
Isolation and Identification of Flavonoids from Black Cumin (Nigella sativa) by HPLC-MS and In Silico Molecular Interactions of Their Major Compounds with Fusarium oxysporum Trypsin-like Serine Protease

 ,
,  , ,
, ,  , , , , ,
, , , , ,
Abstract
1. Introduction
2. Results and Discussion
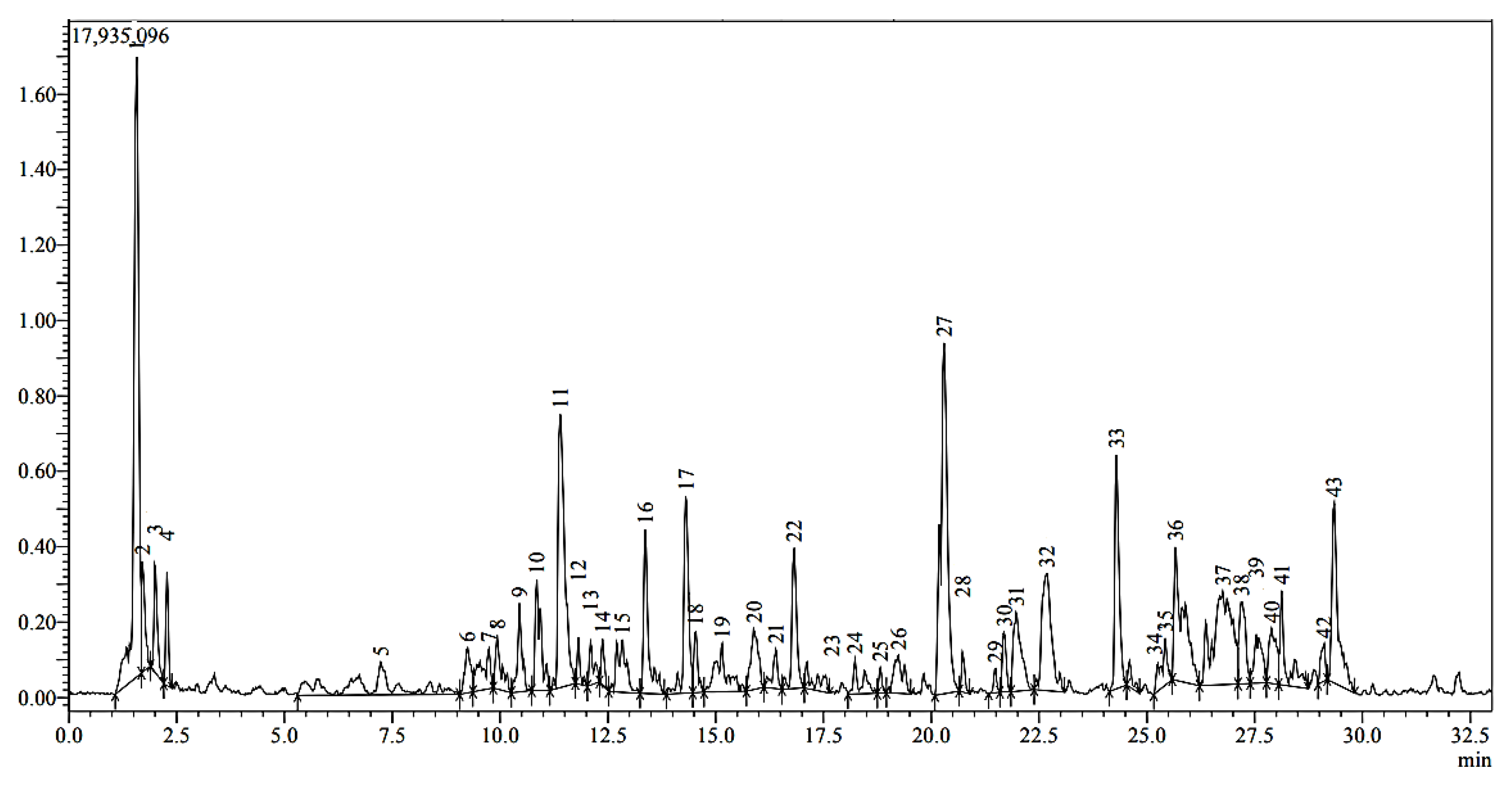
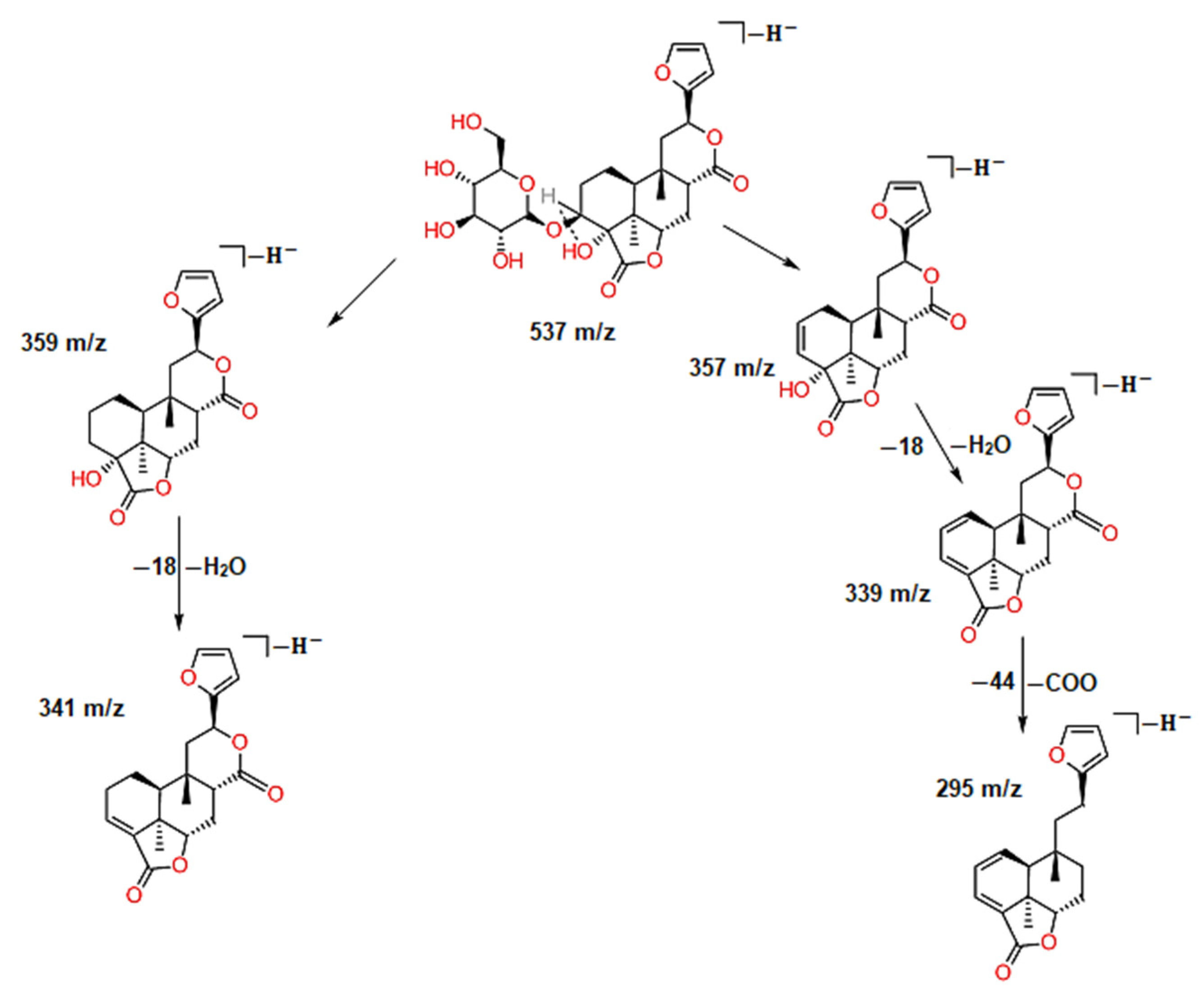
2.1. Analysis of Black Cumin Seeds’ Methanolic Extract by HPLC-MS
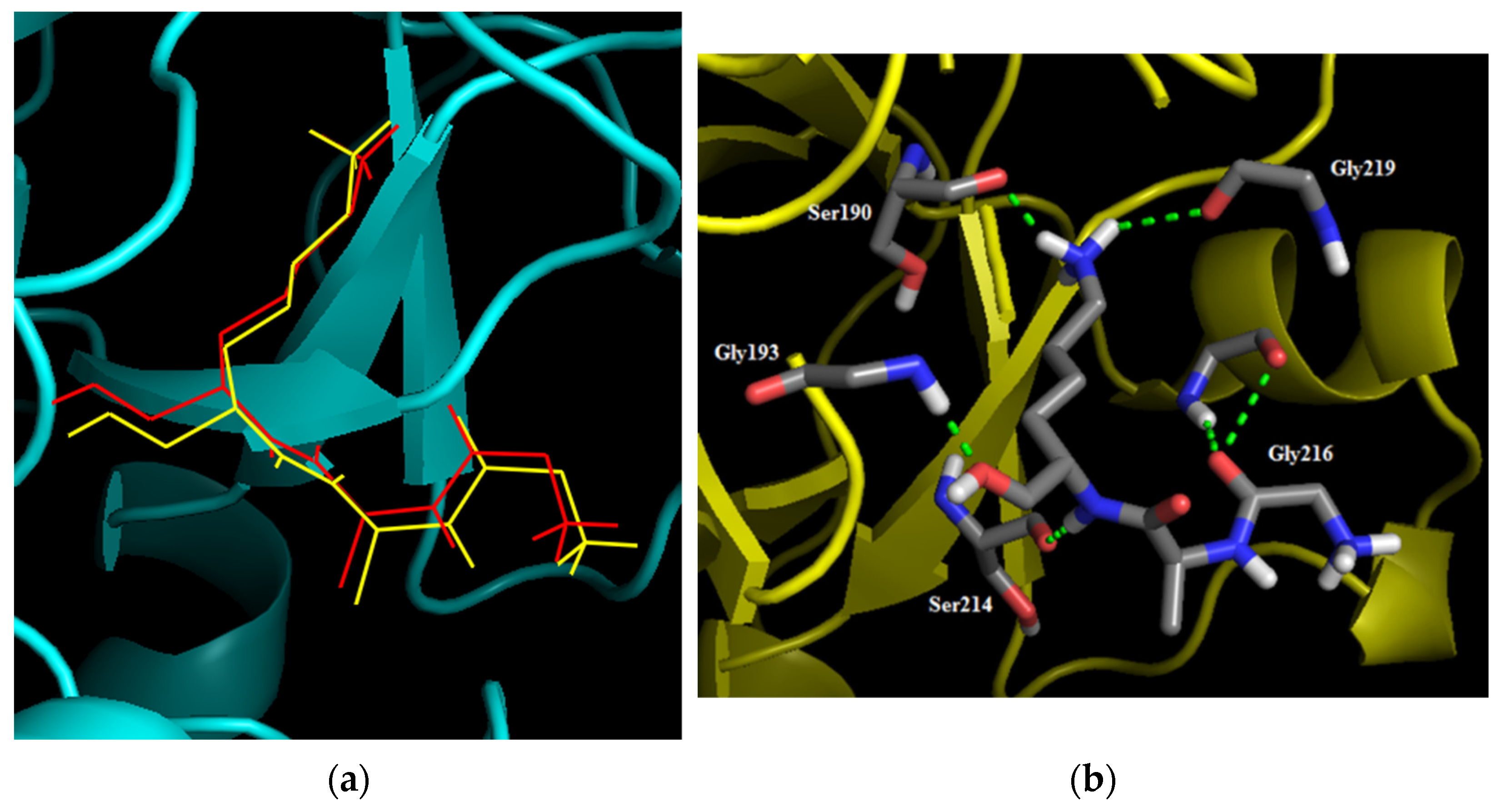
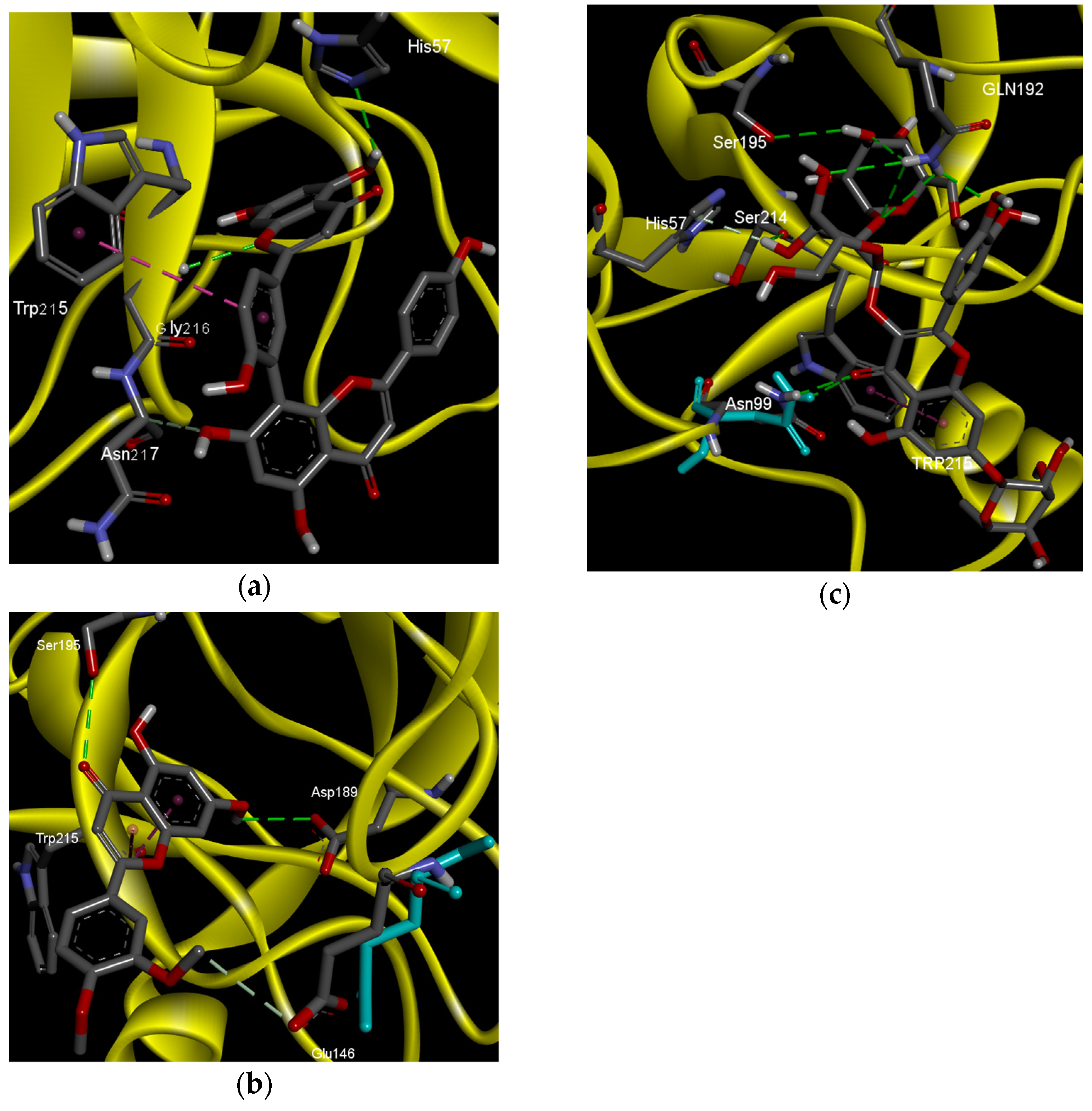
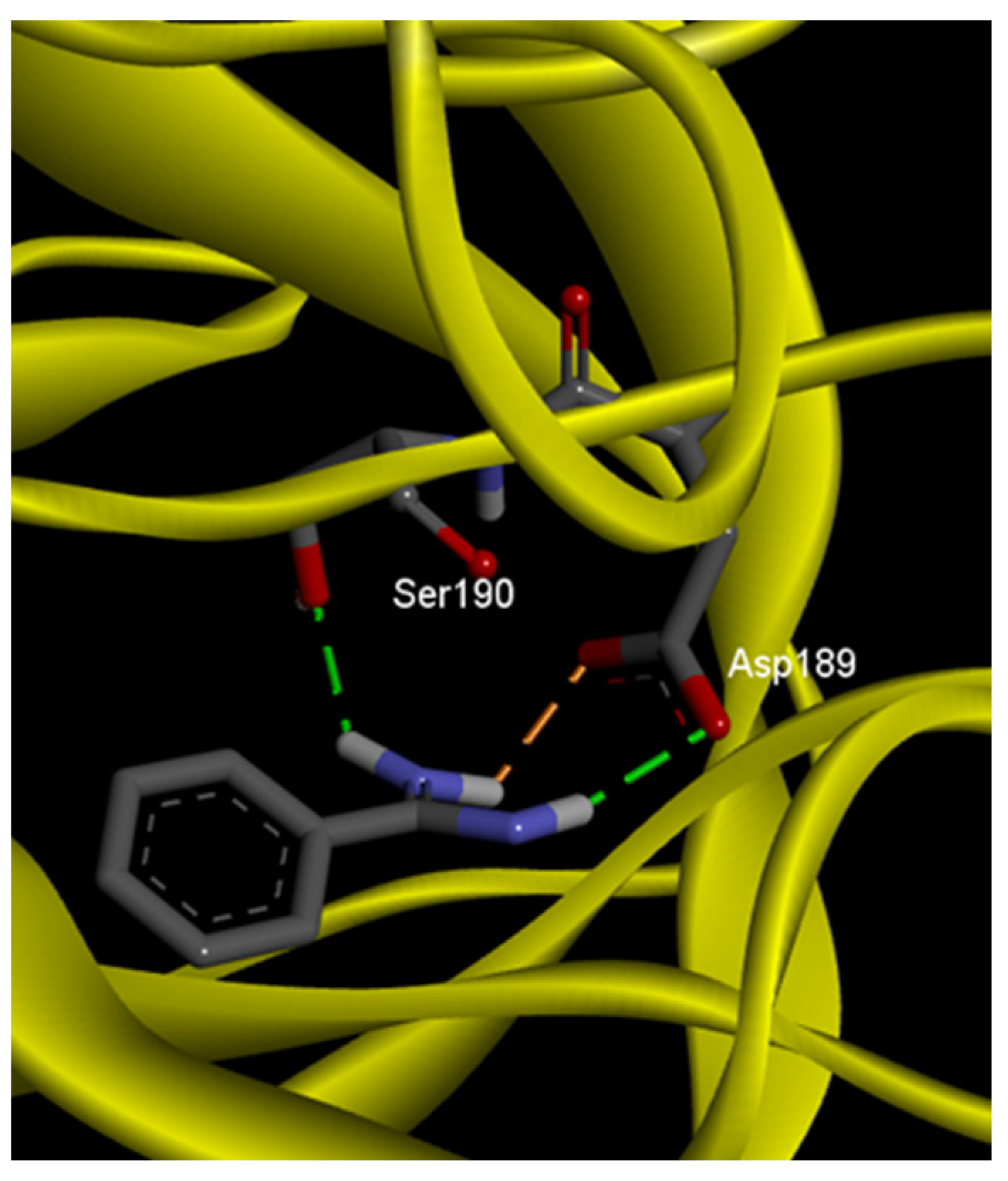
2.2. Molecular Docking Analysis
3. Materials and Methods
3.1. Materials
3.2. Preparation of Extract
3.3. High-Pressure Liquid Chromatography–Mass Spectrometry (HPLC-MS) Analysis
3.4. Ligand Preparation
3.5. Target Protein Preparation
3.6. Molecular Docking Procedures
3.7. Analysis and Visualization of Protein–Ligand Interactions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El-Abyad, M.S.; Abu-Taleb, A.M.; Abdel-Mawgoud, T. Response of host cultivar to cell wall-degrading enzymes of the sugarbeet pathogens Rhizoctonia solani Kühn and Sclerotium rolfsii Sacc. under salinity stress. Microbiol. Res. 1997, 152, 9–17. [Google Scholar] [CrossRef]
- Gvozdeva, E.L.; Volotskaya, A.V.; Sof’in, A.V.; Kudryavtseva, N.N.; Revina, T.A.; Valueva, T.A. Interaction of proteinases secreted by the fungal plant pathogen Rhizoctonia solani with natural proteinase inhibitors produced by plants. Appl. Biochem. Microbiol. 2006, 42, 502–507. [Google Scholar] [CrossRef]
- Ekici, Ö.D.; Paetzel, M.; Dalbey, R.E. Unconventional serine proteases: Variations on the catalytic Ser/His/Asp triad configuration. Protein Sci. 2008, 17, 2023–2037. [Google Scholar] [CrossRef]
- Rypniewski, W.R.; Østergaard, P.R.; Nørregaard-Madsen, M.; Dauter, M.; Wilson, K.S. Fusarium oxysporum trypsin at atomic resolution at 100 and 283 K: A study of ligand binding. Acta Crystallogr. Sect. D Biol. Crystallogr. 2001, 57, 8–19. [Google Scholar] [CrossRef]
