Abstract
Edible flowers are a new “old” trend in modern gastronomy and healthy nutrition aimed at increasing food attractiveness and improving the consumption of beneficial phytocompounds that have valuable properties. Cucumber (Cucumis sativus L., Cucurbitaceae family) is a well-known agricultural product; it is an important crop that is commonly used across the globe. Despite sufficient knowledge of nutrients and secondary metabolites in cucumber fruits, the use of cucumber flowers for food purposes is still understudied. In this study, the total flavonoid fraction of cucumber flowers was isolated and purified after liquid–liquid and solid-phase extraction and polyamide and Amberlite XAD-2 separation followed by high-performance liquid chromatography with photodiode array detection and electrospray ionization triple quadrupole mass spectrometric detection (HPLC–PDA–ESI–tQ–MS/MS), resulting in the detection of 47 compounds. Identified flavonoids included luteolin, apigenin, and chrysoeriol-based flavones such as C-, O-, and C,O-glycosides, most of which were detected for the first time in C. sativus. Flower extracts from ten cucumber cultivars were quantified, and isoorientin, isoorientin-4′-O-glucoside, and cucumerins B and D were predominant. The use of in vitro assay potential to inhibit mammalian pancreatic lipases was demonstrated for the C. sativus extracts. The dominant compounds were studied separately, and luteolin derivatives were determined to be responsible for the bioactivity. To the best of our knowledge, this is the first study on cucumber flowers that has shown their wide potential for use as a beneficial dietary supplement.
Keywords:
cucumber; Cucumis sativus; edible flowers; flavonoids; human pancreatic lipase; inhibition 1. Introduction
Currently, dietics and related areas of nutritional science are experiencing a boom in ideas for ways to enrich the habitual human diet with useful components, including nutrients, minerals, vitamins, and bioactive compounds [1]. Most of the proposed ideas are based on scientific information regarding the presence of certain substances in food additives, the introduction of which makes it possible to adjust the composition of consumed products [2]. The most common food additional components are plants containing various functional components that transform ordinary food into a functional product [3]. Because plants are complex organisms consisting of various parts, we can use the leaves, roots, and flowers to functionalize products and increase the attractiveness of food [4]. The use of edible flowers in cooking is now being widely introduced as common practice, not only in cuisine, but also in dietics [5]. This morphological group of plants is distinguished by the greatest external attractiveness and its enriched, and sometimes unique, chemical composition [6]. To date, a large amount of scientific data has been collected on the metabolites—and their biological activity—of edible flowers used in the cuisines of Europe [7], Asia [8] and the Americas [9], indicating the indisputable advantages of flowers as functional components.
Among the variety of edible flowers used, plants of the Cucurbitaceae family are used in cooking, among which flowers of pumpkin, zucchini, and cucumber are especially popular owing to their wide availability [10]. Cucumber fruits are one of the most widely included crops in the human diet, the production of which has reached up to 90 million tons worldwide [11]. When cultivating cucumber, various secondary products are formed, which include cucumber flowers that do not require additional production conditions because they can be obtained during the process of growing cucumber fruits [12]. This circumstance makes cucumber flowers commercially attractive because at least two products can be produced simultaneously on the same territory of a greenhouse farm.
The known information about cucumber metabolites refers mainly to the fruit, which is logical owing to their wide use as a food product [13]. There are far fewer data available on the chemical components of vegetative biomass (leaves, stems, and flowers). Flavonoids [14,15], megastigmanes [16], and carotenoids [13] have been found in cucumber leaves, and photosynthetic pigments have been found in stems [17]. Some flavonoid glycosides, derivatives of quercetin, isorhamnetin, and kaempferol, have been found in cucumber flowers [18]. Data on the biological activity of cucumber flowers are not available. Given the inclusion of this plant product in the human diet, it is necessary to expand the database on the chemodiversity of cucumber flowers, as well as other edible flowers.
Studies on plant metabolites have confirmed their ability to have biological effects on living organisms [19]. The main compounds that possess a beneficial effect are phenolics, including flavonoids, which have various bioactivities [20]. Flavonoids can inhibit the activity of digestive enzymes [21], including pancreatic lipase [22], which makes this group of natural compounds promising for the treatment and prevention of lipid metabolism disorders. This activity was found for derivatives of various structures (flavonols, flavones, flavans, etc.) [23], among which the inhibition efficiency increased for luteolin and apigenin-based flavones [24]. In cucumber leaves, various derivatives of these two flavones were also previously found and characterized [14,15], and for several compounds, as well as for the total extract, they can be considered active inhibitors of mammalian pancreatic lipases [25,26]. Owing to the lack of scientific data on the metabolites of cucumber flowers, in-depth studies of the phenolic compounds in this product are necessary; in particular, the composition of flavonoids should be studied, and their potential to inhibit digestive lipases should be determined.
In addition to studying the chemical composition and biological activity of total plant extracts, studies on the purified fractions of individual compounds are of great importance, because they have a greater pronounced beneficial effect than crude extracts [27]. These target fractions make it possible to study the composition of metabolites in more detail because they do not contain interfering and ballast substances, which is very important for highly efficient research methods, such as LC–MS and GC–MS [28]. Cucumber flowers have never been studied using LC–MS profiling, which could fill gaps in knowledge regarding the composition of this important food product.
Thus, as part of an ongoing study of cucumber metabolites [25,26], an in-depth study of cucumber flower flavonoids was performed, including LLE and SPE separation of cucumber flower metabolites, macroporous resin separation of flavonoid fraction, LC–MS profiling of flavonoids by high-performance liquid chromatography with photodiode array detection and electrospray ionization triple quadrupole mass spectrometric detection (HPLC–PDA–ESI–tQ–MS/MS), quantification of flavonoids in cucumber flowers, and studying the inhibitory potential of cucumber flavonoids on mammalian lipases.
2. Materials and Methods
2.1. Plant Material and Chemicals
Cucumber flowers were harvested in Buryat Fruit and Plant Nursery (Ulan-Ude, Russia) using authenticated cucumber seeds (cv. Arcadia F1, April F1, Madhur F1, Parisian Gherkin F1, Perseus F1, Secret F1, Shruti F1, Titus F1, Tulsi F1, Zozula F1) purchased in the National Seed Repository (Moscow, Russia). All plants were authenticated by Prof. N.I. Kashchenko (IGEB SB RAS, Ulan-Ude, Russia). Fresh flowers were transported to the laboratory at 4 °C within 1–2 h and microwave dried to a moisture content <5%. Dried flowers were stored at a desiccator containing moisture-indicating silica gel as a desiccant before extraction and analysis. The reference standards were purchased from ChemFaces (Wuhan, Hubei, China): juncein (≥98%; cat. No CFN93776), isoscoparin-2″-O-glucoside (≥98%; cat. No CFN90900); Pharmaffiliates (Panchkula, Haryana, India): isoorientin-2″-O-glucoside (≥98%; cat. No PA270020766); Sigma-Aldrich (St. Louis, MO, USA): orientin (≥98%; cat. No O9765), isoorientin (≥98%; cat. No I1536). Selected flavones were plant-derived metabolites isolated in our laboratory including cucumerins A–D [25], isoorientin-7-O-glucoside [29], isoorientin-6″-O-glucoside [30], isoorientin-4′-O-glucoside [31], luteolin-6-C-arabinoside, isoscoparin-7-O-glucoside [32], and saponarin-6″-O-ferulate [33].
