
Bio-Electrochemical System Depollution Capabilities and Monitoring Applications: Models, Applicability, Advanced Bio-Based Concept for Predicting Pollutant Degradation and Microbial Growth Kinetics via Gene Regulation Modelling


 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Microbial Fuel Cells (MFCs) as a Depollution System for Recalcitrant Pollutants and Specific Pollutants
2.1. Copper (Cu)
2.2. Chromium (Cr)
2.3. Lead (Pb) and Zinc (Zn)
2.4. Cadmium (Cd)
2.5. Dichlorodiphenyl-Dichloroethylene (DDE)
3. Pure and Mixed Microbial Cultures in MFC Systems
4. Release of Genome Sequence and Gene Regulatory Mechanisms Related to Substrate(s) Biodegradation in MFCs
5. Comprehensive Mathematical Models to Predict MFC Output
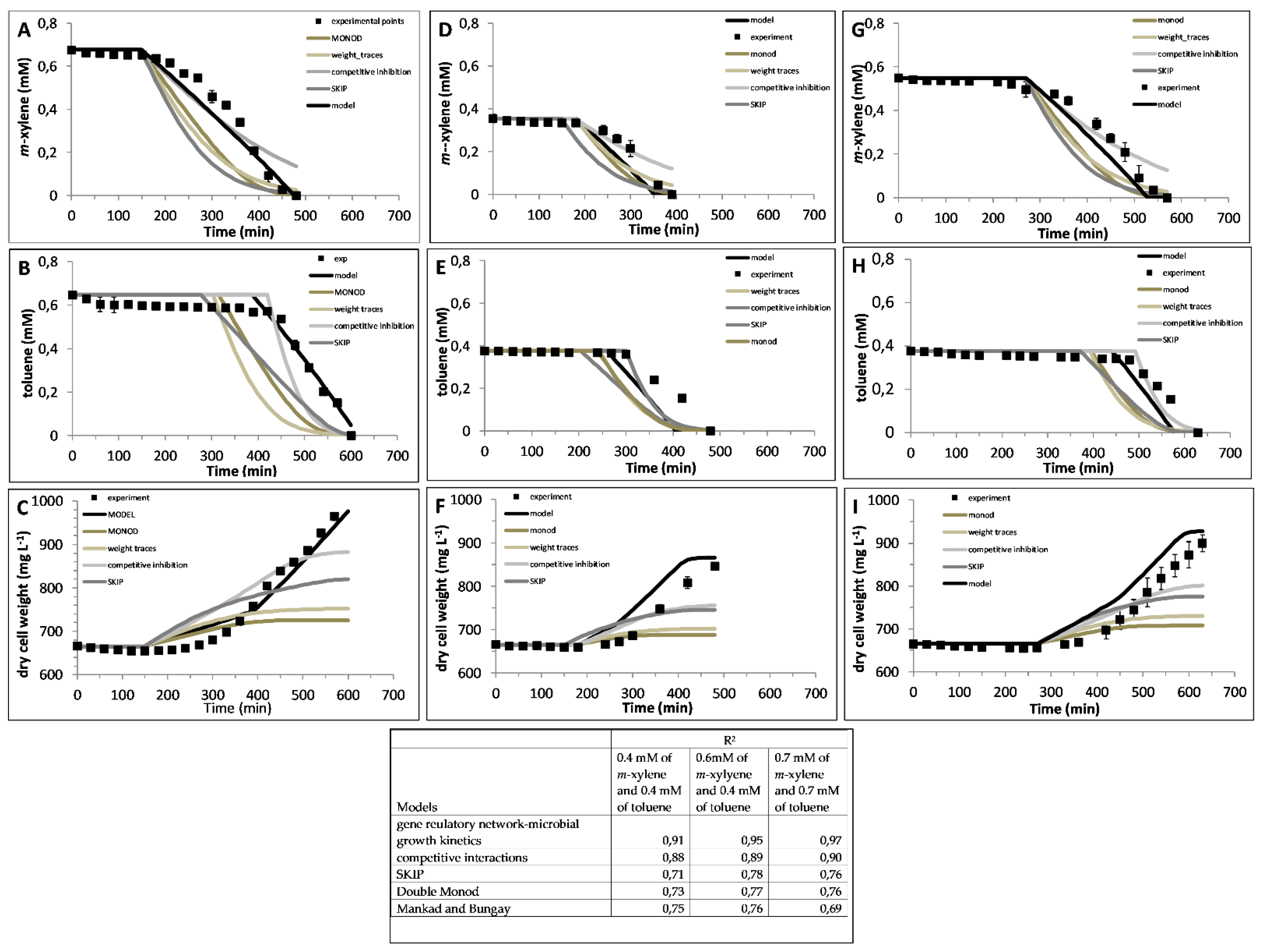
6. Models Used for Microbial Growth Kinetics
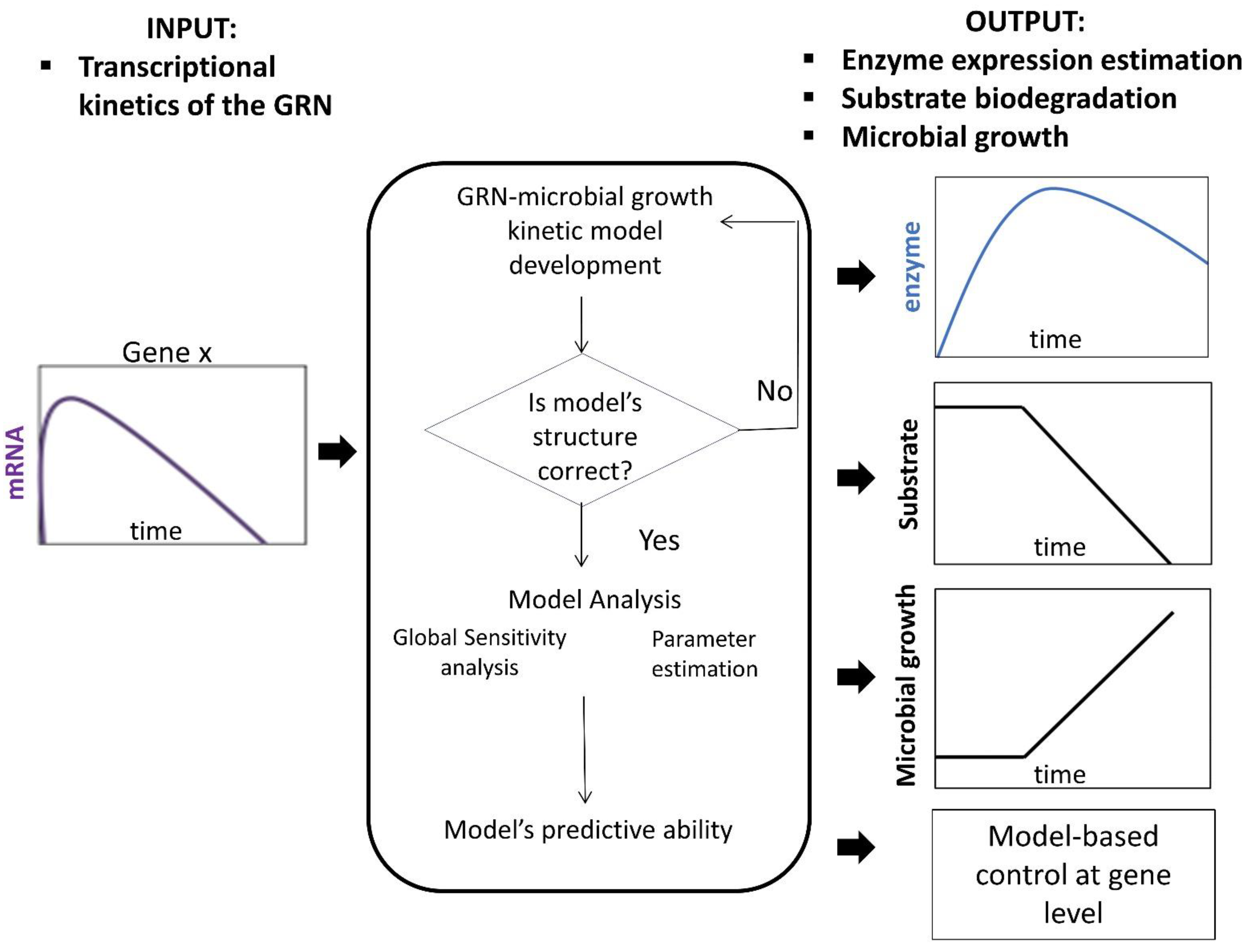
7. Proposed Concept
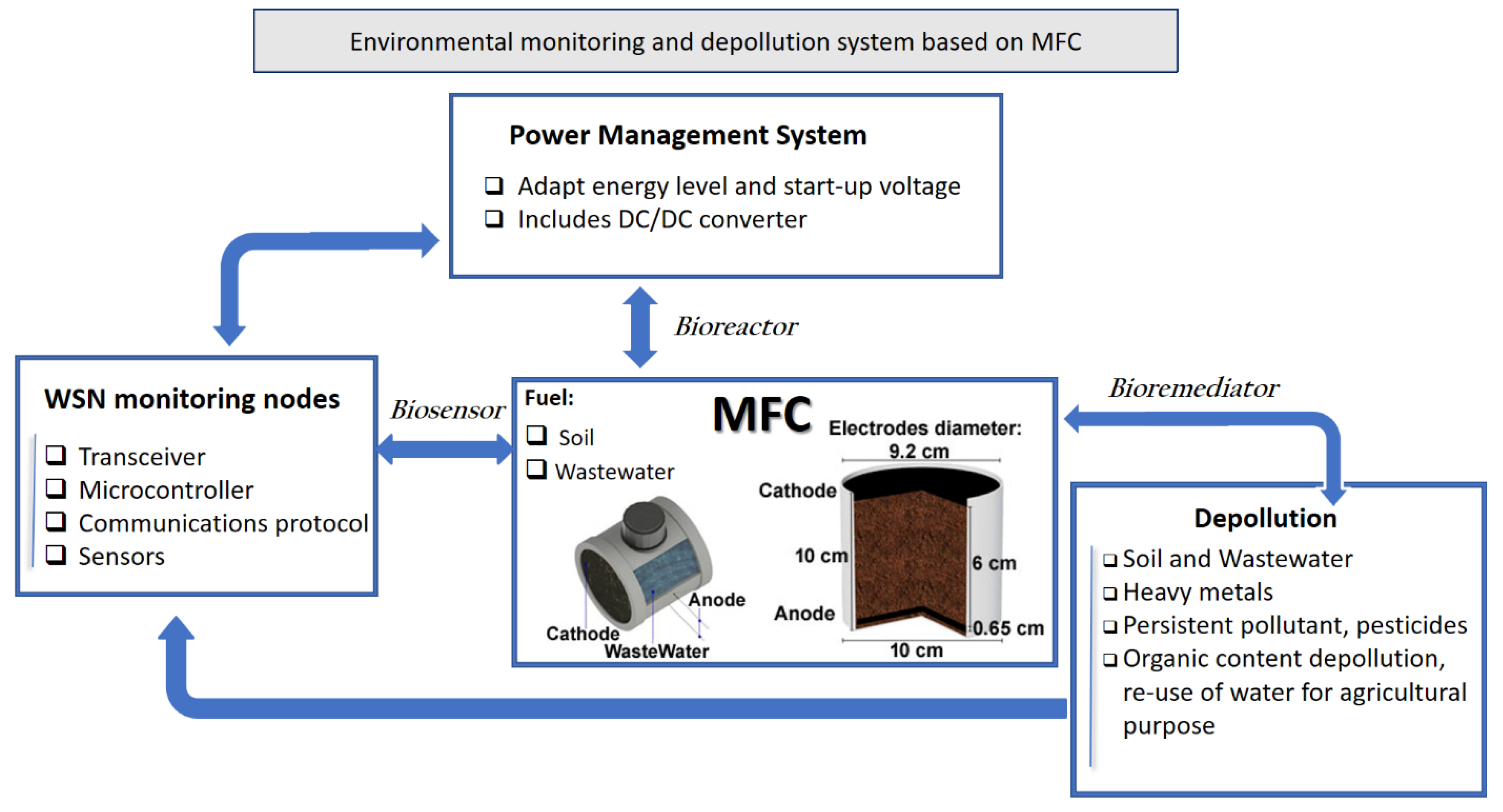
8. MFC Control, Monitoring and Applications
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hassan, H.; Dai, S.; Jin, B. Bioelectrochemical Reaction Kinetics, Mechanisms, and Pathways of Chlorophenol Degradation in MFC Using Different Microbial Consortia. ACS Sustain. Chem. Eng. 2019, 7, 17263–17272. [Google Scholar] [CrossRef]
- Walter, X.A.; Greenman, J.; Ieropoulos, I.A. Microbial fuel cells directly powering a microcomputer. J. Power Sources 2020, 446, 227328. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, D.A.; Carmona-Martínez, A.A.; Chendake, A.D.; Pandit, S.; Pant, D. Modeling and optimization strategies towards performance enhancement of microbial fuel cells. Bioresour. Technol. 2021, 320, 124256. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.E. Biochemical Engineering Fundamentals; McGAraw-Hill: New York, NY, USA, 1986. [Google Scholar]
- Monod, J. The growth of bacterial cultures. Annu. Rev. Microbiol. 1949, 3, 371–394. [Google Scholar] [CrossRef]
- Zhang, J.; Zheng, P.; Zhang, M.; Chen, H.; Chen, T.; Xie, Z.; Cai, J.; Abbas, G. Kinetics of substrate degradation and electricity generation in anodic denitrification microbial fuel cell (AD-MFC). Bioresour. Technol. 2013, 149, 44–50. [Google Scholar] [CrossRef]
- Gadkari, S.; Shemfe, M.; Sadhukhan, J. Microbial fuel cells: A fast converging dynamic model for assessing system performance based on bioanode kinetics. Int. J. Hydrogen Energy 2019, 44, 15377–15386. [Google Scholar] [CrossRef]
- Miroliaei, M.R.; Samimi, A.; Mohebbi-Kalhori, D.; Khorram, M. Kinetics investigation of diversity cultures of E. coli and Shewanella sp., and their combined effect with mediator on MFC performance. J. Ind. Eng. Chem. 2015, 25, 42–50. [Google Scholar] [CrossRef]
- King, R. A structured mathematical model for a class of organisms: I. Development of a model for Streptomyces tendae and application of model-based control. J. Biotechnol. 1997, 52, 219–234. [Google Scholar] [CrossRef]
- Kovarova-Kovar, K.; Egli, T. Growth Kinetics of Suspended Microbial Cells: From Single- Substrate-Controlled Growth to Mixed-Substrate Kinetics. Microbiol. Mol. Biol. Rev. 1998, 62, 646–666. [Google Scholar] [CrossRef]
- Tsipa, A.; Koutinas, M.; Usaku, C.; Mantalaris, A. Optimal bioprocess design through a gene regulatory network—Growth kinetic hybrid model: Towards replacing Monod kinetics. Metab. Eng. 2018, 48, 129–137. [Google Scholar] [CrossRef]