- Abd-El-Khair, H.; El-Gamal, N.G. Effects of aqueous extracts of some plant species against Fusarium solani and Rhizoctonia solani in Phaseolus vulgaris plants. Arch. Phytopathol. Plant Prot. 2011, 44, 1–16. [Google Scholar] [CrossRef]
- Marei, G.I.K.; Abdelgaleil, S.A.M. Antifungal potential and biochemical effects of monoterpenes and phenylpropenes on plant. Plant Prot. Sci. 2018, 54, 9–16. [Google Scholar] [CrossRef]
- Hamza, A.; Mohamed, A.; Derbalah, A. Unconventional alternatives for control of tomato root rot caused by Rhizoctonia solani under greenhouse conditions. J. Plant Prot. Res. 2016, 56, 298–305. [Google Scholar] [CrossRef]
- Aftab, A.; Yousaf, Y.; Javaid, A.; Riaz, N.; Younas, A.; Rashid, M.; Shamsher, B.; Arif, A. Antifungal activity of vegetative methanolic extracts of Nigella sativa against Fusarium oxysporum and Macrophomina phaseolina and its phytochemical profiling by GC-MS analysis. Int. J. Agric. Biol. 2019, 21, 569–576. [Google Scholar]
- Kadam, D.; Lele, S.S. Extraction, characterization and bioactive properties of Nigella sativa seedcake. J. Food Sci. Technol. 2017, 54, 3936–3947. [Google Scholar] [CrossRef] [PubMed]
- Soleimanifar, M.; Niazmand, R.; Jafari, S.M. Evaluation of oxidative stability, fatty acid profile, and antioxidant properties of black cumin seed oil and extract. J. Food Meas. Charact. 2019, 13, 383–389. [Google Scholar] [CrossRef]
- Muthu Kumara, S.S.; Kwong Huat, B.T. Extraction, Isolation and Characterisation of Antitumor Principle, α-Hederin, from the Seeds of Nigella sativa. Planta Med. 2001, 67, 29–32. [Google Scholar] [CrossRef]
- Ahmad, A.; Husain, A.; Mujeeb, M.; Alam Khan, S.; Najmi, A.K.; Siddique, N.A.; Damanhouri, Z.A.; Anwar, F. A review on therapeutic potential of Nigella sativa: A miracle herb. Asian Pac. J. Trop. Biomed. 2013, 3, 337–352. [Google Scholar] [CrossRef]