2.2. Liquid–Liquid Extraction (LLE) and Solid-Phase Extraction (SPE) of Cucumber Flowers
Dried flowers (100 g; cv. Secret F1) were ground and treated by methanol (1.5 L) with sonication three times (ultrasonic bath, 20 min, 40 °C, ultrasound power 100 W, frequency 35 kHz). Methanolic extract centrifuged (6000 rpm, 20 min) and supernatant was additionally filtered through cellulose filter. Resultant solution was concentrated in vacuum, the residue was suspended in water (1:5) and extracted with hexane, chloroform and n-butanol to give hexane LLE-fraction (4.2 g; A), chloroform LLE-fraction (1.2 g; B) and n-butanol LLE-fraction (12.7 g; C). Mother liquor was concentrated in vacuum to give water LLE-fraction (15.3 g; D). The n-butanol LLE-fraction (10 g) was separated by solid-phase extraction on polyamide sorbent (250 g; Sigma-Aldrich, No 02395) preconditioned with methanol (400 mL) and water (1 L) using water (5 L) and 0.5% NH3 in ethanol (5 L) to give SPE-1 subfraction (5.76 g; C1) and SPE-2 subfraction (2.27 g; C2), respectively.
2.3. Chemical Composition of LLE and SPE Fractions
Ready-to-use kits for spectrophotometric assays were applied to measure the total lipids (Lipid Assay Kit, Abcam plc., Shanghai, China; cat. No ab242307), carotenoids (Plant Carotenoid Content Assay Kit, SunLong Biotech Co., LTD, Hangzhou, Zhejiang, China; cat. No AK0079), flavonoids (OxiSelect™ Flavonoid Assay Kit, Cell Biolabs, Inc., San Diego, CA, USA; cat. No XAN-5077), phenolics (Phenolic Compounds Assay Kit, Sigma-Aldrich; cat. MAK365), carbohydrate content (High Sensitivity Carbohydrate Assay Kit, BioVision, Inc., Milpitas, CA, USA; cat. No. K2049-100), and protein (Pierce™ BCA Protein Assay Kit, Thermo Fisher Scientific, Waltham, MA, USA). Total triterpene content was determined using spectrophotometric assay and β-sitosterol as a reference compound [34]. Ash content was determined by the AOAC Official MethodSM to be 942.05 using muffle furnace ignition at 600 °C [35]. All analyses were performed five times, and the data were expressed as the mean value ± standard deviation (S.D.).
2.4. Macroporous Resin Separation of SPE-2 Subfraction
Macroporous resins (D-101, AD-8, Amberlites XAD-2, XAD-4, XAD-16, XAD-7, XAD-8) were immersed in methanol for 48 h, and washed with water. Then, the resins were washed by 5% HCl, water, 5% NaOH, and water to neutral eluate and dried at 70 °C. To screening, 1 g of dry resin was pretreated with SPE-2 subfraction solution in 20% ethanol (2 mg/mL, 60 mL) in a conical flask (250 mL) with stirring (150 rpm) for 14 h at 20 °C, and then the resin was desorbed with 100% ethanol with stirring (150 rpm) for 14 h at 20 °C. Flavonoid content in liquid phases was monitored using OxiSelect™ Flavonoid Assay Kit (Cell Biolabs, Inc.) before and after SPE-2 subfraction loading of resins and after resin desorption. To calculate the values of adsorption capacity (Qa, mg/g), desorption capacity (Qd, mg/g) and desorption ratio (Rd, %), a known equation was used [27]. Adsorption kinetic curve for the resins brands was build using the data of monitoring the samples at equal time intervals till equilibrium (ca. 14 h). The absorption capacity of the Amberlite XAD-2 was determined using 0–10 mg/mL SPE-2 subfraction solution after 6 h phase contact. Desorption ability of the eluents was studies with 0–100% water solutions of methanol. ethanol, and isopropanol with 5 mg/mL SPE-2 subfraction solution and 6 h phase contact. The value of elution flow was determined in a dynamic experiment used Amberlite XAD-2 resin (75 g) loaded to the glass column, saturated with 5 mg/mL SPE-2 subfraction solution (80 mL, 6 h) and pre-washed with water (5 BV). Flow rates 1–5 BV/h were studied with elution 1–7 BV through the column.
2.5. High-Performance Liquid Chromatography with Photodiode Array Detection and Electrospray Ionization Triple Quadrupole Mass Spectrometric Detection (HPLC–PDA–ESI–tQMS)
Metabolite profiling was performed using high-performance liquid chromatography with photodiode array detection and electrospray ionization triple quadrupole mass spectrometric detection (HPLC-PDA-ESI-tQ-MS) with an LC-20 Prominence liquid chromatograph (Shimadzu, Columbia, MD, USA). A photodiode array detector SPD-M30A (wavelength range 200–600 nm) and triple quadrupole mass spectrometer LCMS 8050 (all Shimadzu, Columbia, MD, USA) were coupled with chromatography. Separation was managed using GLC Mastro column (2.1 × 150 mm, 3 μm; Shimadzu, Kyoto, Japan) eluted with 0.5% HCOOH in water (eluent A) and 0.5% HCOOH in MeCN (eluent B) and gradient program (0–2 min 5–8% B, 2–5 min 8–9% B, 5–12 min 9–36% B, 12–13 min 36–59% B, 13–15 min 59–78% B, 15–20 min 78–90% B, 20–25 min 90–5% B). The values of flow rate, column temperature and injection volume were 100 μL/min, 30 °C, and 1 μL, respectively. The negative electrospray ionization mode (source voltage 3 kV, collision energy −35 eV, scanning range m/z 80–2000) was used for the mass spectrometric detection and three temperature levels used in ESI interface, desolvation line, and heat block equal to 300 °C, 250 °C, and 400 °C, respectively. The flow rates of nebulizing gas (N2), heating gas (air), and collision-induced dissociation gas (Ar) were 3 L/min, 10 L/min, and 0.3 mL/min, respectively. Metabolite identification was performed using retention time, ultraviolet spectra, and mass spectra data after compared to reference standards and literature data exploiting LabSolution (Shimadzu) workstation software equipped with an internal LC–MS library.
To quantify the selected compounds the sample was separated as described above, and the full scan MS peak area was used for calculation. The following reference substances were used to build calibration curves: isoorientin (for quantification of isoorientin-4′-O-glucoside, isoorientin, and luteolin-6-C-arabinoside), isovitexin (for quantification of cucumerins B, C, and D), orientin, juncein and isoscoparin (for quantification of chrysoeriol-C-hexoside-O-hexoside-O-ferulate). Stock solutions were prepared after careful weighing (10 mg) of reference substances and dissolving them in volumetric flasks (10 mL) using a methanol–DMSO mixture (1:1) followed by the creation of ‘concentration–peak area’ graphs (1–100 µg/mL). The values of correlation coefficient (r2), standard deviation (SYX), limit of detection (LOD), limit of quantification (LOQ), and linear range were calculated in Advanced Grapher 2.2 (Alentum Software Inc., Ramat-Gan, Israel) using calibration curve data [36] and the results of three sufficient HPLC runs (Table 1). Intra-day, inter-day precisions and recovery of spiked sample were studied using the known assay [37]. The results are expressed as mean values ± standard deviation (S.D.).

Table 1.
Regression equations, correlation coefficients (r2), standard deviation (SYX), limits of detection (LOD), limits of quantification (LOQ), and linear ranges for six reference standards.
2.6. Mammal Lipase Inhibition In Vitro Assay
Spectrophotometric assay was used to study inhibitory potential of extracts and selected compounds on porcine pancreatic lipase (type II, ≥125 U/mg protein; Sigma-Aldrich, cat. No L3126) and recombinant human pancreatic lipase (Sigma-Aldrich, cat. No BCR694) [38]. p-Nitrophenol palmitate was used as a substrate and orlistat (tetrahydrolipstatin) was used as a reference substance [22,23,24]. All analyses were carried out three times, and the data are expressed as mean value ± standard deviation (S.D.).
