
A Theoretical Model of Mitochondrial ATP Synthase Deficiencies. The Role of Mitochondrial Carriers


Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Determination of EFMs with 2-Oxoglutarate (AKG) as a Respiratory Substrate
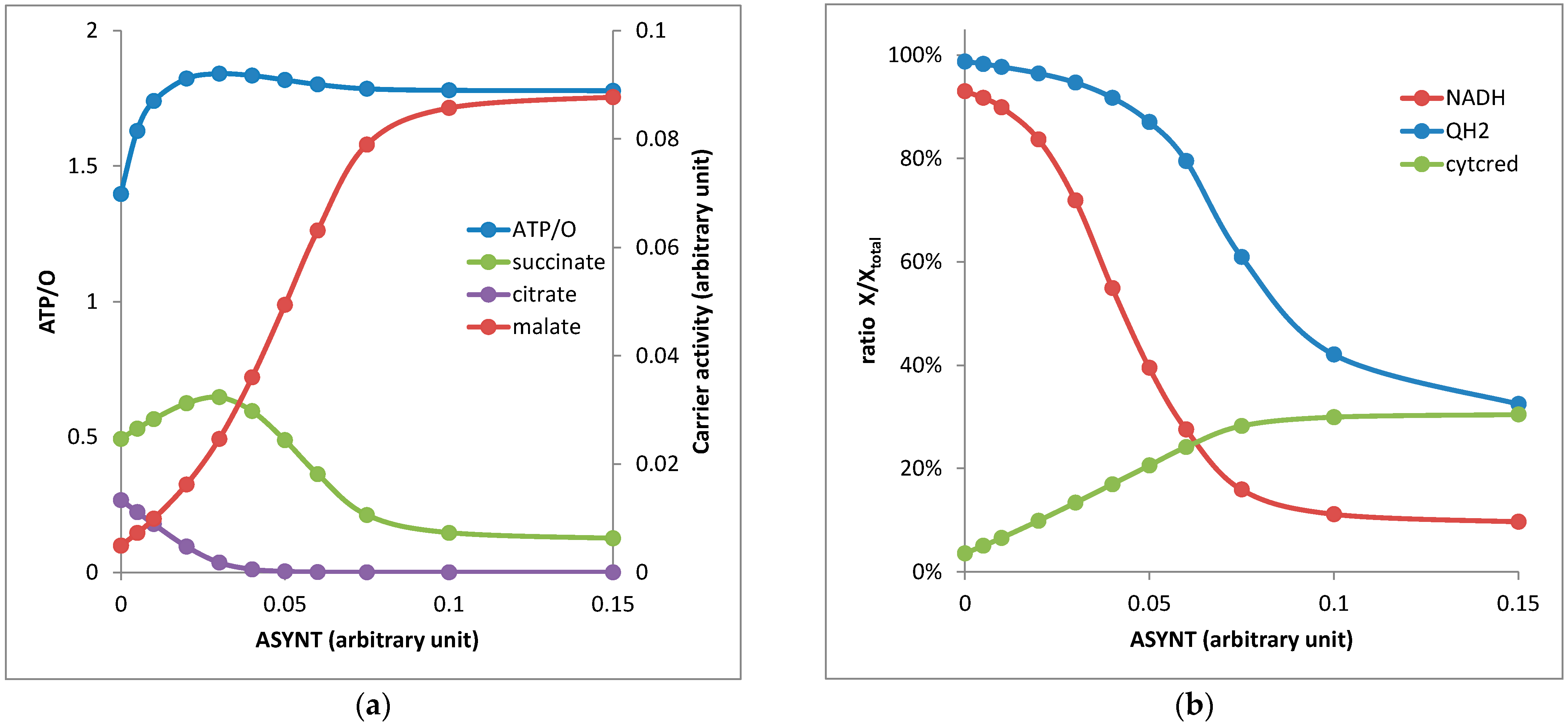
3.2. Dynamics of Oxygen Consumption and ATP Generation and Determination of the Ratio ATP/O as a Function of ATP Synthase Activity
- (1)
- As the thermodynamic span between the pair NADH/NAD and QH2/Q was much greater than the span between succinate/fumarate and QH2/Q, NADH was oxidized before QH2 (Figure 2b) and the accumulation of QH2 blocked the respiratory complex II (RCII or SDH).