- Gadkari, S.; Gu, S.; Sadhukhan, J. Towards automated design of bioelectrochemical systems: A comprehensive review of mathematical models. Chem. Eng. J. 2018, 343, 303–316. [Google Scholar] [CrossRef]
- Fang, C.; Achal, V. The potential of microbial fuel cells for remediation of heavy metals from soil and water—Review of application. Microorganisms 2019, 7, 697. [Google Scholar] [CrossRef]
- Yang, K.; Ji, M.; Liang, B.; Zhao, Y.; Zhai, S.; Ma, Z.; Yang, Z. Bioelectrochemical degradation of monoaromatic compounds: Current advances and challenges. J. Hazard. Mater. 2020, 398, 122892. [Google Scholar] [CrossRef] [PubMed]
- Kronenberg, M.; Trably, E.; Bernet, N.; Patureau, D. Biodegradation of polycyclic aromatic hydrocarbons: Using microbial bioelectrochemical systems to overcome an impasse. Environ. Pollut. 2017, 231, 509–523. [Google Scholar] [CrossRef]
- Mandal, S.K.; Das, N. Application of microbial fuel cells for bioremediation of environmental pollutants: An overview. J. Microbiol. Biotechnol. Food Sci. 2018, 7, 437–444. [Google Scholar] [CrossRef]
- Wuana, R.A.; Okieimen, F.E. Heavy Metals in Contaminated Soils: A Review of Sources, Chemistry, Risks and Best Available Strategies for Remediation. ISRN Ecol. 2011, 2011, 1–20. [Google Scholar] [CrossRef]
- Saba, B.; Khan, M.; Christy, A.D.; Kjellerup, B.V. Microbial phyto-power systems—A sustainable integration of phytoremediation and microbial fuel cells. Bioelectrochemistry 2019, 127, 1–11. [Google Scholar] [CrossRef]
- Zhang, J.; Cao, X.; Wang, H.; Long, X.; Li, X. Simultaneous enhancement of heavy metal removal and electricity generation in soil microbial fuel cell. Ecotoxicol. Environ. Saf. 2020, 192, 110314. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhou, S.; Xu, Y.; Liu, Z.; Ma, F.; Zhi, L.; Zhou, X. Simultaneous Cr (VI) reduction and bioelectricity generation in a dual chamber microbial fuel cell. Chem. Eng. J. 2018, 334, 1621–1629. [Google Scholar] [CrossRef]
- Zhang, T.; Hu, L.; Zhang, M.; Jiang, M.; Fiedler, H. Cr (VI) removal from soils and groundwater using an integrated adsorption and microbial fuel cell (A-MFC) technology. Environ. Pollut. 2019, 252, 1399–1405. [Google Scholar] [CrossRef]
- Ali, J.; Wang, L.; Waseem, H.; Djellabi, R.; Oladoja, N.A.; Pan, G. FeS@rGO nanocomposites as electrocatalysts for enhanced chromium removal and clean energy generation by microbial fuel cell. Chem. Eng. J. 2020, 384, 123335. [Google Scholar] [CrossRef]
- Habibul, N.; Hu, Y.; Sheng, G. Microbial fuel cell driving electrokinetic remediation of toxic metal contaminated soils. J. Hazard. Mater. 2016, 318, 9–14. [Google Scholar] [CrossRef]
- Song, T.S.; Zhang, J.; Hou, S.; Wang, H.; Zhang, D.; Li, S.; Xie, J. In situ electrokinetic remediation of toxic metal-contaminated soil driven by solid phase microbial fuel cells with a wheat straw addition. J. Chem. Technol. Biotechnol. 2018, 93, 2860–2867. [Google Scholar] [CrossRef]
- Abourached, C.; Catal, T.; Liu, H. Efficacy of single-chamber microbial fuel cells for removal of cadmium and zinc with simultaneous electricity production. Water Res. 2014, 51, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Colantonio, N.; Kim, Y. Cadmium (II) removal mechanisms in microbial electrolysis cells. J. Hazard. Mater. 2016, 311, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.; Hu, N.; Lim, B. Bioresource Technology Cadmium recovery by coupling double microbial fuel cells. Bioresour. Technol. 2014, 170, 361–369. [Google Scholar] [CrossRef]
