1. Introduction
Clinacanthus nutans (Burm. f.) Lindau from the Acanthaceae family is a small shrub which is naturally found in tropical and subtropical Asian countries, including Malaysia, Thailand, Indonesia, Vietnam, and China [
1]. In Malaysia, the plant is commonly known as Belalai Gajah and Sabah snake grass [
2]. The aerial part of this plant, i.e., the leaf (
Figure 1) has been shown to possess a vast spectrum of pharmacological activities, in which the anti-proliferative activities on various human cancer cell lines have been extensively studied. For instance,
C. nutans leaf extracted with chloroform, methanol, and water has been reported to demonstrate anti-proliferative effects against various human cancer cell lines, such as the liver hepatocellular carcinoma cell line (HepG2), neuroblastoma cell line (IMR-32), lung cancer cell line (NCI-H23), gastric cancer cell line (SNU-1), colon adenocarcinoma cell line (LS-174T), erythroleukemia cell line (K-562), human cervical cancer cell line (HeLa), and Burkitt’s lymphoma cell line (Raji) [
3]. On the other hand, the leaf extracted with ethanol and ethyl acetate has also been reported to exhibit strong cytotoxic effects against the estrogen-dependent human breast cancer cell line, MCF-7 [
4]. Meanwhile, Fong and colleagues have collected
C. nutans leaves from different countries, such as Malaysia, Thailand and Vietnam. They extracted the collected leaves with methanol and found that the leaf extracts possess cytotoxicity and apoptotic activities against the human skin cancer cell line, D24 melanoma cells [
5]. In addition to the anti-proliferative effects on various cancer cell lines, the aerial parts of the plant have also been reported to exhibit significant antitumor and apoptotic activities on the in vivo mice cancer model [
6]. While most of the anticancer studies on this plant are focused on the aerial part, our group have conducted an anti-proliferative study on the underground part, i.e., the root (
Figure 1) [
7]. The crude methanolic and ethyl acetate extracts of
C. nutans roots were found to exhibit anti-proliferative and apoptotic effects on breast cancer (MCF-7) and cervical cancer (HeLa) cell lines [Teoh].
Owing to its promising anti-proliferative activities, several attempts have been conducted to isolate the anti-proliferative compounds and/or active fractions from the
C. nutans leaf. Huang and colleagues [
6] have extracted the aerial parts of
C. nutans with ethanol, followed by further fractionation on a Diaion HP-20 macroporous adsorption resin to produce a 30% ethanol fraction (CN30) and other fractions. The CN30 fraction was found to be able to induce inhibition in tumor size and weight of the HepA xenograft model mouse (hepatoma cell-injected mice). The active components in CN30 were therefore further fractionated and purified. High-performance liquid chromatography (HPLC) and mass spectrometry (LC/MS/MS) analyses revealed that there were seven active compounds present in CN30. The seven identified active components were shaftoside, apigenin 6,8-C-α-L-pyranarabinoside, orientin, isoorientin, vitexin, isovitexin, and gallic acid. After that, the same research group conducted another study to isolate a novel polysaccharide–peptide complex, which they named CNP-1-2, by a series of complex purification methods from
C. nutans leaves [
8]. This CNP-1-2 complex was found to show the strongest growth inhibition effect on the human gastric cancer cell line (SGC-7901) and was able to stimulate the activation of macrophages. On the other hand, another research group led by Mutazah and colleagues [
9] has managed to fractionate two active fractions from the bark powder of
C. nutans. These two fractions, which they named A12 and A17, were found to contain sulfur-containing compounds, including entadamide C and clinamide D, that possessed strong cytotoxic activities against MDA-MB-231 and MCF-7 breast cancer cell lines.
From the studies above, it can be postulated that the isolation of anti-proliferative compounds from C. nutans is not easy. This may be due to the complexities of the plant’s nature, and also various fractionation/chromatographic steps which need to be optimized. Our study therefore aimed to pre-screen (profile) the volatile compounds present in every part of the plants first, followed by the selection and isolation of target compound. The details of the study are described in the subsequent sections.
