
Impact of Dietary Potassium Nitrate on the Life Span of Drosophila melanogaster


 ,
, 
Abstract
:1. Introduction
- (a)
- enzymatic pathway—when the special enzyme NO-synthase endogenously converts L-arginine into NO [10];
- (b)
- non-enzymatic recently discovered nitrate (KNO3−)-nitrite (NO2)-nitric oxide (NO) pathway.
2. Materials and Methods
2.1. Fruit Flies Culture
2.2. Feeding Assay
2.3. RING Protocol (Locomotor Behavior)
2.4. Statistical Analysis
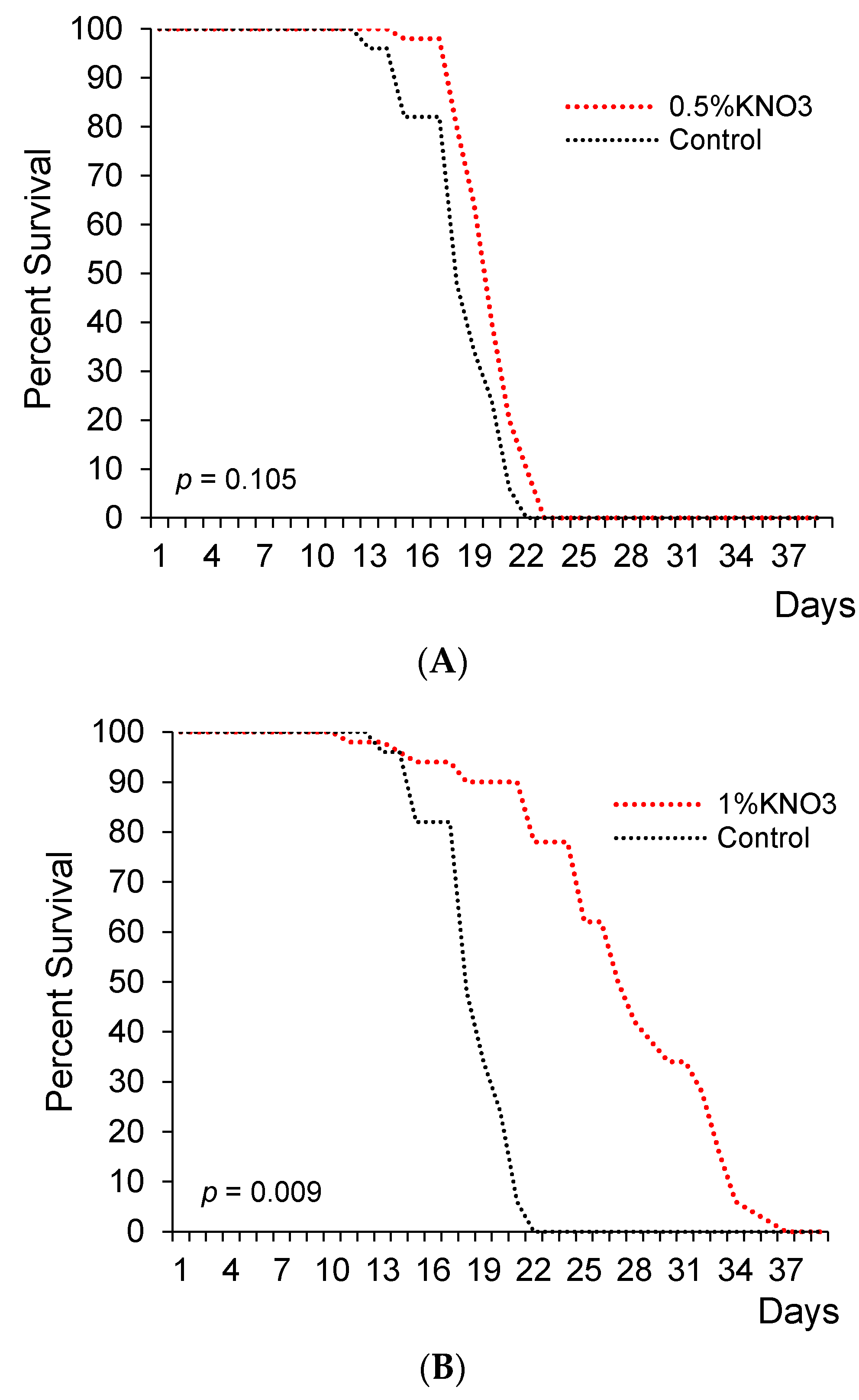
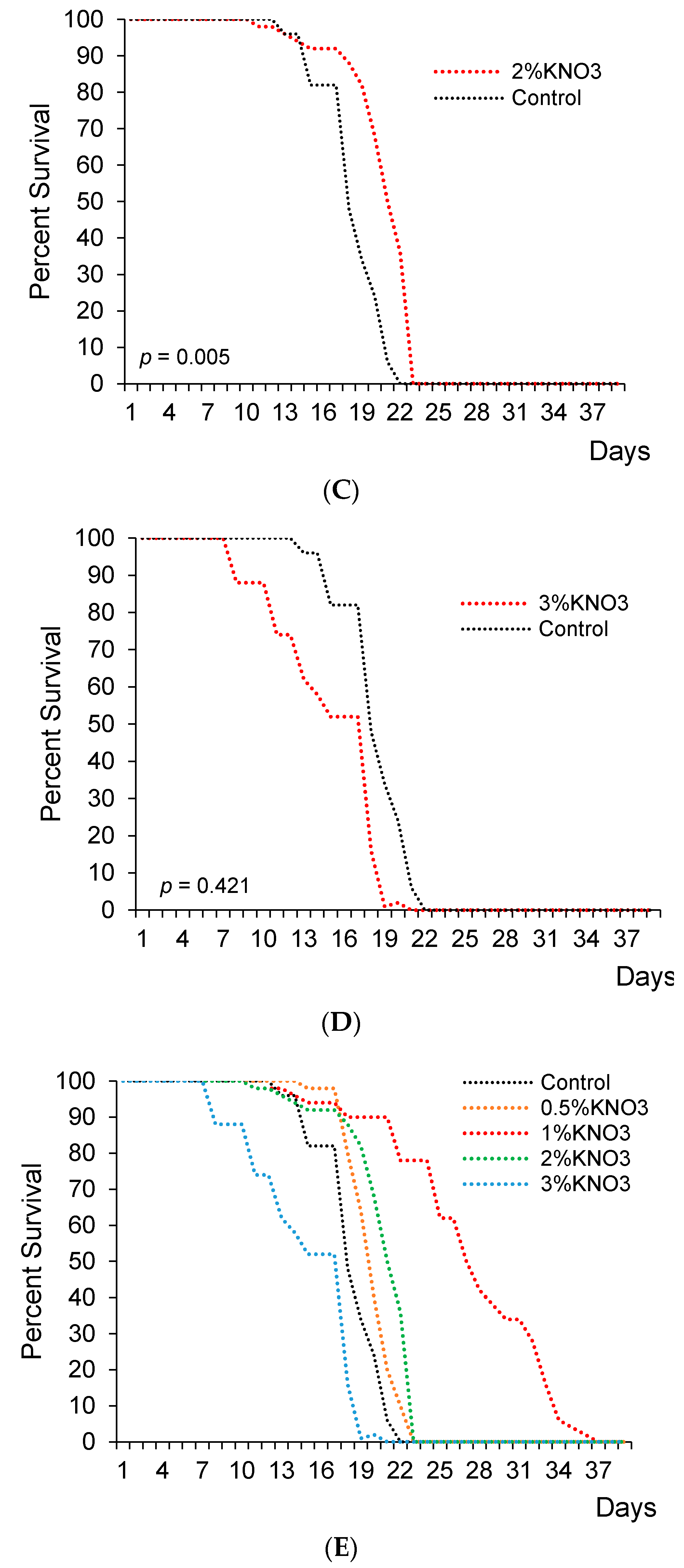
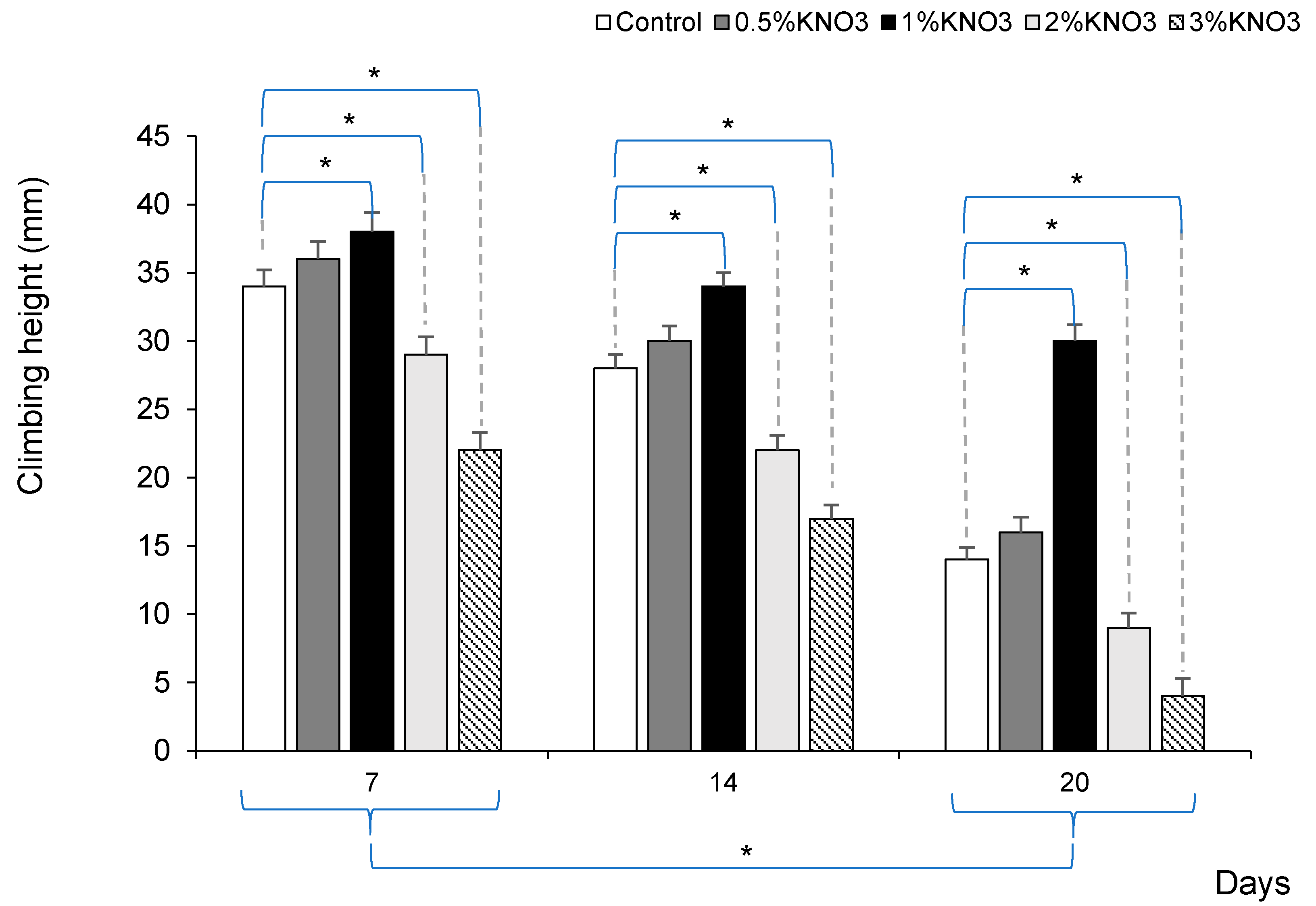
3. Results
4. Discussion & Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaeberlein, M. Longevity and aging. F1000prime Rep. 2013, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Niccoli, T.; Partridge, L. Ageing as a risk factor for disease. Curr. Biol. 2012, 22, R741–R752. [Google Scholar] [CrossRef] [Green Version]
- Lloyd-Sherlock, P.; Kalache, A.; Kirkwood, T.; McKee, M.; Prince, M. WHO’s proposal for a decade of healthy ageing. Lancet 2019, 394, 2152–2153. [Google Scholar] [CrossRef] [Green Version]
- Liubertas, T.; Kairaitis, R.; Stasiule, L.; Capkauskiene, S.; Stasiulis, A.; Viskelis, P.; Viškelis, J.; Urbonaviciene, D. The influence of amaranth (Amaranthus hypochondriacus) dietary nitrates on the aerobic capacity of physically active young persons. J. Int. Soc. Sports Nutr. 2020, 17. [Google Scholar] [CrossRef]
- Simmons, D. Epigenetic influence and disease. Nat. Educ. 2008, 1, 6. [Google Scholar]
- Kaelin, W.G., Jr.; McKnight, S.L. Influence of metabolism on epigenetics and disease. Cell 2013, 153, 56–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Cabo, R.; Carmona-Gutierrez, D.; Bernier, M.; Hall, M.N.; Madeo, F. The search for antiaging interventions: From elixirs to fasting regimens. Cell 2014, 157, 1515–1526. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.M. Dietary nitrate supplementation and exercise performance. Sports Med. (Auckl. N.Z.) 2014, 44 (Suppl. 1), S35–S45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.M.; Vanhatalo, A.; Seals, D.R.; Rossman, M.J.; Piknova, B.; Jonvik, K.L. Dietary Nitrate and Nitric Oxide Metabolism: Mouth, Circulation, Skeletal Muscle, and Exercise Performance. Med. Sci. Sports Exerc. 2021, 53, 280–294. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Weitzberg, E.; Gladwin, M.T. The nitrate–nitrite–nitric oxide pathway in physiology and therapeutics. Nat. Rev. Drug Discov. 2008, 7, 156–167. [Google Scholar] [CrossRef]
