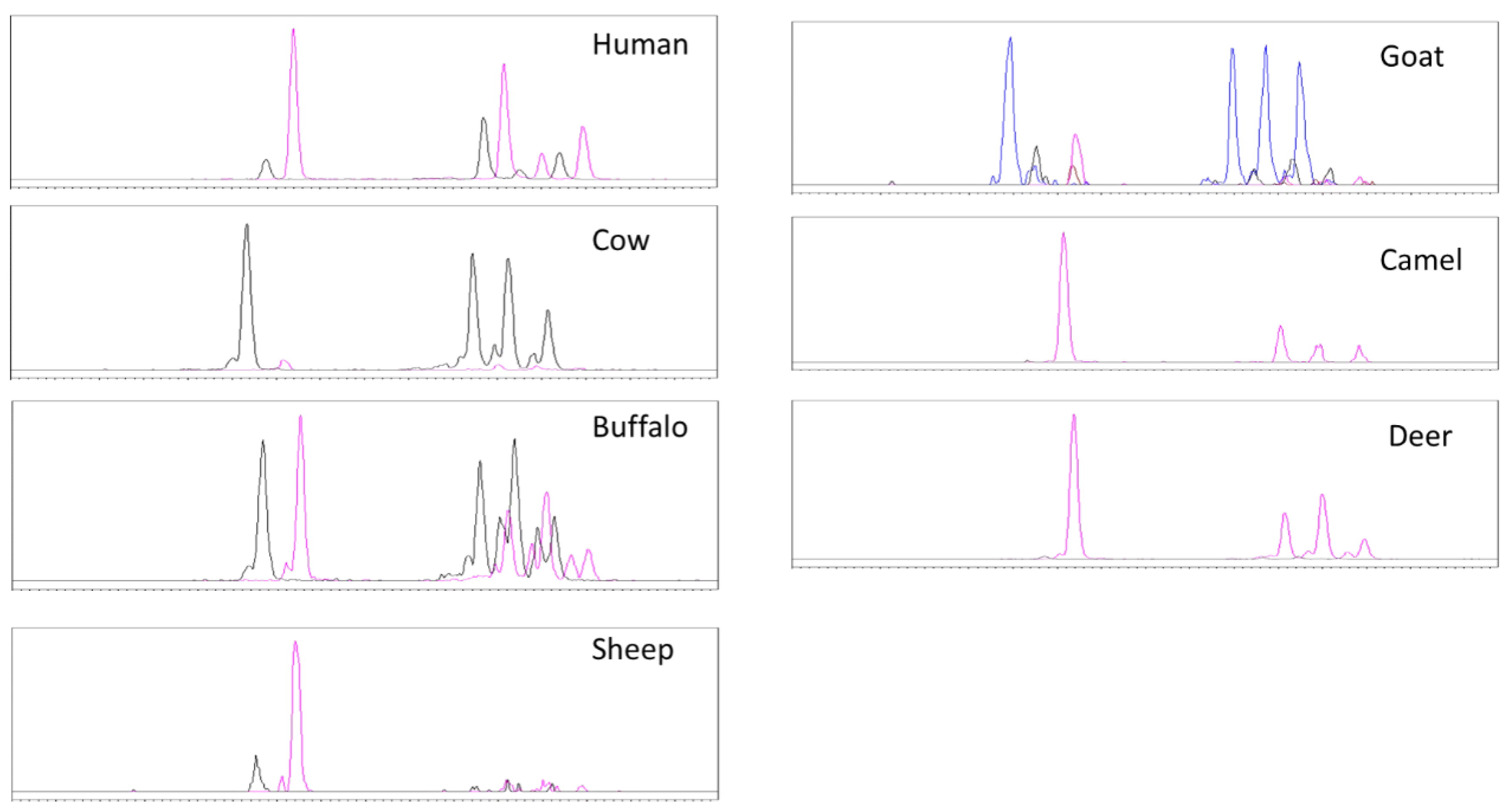
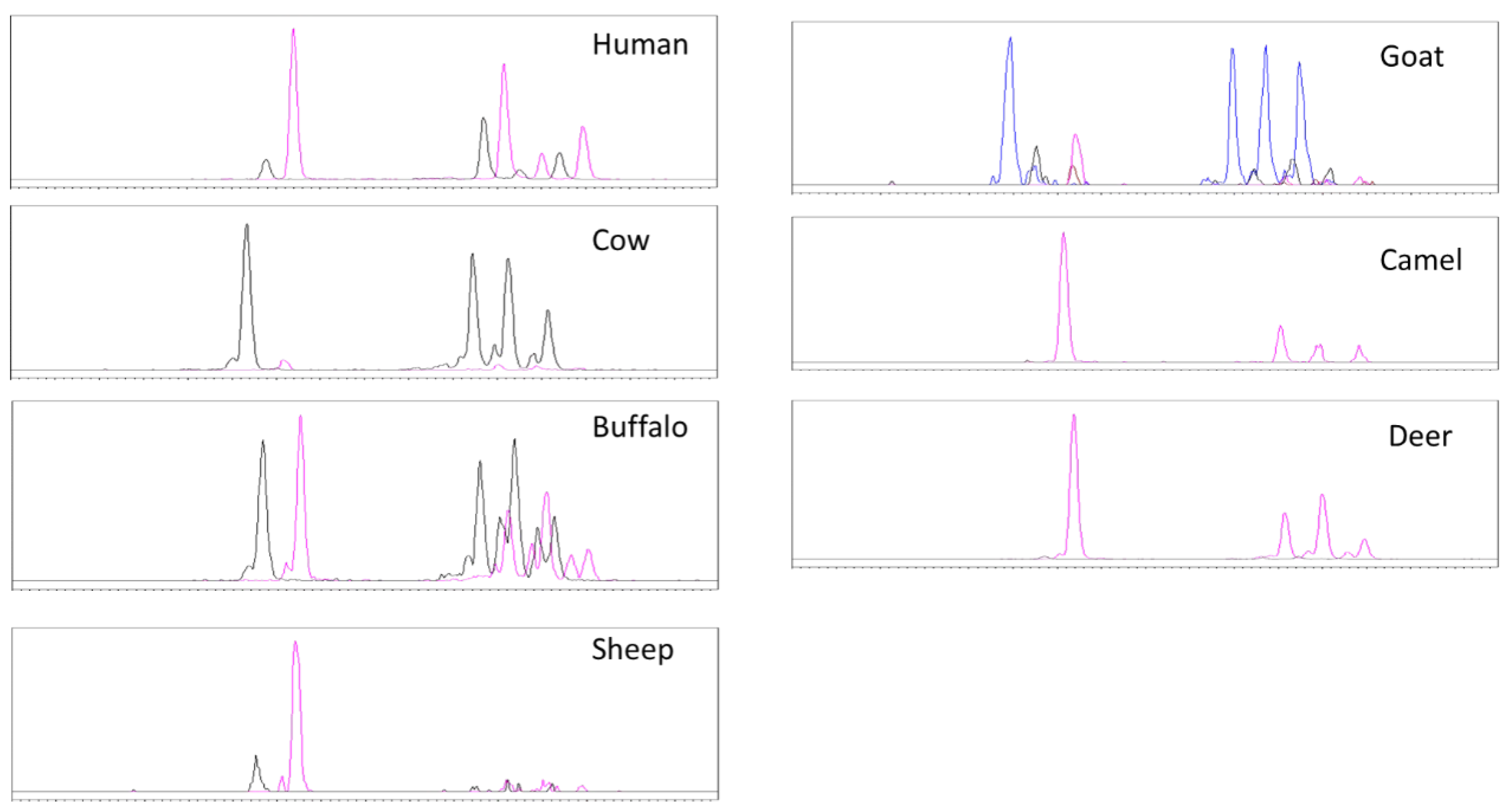
3.4. Neutral Glycosphingolipids by HPLC-MS
3.5. Gangliosides by HPLC-MS
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Jensen, R.G. The Composition of Bovine Milk Lipids: January 1995 to December 2000. J. Dairy Sci. 2002, 85, 295–350. [Google Scholar] [CrossRef]
- Schoenfeld, P.; Wojtczak, L. Short- and medium-chain fatty acids in energy metabolism: The cellular perspective. J. Lipid Res. 2016, 57, 943–954. [Google Scholar] [CrossRef] [Green Version]
- Cunnane, S.C.; Courchesne-Loyer, A.; St-Pierre, V.; Vandenberghe, C.; Pierotti, T.; Fortier, M.; Croteau, E.; Castellano, C. Can ketones compensate for deteriorating brain glucose uptake during aging? Implications for the risk and treatment of Alzheimer’s disease. Ann. N. Y. Acad. Sci. 2016, 1367, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Kim, Y.; Kim, Y.J.; Park, Y. Conjugated Linoleic Acid: Potential Health Benefits as a Functional Food Ingredient. Annu. Rev. Food Sci. Technol. 2016, 7, 221–244. [Google Scholar] [CrossRef]
- Spector, A.A.; Kim, H.-Y. Discovery of essential fatty acids. J. Lipid Res. 2015, 56, 11–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delplanque, B.; Gibson, R.; Koletzko, B.; Lapillonne, A.; Strandvik, B. Lipid Quality in Infant Nutrition. J. Pediatr. Gastroenterol. Nutr. 2015, 61, 8–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koletzko, B.; Agostoni, C.; Carlson, S.; Clandinin, T.; Hornstra, G.; Neuringer, M.; Uauy, R.; Yamashiro, Y.; Willatts, P. Long chain polyunsaturated fatty acids (LC-PUFA) and perinatal development. Acta Paediatr. 2007, 90, 460–464. [Google Scholar] [CrossRef]
- Contarini, G.; Povolo, M. Phospholipids in Milk Fat: Composition, Biological and Technological Significance, and Analytical Strategies. Int. J. Mol. Sci. 2013, 14, 2808–2831. [Google Scholar] [CrossRef] [Green Version]
- Brink, L.R.; Lönnerdal, B. Milk fat globule membrane: The role of its various components in infant health and development. J. Nutr. Biochem. 2020, 85, 108465. [Google Scholar] [CrossRef] [PubMed]