2. Materials and Methods
2.1. Solvents, Chemicals and Cell Lines
All the solvents used in this study were of analytical and/or high performance liquid chromatography (HPLC) grade, and purchased from Fisher Scientific (Pittsburgh, PA, USA). The solvents used were methanol, ethyl acetate and dimethylsulfoxide (DMSO). The authentic reference standard, lupeol (L5632-25 mg, ≥94% purity) for characterization purposes was obtained from Sigma-Aldrich (St. Louis, MO, USA). Roswell Park Memorial Institute (RPMI) 1640, Phosphate-Buffered Saline (PBS), 2.5 g/L trypsin-1mmol/L EDTA, and 0.5% trypan blue stain solution were purchased from Nacalai Tesque (Kyoto, Japan), whereas the Fetal Bovine Serum (FBS) was purchased from JR Scientific (Woodland, CA, USA). The cell proliferation kit was purchased from Roche Diagnostics (Mannheim, Germany).
2.2. Plant Materials
Clinacanthus nutans (Burm. f.) Lindau plants were purchased from herbal suppliers at a local market in Kota Kinabalu, Sabah, Malaysia. The plants were verified by a botanist from the Faculty of Science and Natural Resources, Assoc. Prof. Dr. Berhaman Ahmad, and a voucher specimen (ACCN 001/2013) was deposited in the herbarium of Universiti Malaysia Sabah. The plants were first cleaned thoroughly with tap water to remove the dirt, followed by rinsing several times with distilled water and dried with a paper towel. The leaves, stems and roots were excised from the plants, freeze-dried for five days and ground into powder using a heavy-duty blender. The plant samples in powder form were stored at −80 °C until further analysis. Some of the stems of the purchased plants were re-grown in soil for a duration of 6 months for photo shooting purposes.
2.3. Extraction of C. nutans Leaf, Stem, and Root for Volatile Profiling
Approximately 0.5 g of each freeze-dried C. nutans leaf, stem, and root sample was weighed and added with 10 mL of absolute ethyl acetate, followed by maceration at 37 °C with 180 rpm agitation for three days in dark conditions. After three days of extraction, the plant extracts were filtered with Whatman filter paper, followed by drying under vacuum pressure by using a rotary evaporator at 40 °C, and then stored at −80 °C until further analysis. For each plant’s part, the whole extraction process was repeated another two times to produce three replicates (n = 3) per leaf, stem and root.
2.4. Volatile Profiling of the Crude Leaf, Stem, and Root Extracts Using GC-MS
The crude
C. nutans leaf, stem, and root ethyl acetate extracts were re-dissolved in HPLC-grade ethyl acetate to appropriate concentrations and filtered using 0.22 μm syringe filters prior to GC-MS analyses. The GC-MS analysis was carried out according to [