- Li, M.; Zhou, S.; Xu, Y. Performance of Pb (II) reduction on di ff erent cathodes of microbial electrolysis cell driven by Cr (VI)-reduced microbial fuel cell. J. Power Sources 2019, 418, 1–10. [Google Scholar] [CrossRef]
- Zhang, Y.; He, Q.; Xia, L.; Li, Y.; Song, S. Algae cathode microbial fuel cells for cadmium removal with simultaneous electricity production using nickel foam/graphene electrode. Biochem. Eng. J. 2018, 138, 179–187. [Google Scholar] [CrossRef]
- Ancona, V.; Caracciolo, A.B.; Borello, D.; Ferrara, V.; Grenni, P.; Pietrelli, A. Microbial fuel cell: An energy harvesting technique for environmental remediation. Int. J. Environ. Impacts 2020, 3, 168–179. [Google Scholar] [CrossRef]
- Borello, D.; Gagliardi, G.; Aimola, G.; Ancona, V.; Grenni, P.; Bagnuolo, G.; Garbini, G.L.; Rolando, L.; Barra Caracciolo, A. Use of microbial fuel cells for soil remediation: A preliminary study on DDE. Int. J. Hydrogen Energy 2021, 46, 10131–10142. [Google Scholar] [CrossRef]
- Izallalen, M.; Mahadevan, R.; Burgard, A.; Postier, B.; Didonato, R.; Sun, J.; Schilling, C.H.; Lovley, D.R. Geobacter sulfurreducens strain engineered for increased rates of respiration. Metab. Eng. 2008, 10, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Zhang, H.; Li, Y.; Zhang, H.; Wang, O.; Zhang, J.; Liu, F. Co-occurrence of Methanosarcina mazei and Geobacteraceae in an iron (III)-reducing enrichment culture. Front. Microbiol. 2015, 6, 942. [Google Scholar] [CrossRef] [PubMed]
- Logan, B.E. Extracting Hydrogen and Electricity from Renewable Resources. Environ. Sci. Technol. 2004, 38, 160A–167A. [Google Scholar] [CrossRef] [PubMed]
- Orellana, R.; Leavitt, J.J.; Comolli, L.R.; Csencsits, R.; Janot, N.; Flanagan, K.A.; Gray, A.S.; Leang, C.; Izallalen, M.; Mester, T.; et al. U(VI) reduction by diverse outer surface c-type cytochromes of Geobacter sulfurreducens. Appl. Environ. Microbiol. 2013, 79, 6369–6374. [Google Scholar] [CrossRef]
- Inoue, K.; Leang, C.; Franks, A.E.; Woodard, T.L.; Nevin, K.P.; Lovley, D.R. Specific localization of the c-type cytochrome OmcZ at the anode surface in current-producing biofilms of Geobacter sulfurreducens. Environ. Microbiol. Rep. 2011, 3, 211–217. [Google Scholar] [CrossRef]
- Malvankar, N.S.; Lovley, D.R. Microbial nanowires: A new paradigm for biological electron transfer and bioelectronics. ChemSusChem 2012, 5, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Nevin, K.P.; Richter, H.; Covalla, S.F.; Johnson, J.P.; Woodard, T.L.; Orloff, A.L.; Jia, H.; Zhang, M.; Lovley, D.R. Power output and columbic efficiencies from biofilms of Geobacter sulfurreducens comparable to mixed community microbial fuel cells. Environ. Microbiol. 2008, 10, 2505–2514. [Google Scholar] [CrossRef] [PubMed]
- Ishii, S.; Watanabe, K.; Yabuki, S.; Logan, B.E.; Sekiguchi, Y. Comparison of electrode reduction activities of Geobacter sulfurreducens and an enriched consortium in an air-cathode microbial fuel cell. Appl. Environ. Microbiol. 2008, 74, 7348–7355. [Google Scholar] [CrossRef] [PubMed]