- Palmano, K.; Rowan, A.; Guillermo, R.; Guan, J.; Mc Jarrow, P. The Role of Gangliosides in Neurodevelopment. Nutrients 2015, 7, 3891–3913. [Google Scholar] [CrossRef] [Green Version]
- Muehlhoff, E.; Bennett, A.; McMahon, D. Milk and Dairy Products in Human Nutrition; Food and Agriculture Organisation of the United Nations (FAO): Rome, Italy, 2013. [Google Scholar]
- Vyssotski, M.; Lagutin, K.; Bloor, S.; MacKenzie, A.; Scott, D.; Broadhurst, M.; Day, L.; Samuelsson, L. Lipid profile of New Zealand Sheep Milk. Food New Zealand 2017, 17, 32–33. [Google Scholar]
- Svennerholm, L.; Fredman, P. A procedure for the quantitative isolation of brain gangliosides. Biochim. Biophys. Acta (BBA)—Lipids Lipid Metab. 1980, 617, 97–109. [Google Scholar] [CrossRef]
- Carreau, J.; Dubacq, J. Adaptation of a macro-scale method to the micro-scale for fatty acid methyl transesterification of biological lipid extracts. J. Chromatogr. A 1978, 151, 384–390. [Google Scholar] [CrossRef]
- MacKenzie, A.; Vyssotski, M.; Nekrasov, E. Quantitative Analysis of Dairy Phospholipids by 31P NMR. J. Am. Oil Chem. Soc. 2009, 86, 757–763. [Google Scholar] [CrossRef]
- Clarke, N.G.; Dawson, R.M.C. Alkaline O→N-transacylation. A new method for the quantitative deacylation of phospholipids. Biochem. J. 1981, 195, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Bitman, J.; Wood, D. Changes in Milk Fat Phospholipids During Lactation. J. Dairy Sci. 1990, 73, 1208–1216. [Google Scholar] [CrossRef]
- Le, T.T.; Miocinovic, J.; Nguyen, T.M.; Rombaut, R.; van Camp, J.; Dewettinck, K. Improved Solvent Extraction Procedure and High-Performance Liquid Chromatography–Evaporative Light-Scattering Detector Method for Analysis of Polar Lipids from Dairy Materials. J. Agric. Food Chem. 2011, 59, 10407–10413. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; MacGibbon, A.K.; Mohamed, H.J.B.J.; Loy, S.; Rowan, A.; McJarrow, P.; Fong, B.Y. Determination of ganglioside concentrations in breast milk and serum from Malaysian mothers using a high performance liquid chromatography-mass spectrometry-multiple reaction monitoring method. Int. Dairy J. 2015, 49, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Rivas-Serna, I.M.; Polakowski, R.; Shoemaker, G.K.; Mazurak, V.C.; Clandinin, M.T. Profiling gangliosides from milk products and other biological membranes using LC/MS. J. Food Compos. Anal. 2015, 44, 45–55. [Google Scholar] [CrossRef]