2.7. Statistical Analysis
Statistical analyses were performed by one-way analysis of variance, and the significance of the mean difference was determined by Duncan’s multiple range test. Differences at p < 0.05 were considered statistically significant. The results are presented as the mean ± S.D. The linear regression analysis and generation of calibration graphs were conducted using Advanced Grapher 2.2 (Alentum Software, Inc., Ramat-Gan, Israel).
3. Results and Discussion
3.1. LLE and SPE Separation of Cucumber Flower Metabolites and Mammal Pancreatic Lipase Inhibition by CF Fractions
Preliminary separation of cucumber flower extractives was performed for the methanol extract using liquid–liquid extraction (LLE) [39] and solid-phase extraction (SPE) [40]. The LLE sequence included hexane, chloroform, and n-butanol extraction and resulted in the isolation of fractions A, B, and C, respectively, and the water residue was labeled as water fraction D (Table 2).
Table 2.
Yield, composition, and mammal pancreatic lipase inhibition (PLI) of cucumber flower fractions.
Chemical analysis demonstrated the presence of lipids, carotenoids, and triterpenes in fractions A and B, representing the largest part of the fractions, whereas mid- (flavonoids, phenolics, triterpenes) and high-hydrophilic components (carbohydrates, proteins) were found in fractions C and D. Only the n-butanol LLE-fraction (C) showed inhibitory potential against mammalian pancreatic lipase with an IC50 value <100 μg/mL (89.7–93.4 μg/mL), which we chose for its following separation by SPE. The use of a polyamide sorbent [41] allowed us to divide fraction C into two subfractions eluted with water (C1) and a 0.5% NH3/ethanol mixture (C2). Chemical analysis showed that triterpenes, carbohydrate, and ash were concentrated in subfraction C1, and phenolics detected at a non-trace level only in subfraction C2 (51.9 g/100 g) consisted mostly of flavonoids (48.9 g/100 g). The IC50 in the bioactivity assay for subfraction C2 was 42.7–45.9 μg/mL, which is likely because of the high flavonoid content, which are known inhibitors of pancreatic lipase [42]. Early studies demonstrated the expressed inhibitory potential of the plant LLE fraction with a high phenolic content against pancreatic lipase for the Korean yam (Dioscorea opposita) [43], root of Rhodiola crenulata [44], aniseed (Pimpinella anisum) [45], yellow cassia (Cassia siamea) [46], and many others.
3.2. Amberlite XAD-2 Separation of Cucumber Flower Flavonoids
To purify the flavonoid fraction, macroporous resin separation of the SPE-2 subfraction was applied. Non-ionic polymeric resins are known for their excellent performance for flavonoid separation and concentration [47]. Screening of various resins was performed using seven brands, including polystyrene resins, such as D-101, AD-8, and the Amberlites XAD-2, XAD-4, XAD-16, and polyacrylic resins, such as Amberlites XAD-7 and XAD-8, which were loaded with an SPE-2 subfraction solution (2 mg/mL) which was used as a sample for testing the adsorption and desorption properties. Among the studied resins, the polystyrene Amberlite XAD-2 showed the highest level of adsorption capacity (43.2 mg/g), desorption capacity (37.5 mg/g), and desorption ratio (86.8%) (Table 3).
Table 3.
Adsorption and desorption properties of seven macroporous resins.
The adsorption capacity of cucumber flower total flavonoids increased until 6 h and reached equilibrium after 6 h of phase contact for all studied resins (Figure 1a). Based on the obtained results, the resin Amberlite XAD-2 was chosen for further experimental work. The absorption capacity of Amberlite XAD-2 determined through the equilibrium adsorption curve increased from 18 mg/g at 1 mg/mL of flavonoid solution to 190 mg/g at 6 mg/mL of flavonoid solution (Figure 1b). Upon increasing the sample concentration by more than 6 mg/mL, there was no resin capacity increase.
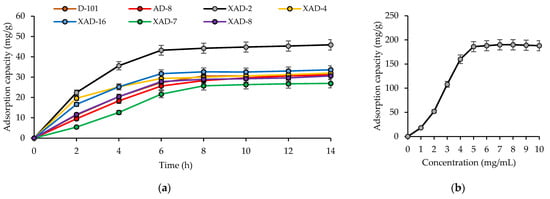
Figure 1.
Adsorption curves for total cucumber flower flavonoids on seven macroporous resins (a) and equilibrium adsorption curve for total cucumber flower flavonoids on Amberlite XAD-2 (b).
In the process of selecting the best eluting system, alcohol–water mixtures have been traditionally used as macroporous resin eluents for flavonoid purification [27]. Low-molecular-weight alcohols showed effective flavonoid desorption for 40–50% solutions with the best performance from 40% ethanol, which gave 92.6% desorption (Table 4).
Table 4.
Desorption ratio of total cucumber flower flavonoids on Amberlite XAD-2 after elution with alcohol-water mixtures.
The elution flow rate directly affects the effectiveness of flavonoid desorption from macroporous resins and is therefore a significant factor influencing the quality of the final phenolic extract [48]. Elution of the cucumber flower flavonoids from Amberlite XAD-2 proceeded under optimal conditions at a flow rate equal to 2 bed volumes per hour (BV/h), which resulted in 92.8% flavonoid desorption with 5 BV eluted through the column (Table 5). A reduced flow rate (1 BV/h) gave 91.3% flavonoid desorption with 7 BV of eluents, and an increase in the flow rate (3–5 BV/h) showed 91–92% flavonoid desorption with 6 BV of eluents.
Table 5.
Desorption ratio of total cucumber flower flavonoids on Amberlite XAD-2 at various flow rate.
Spectrophotometric data of the cucumber flower flavonoid fraction obtained after Amberlite XAD-2 purification of the n-butanol LLE fraction (C2) under optimal conditions indicated significant increase in total flavonoid content to 92–93% of dry fraction weight. The effectiveness of macroporous resins for purification of flavonoids was demonstrated early for Acanthopanax senticosus [48], Elaeis guineensis [49], Lycium barbarum [50], Scorzonera austriaca [27], and Smilax glabra [51].
3.3. LC-MS Profile of Flavonoids in the Amberlite XAD-2 Fraction of Cucumber Flowers
Application of high-performance liquid chromatography with photodiode array detection and electrospray ionization triple quadrupole mass spectrometric detection (HPLC–PDA–ESI–tQ–MS/MS) resulted in the separation of 47 compounds (Figure 2). Identification of analytes was performed by comparing retention times, UV, and mass-spectral patterns with those of reference substances and literature data [15,25,52,53,54,55,56,57,58,59] (Table 6). Forty-seven compounds found in the Amberlite XAD-2 fraction of cucumber flowers were characterized as flavonoids, including fifteen flavones identified using reference standards (Figure 3 and Figure 4) and thirty-two metabolites with tentative structures. All compounds were flavones originating from luteolin, apigenin, and chrysoeriol.
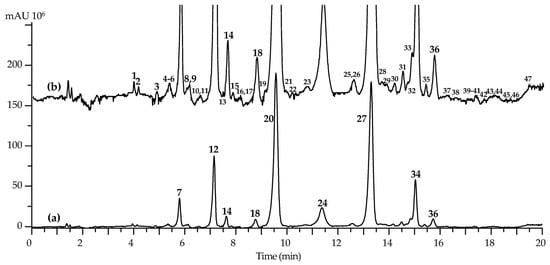
Figure 2.
High-performance liquid chromatography data of Amberlite XAD-2 fraction of cucumber flowers with photodiode array detection ((a); 330 nm) and electrospray ionization triple quadrupole mass spectrometric detection ((b); negative detection, base peak chromatogram). Compounds are numbered as listed in Table 6.
Table 6.
Chromatographic (t) and ultraviolet (UV) and mass-spectrometric (ESI-MS) data of compounds 1–47 found in Amberlite XAD-2 fraction of cucumber flowers.