- Reguera, G.; Nevin, K.P.; Nicoll, J.S.; Covalla, S.F.; Woodard, T.L.; Lovley, D.R. Biofilm and nanowire production leads to increased current in Geobacter sulfurreducens fuel cells. Appl. Environ. Microbiol. 2006, 72, 7345–7348. [Google Scholar] [CrossRef]
- Reguera, G.; McCarthy, K.D.; Mehta, T.; Nicoll, J.S.; Tuominen, M.T.; Lovley, D.R. Extracellular electron transfer via microbial nanowires. Nature 2005, 435, 1098–1101. [Google Scholar] [CrossRef]
- Richter, H.; McCarthy, K.; Nevin, K.P.; Johnson, J.P.; Rotello, V.M.; Lovley, D.R. Electricity generation by Geobacter sulfurreducens attached to gold electrodes. Langmuir 2008, 24, 4376–4379. [Google Scholar] [CrossRef]
- Smith, J.A.; Lovley, D.R.; Tremblay, P.L. Outer cell surface components essential for Fe (III) oxide reduction by Geobacter metallireducens. Appl. Environ. Microbiol. 2013, 79, 901–907. [Google Scholar] [CrossRef]
- Min, B.; Cheng, S.; Logan, B.E. Electricity generation using membrane and salt bridge microbial fuel cells. Water Res. 2005, 39, 1675–1686. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Wang, A.; Cheng, S.; Yates, M.; Logan, B.E. Geobacter anodireducens sp. nov., an exoelectrogenic microbe in bioelectrochemical systems. Int. J. Syst. Evol. Microbiol. 2014, 64, 3485–3491. [Google Scholar] [CrossRef] [PubMed]
- Summers, Z.M.; Fogarty, H.E.; Leang, C.; Franks, A.E.; Malvankar, N.S.; Lovley, D.R. Direct exchange of electrons within aggregates of an evolved syntrophic coculture of anaerobic bacteria. Science 2010, 330, 1413–1415. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.A.; Nevin, K.P.; Lovley, D.R. Syntrophic growth via quinone-mediated interspecies electron transfer. Front. Microbiol. 2015, 6, 121. [Google Scholar] [CrossRef]
- Shrestha, P.M.; Rotaru, A.E.; Aklujkar, M.; Liu, F.; Shrestha, M.; Summers, Z.M.; Malvankar, N.; Flores, D.C.; Lovley, D.R. Syntrophic growth with direct interspecies electron transfer as the primary mechanism for energy exchange. Environ. Microbiol. Rep. 2013, 5, 904–910. [Google Scholar] [CrossRef]
- Rotaru, A.E.; Shrestha, P.M.; Liu, F.; Shrestha, M.; Shrestha, D.; Embree, M.; Zengler, K.; Wardman, C.; Nevin, K.P.; Lovley, D.R. A new model for electron flow during anaerobic digestion: Direct interspecies electron transfer to Methanosaeta for the reduction of carbon dioxide to methane. Energy Environ. Sci. 2014, 7, 408–415. [Google Scholar] [CrossRef]
- Liu, F.; Rotaru, A.E.; Shrestha, P.M.; Malvankar, N.S.; Nevin, K.P.; Lovley, D.R. Promoting direct interspecies electron transfer with activated carbon. Energy Environ. Sci. 2012, 5, 8982–8989. [Google Scholar] [CrossRef]
- Kotloski, N.J.; Gralnick, J.A. Flavin electron shuttles dominate extracellular electron transfer by Shewanella oneidensis. MBio 2013, 4, e00553-12. [Google Scholar] [CrossRef] [PubMed]
- Gorby, Y.A.; Yanina, S.; McLean, J.S.; Rosso, K.M.; Moyles, D.; Dohnalkova, A.; Beveridge, T.J.; Chang, I.S.; Kim, B.H.; Kim, K.S.; et al. Electrically conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and other exoelectrogens. Proc. Natl. Acad. Sci. USA 2006, 103, 11358–11363. [Google Scholar] [CrossRef]
- Baron, D.; LaBelle, E.; Coursolle, D.; Gralnick, J.A.; Bond, D.R. Electrochemical measurement of electron transfer kinetics by Shewanella oneidensis MR-1. J. Biol. Chem. 2009, 284, 28865–28873. [Google Scholar] [CrossRef]