- Torregrossa, A.C.; Aranke, M.; Bryan, N.S. Nitric oxide and geriatrics: Implications in diagnostics and treatment of the elderly. J. Geriatr. Cardiol. 2011, 8, 230. [Google Scholar]
- Lundberg, J.O.; Govoni, M. Inorganic nitrate is a possible source for systemic generation of nitric oxide. Free Radic. Biol. Med. 2004, 37, 395–400. [Google Scholar] [CrossRef]
- Duncan, C.; Dougall, H.; Johnston, P.; Green, S.; Brogan, R.; Leifert, C.; Smith, L.; Golden, M.; Benjamin, N. Chemical generation of nitric oxide in the mouth from the enterosalivary circulation of dietary nitrate. Nat. Med. 1995, 1, 546–551. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Gladwin, M.T.; Ahluwalia, A.; Benjamin, N.; Bryan, N.S.; Butler, A.; Cabrales, P.; Fago, A.; Feelisch, M.; Ford, P.C.; et al. Nitrate and nitrite in biology, nutrition and therapeutics. Nat. Chem. Biol. 2009, 5, 865–869. [Google Scholar] [CrossRef]
- Bryan, N.S.; Loscalzo, J. Nitrite and Nitrate in Human Health and Disease; Springer International Publishing: Cham, Switzerland, 2017. [Google Scholar]
- Van Der Loo, B.; Labugger, R.; Skepper, J.N.; Bachschmid, M.; Kilo, J.; Powell, J.M.; Palacios-Callender, M.; Erusalimsky, J.D.; Quaschning, T.; Malinski, T.; et al. Enhanced peroxynitrite formation is associated with vascular aging. J. Exp. Med. 2000, 192, 1731–1744. [Google Scholar] [CrossRef] [Green Version]
- Seals, D.R.; Jablonski, K.L.; Donato, A.J. Aging and vascular endothelial function in humans. Clin. Sci. 2011, 120, 357–375. [Google Scholar] [CrossRef] [Green Version]
- Toda, N.; Ayajiki, K.; Okamura, T. Cerebral blood flow regulation by nitric oxide in neurological disorders. Can. J. Physiol. Pharmacol. 2009, 87, 581–594. [Google Scholar] [CrossRef]