9], with some modifications. Briefly, 1 µL aliquot of each sample was injected into the GC-MS equipment (GC model 7890 and MS model 5975C; from Agilent Technologies, Santa Clara, CA, USA). An HP-5ms capillary column (30 m × 0.250 mm inner diameter × 0.25 µm of film thickness) from Agilent Technologies was used for separation. GC-MS was performed in a splitless mode and pure helium was used as the carrier gas with a flow rate of 1 mL/min. The MS source was pre-set to 230 °C and MS quad temperature was recorded at 150 °C. The injector temperature was set to 250 °C with an injection volume of 1 µL per sample. The oven temperature was programmed from 220 °C for 10 min and continuously increased at a rate of 5 °C/min until a temperature of 300 °C was reached and then retained for 10 min. The total run time was 36 min. The post-run was fixed at 200 °C for 5 min, and the ion source temperature was constant at 200 °C. The mass analyzer was performed in a full scan mode from
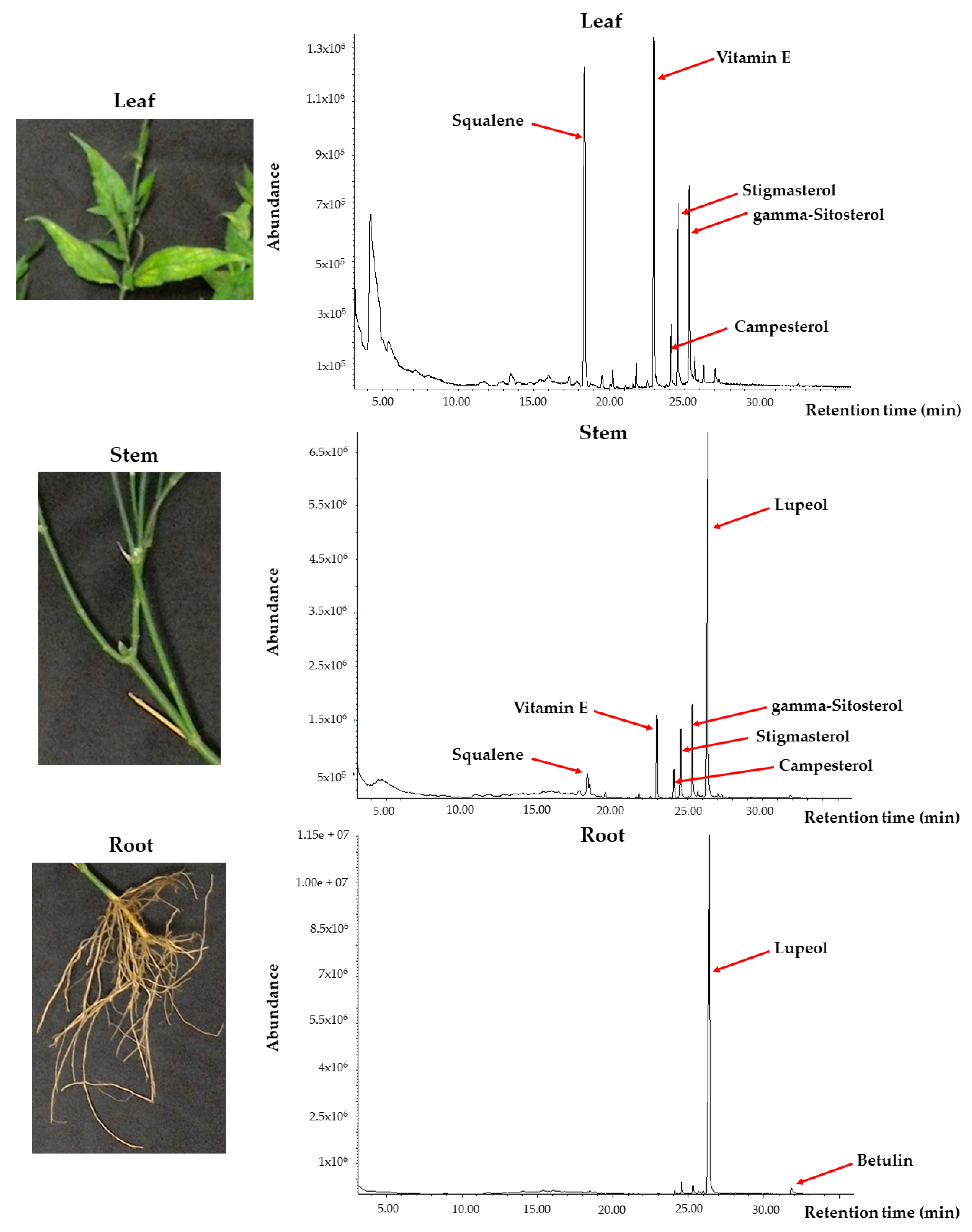
m/z 40 to 550, and mass spectra were obtained at EI = 70 eV. The identification of the compounds was performed by spectra matching using the National Institute Standard and Technology (NIST) library version 11 (NIST11). The leaf, stem, and root profiles obtained were compared to select for the most suitable plant’s part to be used for the subsequent isolation of target compound.
2.5. Extraction of C. nutans Root for the Isolation of Lupeol
The selected plant part in this study was the roots, and the target compound was lupeol. First of all, 80 g of freeze-dried C. nutans roots were weighed and added into a conical flask containing 200 mL of absolute ethyl acetate. The extract was put inside a rotary shaker and macerated at 37 °C with 180 rpm agitation for a consecutive five days in dark conditions. After five days of extraction, the crude root extract was filtered with Whatman filter paper, transferred to a new flask, and stored at 4 °C. A repeated extraction was performed on the remaining root residue in the extraction flask with the addition of another 200 mL of ethyl acetate but only for three days. The second crude root extract was filtered and combined with the first extract, followed by drying under vacuum pressure using a rotary evaporator at 40 °C and stored at −80 °C prior to the recrystallization step.
2.6. Recrystallization of C. nutans Root Extract for the Isolation of Lupeol
The dried C. nutans crude root extract in Part 2.5 above was re-dissolved in 100 mL of absolute methanol and added into an Erlenmeyer flask, followed by heat-stirring on a hot plate until all the extract was fully dissolved and the methanol solvent boiled. The Erlenmeyer flask was then left to cool down inside a huge glass beaker filled with ice. While cooling down, the inner side of the Erlenmeyer flask was gently stirred by using a glass rod to induce crystal formation (nucleation). After ten minutes, the formed crystals were filtered out from the methanol solvent using a Buchner funnel under vacuum pressure. The filtered crystals were collected and left to dry inside a laminar flow for several minutes until a white amorphous powder was obtained. The collected white amorphous powder was weighed, followed by identification and characterization using various analytical instruments.