- Chaieb, K.; Kouidhi, B.; Jrah, H.; Mahdouani, K.; Bakhrouf, A. Antibacterial activity of Thymoquinone, an active principle of Nigella sativa and its potency to prevent bacterial biofilm formation. BMC Complement. Altern. Med. 2011, 11, 29. [Google Scholar] [CrossRef]
- Bordoni, L.; Fedeli, D.; Nasuti, C.; Maggi, F.; Papa, F.; Wabitsch, M.; De Caterina, R.; Gabbianelli, R. Antioxidant and Anti-Inflammatory Properties of Nigella sativa Oil in Human Pre-Adipocytes. Antioxidants 2019, 8, 51. [Google Scholar] [CrossRef]
- El Aziz, S.M.S.A.; Abo-Shady, A.; Ibrahim, M.; Helmy, M.M. Inhibition of Rhizoctonia solani Growth and Its Extracellular Hydrolytic Enzymes by Different Extracts of Cinnamon (Cinnamomum cassia) and Black Cumin Seeds (Nigella sativa). Arab. Univ. J. Agric. Sci. 2022, 30, 1–18. [Google Scholar] [CrossRef]
- Li, H.; DaSilva, N.A.; Liu, W.; Xu, J.; Dombi, G.W.; Dain, J.A.; Li, D.; Chamcheu, J.C.; Seeram, N.P.; Ma, H. Thymocid®, a Standardized Black Cumin (Nigella sativa) Seed Extract, Modulates Collagen Cross-Linking, Collagenase and Elastase Activities, and Melanogenesis in Murine B16F10 Melanoma Cells. Nutrients 2020, 12, 2146. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Marguerita, L.-W. Molecular docking. In Molecular Modeling of Proteins, 1st ed.; Kukol, A., Ed.; Humana: Totowa, NJ, USA, 2008; pp. 365–382. [Google Scholar] [CrossRef]
- Yao, H.; Chen, B.; Zhang, Y.; Ou, H.; Li, Y.; Li, S.; Shi, P.; Lin, X. Analysis of the Total Biflavonoids Extract from Selaginella doederleinii by HPLC-QTOF-MS and Its In Vitro and In Vivo Anticancer Effects. Molecules 2017, 22, 325. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, H.; Takahashi, S.; Takeda, S.; Hatta, H. Distribution of Flavan-3-ol Species in Ripe Strawberry Fruit Revealed by Matrix-Assisted Laser Desorption/Ionization-Mass Spectrometry Imaging. Molecules 2019, 25, 103. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Nakabayashi, R.; Mori, T.; Yamada, Y.; Takahashi, M.; Rai, A.; Sugiyama, R.; Yamamoto, H.; Nakaya, T.; Yamazaki, M.; et al. A cheminformatics approach to characterize metabolomes in stable-isotope-labeled organisms. Nat. Methods 2019, 16, 295–298. [Google Scholar] [CrossRef]