- Von Canstein, H.; Ogawa, J.; Shimizu, S.; Lloyd, J.R. Secretion of flavins by Shewanella species and their role in extracellular electron transfer. Appl. Environ. Microbiol. 2008, 74, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Ringeisen, B.R.; Henderson, E.; Wu, P.K.; Pietron, J.; Ray, R.; Little, B.; Biffinger, J.C.; Jones-Meehan, J.M. High power density from a miniature microbial fuel cell using Shewanella oneidensis DSP10. Environ. Sci. Technol. 2006, 40, 2629–2634. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Zhang, X.; Pastorella, G.; Connolly, J.O.; Barry, N.; Woolley, R.; Krishnamurthy, S.; Marsili, E. Electron transfer mechanism in Shewanella loihica PV-4 biofilms formed at graphite electrode. Bioelectrochemistry 2012, 87, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Marsili, E.; Baron, D.B.; Shikhare, I.D.; Coursolle, D.; Gralnick, J.A.; Bond, D.R. Shewanella secretes flavins that mediate extracellular electron transfer. Proc. Natl. Acad. Sci. USA 2008, 105, 3968–3973. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Yang, F.; Liu, X.; Bai, L. Influence of substrate on electricity generation of Shewanella loihica PV-4 in microbial fuel cells. Microb. Cell Fact. 2014, 13, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Pandit, S.; Khilari, S.; Roy, S.; Pradhan, D.; Das, D. Improvement of power generation using Shewanella putrefaciens mediated bioanode in a single chambered microbial fuel cell: Effect of different anodic operating conditions. Bioresour. Technol. 2014, 166, 451–457. [Google Scholar] [CrossRef]
- Pham, C.A.; Jung, S.J.; Phung, N.T.; Lee, J.; Chang, I.S.; Kim, B.H.; Yi, H.; Chun, J. A novel electrochemically active and Fe (III)-reducing bacterium phylogenetically related to Aeromonas hydrophila, isolated from a microbial fuel cell. FEMS Microbiol. Lett. 2003, 223, 129–134. [Google Scholar] [CrossRef]
- Dietrich, L.E.P.; Price-Whelan, A.; Petersen, A.; Whiteley, M.; Newman, D.K. The phenazine pyocyanin is a terminal signalling factor in the quorum sensing network of Pseudomonas aeruginosa. Mol. Microbiol. 2006, 61, 1308–1321. [Google Scholar] [CrossRef]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as complex differentiated communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef]
- Rabaey, K.; Boon, N.; Höfte, M.; Verstraete, W. Microbial phenazine production enhances electron transfer in biofuel cells. Environ. Sci. Technol. 2005, 39, 3401–3408. [Google Scholar] [CrossRef]
- Fazli, M.; Almblad, H.; Rybtke, M.L.; Givskov, M.; Eberl, L.; Tolker-Nielsen, T. Regulation of biofilm formation in Pseudomonas and Burkholderia species. Environ. Microbiol. 2014, 16, 1961–1981. [Google Scholar] [CrossRef]
- Park, H.S.; Kim, B.H.; Kim, H.S.; Kim, H.J.; Kim, G.T.; Kim, M.; Chang, I.S.; Park, Y.K.; Chang, H.I. A novel electrochemically active and Fe (III)-reducing bacterium phylogenetically related to Clostridium butyricum isolated from a microbial fuel cell. Anaerobe 2001, 7, 297–306. [Google Scholar] [CrossRef]
- Singh, L.; Wahid, Z. Enhancement of hydrogen production from palm oil mill effluent via cell immobilisation technique. Int. J. Energy Res. 2014, 39, 215–222. [Google Scholar] [CrossRef]