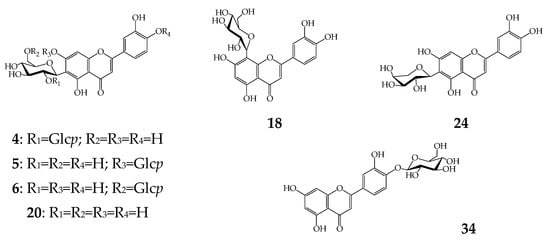
Figure 3.
Known luteolin glycosides found in cucumber flowers. Glcp—glucopyranose.
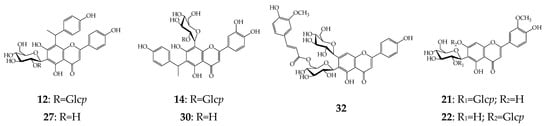
Figure 4.
Known apigenin and chrysoeriol glycosides found in cucumber flowers. Glcp—glucopyranose.
3.3.1. Luteolin Glycosides
Twenty-one derivatives of luteolin were C-, O-, and C,O-glycosides with non-acylated and acylated structures. Luteolin-4′-glycoside (34) was the only O-glycoside found among the cucumber flavonoids and identified by direct comparison of tR, UV, and MS data with the reference standard. Three C-glycosides, orientin (luteolin-8-C-glucoside, 18), isoorientin (luteolin-6-C-glucoside, 20), and luteolin-6-C-arabinoside (24), gave specific mass-spectrometric patterns that are typical for flavone-C-glycosides [52] and similar to those of the reference standards.
The largest group of luteolins was C,O-glycosides with 17 compounds, including the known flavones of isoorientin-2″-O-glycoside (4) [53], isoorientin-7-O-glycoside (lutonarin, 5) [60], isoorientin-6″-O-glycoside (6) [61], and isoorientin-4′-O-glycoside (7) [62]. Tentative structures were attributed to three isomeric flavones, 1–3, which gave a deprotonated ion with m/z 771, sequential loss of two hexose fragments (m/z 771→609→447) [63], and an ion cascade typical for luteolin-C-glycosides such as orientin and isoorientin (m/z 357, 329, 327, 299) [52]. The abovementioned results indicated the structures of 1–3 as possibly being luteolin-C-hexoside-di-O-hexosides. The UV spectral data for 1 and 2 (λmax 255, 270, 350 nm) were close to 6/8-C- and 7-O-glucosides of luteolin [64], suggesting the presence of this type of substitution as in the case of isoorientin-7,2″-di-O-glycoside or isoorientin-7,2‴-di-O-glycoside putatively assigned in barley leaves [65], but thus far not isolated from any plant. The spectral pattern of 3 showed a hypsochromic shift of band I (1, 2: 350 nm→3: 338 nm), which is possible in the case of 4′-O-substituted luteolin glycosides [63] as for isoorientin-4′,2″-di-O-glycoside [66].
The remaining luteolins were acylated C,O-glycosides incorporating fragments of acetic and ferulic acids. Isomeric acetates of luteolin-C-hexoside-di-O-hexoside 13, 15, and 17 showed the loss of an acyl group with m/z 42 followed by the series of ions found in the cleavage products of 1–3. Most likely, 13, 15, and 17 are the mono-acetates of 1–3, although to date there is still no information on natural flavones with similar structures. Compounds 28, 29, and 31 gave ions with m/z 785 ([M − H]−) and 609, indicating the loss of the 176 amu fragment. The hypsochromic shift of bands I compared with isoorientin (349→333 nm) is possible in the case of ferulic fragment substitution [33,52]. The other ions were close to luteolin-C-hexoside-O-hexosides, such as compounds 4–6, suggesting the structures of 28, 29, and 31 as possibly being mono-feruloylated analogs of 4–6, for which the natural equivalents are still unknown.
Compounds 35 and 36 were concluded to be luteolin-C-hexoside-O-ferulates owing to the UV and MS data showing similarities to isoorientin-2″-O-ferulate and orientin-2″-O-ferulate [57]. Flavone 41 was luteolin-C-pentoside-O-ferulate because its mass spectrum was similar to that of 24 (luteolin-6-C-arabinoside) from which the feruloyl fragment has been previously removed. The only acylated C,O-glycoside 38 contained the residues of both acetic and ferulic acids. Evidence of this included the loss of ferulic (m/z 665→489) and acetic acid (m/z 489→447), followed by the cleavage pattern typical for isoorientin/orientin [52]. The natural compounds close to 38 and 41 are unknown.
3.3.2. Apigenin Glycosides
Sixteen apigenins were found in cucumber flowers, including the reference identified as saponarin-6″-O-ferulate [29] and specific Cucumis flavones constituting apigenin fused with the 4-hydroxy-1-ethylbenzene residue at the C-6 or C-8 positions. Among the latter compounds, we detected apigenin-6-C-(4-hydroxy-1-ethylbenzene)-8-C-glucoside or cucumerin A (30) and apigenin-6-C-glucoside-8-C-(4-hydroxy-1-ethylbenzene) or cucumerin B (27), which were first isolated from the leaves of C. sativus infected by a plant pathogen that caused powdery mildew (Podosphaera fuliginea (Schltdl.) U. Braun & S. Takam.) [15], and glucosides of 27 (cucumerin C) and 30 (cucumerin D) were isolated by us from healthy cucumber leaves [25].
Flavones 16 and 19 were the isomers of 12 and 14 and are likely to contain the isomeric to glucose fragment of galactose. Compounds 8–11 showed similar mass-spectral patterns to those of cucumerins C and D in which the presence of an additional hexose fragment represented apigenin-C-hexoside-C-(4-hydroxy-1-ethylbenzene) di-O-hexosides. The nature of compounds 8–11, 16, and 19 is unknown, and more research is needed to determine their structures. The approach used for identification of luteolin glycosides made it possible to determine the tentative structures of acylated glycoside. Compound 33 was found to be apigenin-C-hexoside-O-hexoside-O-ferulate isomeric to 32, compounds 40 and 42 were acylated cucumerins or apigenin-C-hexoside-C-(4-hydroxy-1-ethylbenzene)-O-ferulate and apigenin-C-hexoside-C-(4-hydroxy-1-ethylbenzene)-O-ferulate-O-acetate, respectively, and isomeric apigenin-C-hexoside-O-ferulates 45 and 46 were most likely ferulates of isovitexin and vitexin [29,33].
3.3.3. Chrysoeriol Glycosides
Derivatives of chrysoeriol were the smallest group of flavones detected in cucumber flowers that are biogenetically related to luteolins. Two known compounds identified after comparison of chromatographic and spectral data with reference standards included isoscoparin-2″-O-glucoside (21) and isoscoparin-7-O-glucoside (22). Flavones 23, 25, and 26 were concluded to be chrysoeriol-C-hexoside-O-hexosides and had similar spectral patterns to 21 and 22. Elimination of the feruloyl fragment was detected in the mass spectra of 37, 39, and 43 (m/z 799→623), and after that, cleavage was similar to isoscoparin O-glucosides 21 and 22, indicating the probable structure of chrysoeriol-C-hexoside-O-hexoside-O-ferulates, which are still unidentified in plants. The deprotonated ion of 43 was 42 amu larger, which indicated the presence of an additional acetyl group upon evaluating the structure of chrysoeriol-C-hexoside-O-hexoside-O-ferulate-O-acetate. Compound 47 was found to be chrysoeriol-C-hexoside-O-ferulate owing to its spectral similarity with isoscoparin/scoparin (chrysoeriol-6/8-C-glucosides) and arguably is isoscoparin-2″-O-ferulate isolated earlier from other cucurbits, Bryonia alba and B. dioica [67].