- Fabre, N.; Rustan, I.; de Hoffmann, E.; Quetin-Leclercq, J. Determination of flavone, flavonol, and flavanone aglycones by negative ion liquid chromatography electrospray ion trap mass spectrometry. J. Am. Soc. Mass Spectrom. 2001, 12, 707–715. [Google Scholar] [CrossRef]
- Lewars, E.G.; March, R.E. Fragmentation of 3-hydroxyflavone; a computational and mass spectrometric study. Rapid Commun. Mass Spectrom. 2007, 21, 1669–1679. [Google Scholar] [CrossRef]
- Benayad, Z.; Gómez-Cordovés, C.; Es-Safi, N.E. Characterization of Flavonoid Glycosides from Fenugreek (Trigonella foenum-graecum) Crude Seeds by HPLC–DAD–ESI/MS Analysis. Int. J. Mol. Sci. 2014, 15, 20668–20685. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-H.; Guo, H.; Xu, W.-B.; Ge, J.; Li, X.; Alimu, M.; He, D.-J. Rapid Identification of Flavonoid Constituents Directly from PTP1B Inhibitive Extract of Raspberry (Rubus idaeus L.) Leaves by HPLC–ESI–QTOF–MS-MS. J. Chromatogr. Sci. 2016, 54, 805–810. [Google Scholar] [CrossRef]
- Singh, A.; Bajpai, V.; Kumar, S.; Sharma, K.R.; Kumar, B. Profiling of Gallic and Ellagic Acid Derivatives in Different Plant Parts of Terminalia arjuna by HPLC-ESI-QTOF-MS/MS. Nat. Prod. Commun. 2016, 11, 239–244. [Google Scholar] [CrossRef]
- Brito, A.; Ramirez, J.E.; Areche, C.; Sepúlveda, B.; Simirgiotis, M.J. HPLC-UV-MS Profiles of Phenolic Compounds and Antioxidant Activity of Fruits from Three Citrus Species Consumed in Northern Chile. Molecules 2014, 19, 17400–17421. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, Y.M.; Abreu, L.S.; Lima, R.L.; Costa, V.C.O.; de Melo, J.I.M.; Braz-Filho, R.; Silva, M.S.; Tavares, J.F. Rapid Characterization of Triterpene Saponins from Zornia brasiliensis by HPLC-ESI-MS/MS. Molecules 2019, 24, 2519. [Google Scholar] [CrossRef]