- Moretti, C.; Zhuge, Z.; Zhang, G.; Haworth, S.M.; Paulo, L.L.; Guimarães, D.D.; Cruz, J.; Montenegro, M.F.; Cordero-Herrera, I.; Braga, V.; et al. The obligatory role of host microbiota in bioactivation of dietary nitrate. Free Radic. Biol. Med. 2019, 145, 342–348. [Google Scholar] [CrossRef]
- Carlström, M.; Lundberg, J.O.; Weitzberg, E. Mechanisms underlying blood pressure reduction by dietary inorganic nitrate. Acta Physiol. 2018, 224, e13080. [Google Scholar] [CrossRef]
- Grotewiel, M.S.; Martin, I.; Bhandari, P.; Cook-Wiens, E. Functional senescence in Drosophila melanogaster. Ageing Res. Rev. 2005, 4, 372–397. [Google Scholar] [CrossRef]
- Iliadi, K.G.; Boulianne, G.L. Age-related behavioral changes in Drosophila. Ann. N. Y. Acad. Sci. 2010, 1197, 9–18. [Google Scholar] [CrossRef]
- Davies, S.A.; Stewart, E.J.; Huesmann, G.R.; Skaer, N.J.; Maddrell, S.H.; Tublitz, N.J.; Dow, J.A. Neuropeptide stimulation of the nitric oxide signaling pathway in Drosophila melanogaster Malpighian tubules. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1997, 273, R823–R827. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Kounatidis, I.; Ligoxygakis, P. Drosophila as a model to study the role of blood cells in inflammation, innate immunity and cancer. Front. Cell. Infect. Microbiol. 2014, 3, 113. [Google Scholar] [CrossRef]
- Pandey, U.B.; Nichols, C.D. Human disease models in Drosophila melanogaster and the role of the fly in therapeutic drug discovery. Pharmacol. Rev. 2011, 63, 411–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moretti, C.H.; Schiffer, T.A.; Montenegro, M.F.; Larsen, F.J.; Tsarouhas, V.; Carlström, M.; Lundberg, J.O. Dietary nitrite extends lifespan and prevents age-related locomotor decline in the fruit fly. Free Radic. Biol. Med. 2020, 160, 860–870. [Google Scholar] [CrossRef] [PubMed]
- Peat, J.; Barton, B. Medical Statistics: A Guide to Data Analysis and Critical Appraisal; Blackwell Publishing: Hoboken, NJ, USA, 2005. [Google Scholar]
- McDonald, J.H. Handbook of Biological Statistics, 3rd ed.; Sparky House Publishing: Baltimore, MD, USA, 2014; pp. 157–164. [Google Scholar]
- Richardson, J.T.E. Mann-Whitney Test. In The SAGE Encyclopedia of Educational Research, Measurement, and Evaluation; Frey, B.B., Ed.; SAGE Publications: Thousand Oaks, CA, USA, 2018; pp. 1005–1008. [Google Scholar]