2.7. GC-MS Analyses and Comparisons of the Isolated and Authentic Reference Standard Lupeols
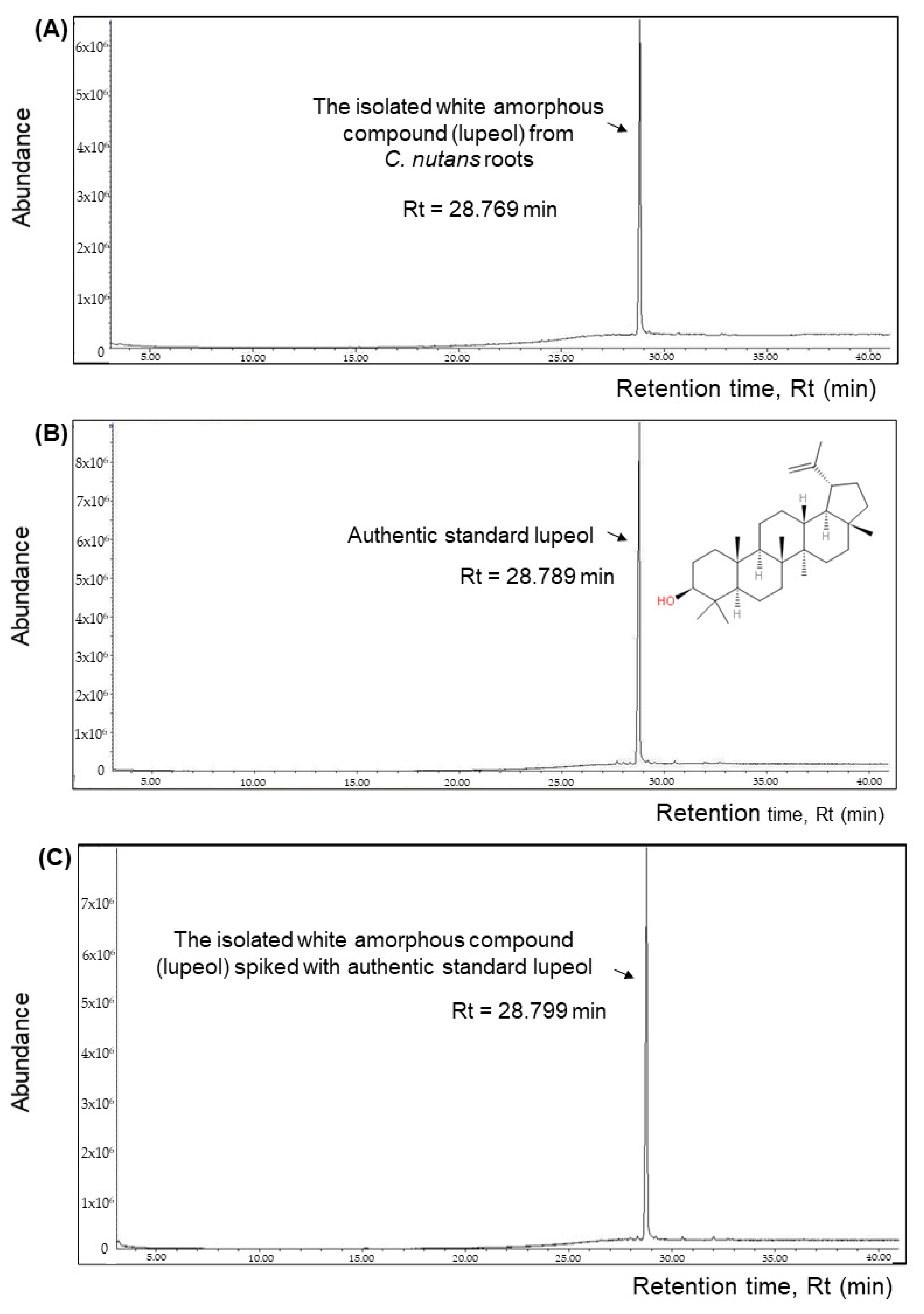
The isolated white amorphous compound from C. nutans roots and the purchased authentic standard lupeol were dissolved in HPLC-grade ethyl acetate to appropriate concentrations and were filtered using 0.22 μm syringe filters prior to GC-MS analyses. The GC-MS analysis was carried out according to Part 2.4, with some modifications on the ramping program. Briefly, the oven temperature was programmed from 220 °C for 10 min and continuously increased at a rate of 5 °C/min until a temperature of 300 °C was reached and then retained for 10 min. The total run time was 41 min. The identification of the compound was determined using the NIST11 library (Cas number for lupeol is 000545-47-1, entry number in library is #221171) and also compared with the MS spectra of the reference standard lupeol. In addition, further confirmation of the identity and purity of lupeol isolated from C. nutans roots was carried out by spiking the standard lupeol into the isolated lupeol sample, followed by a re-analysis with GC-MS to observe if only one overlapped peak (lupeol) was present in the chromatogram or not.