- Wang, H.-Y.; Liu, K.; Wang, R.-X.; Qin, S.-H.; Wang, F.-L.; Sun, J.-Y. Two new triterpenoids from Nauclea officinalis. Nat. Prod. Res. 2015, 29, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Mad, T.; Sterk, H.; Mittelbach, M.; Rechberger, G.N. Tandem mass spectrometric analysis of a complex triterpene saponin mixture of Chenopodium quinoa. J. Am. Soc. Mass Spectrom. 2006, 17, 795–806. [Google Scholar] [CrossRef]
- Xu, L.-L.; Guo, F.-X.; Chi, S.-S.; Wang, Z.-J.; Jiang, Y.-Y.; Liu, B.; Zhang, J.-Y. Rapid Screening and Identification of Diterpenoids in Tinospora sinensis Based on High-Performance Liquid Chromatography Coupled with Linear Ion Trap-Orbitrap Mass Spectrometry. Molecules 2017, 22, 912. [Google Scholar] [CrossRef]
- Hassan, W.H.B.; Abdelaziz, S.; Al Yousef, H.M. Chemical Composition and Biological Activities of the Aqueous Fraction of Parkinsonea aculeata L. Growing in Saudi Arabia. Arab. J. Chem. 2019, 12, 377–387. [Google Scholar] [CrossRef]
- Fischer, U.A.; Carle, R.; Kammerer, D.R. Identification and quantification of phenolic compounds from pomegranate (Punica granatum L.) peel, mesocarp, aril and differently produced juices by HPLC-DAD–ESI/MSn. Food Chem. 2011, 127, 807–821. [Google Scholar] [CrossRef]
- Hsieh, Y.-J.; Lin, L.-C.; Tsai, T.-H. Determination and identification of plumbagin from the roots of Plumbago zeylanica L. by liquid chromatography with tandem mass spectrometry. J. Chromatogr. A 2005, 1083, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Steinmetzer, T.; Hardes, K. The antiviral potential of host protease inhibitors, in Activation of Viruses by Host Proteases. Act. Viruses Host Proteases 2018, 16, 279–325. [Google Scholar] [CrossRef]
- Selim, S.; Almuhayawi, M.S.; Alharbi, M.T.; Al Jaouni, S.K.; Alharthi, A.; Abdel-Wahab, B.A.; Ibrahim, M.A.R.; Alsuhaibani, A.M.; Warrad, M.; Rashed, K. Insights into the Antimicrobial, Antioxidant, Anti-SARS-CoV-2 and Cytotoxic Activities of Pistacia lentiscus Bark and Phytochemical Profile; In Silico and In Vitro Study. Antioxidants 2022, 11, 930. [Google Scholar] [CrossRef]