- Yao, Y.; Zhao, G.; Xiang, J.; Zou, X.; Jin, Q.; Wang, X. Lipid composition and structural characteristics of bovine, caprine and human milk fat globules. Int. Dairy J. 2016, 56, 64–73. [Google Scholar] [CrossRef]
- Barłowska, J.; Szwajkowska, M.; Litwińczuk, Z.; Król, J. Nutritional Value and Technological Suitability of Milk from Various Animal Species Used for Dairy Production. Compr. Rev. Food Sci. Food Saf. 2011, 10, 291–302. [Google Scholar] [CrossRef]
- Innis, S.M. Impact of maternal diet on human milk composition and neurological development of infants. Am. J. Clin. Nutr. 2014, 99, 734S–741S. [Google Scholar] [CrossRef] [Green Version]
- Jensen, R.G. Lipids in human milk. Lipids 1999, 34, 1243–1271. [Google Scholar] [CrossRef]
- Aitchison, J.M.; Dunkley, W.L.; Canolty, N.L.; Smith, L.M. Influence of diet on trans fatty acids in human milk. Am. J. Clin. Nutr. 1977, 30, 2006–2015. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Chen, H.; Stanton, C.; Ross, R.P.; Zhang, H.; Chen, Y.Q.; Chen, W. Review of the roles of conjugated linoleic acid in health and disease. J. Funct. Foods 2015, 15, 314–325. [Google Scholar] [CrossRef]
- Bauman, D.; Perfield, J.; Lock, A. Effect of Trans Fatty Acids on Milk Fat and Their Impact on Human Health. In Proceedings of the Southwest Nutrition and Management Conference, Tempe, AZ, USA, 26–27 February 2004; pp. 41–52. [Google Scholar]
- Chilliard, Y.; Ferlay, A.; Mansbridge, R.M.; Doreau, M. Ruminant milk fat plasticity: Nutritional control of saturated, polyunsaturated, trans and conjugated fatty acids. Anim. Res. 2000, 49, 181–205. [Google Scholar] [CrossRef] [Green Version]
- Siurana, A.; Calsamiglia, S. A metaanalysis of feeding strategies to increase the content of conjugated linoleic acid (CLA) in dairy cattle milk and the impact on daily human consumption. Anim. Feed Sci. Technol. 2016, 217, 13–26. [Google Scholar] [CrossRef]