- Nichols, C.D.; Becnel, J.; Pandey, U.B. Methods to assay Drosophila behavior. J. Vis. Exp. 2012, 61, e3795. [Google Scholar]
- Koliada, A.; Gavrilyuk, K.; Burdylyuk, N.; Strilbytska, O.; Storey, K.B.; Kuharskii, V.; Lushchak, O.; Vaiserman, A. Mating status affects Drosophila lifespan, metabolism and antioxidant system. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2020, 246, 110716. [Google Scholar] [CrossRef]
- Ziehm, M.; Thornton, J.M. Unlocking the potential of survival data for model organisms through a new database and online analysis platform: SurvCurv. Aging Cell 2013, 12, 910–916. [Google Scholar] [CrossRef] [Green Version]
- Ziehm, M.; Piper, M.D.; Thornton, J.M. Analysing variation in Drosophila aging across independent experimental studies: A meta-analysis of survival data. Aging Cell 2013, 12, 917–922. [Google Scholar] [CrossRef] [Green Version]
- Chhabra, R.; Kolli, S.; Bauer, J.H. Organically grown food provides health benefits to Drosophila melanogaster. PLoS ONE 2013, 8, e52988. [Google Scholar] [CrossRef] [Green Version]
- Van Dam, E.; Van Leeuwen, L.A.; Dos Santos, E.; James, J.; Best, L.; Lennicke, C.; Vincent, A.J.; Marinos, G.; Foley, A.; Cochemé, H.M.; et al. Sugar-induced obesity and insulin resistance are uncoupled from shortened survival in Drosophila. Cell Metab. 2020, 31, 710–725. [Google Scholar] [CrossRef]
- Duranski, M.R.; Greer, J.J.; Dejam, A.; Jaganmohan, S.; Hogg, N.; Langston, W.; Patel, R.P.; Yet, S.-F.; Wang, X.; Lefer, D.J.; et al. Cytoprotective effects of nitrite during in vivo ischemia-reperfusion of the heart and liver. J. Clin. Investig. 2005, 115, 1232–1240. [Google Scholar] [CrossRef] [Green Version]
- Vitturi, D.A.; Patel, R.P. Current perspectives and challenges in understanding the role of nitrite as an integral player in nitric oxide biology and therapy. Free Radic. Biol. Med. 2011, 51, 805–812. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Meng, Z.; Liu, Y.; Patel, R.P.; Lang, J.D. The hepatoprotective effect of sodium nitrite on cold ischemia-reperfusion injury. J. Transpl. 2012, 2012, 635179. [Google Scholar] [CrossRef]