- Wang, H.; Lu, L.; Cui, F.; Liu, D.; Zhao, Z.; Xu, Y. Simultaneous bioelectrochemical degradation of algae sludge and energy recovery in microbial fuel cells. RSC Adv. 2012, 7228–7234. [Google Scholar] [CrossRef]
- Cui, Y.; Rashid, N.; Hu, N.; Saif, M.; Rehman, U.; Han, J. Electricity generation and microalgae cultivation in microbial fuel cell using microalgae-enriched anode and bio-cathode. Energy Convers. Manag. 2014, 79, 674–680. [Google Scholar] [CrossRef]
- Kim, G.T.; Hyun, M.S.; Chang, I.S.; Kim, H.J.; Park, H.S.; Kim, B.H.; Kim, S.D.; Wimpenny, J.W.T.; Weightman, A.J. Dissimilatory Fe (III) reduction by an electrochemically active lactic acid bacterium phylogenetically related to Enterococcus gallinarum isolated from submerged soil. J. Appl. Microbiol. 2005, 99, 978–987. [Google Scholar] [CrossRef]
- Keogh, D.; Lam, L.N.; Doyle, L.E.; Matysik, A.; Pavagadhi, S.; Umashankar, S.; Low, P.M.; Dale, J.L.; Song, Y.; Ng, S.P.; et al. Extracellular Electron Transfer Powers Enterococcus faecalis Biofilm Metabolism. MBio 2018, 9, e00626-17. [Google Scholar] [CrossRef]
- El-Naggar, M.Y.; Wanger, G.; Leung, K.M.; Yuzvinsky, T.D.; Southam, G.; Yang, J.; Lau, W.M.; Nealson, K.H.; Gorby, Y.A. Electrical transport along bacterial nanowires from Shewanella oneidensis MR-1. Proc. Natl. Acad. Sci. USA 2010, 107, 18127–18131. [Google Scholar] [CrossRef]
- Zuo, Y.; Xing, D.; Regan, J.M.; Logan, B.E. Isolation of the exoelectrogenic bacterium Ochrobactrum anthropi YZ-1 by using a U-tube microbial fuel cell. Appl. Environ. Microbiol. 2008, 74, 3130–3137. [Google Scholar] [CrossRef] [PubMed]
- Parameswaran, P.; Bry, T.; Popat, S.C.; Lusk, B.G.; Rittmann, B.E.; Torres, C.I. Kinetic, electrochemical, and microscopic characterization of the thermophilic, anode-respiring bacterium Thermincola ferriacetica. Environ. Sci. Technol. 2013, 47, 4934–4940. [Google Scholar] [CrossRef]
- Wrighton, K.C.; Thrash, J.C.; Melnyk, R.A.; Bigi, J.P.; Byrne-Bailey, K.G.; Remis, J.P.; Schichnes, D.; Auer, M.; Chang, C.J.; Coates, J.D. Evidence for direct electron transfer by a gram-positive bacterium isolated from a microbial fuel cell. Appl. Environ. Microbiol. 2011, 77, 7633–7639. [Google Scholar] [CrossRef]
- Bond, D.R.; Lovley, D.R. Evidence for involvement of an electron shuttle in electricity generation by Geothrix fermentans. Appl. Environ. Microbiol. 2005, 71, 2186–2189. [Google Scholar] [CrossRef] [PubMed]
- Mehta-Kolte, M.G.; Bond, D.R. Geothrix fermentans secretes two different redox-active compounds to utilize electron acceptors across a wide range of redox potentials. Appl. Environ. Microbiol. 2012, 78, 6987–6995. [Google Scholar] [CrossRef]
- Keller, K.L.; Rapp-Giles, B.J.; Semkiw, E.S.; Porat, I.; Brown, S.D.; Wall, J.D. New model for electron flow for sulfate reduction in Desulfovibrio alaskensis G20. Appl. Environ. Microbiol. 2014, 80, 855–868. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.S.; Eaktasang, N.; Kwon, D.Y.; Kim, H.S. Enhanced current production by Desulfovibrio desulfuricans biofilm in a mediator-less microbial fuel cell. Bioresour. Technol. 2014, 165, 27–30. [Google Scholar] [CrossRef]
- Freguia, S.; Masuda, M.; Tsujimura, S.; Kano, K. Lactococcus lactis catalyses electricity generation at microbial fuel cell anodes via excretion of a soluble quinone. Bioelectrochemistry 2009, 76, 14–18. [Google Scholar] [CrossRef]