- Decsi, T. Nutritional relevance of trans isomeric fatty acids in human milk. Acta Paediatr. 2003, 92, 1369–1371. [Google Scholar] [CrossRef]
- Bhavsar, N.; St-Onge, M.-P. The diverse nature of saturated fats and the case of medium-chain triglycerides. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 81–87. [Google Scholar] [CrossRef]
- Augustin, K.; Khabbush, A.; Williams, S.; Eaton, S.; Orford, M.; Cross, H.; Heales, S.J.R.; Walker, M.C.; Williams, R.S.B. Mechanisms of action for the medium-chain triglyceride ketogenic diet in neurological and metabolic disorders. Lancet Neurol. 2018, 17, 84–93. [Google Scholar] [CrossRef]
- Garcia, C.; Lutz, N.W.; Confort-Gouny, S.; Cozzone, P.J.; Armand, M.; Bernard, M. Phospholipid fingerprints of milk from different mammalians determined by 31P NMR: Towards specific interest in human health. Food Chem. 2012, 135, 1777–1783. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, S.; Hashimoto, M.; Haque, A.M.; Nakagawa, K.; Kinoshita, M.; Shido, O.; Miyazawa, T. Oral Administration of Ethanolamine Glycerophospholipid Containing a High Level of Plasmalogen Improves Memory Impairment in Amyloid β-Infused Rats. Lipids 2017, 52, 575–585. [Google Scholar] [CrossRef]
- Farah, Z. Composition and characteristics of camel milk. J. Dairy Res. 1993, 60, 603–626. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Alcalá, L.; Fontecha, J. Major lipid classes separation of buttermilk, and cows, goats and ewes milk by high performance liquid chromatography with an evaporative light scattering detector focused on the phospholipid fraction. J. Chromatogr. A 2010, 1217, 3063–3066. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Xu, J.; Xue, C. Sphingolipids in food and their critical roles in human health. Crit. Rev. Food Sci. Nutr. 2020, 61, 462–491. [Google Scholar] [CrossRef]
- Schmelz, E.M.; Zhou, H.; Roberts, P.C. Dietary sphingolipids in colon cancer prevention. In Bioactive Sphingolipids in Cancer Biology and Therapy; Springer: Berlin/Heidelberg, Germany, 2015; pp. 161–188. [Google Scholar]
- Rombaut, R.; Dewettinck, K. Properties, analysis and purification of milk polar lipids. Int. Dairy J. 2006, 16, 1362–1373. [Google Scholar] [CrossRef]