2.8. Characterization of the Isolated Lupeol from C. nutans Roots
Firstly, the melting point of the isolated lupeol was determined with SMP40 Automatic Melting Point Apparatus (Stuart, UK). Next, the infrared (IR) spectra of the isolated and standard lupeol were determined using a Cary 630 FTIR Spectrometer (Agilent Technologies, CA, USA). Finally, structural elucidation of the isolated compound was determined using 1D-FT-NMR in 600 MHz (Bruker, Germany) for both 1H-NMR and 13C-NMR.
2.9. Anti-Proliferative Activities of the Isolated Lupeol Compared to the Authentic Standard Lupeol
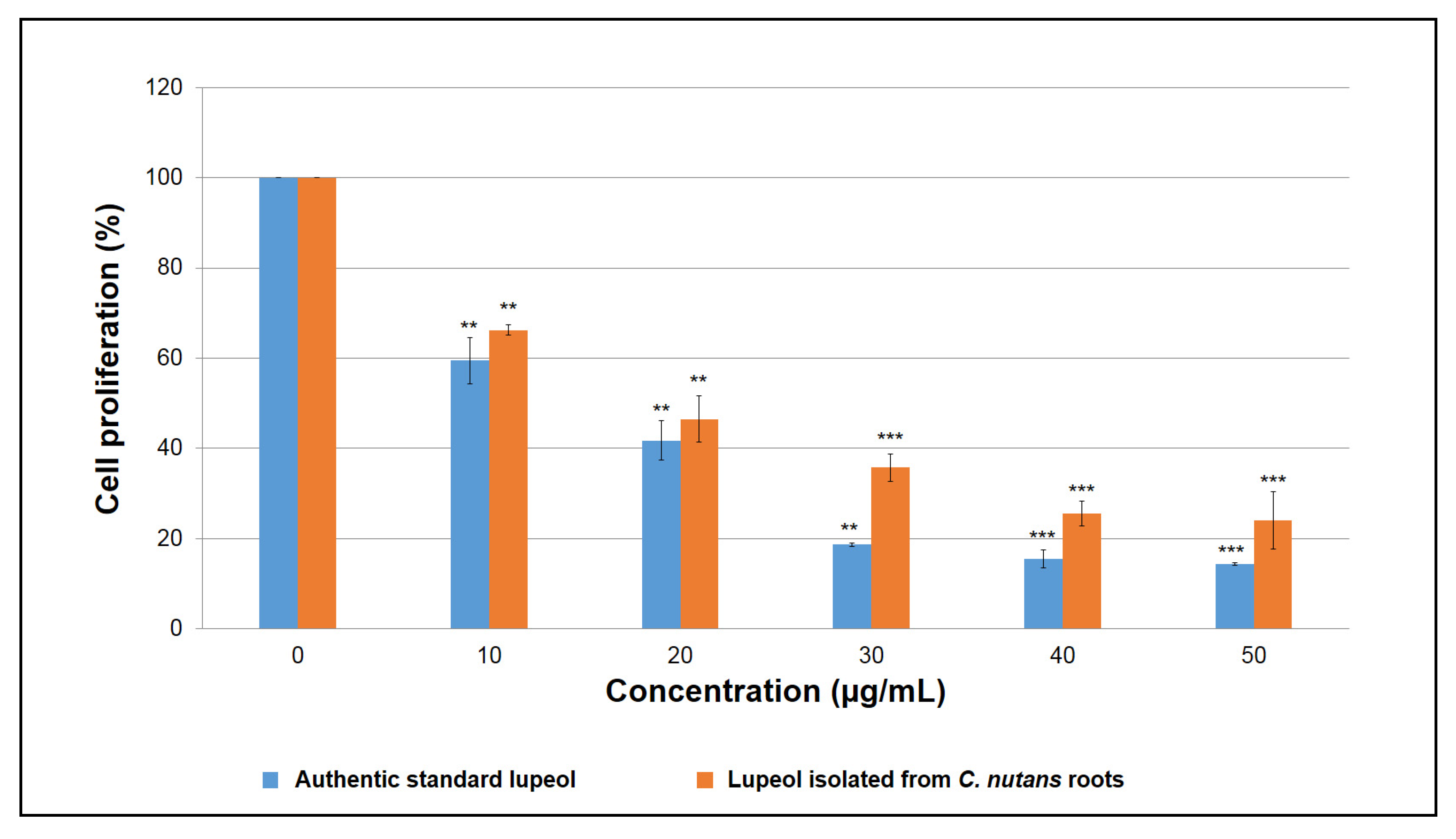
The anti-proliferative activities of isolated and standard lupeol were determined using MTT assay, which was performed using the Cell Proliferation Kit I (Roche Diagnostics, Mannheim, Germany) according to the manufacturer’s protocol. Human breast cancer (MCF-7) cell line was seeded in a 96-well plate (3 × 104 cells/well) with 100 µL of RPMI 1640 medium and incubated at 37 °C with 5% CO2 of humidified atmosphere overnight. After 24 h, cells were treated with different concentrations of the isolated compound and standard lupeol ranging from 10 to 50 µg/mL for 72 h. For positive control, cells were treated with camptothecin at the concentrations of 0.5–2.0 µg/mL for 72 h. Cells treated with DMSO were used as a control. The cell proliferation was measured at 570 nm using a microplate reader (Molecular Devices, Sunnyvale, CA, USA). The IC50 values (50% growth inhibitory concentration) were calculated from three independent experiments, in which there were three technical replicates (n = 3) per concentration per experiment.
2.10. Statistical Analysis