- Kumar, R.; Singh, L.; Wahid, Z.A.; Din, M.F. Exoelectrogens in microbial fuel cells toward bioelectricity generation: A review. Int. J. Energy Res. 2015, 39, 1048–1067. [Google Scholar] [CrossRef]
- Qiao, Y.; Li, C.M.; Bao, S.J.; Lu, Z.; Hong, Y. Direct electrochemistry and electrocatalytic mechanism of evolved Escherichia coli cells in microbial fuel cells. Chem. Commun. 2008, 1290–1292. [Google Scholar] [CrossRef]
- Fuller, S.J.; McMillan, D.G.G.; Renz, M.B.; Schmidt, M.; Burke, I.T.; Stewart, D.I. Extracellular electron transport-mediated Fe (III) reduction by a community of alkaliphilic bacteria that use flavins as electron shuttles. Appl. Environ. Microbiol. 2014, 80, 128–137. [Google Scholar] [CrossRef]
- Lifang, D.; FrangBai, L.; Shungui, Z.; Yin, H.D.; Jinren, N.I. A study of electron-shuttle mechanism in Klebsiella pneumoniae based microbial fuel cells. Environ. Sci. Technol. 2010, 55, 99–104. [Google Scholar]
- Xing, D.; Zuo, Y.; Cheng, S.; Regan, J.M.; Logan, B.E. Electricity generation by Rhodopseudomonas palustris DX-1. Environ. Sci. Technol. 2008, 42, 4146–4151. [Google Scholar] [CrossRef] [PubMed]
- Inglesby, A.E.; Beatty, D.A.; Fisher, A.C. Rhodopseudomonas palustris purple bacteria fed Arthrospira maxima cyanobacteria: Demonstration of application in microbial fuel cells. RSC Adv. 2012, 2, 4829–4838. [Google Scholar] [CrossRef]
- Raghavulu, S.V.; Goud, R.K.; Sarma, P.N.; Mohan, S.V. Saccharomyces cerevisiae as anodic biocatalyst for power generation in biofuel cell: Influence of redox condition and substrate load. Bioresour. Technol. 2011, 102, 2751–2757. [Google Scholar] [CrossRef] [PubMed]
- Prasad, D.; Arun, S.; Murugesan, M.; Padmanaban, S.; Satyanarayanan, R.S.; Berchmans, S.; Yegnaraman, V. Direct electron transfer with yeast cells and construction of a mediatorless microbial fuel cell. Biosens. Bioelectron. 2007, 22, 2604–2610. [Google Scholar] [CrossRef]
- Hubenova, Y.; Mitov, M. Potential application of Candida melibiosica in biofuel cells. Bioelectrochemistry 2010, 78, 57–61. [Google Scholar] [CrossRef]
- Nandy, A.; Kumar, V.; Kundu, P.P. Utilization of proteinaceous materials for power generation in a mediatorless microbial fuel cell by a new electrogenic bacteria Lysinibacillus sphaericus VA5. Enzym. Microb. Technol. 2013, 53, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Liu, H. New exoelectrogen Citrobacter sp. SX-1 isolated from a microbial fuel cell. J. Appl. Microbiol. 2011, 111, 1108–1115. [Google Scholar] [CrossRef]
- Kimura, Z.I.; Chung, K.M.; Itoh, H.; Hiraishi, A.; Okabe, S. Raoultella electrica sp. nov., isolated from anodic biofilms of a glucose-fed microbial fuel cell. Int. J. Syst. Evol. Microbiol. 2014, 64, 1384–1388. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhong, G.Z.; Qiao, Y.; Huang, J.; Hu, W.H.; Wang, X.G.; Li, C.M. A high performance xylose microbial fuel cell enabled by Ochrobactrum sp. 575 cells. RSC Adv. 2014, 4, 39839–39843. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Khawdas, W.; Aso, Y.; Ohara, H. Microbial fuel cells using Cellulomonas spp. with cellulose as fuel. J. Biosci. Bioeng. 2017, 123, 358–363. [Google Scholar] [CrossRef]