- Bouhours, J.-F.; Bouhours, D. Galactosylceramide is the major cerebroside of human milk fat globule membrane. Biochem. Biophys. Res. Commun. 1979, 88, 1217–1222. [Google Scholar] [CrossRef]
- Newburg, D.S.; Chaturvedi, P. Neutral glycolipids of human and bovine milk. Lipids 1992, 27, 923–927. [Google Scholar] [CrossRef]
- Kuchta-Noctor, A.M.; Murray, B.A.; Stanton, C.; Devery, R.; Kelly, P.M. Anticancer Activity of Buttermilk Against SW480 Colon Cancer Cells is Associated with Caspase-Independent Cell Death and Attenuation of Wnt, Akt, and ERK Signaling. Nutr. Cancer 2016, 68, 1234–1246. [Google Scholar] [CrossRef]
- Kotaro, T.; Masao, I.; Masahiko, M.; Yoshitaka, N. Selective changes in gangliosides of human milk during lactation: A molecular indicator for the period of lactation. Biochim. Biophys. Acta (BBA)—Lipids Lipid Metab. 1986, 879, 73–77. [Google Scholar] [CrossRef]
- Lægreid, A.; Kolstø Otnæss, A.-B.; Fuglesang, J. Human and Bovine Milk: Comparison of Ganglioside Composition and Enterotoxin- Inhibitory Activity. Pediatric Res. 1986, 20, 416–421. [Google Scholar] [CrossRef] [Green Version]
- Simpson, M.A.; Cross, H.; Proukakis, C.; Priestman, D.A.; Neville, D.C.A.; Reinkensmeier, G.; Wang, H.; Wiznitzer, M.; Gurtz, K.; Verganelaki, A.; et al. Infantile-onset symptomatic epilepsy syndrome caused by a homozygous loss-of-function mutation of GM3 synthase. Nat. Genet. 2004, 36, 1225–1229. [Google Scholar] [CrossRef] [Green Version]
- Gallier, S.; Tolenaars, L.; Prosser, C. Whole Goat Milk as a Source of Fat and Milk Fat Globule Membrane in Infant Formula. Nutrients 2020, 12, 3486. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Human | Cow | Buffalo | Sheep | Goat | Camel | Deer | |
|---|---|---|---|---|---|---|---|
| 4:0 | 0.1 ± 0.01 | 2.7 ± 0.20 | 4.2 ± 0.07 | 2.9 ± 0.04 | 2.7 ± 0.13 | 0.1 ± 0.01 | 4.8 ± 0.39 |
| 6:0 | tr. | 1.7 ± 0.18 | 2.1 ± 0.12 | 2.0 ± 0.07 | 3.0 ± 0.07 | 0.1 ± 0.02 | 1.9 ± 0.24 |
| 8:0 | 0.1 ± 0.002 | 1.1 ± 0.04 | 1.0 ± 0.03 | 1.8 ± 0.05 | 3.5 ± 0.10 | 0.1 ± 0.01 | 1.0 ± 0.03 |
| 10:0 | 1.4 ± 0.01 | 3.4 ± 0.03 | 2.6 ± 0.02 | 6.5 ± 0.02 | 12.7 ± 0.01 | 0.2 ± 0.01 | 2.1 ± 0.02 |