Previously, in flowers of cultivars of C. sativus (Olimp F1, Hela F1, Cezar F1, Polan F1, Monastyrski, and Delicius) from Poland, after HPLC and TLC analyses, only flavonols were detected, including kaempferol 3-O-glucoside, isorhamnetin 3-O-glucoside, quercetin 3-O-glucoside, and kaempferol 3-O-rhamnoside [18]. In our study, only flavones were identified, which may be the reason for the individual chemical variability. For the other flavonoids, isoorientin was described in the leaves of Polish cucumber cultivars [18] and orientin and cucumerins A and B in the leaves of Canadian cultivars [15]. The remaining compounds were described for C. sativus for the first time. The presence of feruloylated flavone-C,O-glycosides derived from isovitexin and isoscoparin was demonstrated in leaves of UK cucumber cultivars [14], which combined with other data suggests that acylation of the flavonoids in cucumber tissues is characteristic for the species or genus overall, although the last statement requires additional confirmation.
3.4. Quantification of Nine Flavonoids in Cucumber Flowers
To quantify cucumber flowers, nine flavonoids were selected as markers, including isoorientin-4′-O-glucoside (7), cucumerin D (12), cucumerin C (14), orientin (18), isoorientin (20), luteolin-6-C-arabinoside (24), cucumerin B (27), juncein (34), and chrysoeriol-C-hexoside-O-hexoside-O-ferulate (37) (Table 7).
Table 7.
Content of nine flavonoids in flowers of ten cucumber cultivars.
The main criterion included non-trace concentrations of compounds, and the markers completely satisfied this requirement. Ten cultivars were those most commonly used in greenhouses of Baikal Region to produce cucumber fruits and flowers. The obtained results demonstrated variations in total flavonoid content from 6.94 mg/g (cv. Parisian Gherkin F1) to 11.23 mg/g (cv. Perseus F1). The dominant flavonoids included isoorientin (1.52–3.24 mg/g), cucumerin B (1.42–3.33 mg/g), isoorientin-4′-O-glucoside (0.93–1.92 mg/g), and cucumerin D (0.52–1.27 mg/g), which account for more than 80% of total flavonoids in cucumber flowers.
The known data on flavonoid content in cucumber flowers was found for Chinese cultivars containing 5.45 mg/g [8]. Edible flowers of varying botanical origin may contain fewer flavonoids, as in the flowers of Malus spectabilis (1.6 mg/g), Dianthus caryophyllus, Rosa rugosa (2.5 mg/g), Crocus sativus (2.6 mg/g), and Oroxylum indicum (3 mg/g), or a similar content, as in the flowers of Matthiola incana (7.5 mg/g), Nelumbo nucifera (7.8 mg/g), Rosa centifolia (10.9 mg/g), and Myosotis sylvatica (11.1 mg/g) [68,69]. In this regard, edible flowers of C. sativus may be considered a good source of flavonoids.
3.5. Inhibition of Mammal Lipases by Flavonoids of Cucumber Flowers
Fourteen selected flavonoids were studied as inhibitors of two mammalian pancreatic lipases of porcine (PPL) and human origin (HPL), and the obtained results demonstrated a similar mode of action for the studied compounds (Table 8). The group of luteolin derivatives included nine flavone C-, O-, and C,O-glycosides. Isomeric isoorientin and orientin were different with respect to the extent of inhibitory potential, and luteolin-6-C-glucoside has an advantage towards isoorientin (IC50 22.63 μM—PPL; 15.32 μM—HPL). The attachment of an additional glucose moiety to the isoorientin skeleton induced altered bioactivity with the most pronounced effect in the case of isoorientin-2″-O-glucoside (IC50 12.68 μM—PPL; 10.06 μM—HPL). The influence of the 6″-O-attached glucose was insignificant (IC50 22.15 μM—PPL) or resulted in lower activity (IC50 30.63 μM—HPL). The substitution of 7-O- and 4′-O-hydroxyl groups of isoorientin greatly reduced inhibition of PPL (IC50 49.54 μM—isoorientin-7-O-glucoside; 63.18 μM—isoorientin-4′-O-glucoside) and HPL (IC50 38.62 μM—isoorientin-7-O-glucoside; 77.11 μM—isoorientin-4′-O-glucoside). In the transition from isoorientin to its 3′-O-methyl ester (isoscoparin), a significant decrease in the inhibitory activity was observed, regardless of the location of the glucosyl substituent. These results demonstrate that the 6-C-located glucose and its 2″-O-substitution may play a fundamental role in mammalian lipase inhibitory properties of luteolin derivatives. Previously, similar phenomenon was found for isoorientin and isoorientin-2″-O-rhamnoside isolated from centipede grass (Eremochloa ophiuroides) and showed good inhibition of PPL [70]. Comparatively speaking, non-6-C-glucosylated juncein (luteolin-4′-O-glucoside) is a weak lipase inhibitor.
Table 8.
Inhibition of mammal lipase activity by selected flavonoids from cucumber flowers, IC50, μM ± S.D.
Oher flavonoids were derivatives of apigenin, among which the most active was cucumerin B or 6-C-glucoside of apigenin-8-C-(4-hydroxy-1-ethylbenzene) (IC50 25.63 μM—PPL; 20.89 μM—HPL) and its 2″-O-glucoside or cucumerin D (IC50 12.53 μM—PPL; 10.35 μM—HPL). 8-C-glucosylation reduced the bioactivity of flavones, as in the case of cucumerins A and C. Saponarin-6″-O-ferulate, despite the presence of 6-C-glucose, was also 7-O-substituted, which negatively affected the bioactivity of the compound. Thus, the structure–activity relationships found for luteolin derivatives have also been observed in apigenin glycosides.
Flavonoids, owing to their wide distribution in plants and the wide variety of structural types, have been studied many times and are still being studied for their influence on the activity of mammalian pancreatic lipases [22]. The interest in this problem is associated with evidence of the inhibitory effect of flavonoids on this group of digestive enzymes [71]. Among the key structural features, the presence of which leads to the appearance of the biological effect, are the presence or absence of additional fragments of monosaccharides and/or benzoic acids or hydroxycinnamates attached to the main skeleton of the molecule [72]. These regions of the molecule can form hydrogen bonds or participate in the van der Waals interaction between the flavonoid molecule and fragments of the lipase molecule containing residues of phenylalanine, tyrosine, and tryptophan [73]. Because of these interactions, the microenvironment of regions of the lipase molecule is transformed, and its conformation is changed, which negatively affects the ability of the enzyme to perform lytic functions. The molecules of hesperidin [74], catechins [75], myricetin [76], and luteolin [70] have a similar effect on lipases. Our data on the pronounced inhibitory effect of 2″-O-glucosylated derivatives of luteolin and apigenin are most likely associated with this phenomenon of changing the configuration of the enzyme macromolecule. To obtain more rigorous evidence for this hypothesis, additional studies are needed, which will be performed in the future.
The data obtained in the bioactivity study revealed the significance of C. sativus flavones for the anti-mammalian lipase effect of the total flavonoid fraction, which in turn can affect the activity of the total flower extracts. New information about the chemical composition and biological activity of C. sativus indicates that this agricultural species is not only a source for obtaining traditional food products, which are fruits, but also clearly confirms the possibility of using cucumber flowers as a culinary and dietary product. The variety of glycosylflavones found in cucumber flowers suggests the presence of “usual” types of activity for flavonoids, such as antioxidant, antimicrobial, anti-inflammatory, and others [77,78,79,80], which has been repeatedly confirmed for various representatives of the luteolin and apigenin series [80,81]. Of great importance are data on the high quantitative content of flavones in cucumber flowers (~700–1100 mg/100 g), which is close to the values of known food sources of flavonoids, such as those for parsley, black currants, blueberries, blackberries, green tea (~100–300 mg/100 g), elderberries, capers, and chokeberry (~300–1500 mg/100 g) [82], and allows us to consider them as an additional food source of this group of compounds. Separately, the production of cucumber flowers does not require the organization of additional greenhouse space because they can be obtained on the territory occupied by the production of fruits. In most cases, parthenocarpic varieties are used for growing cucumbers, the removal of flowers from which does not affect the quality or productivity of plants; thus, cucumber flowers can easily become a new and affordable food product.