- Pan, X.; Tan, N.; Zeng, G.; Zhang, Y.; Jia, R. Amentoflavone and its derivatives as novel natural inhibitors of human Cathepsin B. Bioorgan. Med. Chem. 2005, 13, 5819–5825. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.-H.; Chen, T.-R.; Fang, H.-B.; Jin, Q.; Zhang, S.-J.; Hou, J.; Yu, Y.; Dou, T.-Y.; Cao, Y.-F.; Guo, W.-Z.; et al. Natural constituents of St. John’s Wort inhibit the proteolytic activity of human thrombin. Int. J. Biol. Macromol. 2019, 134, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.A.; Hossain, R.; Siyadatpanah, A.; Al-Khafaji, K.; Khalipha, A.B.R.; Dey, D.; Asha, U.H.; Biswas, P.; Saikat, A.S.M.; Chenari, H.A.; et al. Diterpenes/Diterpenoids and Their Derivatives as Potential Bioactive Leads against Dengue Virus: A Computational and Network Pharmacology Study. Molecules 2021, 26, 6821. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, A.; Haque, M. Preparation of medicinal plants: Basic extraction and fractionation procedures for experimental purposes. J. Pharm. Bioallied Sci. 2020, 12, 1–10. [Google Scholar] [CrossRef]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | tR (min) | [M–H]− (m/z) | ESI-MS (m/z) | Formula | Identification | % Area |
|---|---|---|---|---|---|---|
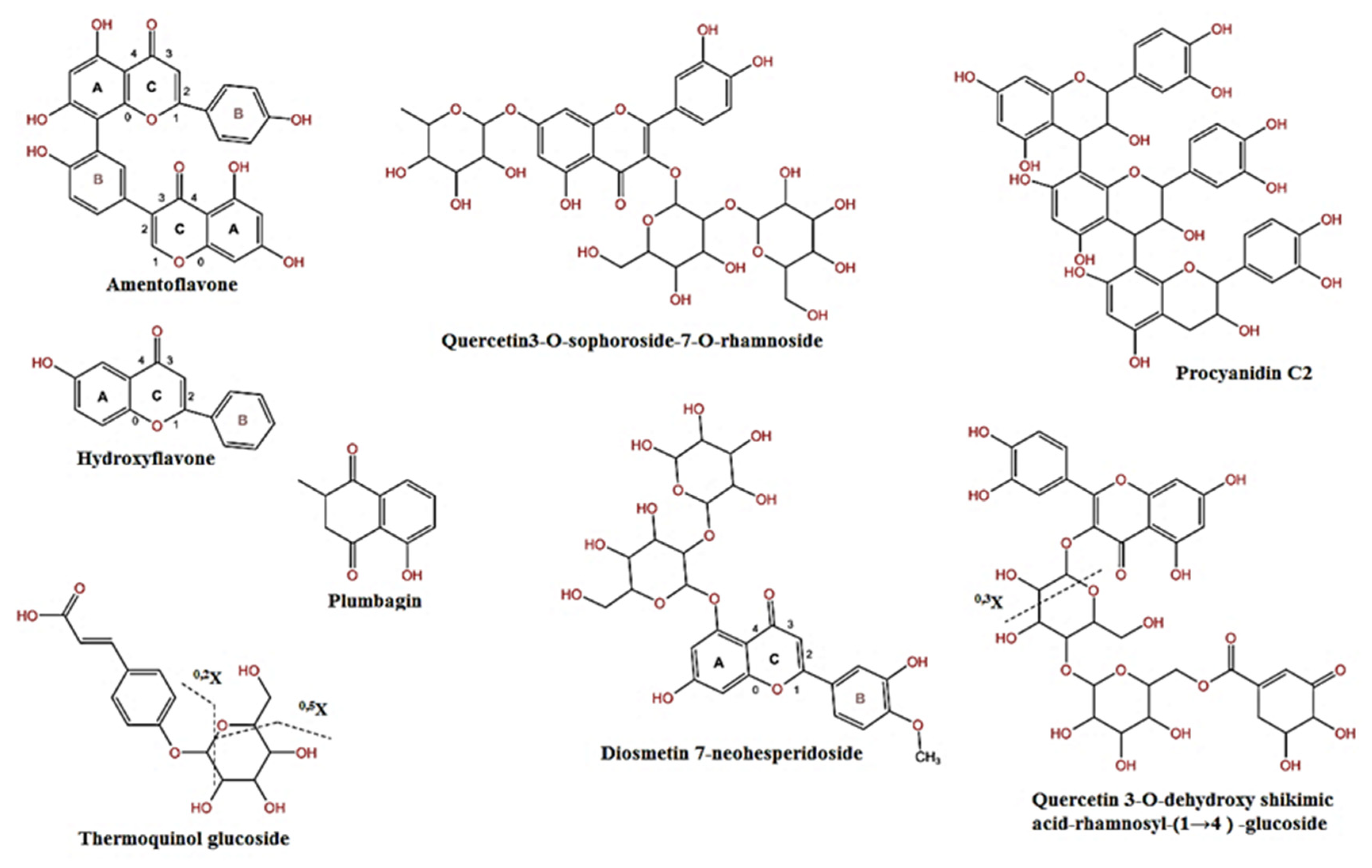
| 1 | 1.57 | 537 | 387, 357, 195 | C30H18O10 | Amentoflavone | 36.8 |
| 4 | 2.28 | 237 | 191, 128 | C15H10O3 | Hydroxyflavone | 0.6 |
| 7 | 9.74 | 779 | 609, 497, 327, 171 | C34H36O21 | Quercetin 3-O-dehydroxy shikimic acid-rhamnosyl-(1)-4→glucoside | 0.3 |
| 8 | 9.93 | 325 | 266, 175, 137 | C15H18O8 | Thermoquinol glucoside | 0.3 |
| 9 | 10.45 | 933 | 535, 489, 239 | C41H26O26 | castalagin | 0.6 |
| 10 | 10.85 | 917 | 431 | C42H46O23 | Flavonol base + 4O, O-dHex-Hex-Hex-Coumaroyl | 1.4 |
| 11 | 11.4 | 771 | C33H40O21 | Quercetin3-O-sophoroside-7-O-rhamnoside | 9.9 | |