| 12:0 | 4.9 ± 0.01 | 4.3 ± 0.01 | 3.2 ± 0.01 | 4.2 ± 0.04 | 5.3 ± 0.02 | 1.2 ± 0.02 | 2.8 ± 0.01 |
| 14:0 | 5.6 ± 0.03 | 12.3 ± 0.08 | 11.2 ± 0.04 | 12.5 ± 0.11 | 10.0 ± 0.03 | 14.4 ± 0.25 | 12.2 ± 0.13 |
| i15:0 | 0.1 ± 0.003 | 0.2 ± 0.002 | 0.5 ± 0.002 | 0.4 ± 0.01 | 0.2 ± 0.004 | 0.5 ± 0.01 | 0.4 ± 0.00 |
| 14:1n-5 | 0.2 ± 0.01 | 1.1 ± 0.02 | 0.8 ± 0.003 | 0.3 ± 0.01 | 0.1 ± 0.01 | 1.4 ± 0.02 | 0.5 ± 0.00 |
| ai15:0 | 0.1 ± 0.004 | 0.5 ± 0.01 | 0.8 ± 0.004 | 0.7 ± 0.01 | 0.5 ± 0.003 | 0.8 ± 0.01 | 0.6 ± 0.01 |
| 15:0 | 0.4 ± 0.01 | 1.1 ± 0.004 | 1.4 ± 0.004 | 1.2 ± 0.01 | 0.9 ± 0.01 | 1.4 ± 0.01 | 1.1 ± 0.02 |
| i16:0 | 0.1 ± 0.002 | 0.2 ± 0.002 | 0.4 ± 0.01 | 0.2 ± 0.002 | 0.2 ± 0.002 | 0.4 ± 0.01 | 0.3 ± 0.002 |
| 16:0 | 21.4 ± 0.06 | 36.0 ± 0.26 | 26.4 ± 0.03 | 26.6 ± 0.12 | 23.4 ± 0.06 | 29.7 ± 0.22 | 25.7 ± 0.46 |
| 16:1n-7 | 2.4 ± 0.02 | 1.7 ± 0.003 | 1.5 ± 0.01 | 1.1 ± 0.01 | 0.3 ± 0.02 | 9.3 ± 0.10 | 1.1 ± 0.03 |
| i17:0 | 0.1 ± 0.01 | 0.3 ± 0.01 | 0.4 ± 0.005 | 0.4 ± 0.01 | 0.3 ± 0.01 | 0.4 ± 0.02 | 0.4 ± 0.01 |
| ai17:0 | 0.2 ± 0.02 | 0.6 ± 0.01 | 0.5 ± 0.002 | 0.7 ± 0.04 | 0.5 ± 0.02 | 0.6 ± 0.02 | 0.4 ± 0.02 |
| 17:0 | 0.3 ± 0.004 | 0.5 ± 0.002 | 0.7 ± 0.01 | 0.5 ± 0.02 | 0.6 ± 0.01 | 0.7 ± 0.002 | 0.6 ± 0.02 |
| 18:0 | 5.6 ± 0.01 | 9.5 ± 0.05 | 10.8 ± 0.02 | 9.1 ± 0.03 | 10.8 ± 0.03 | 13.2 ± 0.01 | 14.9 ± 0.61 |
| 18:1 9t + (6t–8t) | 0.1 ± 0.01 | 0.4 ± 0.01 | 0.5 ± 0.01 | 0.4 ± 0.02 | 0.5 ± 0.01 | 0.8 ± 0.02 | 0.4 ± 0.01 |
| 18:1 10t & 11t | 0.6 ± 0.01 | 2.1 ± 0.005 | 3.7 ± 0.01 | 2.5 ± 0.03 | 2.4 ± 0.004 | 1.3 ± 0.01 | 2.6 ± 0.02 |
| Other 18:1 trans | 0.1 ± 0.01 | 0.3 ± 0.02 | 0.2 ± 0.002 | 0.3 ± 0.02 | 0.6 ± 0.003 | 0.1 ± 0.01 | 0.3 ± 0.01 |
| 18:1 9c | 37.2 ± 0.02 | 15.7 ± 0.06 | 21.6 ± 0.07 | 20.7 ± 0.04 | 17.0 ± 0.05 | 18.6 ± 0.24 | 20.1 ± 0.44 |
| 18:1 11c | 1.9 ± 0.01 | 0.3 ± 0.01 | 0.6 ± 0.01 | 0.4 ± 0.02 | 0.3 ± 0.003 | 1.0 ± 0.005 | 0.7 ± 0.01 |
| 18:1 12c | tr. | 0.1 ± 0.001 | 0.1 ± 0.003 | 0.1 ± 0.01 | 0.2 ± 0.004 | 0.1 ± 0.001 | 0.1 ± 0.00 |
| 18:2n-6 | 12.5 ± 0.04 | 1.2 ± 0.03 | 1.7 ± 0.004 | 1.1 ± 0.06 | 1.3 ± 0.02 | 1.0 ± 0.06 | 1.5 ± 0.04 |
| 18:3n-3 | 1.4 ± 0.003 | 1.0 ± 0.01 | 0.8 ± 0.004 | 0.9 ± 0.01 | 1.4 ± 0.01 | 0.9 ± 0.02 | 1.6 ± 0.05 |