4. Conclusions
Chromatographic study of cucumber (Cucumis sativa) flower metabolites indicated the presence of flavonoids, which were accumulated in the total fraction after LLE, SPE, and macroporous resin separation. Application of HPLC–PDA–ESI–tQ–MS/MS assay made it possible to identify 47 flavones that were derivatives of luteolin, apigenin, and chrysoeriol in the form of glycosides. To the best of our knowledge, our study is the first extensive study of cucumber flower flavonoids. Together with data on the high quantitative content of flavonoids, the flowers of C. sativa are a rich source of dietary flavonoids. The flavonoid fraction and selected compounds demonstrated the ability to inhibit mammalian pancreatic lipases in some cases close to the orlistat potential. These results provide new data on the beneficial properties of edible flowers of cucumber as an original functional food.
Funding
This research was funded by Russian Science Foundation, grant number 23-26-00063.
Data Availability Statement
Not applicable.
Acknowledgments
The author acknowledges the Buryat Research Resource Center for the technical support in chromatographic and mass-spectrometric research.
Conflicts of Interest
The author declares no conflict of interest.
References
- Dixon, K.A.; Michelsen, M.K.; Carpenter, C.L. Modern diets and the health of our planet: An investigation into the environmental impacts of food choices. Nutrients 2023, 15, 692. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Yang, J.; Chen, J.; Pu, X.; Li, X.; Yang, X.; Yang, L.; Ding, Y.; Nong, M.; Zhang, S.; et al. Actional mechanisms of active ingredients in functional food adlay for human health. Molecules 2022, 27, 4808. [Google Scholar] [CrossRef]
- Restani, P.; Colombo, F.; Biella, S.; Bani, C.; Mercogliano, F.; Di Lorenzo, C. Diet, polyphenols, and human evolution. Appl. Sci. 2022, 12, 7805. [Google Scholar] [CrossRef]
- Clemente-Villalba, J.; Burló, F.; Hernández, F.; Carbonell-Barrachina, Á.A. Valorization of wild edible plants as food ingredients and their economic value. Foods 2023, 12, 1012. [Google Scholar] [CrossRef]
- Prabawati, N.B.; Oktavirina, V.; Palma, M.; Setyaningsih, W. Edible flowers: Antioxidant compounds and their functional properties. Horticulturae 2021, 7, 66. [Google Scholar] [CrossRef]
- Kandylis, P. Phytochemicals and antioxidant properties of edible flowers. Appl. Sci. 2022, 12, 9937. [Google Scholar] [CrossRef]
- Motti, R.; Paura, B.; Cozzolino, A.; Falco, B.D. Edible flowers used in some countries of the Mediterranean basin: An ethnobotanical overview. Plants 2022, 11, 3272. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Yu, X.; Maninder, M.; Xu, B. Total phenolics and antioxidants profiles of commonly consumed edible flowers in China. Int. J. Food Prop. 2018, 21, 1524–1540. [Google Scholar] [CrossRef]
- Mateos-Maces, L.; Chávez-Servia, J.L.; Vera-Guzmán, A.M.; Aquino-Bolaños, E.N.; Alba-Jiménez, J.E.; Villagómez-González, B.B. Edible leafy plants from Mexico as sources of antioxidant compounds, and their nutritional, nutraceutical and antimicrobial potential: A review. Antioxidants 2020, 9, 541. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, H.; Wang, H.; Feng, S. Research progress in leaf related molecular breeding of Cucurbitaceae. Agronomy 2022, 12, 2908. [Google Scholar] [CrossRef]
- World Cucumber Production by Country. Available online: https://www.atlasbig.com/en-us/countries-cucumber-production (accessed on 30 March 2023).
- Grumet, R.; McCreight, J.D.; McGregor, C.; Weng, Y.; Mazourek, M.; Reitsma, K.; Labate, J.; Davis, A.; Fei, Z. Genetic resources and vulnerabilities of major cucurbit crops. Genes 2021, 12, 1222. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Mishra, A.; Hasan, S.M.; Usmani, A.; Ubaid, M.; Khan, N.; Saidurrahman, M. Biological and medicinal application of Cucumis sativus Linn.—Review of current status with future possibilities. J. Complement. Integr. Med. 2021, 19, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Abou-Zaid, M.M.; Lombardo, D.A.; Kite, G.C.; Grayer, R.J.; Veitch, N.C. Acylated flavone C-glycosides from Cucumis sativus. Phytochemistry 2001, 58, 167–172. [Google Scholar] [CrossRef] [PubMed]
- McNally, D.J.; Wurms, K.V.; Labbé, C.; Quideau, S.; Bélanger, R.R. Complex C-glycosyl flavonoid phytoalexins from Cucumis sativus. J. Nat. Prod. 2003, 66, 1280–1283. [Google Scholar] [CrossRef]
- Kai, H.; Baba, M.; Okuyama, T. Two new megastigmanes from the leaves of Cucumis sativus. Chem Pharm. Bull. 2007, 55, 133–136. [Google Scholar] [CrossRef]
- Sun, W.; Ma, N.; Huang, H. Photosynthetic contribution and characteristics of cucumber stems and petioles. BMC Plant Biol. 2021, 21, 454. [Google Scholar] [CrossRef] [PubMed]
- Krauze-Baranowska, M.; Cisowski, W. Flavonoids from some species of the genus Cucumis. Biochem. Syst. Ecol. 2001, 29, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Elshafie, H.S.; Camele, I.; Mohamed, A.A. A Comprehensive review on the biological, agricultural and pharmaceutical properties of secondary metabolites based-plant origin. Int. J. Mol. Sci. 2023, 24, 3266. [Google Scholar] [CrossRef]
- Li, M.; Qian, M.; Jiang, Q.; Tan, B.; Yin, Y.; Han, X. Evidence of flavonoids on disease prevention. Antioxidants 2023, 12, 527. [Google Scholar] [CrossRef]
- Martinez-Gonzalez, A.I.; Díaz-Sánchez, Á.G.; Rosa, L.A.D.L.; Vargas-Requena, C.L.; Bustos-Jaimes, I.; Alvarez-Parrilla, A.E. Polyphenolic compounds and digestive enzymes: In vitro non-covalent interactions. Molecules 2017, 22, 669. [Google Scholar] [CrossRef]
- Buchholz, T.; Melzig, M. Polyphenolic compounds as pancreatic lipase inhibitors. Planta Med. 2015, 81, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Birari, R.B.; Bhutani, K.K. Pancreatic lipase inhibitors from natural sources: Unexplored potential. Drug Discov. Today 2007, 12, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Rahim, A.T.M.A.; Takahashi, Y.; Yamaki, K. Mode of pancreatic lipase inhibition activity in vitro by some flavonoids and non-flavonoid polyphenols. Food Res. Int. 2015, 75, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Olennikov, D.N.; Kashchenko, N.I. New flavonoids from Cucumis sativus. Chem. Nat. Compd. 2023, 58, 356–359. [Google Scholar]
- Olennikov, D.N.; Kashchenko, N.I. Acylated flavonoids from Cucumis sativus inhibit activity of human pancreatic lipase. Appl. Biochem. Microbiol. 2023, 59, 487–493. [Google Scholar] [CrossRef]