| 12 | 11.82 | 607 | 543, 461, 415, 149 | C28H32O15 | Diosmetin 7-neohesperidoside | 0.1 |
| 13 | 12.1 | 765 | 439, 407, 371, 321, 233 | C41H66O13 | Soyasaponin IV | 0.2 |
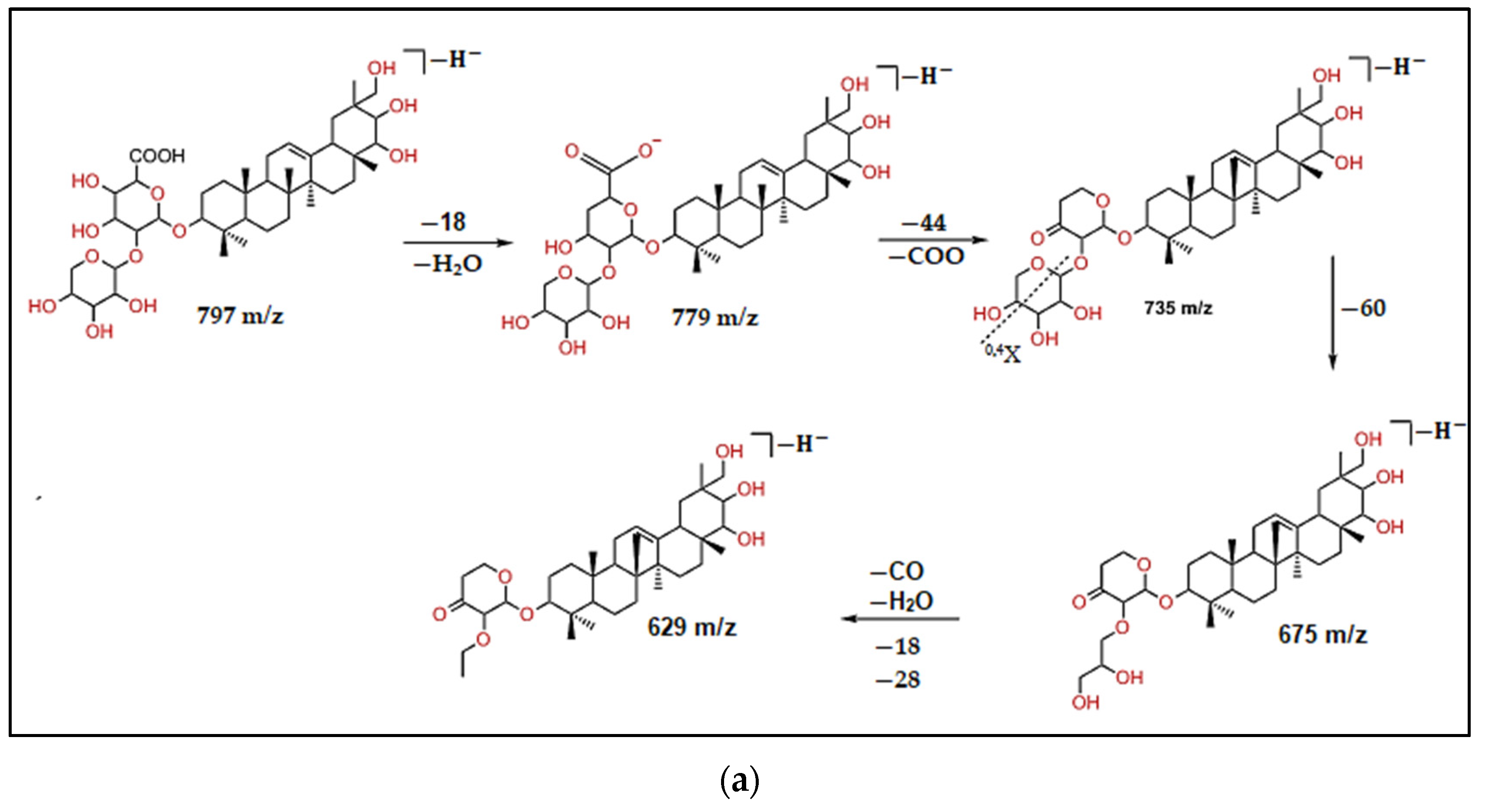
| 16 | 13.37 | 797 | 675, 629 | C41H65O15 | Kudzusapongenol A-hexA-pen | 2.4 |
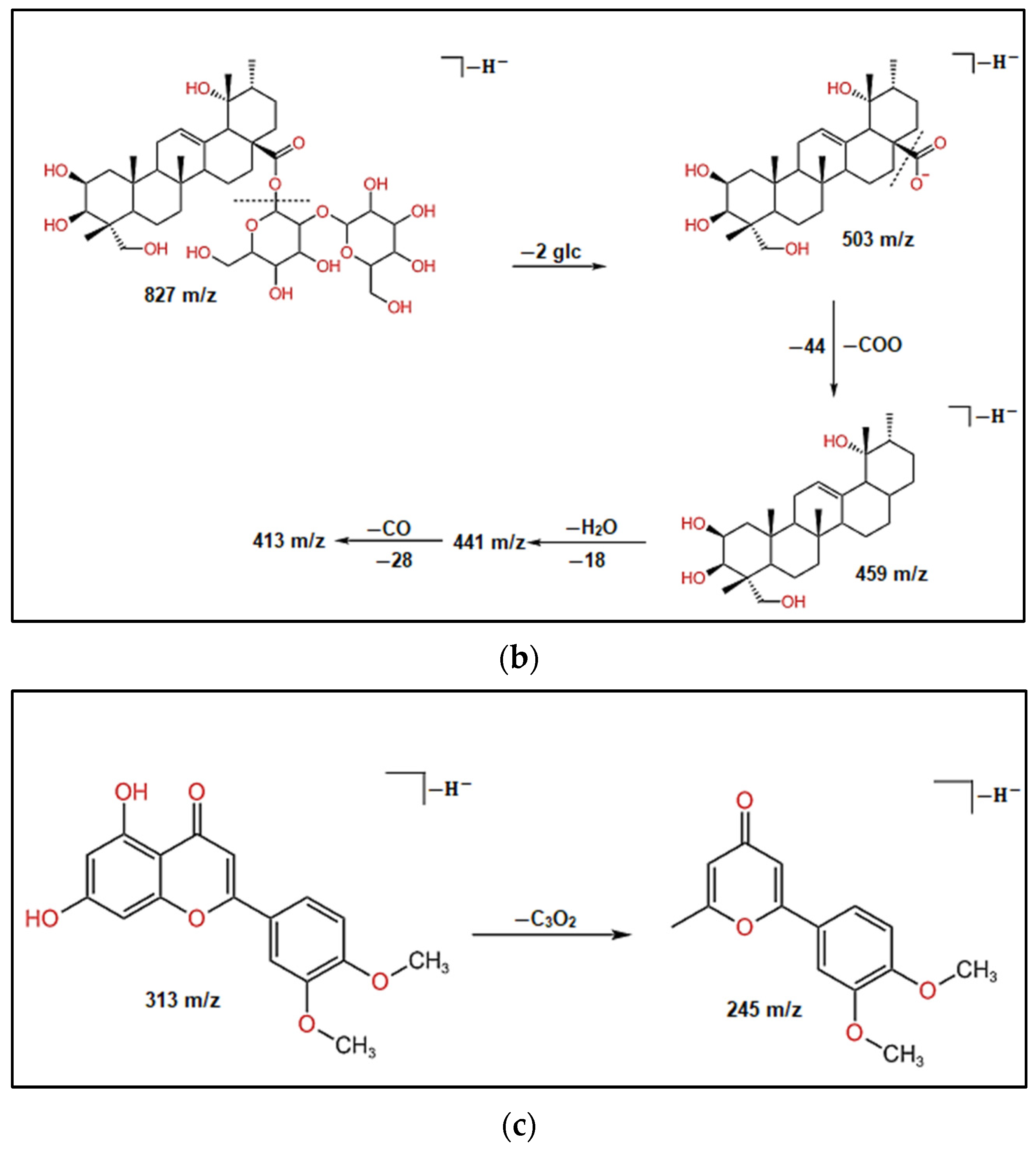
| 17 | 14.31 | 827 | 413 | C42H67O16 | Tetrahydroxy-urs-12-en-28-O-[b-D-glucopyranosyl (1-2)-b-D-glucopyranosyl] ester | 3.2 |
| 18 | 14.53 | 187 | C11H8O3 | Plumbagin | 0.3 | |
| 22 | 16.82 | 791 | 395 | Oleanoic acid-hex A–pent | 1.8 | |
| 27 | 20.29 | 865 | 525, 251 | C45H38O18 | Procyanidin C2 | 16.7 |
| 33 | 24.29 | 313 | 245 | C17H14O6 | 5,7-Dihydroxy-3,4-dimethoxyflavone | 4.5 |
| 42 | 29.1 | 555 | 393, 327, 295 | C26H36O13 | Tinosineside A | 0.1 |
| 43 | 29.34 | 537 | 341, 295 | C26H34O12 | Borapetoside A | 4.1 |
| Total | 83.3 |
| Inhibitor | 2D Interactions | Type of Bonds | Interacted Residues | Bond Length (Å) | BE (Kcal/mol) |
|---|---|---|---|---|---|
| Amentoflavone |  | Hydrogen bonds | His57 | 2.52 | −6.4 |
| Gly216 | 2.5 | ||||
| Carbon hydrogen bonds | Asn217 | 3.37 | |||
| Pi–Pi stacked | Trp215 | 5.89 | |||
| Benzamidine |  | Hydrogen bonds | Asp189 | 2.13 | −5.5 |
| Ser190 | 2.05 | ||||
| Salt bridge | Asp189 | 2.22 | |||
| 5,7-Dihydroxy-3,4-dimethoxyflavone |  | Hydrogen bonds | Ser195 | 3.18 | −6.5 |
| Asp189 | 3.02 | ||||
| Carbon hydrogen bond | Glu146 | 3.5 | |||
| Amid-Pi stacked | Trp215 (2) | 3.64 8.08 | |||
| Borapetoside A |  | Hydrogen bond | Asn99 | 2.91 | −4.2 |
| Gly216 (2) | 2.66 2.84 | ||||
| Carbon hydrogen bond | Trp41 | 3.63 | |||
| Pi–Pi T shaped | His57 | 5.07 | |||
| Trp41 (2) | 4.78 5.14 | ||||
| Pi alkyl | Cys42 | 5.08 | |||
| Plumbagin |  | −5.3 | |||
| Quercetin3-O-sophoroside-7-O-rhamnoside |  | hydrogen bonds | Gln192 (5) | 2.92 2.9 2.99 2.89 2.4 | −6.5 |
| Ser214 | 2.67 | ||||
| Asn99 | 2.43 | ||||
| Ser195 | 2.85 | ||||
| Pi–donor hydrogen bonds | His57 | 2.43 | |||