- (2)
- When the ratio NADH/NAD was high (low ATP synthase activity), the reversion of IDH3 associated with NADH reoxidation was possible.
- (3)
- When both NADH/NAD and QH2/Q ratios were low (high activity of the respiratory chain and ATP synthase), RCII was active and succinate was easily transformed in fumarate competing with the output of succinate against AKG through Odc1p. The AKG/succinate exchange thus decreased and was largely replaced by the AKG/malate exchange (Figure 2a; high ASYNT activity).
3.3. Decomposition of the Steady States in EFMs
3.4. Overexpression of Odc1p
4. Discussion
4.1. EFMs
4.2. Kinetic Model and Decomposition of EFMs
4.3. Odc1p Rescue
4.4. Implications for NARP Syndrome Human Odc1p Counterpart
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AKG | α-ketoglutarate/2-oxoglutarate |
| ASYNT-ATP | ATP synthesized by the mitochondrial ATP-synthase |
| PMF | Proton motive force (ΔµH+) |
| SLP | Substrate-level phosphorylation |
| SLP-ATP | ATP synthesized by Succinyl-CoA synthetase (succinate thiokinase) |
References
- Zhou, A.; Rohou, A.; Schep, D.G.; Bason, J.V.; Montgomery, M.G.; Walker, J.E.; Grigorieff, N.; Rubinstein, J.L. Structure and conformational states of the bovine mitochondrial ATP synthase by cryo-EM. eLife 2015, 4, e10180. [Google Scholar] [CrossRef]
- Dautant, A.; Velours, J.; Giraud, M.-F. Crystal Structure of the Mg·ADP-inhibited State of the Yeast F1c10-ATP Synthase. J. Biol. Chem. 2010, 285, 29502–29510. [Google Scholar] [CrossRef] [Green Version]
- Spikes, T.E.; Montgomery, M.G.; Walker, J.E. Structure of the dimeric ATP synthase from bovine mitochondria. Proc. Natl. Acad. Sci. USA 2020, 117, 23519–23526. [Google Scholar] [CrossRef]
- Guo, H.; Rubinstein, J.L. Cryo-EM of ATP synthases. Curr. Opin. Struct. Biol. 2018, 52, 71–79. [Google Scholar] [CrossRef]
- Dautant, A.; Meier, T.; Hahn, A.; Tribouillard-Tanvier, D.; Di Rago, J.-P.; Kucharczyk, R. ATP Synthase Diseases of Mitochondrial Genetic Origin. Front. Physiol. 2018, 9, 329. [Google Scholar] [CrossRef]
- Su, X.; Dautant, A.; Rak, M.; Godard, F.; Ezkurdia, N.; Bouhier, M.; Bietenhader, M.; Mueller, D.M.; Kucharczyk, R.; di Rago, J.-P.; et al. The pathogenic m.8993 T > G mutation in mitochondrial ATP6 gene prevents proton release from the subunit c-ring rotor of ATP synthase. Hum. Mol. Genet. 2021, 30, 381–392. [Google Scholar] [CrossRef]
- Su, X.; Dautant, A.; Godard, F.; Bouhier, M.; Zoladek, T.; Kucharczyk, R.; Di Rago, J.-P.; Tribouillard-Tanvier, D. Molecular Basis of the Pathogenic Mechanism Induced by the m.9191T>C Mutation in Mitochondrial ATP6 Gene. Int. J. Mol. Sci. 2020, 21, 5083. [Google Scholar] [CrossRef]
- Su, X.; Rak, M.; Tetaud, E.; Godard, F.; Sardin, E.; Bouhier, M.; Gombeau, K.; Caetano-Anollés, D.; Salin, B.; Chen, H.; et al. Deregulating mitochondrial metabolite and ion transport has beneficial effects in yeast and human cellular models for NARP syndrome. Hum. Mol. Genet. 2019, 28, 3792–3804. [Google Scholar] [CrossRef] [PubMed]