| 20:0 | 0.1 ± 0.01 | 0.1 ± 0.002 | 0.2 ± 0.01 | 0.2 ± 0.01 | 0.1 ± 0.01 | 0.3 ± 0.01 | 0.2 ± 0.01 |
| 9c,11t CLA | 0.4 ± 0.004 | 0.8 ± 0.004 | 1.6 ± 0.01 | 1.7 ± 0.02 | 0.8 ± 0.01 | 0.5 ± 0.02 | 1.1 ± 0.03 |
| 20:1n-9 | 0.4 ± 0.01 | tr. | tr. | tr. | tr. | 0.2 ± 0.01 | 0.1 ± 0.002 |
| 20:2n-6 | 0.2 ± 0.01 | tr. | tr. | tr. | tr. | tr. | tr. |
| 20:3n-6 | 0.4 ± 0.01 | tr. | 0.1 ± 0.0004 | tr. | tr. | tr. | tr. |
| 20:4n-6 | 0.4 ± 0.004 | 0.1 ± 0.001 | 0.1 ± 0.004 | 0.1 ± 0.005 | 0.1 ± 0.01 | 0.1 ± 0.003 | 0.1 ± 0.005 |
| 11c-22:1 | 0.1 ± 0.004 | 0.1 ± 0.002 | tr. | tr. | tr. | tr. | tr. |
| 20:5n-3 | 0.1 ± 0.003 | 0.1 ± 0.002 | 0.1 ± 0.01 | 0.1 ± 0.004 | 0.1 ± 0.003 | 0.1 ± 0.001 | 0.1 ± 0.004 |
| 24:0 | 0.1 ± 0.01 | tr. | 0.1 ± 0.01 | tr. | tr. | 0.1 ± 0.005 | 0.1 ± 0.01 |
| 22:5n-3 | 0.3 ± 0.004 | 0.2 ± 0.01 | 0.1 ± 0.01 | 0.2 ± 0.01 | 0.2 ± 0.003 | 0.2 ± 0.01 | 0.2 ± 0.01 |
| 22:6n-3 | 0.4 ± 0.004 | tr. | tr. | 0.1 ± 0.01 | 0.1 ± 0.002 | tr. | tr. |
| Saturated | 40.5 | 74.7 | 66.3 | 69.8 | 74.6 | 64.1 | 69.5 |
| Medium-chain | 1.5 | 6.3 | 5.7 | 10.3 | 19.2 | 0.3 | 5.0 |
| Total monoenoic | 43.2 | 21.8 | 29.0 | 25.8 | 21.3 | 32.8 | 25.8 |
| cis-monoenoic | 42.3 | 19.0 | 24.6 | 22.7 | 17.9 | 30.6 | 22.5 |
| trans-monoenoic | 0.9 | 2.8 | 4.4 | 3.1 | 3.4 | 2.2 | 3.3 |
| Polyunsaturated | 16.1 | 3.4 | 4.5 | 4.3 | 4.0 | 3.0 | 4.6 |
| Phospholipids * | Human | Cow | Buffalo | Sheep | Goat | Camel | Deer |
|---|---|---|---|---|---|---|---|
| PC | 23.7 ± 0.1 | 26.1 ± 0.5 | 31.1 ± 1.2 | 24.2 ± 1.4 | 25.1 ± 0.2 | 21.1 ± 0.6 | 23.1 ± 0.0 |
| PI | 7.0 ± 0.1 | 7.9 ± 0.7 | 4.6 ± 0.8 | 5.9 ± 0.6 | 7.2 ± 0.4 | 6.8 ± 0.1 | 6.5 ± 0.3 |
| PS | 9.0 ± 0.1 | 12.6 ± 0.9 | 8.7 ± 0.8 | 11.7 ± 1.0 | 12.1 ± 0.8 | 10.7 ± 1.0 | 10.8 ± 0.2 |
| EPLAS | 7.9 ± 0.1 | 1.5 ± 0.7 | 1.9 ± 0.2 | 2.1 ± 0.7 | 3.1 ± 0.3 | 4.6 ± 0.1 | 2.3 ± 0.2 |
| PE | 18.8 ± 0.1 | 22.9 ± 4.2 | 29.9 ± 0.4 | 25.8 ± 1.9 | 24.9 ± 1.2 | 23.5 ± 1.0 | 27.9 ± 0.7 |
| SM | 32.3 ± 0.01 | 24.6 ± 3.1 | 18.4 ± 0.8 | 28.7 ± 0.4 | 26.0 ± 1.5 | 32.6 ± 0.7 | 24.9 ± 1.0 |
| DHSM | 1.3 ± 0.2 | 4.5 ± 0.5 | 4.9 ± 1.1 | 1.6 ± 0.4 | 1.6 ± 0.2 | 0.7 ± 0.1 | 4.4 ± 0.2 |