- Wright, M.J.; Davison, K.L. Nitrate accumulation in crops and nitrate poisoning in animals. In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 1964; Volume 16, pp. 197–247. [Google Scholar]
- Hord, N.G.; Tang, Y.; Bryan, N.S. Food sources of nitrates and nitrites: The physiologic context for potential health benefits. Am. J. Clin. Nutr. 2009, 90, 1–10. [Google Scholar] [CrossRef]
- Piper, M.D.; Partridge, L. Drosophila as a model for ageing. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 2707–2717. [Google Scholar] [CrossRef]
- Partridge, L. The new biology of ageing. Phil. Trans. R. Soc. 2010, 365, 147–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, A.L. Of worms and women: Sarcopenia and its role in disability and mortality. J. Am. Geriatr. Soc. 2004, 52, 1185–1190. [Google Scholar] [CrossRef]
- Augustin, H.; Partridge, L. Invertebrate models of age-related muscle degeneration. Biochim. Biophys. Acta 2009, 1790, 1084–1094. [Google Scholar] [CrossRef] [PubMed]
- Yanase, T.; Yanagita, I.; Muta, K.; Nawata, H. Frailty in elderly diabetes patients. Endocr. J. 2017, 65, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyère, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Zamboni, M.; et al. Sarcopenia: Revised European consensus on definition and diagnosis. Age Ageing 2019, 48, 16–31. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.S.; Lekkas, P.; Braddock, J.M.; Farman, G.P.; Ballif, B.A.; Irving, T.C.; Maughan, D.W.; Vigoreaux, J.O. Aging enhances indirect flight muscle fiber performance yet decreases flight ability in Drosophila. Biophys. J. 2008, 95, 2391–2401. [Google Scholar] [CrossRef] [Green Version]
- Simon, A.F.; Liang, D.T.; Krantz, D.E. Differential decline in behavioral performance of Drosophila melanogaster with age. Mech. Ageing Dev. 2006, 127, 647–651. [Google Scholar] [CrossRef]
- Martinez, V.G.; Javadi, C.S.; Ngo, E.; Ngo, L.; Lagow, R.D.; Zhang, B. Age-related changes in climbing behavior and neural circuit physiology in Drosophila. Dev. Neurobiol. 2007, 67, 778–791. [Google Scholar] [CrossRef] [PubMed]