- Rak, M.; Tetaud, E.; Duvezin-Caubet, S.; Ezkurdia, N.; Bietenhader, M.; Rytka, J.; di Rago, J.-P. A Yeast Model of the Neurogenic Ataxia Retinitis Pigmentosa (NARP) T8993G Mutation in the Mitochondrial ATP Synthase-6 Gene. J. Biol. Chem. 2007, 282, 34039–34047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwimmer, C.; Lefebvre-Legendre, L.; Rak, M.; Devin, A.; Slonimski, P.P.; di Rago, J.-P.; Rigoulet, M. Increasing Mitochondrial Substrate-level Phosphorylation Can Rescue Respiratory Growth of an ATP Synthase-deficient Yeast. J. Biol. Chem. 2005, 280, 30751–30759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butow, R.A.; Avadhani, N.G. Mitochondrial Signaling: The Retrograde Response. Mol. Cell 2004, 14, 1–15. [Google Scholar] [CrossRef]
- Palmieri, L.; Agrimi, G.; Runswick, M.J.; Fearnley, I.M.; Palmieri, F.; Walker, J. Identification in Saccharomyces cerevisiae of Two Isoforms of a Novel Mitochondrial Transporter for 2-Oxoadipate and 2-Oxoglutarate. J. Biol. Chem. 2001, 276, 1916–1922. [Google Scholar] [CrossRef] [Green Version]
- Lebiedzinska, M.; Karkucińska-Więckowska, A.; Wojtala, A.; Suski, J.M.; Szabadkai, G.; Wilczynski, G.; Wlodarczyk, J.; Diogo, C.V.; Oliveira, P.J.; Tauber, J.; et al. Disrupted ATP synthase activity and mitochondrial hyperpolarisation-dependent oxidative stress is associated with p66Shc phosphorylation in fibroblasts of NARP patients. Int. J. Biochem. Cell Biol. 2013, 45, 141–150. [Google Scholar] [CrossRef]
- Hoops, S.; Sahle, S.; Gauges, R.; Lee, C.; Pahle, J.; Simus, N.; Singhal, M.; Xu, L.; Mendes, P.; Kummer, U. COPASI—A COmplex PAthway SImulator. Bioinformatics 2006, 22, 3067–3074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohnensack, R. Control of energy transformation in mitochondria. Analysis by a quantitative model. Biochim. et Biophys. Acta (BBA) Bioenerg. 1981, 634, 203–218. [Google Scholar] [CrossRef]
- Mitchell, P. Vectorial Chemistry and the Molecular Mechanics of Chemiosmotic Coupling: Power Transmission by Proticity. Biochem. Soc. Trans. 1976, 4, 399–430. [Google Scholar] [CrossRef]
- Toth, A.; Meyrat, A.; Stoldt, S.; Santiago, R.; Wenzel, D.; Jakobs, S.; von Ballmoos, C.; Ott, M. Kinetic coupling of the respiratory chain with ATP synthase, but not proton gradients, drives ATP production in cristae membranes. Proc. Natl. Acad. Sci. USA 2020, 117, 2412–2421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, S.; Hilgetag, C.C. On elementary flux modes in biochemical reaction systems at steady state. J. Biol. Syst. 1994, 2, 165–182. [Google Scholar] [CrossRef]
- Pfeiffer, T.; Sanchez-Valdenebro, I.; Nuño, J.C.; Montero, F.; Schuster, S. METATOOL: For studying metabolic networks. Bioinformatics 1999, 15, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Rigoulet, M.; Velours, J.; Guerin, B. Substrate-level phosphorylation in isolated yeast mitochondria. JBIC J. Biol. Inorg. Chem. 1985, 153, 601–607. [Google Scholar] [CrossRef]
- Schuster, S.; Dandekar, T.; Fell, D.A.; Schuster, S.; Dandekar, T.; Fell, D.A.; Schuster, S.; Dandekar, T.; Fell, D.A.; Schuster, S.; et al. Detection of elementary flux modes in biochemical networks: A promising tool for pathway analysis and metabolic engineering. Trends Biotechnol. 1999, 17, 53–60. [Google Scholar] [CrossRef]
- Chinopoulos, C. Acute sources of mitochondrial NAD+ during respiratory chain dysfunction. Exp. Neurol. 2020, 327, 113218. [Google Scholar] [CrossRef]