- Xie, Y.; Guo, Q.-S.; Wang, G.-S. Preparative separation and purification of the total flavonoids in Scorzonera austriaca with macroporous resins. Molecules 2016, 21, 768. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, B.; Ye, M.; Chen, J.; Liu, Y.; Wang, C. An effective chromatography process for simultaneous purification and separation of total lignans and flavonoids from Valeriana amurensis. Molecules 2022, 27, 8598. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I. New C,O-glycosylflavones from the genus Silene. Chem. Nat. Compd. 2020, 56, 1026–1034. [Google Scholar] [CrossRef]
- Olennikov, D.N. Ecdysteroids, flavonoids, and phenylpropanoids from Silene nutans. Chem. Nat. Compd. 2019, 55, 127–130. [Google Scholar] [CrossRef]
- Olennikov, D.N. Chemical investigation of Anagallidium dichotomum and anticholinesterase activity of its components. Chem. Nat. Compd. 2013, 49, 977–979. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Chirikova, N.K. C-Glycosyl flavones from two Eastern Siberian species of Silene. Chem. Nat. Compd. 2019, 55, 642–647. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Chirikova, N.K. New C,O-glycosyl flavones from Melandrium divaricatum. Chem. Nat. Compd. 2019, 55, 1032–1038. [Google Scholar] [CrossRef]
- Pedrosa, A.M.; de Castro, W.V.; Castro, A.H.F.; Duarte-Almeida, J.M. Validated spectrophotometric method for quantification of total triterpenes in plant matrices. DARU 2020, 28, 281–286. [Google Scholar] [CrossRef]
- Thiex, N.; Novotny, L.; Crawford, A. Determination of ash in animal Feed: AOAC Official Method 942.05 revisited. J. AOAC Int. 2012, 95, 1392–1397. [Google Scholar] [CrossRef] [PubMed]
- Olennikov, D.N.; Chirikova, N.K.; Kashchenko, N.I.; Nikolaev, V.M.; Kim, S.-W.; Vennos, C. Bioactive phenolics of the genus Artemisia (Asteraceae): HPLC-DAD-ESI-TQ-MS/MS profile of the Siberian species and their inhibitory potential against α-amylase and α-glucosidase. Front. Pharmacol. 2018, 9, 756. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Zilfikarov, I.N.; Penzina, T.A. Use of microcolumn HPLC for analysis of aloenin in Aloe arborescens raw material and related drugs. Pharm. Chem. J. 2013, 47, 494–497. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Chemposov, V.V.; Chirikova, N.K. Polymeric compounds of lingonberry waste: Characterization of antioxidant and hypolipidemic polysaccharides and polyphenol-polysaccharide conjugates from Vaccinium vitis-idaea press cake. Foods 2022, 11, 2801. [Google Scholar] [CrossRef]
- Zhang, Q.W.; Lin, L.G.; Ye, W.C. Techniques for extraction and isolation of natural products: A comprehensive review. Chin. Med. 2018, 13, 20. [Google Scholar] [CrossRef]
- Kamaruzaman, S.; Nasir, N.M.; Mohd Faudzi, S.M.; Yahaya, N.; Mohamad Hanapi, N.S.; Wan Ibrahim, W.N. Solid-phase extraction of active compounds from natural products by nolecularly imprinted polymers: Synthesis and extraction parameters. Polymers 2021, 13, 3780. [Google Scholar] [CrossRef]
- Khoddami, A.; Wilkes, M.A.; Roberts, T.H. Techniques for analysis of plant phenolic compounds. Molecules 2013, 18, 2328–2375. [Google Scholar] [CrossRef]
- Rajan, L.; Palaniswamy, D.; Mohankumar, S.K. Targeting obesity with plant-derived pancreatic lipase inhibitors: A comprehensive review. Pharm. Res. 2020, 155, 104681. [Google Scholar] [CrossRef]
- Yang, M.H.; Chin, Y.-W.; Yoon, K.D.; Kim, J. Phenolic compounds with pancreatic lipase inhibitory activity from Korean yam (Dioscorea opposita). J. Enzyme Inhibit. Med. Chem. 2013, 29, 1–6. [Google Scholar] [CrossRef]
- Ma, L.-J.; Hou, X.-D.; Qin, X.-Y.; He, R.-J.; Yu, H.-N.; Hu, Q.; Guan, X.-Q.; Jia, S.-N.; Hou, J.; Lei, T.; et al. Discovery of human pancreatic lipase inhibitors from root of Rhodiola crenulata via integrating bioactivity-guided fractionation, chemical profiling and biochemical assay. J. Pharm. Anal. 2022, 12, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Ado, M.A.; Abas, F.; Mohammed, A.S.; Ghazali, H.M. Anti- and pro-lipase activity of selected medicinal, herbal and aquatic plants, and structure elucidation of an anti-lipase compound. Molecules 2013, 18, 14651–14669. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Karmase, A.; Jagtap, S.; Shekhar, R.; Bhutani, K.K. Pancreatic lipase inhibitory activity of cassiamin A, a bianthraquinone from Cassia siamea. Nat. Prod. Commun. 2013, 8, 195–198. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; Blázquez, M.A.; Garcia-Viguera, C.; Ferreres, F.; Tomás-Lorente, F. A comparative study of different amberlite XAD resins in flavonoid analysis. Phytochem. Anal. 1992, 3, 178–181. [Google Scholar] [CrossRef]
- Wang, X.; Su, J.; Chu, X.; Zhang, X.; Kan, Q.; Liu, R.; Fu, X. Adsorption and desorption characteristics of total flavonoids from Acanthopanax senticosus on macroporous adsorption resins. Molecules 2021, 26, 4162. [Google Scholar] [CrossRef]
- Che Zain, M.S.; Lee, S.Y.; Teo, C.Y.; Shaari, K. Adsorption/desorption characteristics and simultaneous enrichment of orientin, isoorientin, vitexin and isovitexin from hydrolyzed oil palm leaf extract using macroporous resins. Processes 2021, 9, 659. [Google Scholar] [CrossRef]
- Liu, J.; Meng, J.; Du, J.; Liu, X.; Pu, Q.; Di, D.; Chen, C. Preparative separation of flavonoids from goji berries by mixed-mode macroporous adsorption resins and effect on Aβ-expressing and anti-aging genes. Molecules 2020, 25, 3511. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Zhang, X.; Wang, C.; Hou, G.; Li, H. Adsorption properties and preparative separation of flavonoids from rhizoma Smilacis glabrae using macroporous resins. Separations 2022, 9, 431. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Gadimli, A.I.; Isaev, J.I.; Kashchenko, N.I.; Prokopyev, A.S.; Kataeva, T.N.; Chirikova, N.K.; Vennos, C. Caucasian Gentiana species: Untargeted LC-MS metabolic profiling, antioxidant and digestive enzyme inhibiting activity of six plants. Metabolites 2019, 9, 271. [Google Scholar] [CrossRef]
- Hostettmann, K.; Jacot-Guillarmod, A. Contribution à la phytochimie du genre Gentiana. X. Etude des composés flavoniques et xanthoniques dans les feuilles de Gentiana verna L. Helv. Chim. Acta 1975, 58, 130–132. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K.; Tankhaeva, L.M. Iridoids and flavonoids of four Siberian gentians: Chemical profile and gastric stimulatory effect. Molecules 2015, 20, 19172–19188. [Google Scholar] [CrossRef]