- Rhodenizer, D.; Martin, I.; Bhandari, P.; Pletcher, S.D.; Grotewiel, M. Genetic and environmental factors impact age-related impairment of negative geotaxis in Drosophila by altering age-dependent climbing speed. Exp. Gerontol. 2008, 43, 739–748. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, M.; Ocorr, K.; Bodmer, R.; Cartry, J. Drosophila as a model to study cardiac aging. Exp. Gerontol. 2011, 46, 326–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, S.L.; De Craen, A.J.; Kerse, N.; Teh, R.; Granic, A.; Davies, K.; Wesnes, K.; Den Elzen, W.P.J.; Gussekloo, J.; Stephan, B.C.; et al. Predicting Risk of Cognitive Decline in Very Old Adults Using Three Models: The Framingham Stroke Risk Profile; the Cardiovascular Risk Factors, Aging, and Dementia Model and Oxi-Inflammatory Biomarkers. J. Am. Geriatr. Soc. 2017, 65, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Sharp, S.I.; Aarsland, D.; Day, S.; Sønnesyn, H.; Alzheimer’s Society Vascular Dementia Systematic Review Group; Ballard, C. Hypertension is a potential risk factor for vascular dementia: Systematic review. Int. J. Geriatr. Psychiatry 2011, 26, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Snyder, H.M.; Corriveau, R.A.; Craft, S.; Faber, J.E.; Greenberg, S.M.; Knopman, D.; Lamb, B.T.; Montine, T.J.; Nedergaard, M.; Carrillo, M.C.; et al. Vascular contributions to cognitive impairment and dementia including Alzheimer’s disease. Alzheimers Dement 2015, 11, 710–717. [Google Scholar] [CrossRef] [Green Version]
- Fasuyi, A.O.; Dairo, F.A.S.; Adeniji, A.O. Tropical vegetable (Amaranthus cruentus) leaf meal as alternative protein supplement in broiler starter diets: Bionutritional evaluation. J. Cent. Eur. Agric. 2008, 9, 23–34. [Google Scholar]
- Johnson, L.C.; DeVan, A.E.; Justice, J.N.; Seals, D.R. Nitrate and Nitrite in Aging and Age-Related Disease. In Nitrite and Nitrate in Human Health and Disease; Humana Press: Cham, Switzerland, 2017; pp. 259–277. [Google Scholar]
- Lundberg, J.O.; Carlström, M.; Larsen, F.J.; Weitzberg, E. Roles of dietary inorganic nitrate in cardiovascular health and disease. Cardiovasc. Res. 2011, 89, 525–532. [Google Scholar] [CrossRef]