The IC50 values of the isolated and standard lupeol were expressed as mean ± SD of the three independent experiments (n = 3), which were calculated using a non-linear dose–response curve fitting analysis with GraphPad Prism 7 (GraphPad Software, Inc., San Diego, CA, USA). The level of significance (* p < 0.05, ** p < 0.01 and *** p < 0.001) was determined using one-way ANOVA followed by a post hoc Dunnett test by comparison to the untreated controls.
4. Discussion
The isolation of anti-proliferative compounds from
C. nutans is not easy. This may be due to the complexities of the plant’s nature and also various fractionation/chromatographic steps which need to be optimized. We came up with a strategy to pre-screen (profile) the volatile compounds present in every part of the plant; firstly, using a GC-MS method, we have optimized for the detection of volatile compounds [
7], then only select the suitable plant part and target compound. In our present study, we found that the aerial part of the plant, i.e., the leaf, was predominated by vitamin E (α-tocopherol), squalene, and phytosterols such as stigmasterol, γ/β-sitosterol, and campesterol. Interestingly, the intensities of these compounds decreased when moving to the lower part of the plant, i.e., the stem. In the stem, lupeol was found to start to accumulate abundantly and the amount was much higher than squalene and other phytosterols. Lupeol is one of the pentacyclic triterpenes with a 30-carbon skeleton comprising four six-membered rings and one five-membered ring [
17]. More intriguingly, when it came to the underground part of the plant, i.e., the root, it was found that this part contained enormous lupeol levels and some betulin. All of the detected phytosterols and pentacyclic terpenoids (lupeol and betulin) in this study have been reported to possess anti-proliferative and apoptotic activities by other researchers [
18,
19,
20,
21,
22,
23,
24,
25,
26,
27].