- Besson, E.; Dellamonica, G.; Chopin, J.; Markham, K.R.; Kim, M.; Koh, H.-S.; Fukami, H. C-Glycosylflavones from Oryza sativa. Phytochemistry 1985, 24, 1061–1064. [Google Scholar] [CrossRef]
- Herrera, A.Y.; Bain, J.F. Flavonoids of the Muhlenbergia montana complex. Biochem. Syst. Ecol. 1991, 19, 665–672. [Google Scholar] [CrossRef]
- Jacot-Guillarmod, A.; Duc, L.M.; Hostettmann, K. Contribution à la phytochimie du genre Gentian. XI. Identification de nouveaux dérivés de l’iso-orientine extraits des feuilles de Gentiana burseri Lapeyr. Helv. Chim. Acta 1975, 58, 1477–1481. [Google Scholar] [CrossRef]
- Ohkawa, M.; Kinjo, J.; Hagiwara, Y.; Hagiwara, H.; Ueyama, H.; Nakamura, K.; Ishikawa, R.; Ono, M.; Nohara, T. Three new anti-oxidative saponarin analogs from young green barley leaves. Chem. Pharm. Bull. 1998, 46, 1887–1889. [Google Scholar] [CrossRef]
- Xie, H.; Wang, T.; Matsuda, H.; Morikawa, T.; Yoshikawa, M.; Tani, T. Bioactive constituents from Chinese natural medicines. XV. Inhibitory effect on aldose reductase and structures of saussureosides A and B from Saussurea medusa. Chem. Pharm. Bull. 2005, 53, 1416–1422. [Google Scholar] [CrossRef]
- Markham, K.R.; Mitchell, K.A. The mis-identification of the major antioxidant Flavonoids in young barley (Hordeum vulgare) leaves. Z. Naturforsch. C 2003, 58, 53–56. [Google Scholar] [CrossRef]
- Chulia, A.J.; Mariotte, A.M. Nouvelles C-glucosylflavones chez Gentiana pedicellata. J. Nat. Prod. 1985, 48, 480–483. [Google Scholar] [CrossRef]
- Williams, C.A.; Murray, B.G. Flavonoid variation in the genus Briza. Phytochemistry 1972, 11, 2507–2512. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Khandy, M.T.; Chirikova, N.K. Oriental strawberry metabolites: LC–MS profiling, antioxidant potential, and postharvest changes of Fragaria orientalis fruits. Horticulturae 2022, 8, 975. [Google Scholar] [CrossRef]
- Li, Q.; Wang, L.; Dai, P.; Zeng, X.; Qi, X.; Zhu, L.; Yan, T.; Wang, Y.; Lu, L.; Hu, M.; et al. A combined strategy of mass fragmentation, post-column cobalt complexation and shift in ultraviolet absorption spectra to determine the uridine 5′-diphospho-glucuronosyltransferase metabolism profiling of flavones after oral administration of a flavone mixture in rats. J. Chromatogr. A 2015, 1395, 116–128. [Google Scholar] [CrossRef]
- Piasecka, A.; Sawikowska, A.; Krajewski, P.; Kachlicki, P. Combined mass spectrometric and chromatographic methods for in-depth analysis of phenolic secondary metabolites in barley leaves. J. Mass Spectrom. 2015, 50, 513–532. [Google Scholar] [CrossRef] [PubMed]
- Goetz, M.; Jacot-Guillarmod, A. Contribution à la phytochimie du genre Gentiana, XX. Identification de nouveaux di-O-glucosides de C-glucosylflavones dans Gentiana asclepiadea L. Helv. Chim. Acta 1977, 60, 1322–1324. [Google Scholar] [CrossRef]
- Krauze-Baranowska, M.; Cisowski, W. High-performance liquid chromatographic determination of flavone C-glycosides in some species of the Cucurbitaceae family. J. Chromatogr. A 1994, 675, 240–243. [Google Scholar] [CrossRef]
- Chen, G.L.; Chen, S.G.; Xie, Y.Q.; Chen, F.; Zhao, Y.Y.; Luo, C.X.; Gao, Y.Q. Total phenolic, flavonoid and antioxidant activity of 23 edible flowers subjected to in vitro digestion. J. Funct. Foods 2015, 17, 243–259. [Google Scholar] [CrossRef]
- Pinedo-Espinoza, J.M.; Gutiérrez-Tlahque, J.; Santiago-Saenz, Y.O.; Aguirre-Mancilla, C.L.; Reyes-Fuentes, M.; López-Palestina, C.U. Nutritional composition, bioactive compounds and antioxidant activity of wild edible flowers consumed in semiarid regions of Mexico. Plant Foods Hum. Nutr. 2020, 75, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.M.; Lee, S.S.; Chung, B.Y.; Cho, J.-Y.; Lee, I.C.; Ahn, S.R.; Jang, S.J.; Kim, T.H. Pancreatic lipase inhibition by C-glycosidic flavones isolated from Eremochloa ophiuroides. Molecules 2010, 15, 8251–8259. [Google Scholar] [CrossRef]
- Li, M.; Chen, Y.; Ruan, J.; Wang, W.; Chen, J.; Zhang, Q. Structure-activity relationship of dietary flavonoids on pancreatic lipase. Curr. Res. Food Sci. 2023, 6, 100424. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.-T.; Liu, X.-T.; Chen, Q.-X.; Shi, Y. Lipase inhibitors for obesity: A review. Biomed. Pharmacother. 2020, 128, 110314. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Que, Q.; Pan, J.; Guo, J. Study of the interaction between icariin and human serum albumin by fluorescence spectroscopy. J. Molec. Struct. 2008, 881, 132–138. [Google Scholar] [CrossRef]
- Huang, R.; Zhang, Y.; Shen, S.; Zhi, Z.; Chen, H.; Chen, S.; Ye, X. Antioxidant and pancreatic lipase inhibitory effects of flavonoids from different citrus peel extracts: An in vitro study. Food Chem. 2020, 326, 126785. [Google Scholar] [CrossRef]
- Nakai, M.; Fukui, Y.; Asami, S.; Toyoda-Ono, Y.; Iwashita, T.; Shibata, H.; Mitsunaga, T.; Hashimoto, F.; Kiso, Y. Inhibitory effects of oolong tea polyphenols on pancreatic lipase in vitro. J. Agric. Food Chem. 2005, 53, 4593–4598. [Google Scholar] [CrossRef]
- Batubara, I.; Kuspradini, H.; Muddathir, A.M.; Mitsunaga, T. Intsia palembanica wood extracts and its isolated compounds as Propionibacterium acnes lipase inhibitor. J. Wood Sci. 2014, 60, 169–174. [Google Scholar] [CrossRef]
- Grassi, D.; Desideri, G.; Ferri, C. Flavonoids: Antioxidants against atherosclerosis. Nutrients 2010, 2, 889–902. [Google Scholar] [CrossRef]
- Fernandes, I.; Pérez-Gregorio, R.; Soares, S.; Mateus, N.; De Freitas, V. Wine flavonoids in health and disease prevention. Molecules 2017, 22, 292. [Google Scholar] [CrossRef] [PubMed]
- Suen, J.; Thomas, J.; Kranz, A.; Vun, S.; Miller, M. Effect of flavonoids on oxidative stress and inflammation in adults at risk of cardiovascular disease: A systematic review. Healthcare 2016, 4, 69. [Google Scholar] [CrossRef]
- Wang, M.; Firrman, J.; Liu, L.; Yam, K. A Review on flavonoid apigenin: Dietary intake, ADME, antimicrobial effects, and interactions with human gut microbiota. BioMed Res. Int. 2019, 2019, 7010467. [Google Scholar] [CrossRef]
- Wang, Z.; Zeng, M.; Wang, Z.; Qin, F.; Chen, J.; He, Z. Dietary luteolin: A narrative review focusing on its pharmacokinetic properties and effects on glycolipid metabolism. J. Agricult. Food Chem. 2021, 69, 1441–1454. [Google Scholar] [CrossRef] [PubMed]
- Kozłowska, A.; Szostak-Węgierek, D. Flavonoids—Food Sources, Health Benefits, and Mechanisms Involved. In Bioactive Molecules in Food. Reference Series in Phytochemistry; Mérillon, J.M., Ramawat, K., Eds.; Springer: Cham, Switzerland, 2019; pp. 53–78. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).