- Rocha, B.S.; Gago, B.; Pereira, C.; Barbosa, R.M.; Bartesaghi, S.; Lundberg, J.O.; Radi, R.; Laranjinha, J. Dietary nitrite in nitric oxide biology: A redox interplay with implications for pathophysiology and therapeutics. Curr. Drug Targets 2011, 12, 1351–1363. [Google Scholar] [CrossRef] [PubMed]
- Carlstrom, M.; Montenegro, M.F. Therapeutic value of stimulating the nitrate-nitrite-nitric oxide pathway to attenuate oxidative stress and restore nitric oxide bioavailability in cardiorenal disease. J. Int. Med. 2019, 285, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Stephan, B.C.; Harrison, S.L.; Keage, H.A.; Babateen, A.; Robinson, L.; Siervo, M. Cardiovascular disease, the nitric oxide pathway and risk of cognitive impairment and dementia. Curr. Cardiol. Rep. 2017, 19, 87. [Google Scholar] [CrossRef] [Green Version]
- Keeton, J.T. History of nitrite and nitrate in food. In Nitrite and Nitrate in Human Health and Disease; Humana Press: Totowa, NJ, USA, 2011; pp. 69–84. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Potassium Nitrate Concentration (%) | Survival Days | Median Lifespan Change (% vs. Control) | ||
|---|---|---|---|---|
| Median | Mean | Max * | ||
| Control | 29.5 | 20.00 | 22 | |
| 0.5 | 30 | 21.60 | 23 | +1.7 |
| 1 | 35 | 31.80 | 37 | +18.6 |
| 2 | 31 | 23.00 | 23 | +5.1 |
| 3 | 29 | 19.20 | 21 | −1.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liubertas, T.; Poderys, J.; Vilma, Z.; Capkauskiene, S.; Viskelis, P. Impact of Dietary Potassium Nitrate on the Life Span of Drosophila melanogaster. Processes 2021, 9, 1270. https://doi.org/10.3390/pr9081270
Liubertas T, Poderys J, Vilma Z, Capkauskiene S, Viskelis P. Impact of Dietary Potassium Nitrate on the Life Span of Drosophila melanogaster. Processes. 2021; 9(8):1270. https://doi.org/10.3390/pr9081270
Chicago/Turabian StyleLiubertas, Tomas, Jonas Poderys, Zigmantaite Vilma, Sandrija Capkauskiene, and Pranas Viskelis. 2021. "Impact of Dietary Potassium Nitrate on the Life Span of Drosophila melanogaster" Processes 9, no. 8: 1270. https://doi.org/10.3390/pr9081270
APA StyleLiubertas, T., Poderys, J., Vilma, Z., Capkauskiene, S., & Viskelis, P. (2021). Impact of Dietary Potassium Nitrate on the Life Span of Drosophila melanogaster. Processes, 9(8), 1270. https://doi.org/10.3390/pr9081270