| % PL w/w in lipid | 0.66 ± 0.1 | 0.61 ± 0.03 | 0.66 ± 0.05 | 0.74 ± 0.1 | 0.87 ± 0.01 | 1.17 ± 0.02 | 0.88 ± 0.01 |
| PL (mg/100g milk) | 24.4 ± 0.5 | 24.4 ± 1.5 | 43.1 ± 2.9 | 50.5 ± 4.0 | 28.9 ± 0.8 | 31.7 ± 0.5 | 65.0 ± 0.0 |
| Hexosylceramides | Lactosylceramide | HexCer/LacCer | |
|---|---|---|---|
| Human | 0.016 ± 0.002 | 0.010 ± 0.002 | 1.6 |
| Cow | 0.034 ± 0.001 | 0.037 ± 0.003 | 0.9 |
| Buffalo | 0.047 ± 0.005 | 0.066 ± 0.008 | 0.7 |
| Sheep | 0.034 ± 0.002 | 0.059 ± 0.002 | 0.6 |
| Goat | 0.058 ± 0.002 | 0.099 ± 0.004 | 0.6 |
| Camel | 0.094 ± 0.003 | 0.103 ± 0.006 | 0.9 |
| Deer | 0.009 ± 0.000 | 0.043 ± 0.002 | 0.2 |
| GD3 | GM3 | GD3 glyc* | GM3/GD3 | |
|---|---|---|---|---|
| Human | 5.25 ± 0.4 | 3.46 ± 0.3 | n.d. | 0.7 |
| Cow | 3.03 ± 0.17 | tr. | n.d. | |
| Buffalo | 0.87 ± 0.01 | 0.12 ± 0.01 | n.d. | 0.1 |
| Sheep | n.d. | n.d. | n.d. | |
| Goat | n.d. | n.d. | 0.30 ± 0.03 | |
| Camel | n.d. | 0.11 ± 0.01 | n.d. | |
| Deer | 0.18 ± 0.02 | 3.43 ± 0.06 | n.d. | 19.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lagutin, K.; MacKenzie, A.; Bloor, S.; Scott, D.; Vyssotski, M. HPLC-MS, GC and NMR Profiling of Bioactive Lipids of Human Milk and Milk of Dairy Animals (Cow, Sheep, Goat, Buffalo, Camel, Red Deer). Separations 2022, 9, 145. https://doi.org/10.3390/separations9060145
Lagutin K, MacKenzie A, Bloor S, Scott D, Vyssotski M. HPLC-MS, GC and NMR Profiling of Bioactive Lipids of Human Milk and Milk of Dairy Animals (Cow, Sheep, Goat, Buffalo, Camel, Red Deer). Separations. 2022; 9(6):145. https://doi.org/10.3390/separations9060145
Chicago/Turabian StyleLagutin, Kirill, Andrew MacKenzie, Stephen Bloor, Dawn Scott, and Mikhail Vyssotski. 2022. "HPLC-MS, GC and NMR Profiling of Bioactive Lipids of Human Milk and Milk of Dairy Animals (Cow, Sheep, Goat, Buffalo, Camel, Red Deer)" Separations 9, no. 6: 145. https://doi.org/10.3390/separations9060145
APA StyleLagutin, K., MacKenzie, A., Bloor, S., Scott, D., & Vyssotski, M. (2022). HPLC-MS, GC and NMR Profiling of Bioactive Lipids of Human Milk and Milk of Dairy Animals (Cow, Sheep, Goat, Buffalo, Camel, Red Deer). Separations, 9(6), 145. https://doi.org/10.3390/separations9060145