- Valle, A.; Oliver, J.O.; Roca, P. Role of Uncoupling Proteins in Cancer. Cancers 2010, 2, 567–591. [Google Scholar] [CrossRef]
- Samudio, I.; Fiegl, M.; Andreeff, M. Mitochondrial Uncoupling and the Warburg Effect: Molecular Basis for the Reprogramming of Cancer Cell Metabolism: Figure 1. Cancer Res. 2009, 69, 2163–2166. [Google Scholar] [CrossRef] [Green Version]
- Tilques, M.D.T.D.; Tribouillard-Tanvier, D.; Tétaud, E.; Testet, E.; di Rago, J.-P.; Lasserre, J.-P. Overexpression of mitochondrial oxodicarboxylate carrier (ODC1) preserves oxidative phosphorylation in a yeast model of the Barth syndrome. Dis. Model. Mech. 2017, 10, 439–450. [Google Scholar] [CrossRef] [Green Version]
- Chinopoulos, C.; Seyfried, T.N. Mitochondrial Substrate-Level Phosphorylation as Energy Source for Glioblastoma: Review and Hypothesis. ASN Neuro 2018, 10, 1759091418818261. [Google Scholar] [CrossRef] [PubMed]
- Sgarbi, G.; Casalena, G.A.; Baracca, A.; Lenaz, G.; DiMauro, S.; Solaini, G. Human NARP Mitochondrial Mutation Metabolism Corrected With α-Ketoglutarate/Aspartate. Arch. Neurol. 2009, 66, 951–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Kirk, K.; Shurubor, Y.I.; Zhao, D.; Arreguin, A.; Shahi, I.; Valsecchi, F.; Primiano, G.; Calder, E.L.; Carelli, V.; et al. Rewiring of Glutamine Metabolism Is a Bioenergetic Adaptation of Human Cells with Mitochondrial DNA Mutations. Cell Metab. 2018, 27, 1007–1025.e5. [Google Scholar] [CrossRef] [Green Version]
- Fiermonte, G.; Dolce, V.; Palmieri, L.; Ventura, M.; Runswick, M.J.; Palmieri, F.; Walker, J.E. Identification of the Human Mitochondrial Oxodicarboxylate Carrier. Bacterial Expression, Reconstitution, Functional Characterization, Tissue Distribution, and Chromosomal Location. J. Biol. Chem. 2001, 276, 8225–8230. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No Odc1p Leak | ATP Synthesis Rate | Respiration Rate | ATP/O |
|---|---|---|---|
| WT | 0.301 | 0.179 | 1.79 |
| NARP | 0.030 | 0.021 | 1.40 |
| NARP + Odc1p (×10) | 0.068 | 0.026 | 2.65 |
| Odc1 Leak | ATP Synthesis Rate | Respiration Rate | ATP/O |
|---|---|---|---|
| WT | 0.300 | 0.169 | 1.77 |
| NARP | 0.031 | 0.034 | 1.11 |
| NARP + Odc1p (×10) | 0.084 | 0.047 | 1.79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazat, J.-P.; Devin, A.; Yoboue, E.; Ransac, S. A Theoretical Model of Mitochondrial ATP Synthase Deficiencies. The Role of Mitochondrial Carriers. Processes 2021, 9, 1424. https://doi.org/10.3390/pr9081424
Mazat J-P, Devin A, Yoboue E, Ransac S. A Theoretical Model of Mitochondrial ATP Synthase Deficiencies. The Role of Mitochondrial Carriers. Processes. 2021; 9(8):1424. https://doi.org/10.3390/pr9081424
Chicago/Turabian StyleMazat, Jean-Pierre, Anne Devin, Edgar Yoboue, and Stéphane Ransac. 2021. "A Theoretical Model of Mitochondrial ATP Synthase Deficiencies. The Role of Mitochondrial Carriers" Processes 9, no. 8: 1424. https://doi.org/10.3390/pr9081424
APA StyleMazat, J.-P., Devin, A., Yoboue, E., & Ransac, S. (2021). A Theoretical Model of Mitochondrial ATP Synthase Deficiencies. The Role of Mitochondrial Carriers. Processes, 9(8), 1424. https://doi.org/10.3390/pr9081424