| Pi–Pi stacked | Trp215 | 5.63 | |||
 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Beltagi, H.S.; Aziz, S.M.S.A.; Aboshady, A.I.; Ibrahim, M.A.R.; Ibrahim, M.F.M.; Alenezi, M.A.; Darwish, D.B.E.; Al-Qahtani, S.M.; Al-Harbi, N.A.; Darwish, H.; et al. Isolation and Identification of Flavonoids from Black Cumin (Nigella sativa) by HPLC-MS and In Silico Molecular Interactions of Their Major Compounds with Fusarium oxysporum Trypsin-like Serine Protease. Separations 2023, 10, 360. https://doi.org/10.3390/separations10060360
El-Beltagi HS, Aziz SMSA, Aboshady AI, Ibrahim MAR, Ibrahim MFM, Alenezi MA, Darwish DBE, Al-Qahtani SM, Al-Harbi NA, Darwish H, et al. Isolation and Identification of Flavonoids from Black Cumin (Nigella sativa) by HPLC-MS and In Silico Molecular Interactions of Their Major Compounds with Fusarium oxysporum Trypsin-like Serine Protease. Separations. 2023; 10(6):360. https://doi.org/10.3390/separations10060360
Chicago/Turabian StyleEl-Beltagi, Hossam S., Seham M. S. Abdel Aziz, A. I. Aboshady, Mervat A. R. Ibrahim, Mohamed F. M. Ibrahim, Muneefah Abdullah Alenezi, Doaa Bahaa Eldin Darwish, Salem Mesfir Al-Qahtani, Nadi Awad Al-Harbi, Hadeer Darwish, and et al. 2023. "Isolation and Identification of Flavonoids from Black Cumin (Nigella sativa) by HPLC-MS and In Silico Molecular Interactions of Their Major Compounds with Fusarium oxysporum Trypsin-like Serine Protease" Separations 10, no. 6: 360. https://doi.org/10.3390/separations10060360
APA StyleEl-Beltagi, H. S., Aziz, S. M. S. A., Aboshady, A. I., Ibrahim, M. A. R., Ibrahim, M. F. M., Alenezi, M. A., Darwish, D. B. E., Al-Qahtani, S. M., Al-Harbi, N. A., Darwish, H., & Srour, H. A. M. (2023). Isolation and Identification of Flavonoids from Black Cumin (Nigella sativa) by HPLC-MS and In Silico Molecular Interactions of Their Major Compounds with Fusarium oxysporum Trypsin-like Serine Protease. Separations, 10(6), 360. https://doi.org/10.3390/separations10060360