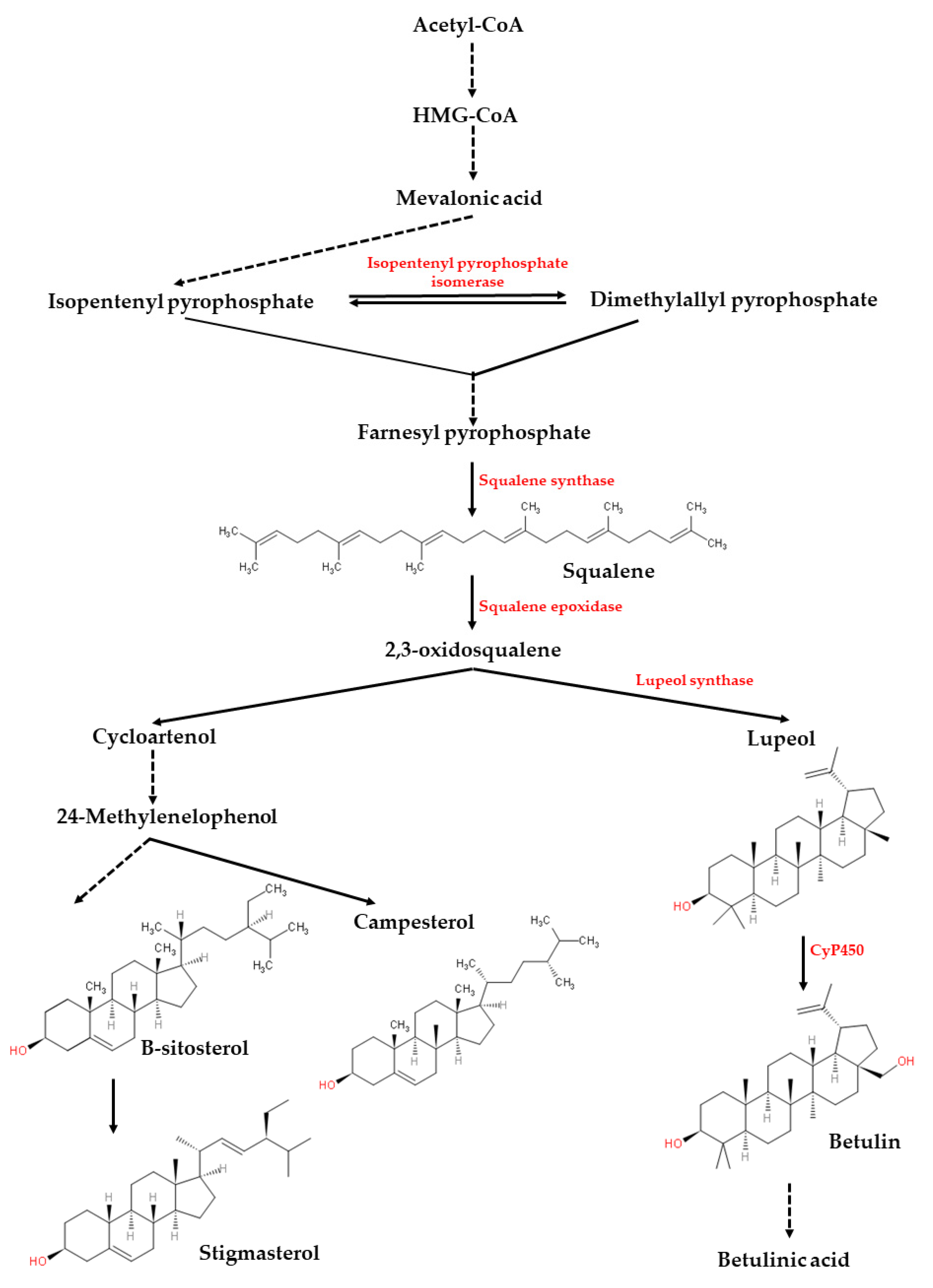
Figure 5 illustrates the biosynthesis of phytosterols and some pentacyclic triterpenoids such as lupeol and betulin. The figure was made by referring to [
28,
29,
30]. The biosynthesis of the compounds begins from the reactions of two precursors called isopentenyl pyrophosphate (IPP) and dimethylallyl pyrophosphate (DMAPP), which are derived from acetyl-CoA via the mevalonic acid (MVA) pathway [
28,
29,
30]. The reaction is catalyzed by the enzyme isopentenyl pyrophosphate isomerase to yield the geranylpyrophosphate (GPP) and farnesyl pyrophosphate (FPP) via the catalysis of prenyltransferase. Then, two molecules of FPPs are catalyzed by the squalene synthase to yield squalene, which is the important precursor for various phytosterols and pentacyclic triterpenoids. Via the squalene epoxidase, squalene is converted to 2,3-oxidosqualene, which is the branch point for the subsequent biosynthesis of phytosterols or pentacyclic triterpenoids [
28,
29,
30]. For phytosterols, multiple enzymatic steps are involved in the plants to convert 2,3-oxidosqualene to cycloartenol and end products such as stigmasterol, sitosterol and campesterol [
30]. For pentacyclic triterpenoids such as lupeol and betulin, the first step is the conversion of 2,3-oxidosqualene to lupeol by lupeol synthase, and then followed by three sequential oxidations catalyzed by cytochrome P450 monooxygenases (CyP450), leading to the generation of betulin, betulinic aldehyde, and betulinic acid [
28,
29]. Based on the results observed in our study, it can be postulated that squalene biosynthesis may actively take part in the aerial part (leaf). This squalene then becomes the precursor for the biosynthesis of lupeol in the stem part. Finally, the root part may either become the synthesis site or main storage site for lupeol.
To date, two studies related to the isolation of lupeol from
C. nutans plant have been reported. One was reported by Dampawan and colleagues [
31] in 1977 in which the plant part used was the stem. Another one was reported by Le and colleagues [
15] in 2017 in which the plant parts used were the leaf and stem. Our study is the first report on the isolation of lupeol from the root part. Owing to its high abundance in the roots and simple root profile obtained via GC-MS analysis, we developed a simpler and straight-forward isolation method for lupeol from the plant’s roots. In our study, we used an initial 80 g of freeze-dried root powder and macerated it with ethyl acetate for five days during the first extraction, followed by three days for the second extraction on the root residues with ethyl acetate, and combined the two extracts and dried them. The vacuum-dried extract was re-dissolved in methanol and underwent recrystallization to yield 1.138 g white amorphous compound, which was identified to be lupeol (yield of extraction = 1.423%). In [
31], an initial two kilograms of dried and milled
C. nutans stems were extracted by using light petroleum in Soxhlet apparatus, followed by silica gel column chromatography using ether and light petroleum for elution, then further purified by using preparative thin layer chromatography and recrystallization to yield 3.7 g of lupeol (yield = 0.185%). In [
15], an initial four kilograms of dried and powdered
C. nutans leaves were macerated with methanol and filtered, followed by solvent–solvent partitioned with hexane to yield a crude extract of 153.9 g. The crude hexane extract was later dissolved in methanol and chromatographed over Diaion HP20SS using ethanol to yield five sub-fractions (H1, H2, H3, H4, and H5). These five sub-fractions were further subjected to column chromatography using different gels to produce four pure compounds (shaftoside, stigmasterol, β-sitosterol, and lupeol). Unfortunately, the final yield for each of the pure compounds was not mentioned in the literature and thus could not compare to our method. However, compared to Dampawan, our method manages to produce a higher extraction yield of lupeol. Additionally, our method is simple and straightforward compared to [
15,
31] as it does not involve solvent–solvent partition, and also without the hassle to go through the column or preparative thin-layer chromatography. The successful isolation of lupeol from
C. nutans roots in this study also suggests the importance of GC-MS- or LC-MS-based profiling of every part of the plant first prior to the selection of potential candidate compounds for the isolation process.
In addition, the extensive characterization of the isolated compound using UV-visible spectrophotometry, Fourier transform infrared spectroscopy, GC-MS, and nuclear magnetic resonance have been able to identify the isolated compound as lupeol. Further analysis of the anti-proliferative activity of the isolated lupeol against human breast cancer cells has revealed that the isolated lupeol was able to demonstrate comparable anti-proliferative activity with the standard lupeol and positive control camptothecin. A study reported by [
22] has postulated the possible mechanism of lupeol on human breast cancer cells by down-regulating the anti-apoptosis proteins, Bcl-2 and Bcl-XL, which lead to cell apoptosis. Our previous study [
7] also shows that the root extract from
C. nutans is capable of causing apoptosis of the breast cancer cells via the condensation of chromatin, down-regulation of the BCL2 gene, and the loss of mitochondrial membrane potential of the cancer cells. We therefore highly speculate that the pro-apoptotic capability of
C. nutans roots could be highly correlated with the lupeol contained inside the roots. We therefore are interested in exploring the molecular mechanisms associated with anti-proliferative activity exerted by the isolated lupeol. Further, the profiling method presented in this study is a simple qualitative study of volatile compounds in different parts of
C. nutans to aid in the isolation of the anti-proliferative compound. Referring to the successful GC-MS-based metabolomics study of
C. nutans leaf extracts on α-glucosidase activity as reported by [
32], we are also interested in applying this similar sophisticated GC-MS-based metabolomics in the future to explore the anti-proliferative mechanism of lupeol isolated from
C. nutans plants harvested from different growing stages or collected from different regions.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