- Khawdas, W.; Watanabe, K.; Karatani, H.; Aso, Y.; Tanaka, T.; Ohara, H. Direct electron transfer of Cellulomonas fimi and microbial fuel cells fueled by cellulose. J. Biosci. Bioeng. 2019, 128, 593–598. [Google Scholar] [CrossRef]
- Aslan, S.; Conghaile, P.Ó.; Leech, D.; Gorton, L.; Timur, S.; Anik, U. Development of an Osmium Redox Polymer Mediated Bioanode and Examination of its Performance in Gluconobacter oxydans Based Microbial Fuel Cell. Electroanalysis 2017, 29, 1651–1657. [Google Scholar] [CrossRef]
- Aslan, S.; Conghaile, P.Ó.; Leech, D.; Gorton, L.; Timur, S.; Anik, U. Development of a Bioanode for Microbial Fuel Cells Based on the Combination of a MWCNT-Au-Pt Hybrid Nanomaterial, an Osmium Redox Polymer and Gluconobacter oxydans DSM 2343 Cells. ChemistrySelect 2017, 2, 12034–12040. [Google Scholar] [CrossRef]
- Adachi, T.; Kitazumi, Y.; Shirai, O.; Kano, K. Direct electron transfer-type bioelectrocatalysis by membrane-bound aldehyde dehydrogenase from Gluconobacter oxydans and cyanide effects on its bioelectrocatalytic properties. Electrochem. Commun. 2021, 123, 106911. [Google Scholar] [CrossRef]
- Liu, X.; Ali, A.; Liu, C.; Liu, Y.; Zhang, P. The first in-depth exploration of the genome of the engineered bacterium, Gluconobacter thailandicus. Biotechnol. Appl. Biochem. 2021, 1–9. [Google Scholar] [CrossRef]
- Park, D.L.M.; Guettler, M.V.; Jain, M.K.; Zeikus, J.G. Microbial utilization of electrically reduced neutral red as the sole electron donor for growth and metabolite production. Appl. Environ. Microbiol. 1999, 65, 2912–2917. [Google Scholar] [CrossRef]
- Choudhury, P.; Uday, U.S.P.; Mahata, N.; Nath Tiwari, O.; Narayan Ray, R.; Kanti Bandyopadhyay, T.; Bhunia, B. Performance improvement of microbial fuel cells for waste water treatment along with value addition: A review on past achievements and recent perspectives. Renew. Sustain. Energy Rev. 2017, 79, 372–389. [Google Scholar] [CrossRef]
- Kouzuma, A.; Kasai, T.; Hirose, A.; Watanabe, K. Catabolic and regulatory systems in shewanella oneidensis MR-1 involved in electricity generation in microbial fuel cells. Front. Microbiol. 2015, 6, 609. [Google Scholar] [CrossRef] [PubMed]
- Aklujkar, M.; Krushkal, J.; Dibartolo, G.; Lapidus, A.; Land, M.L.; Lovley, D.R. The genome sequence of Geobacter metallireducens: Features of metabolism, physiology and regulation common and dissimilar to Geobacter sulfurreducens. BMC Microbiol. 2009, 9, 1–22. [Google Scholar] [CrossRef]
- Heidelberg, J.F.; Paulsen, I.T.; Nelson, K.E.; Gaidos, E.J.; Nelson, W.C.; Read, T.D.; Eisen, J.A.; Seshadri, R.; Ward, N.; Methe, B.; et al. Genome sequence of the dissimilatory metal ion-reducing bacterium Shewanella oneidensis. Nat. Biotechnol. 2002, 20, 1118–1123. [Google Scholar] [CrossRef] [PubMed]
- Serres, M.H.; Riley, M. Genomic analysis of carbon source metabolism of Shewanella oneidensis MR-1: Predictions versus experiments. J. Bacteriol. 2006, 188, 4601–4609. [Google Scholar] [CrossRef] [PubMed]
- Deb, D.; Patel, R.; Balas, V.E. A Review of Control-Oriented Bioelectrochemical Mathematical Models of Microbial Fuel Cells. Processes 2020, 8, 583. [Google Scholar] [CrossRef]
- Jadhav, G.S.; Ghangrekar, M.M. Performance of microbial fuel cell subjected to variation in pH, temperature, external load and substrate concentration. Bioresour. Technol. 2009, 100, 717–723. [Google Scholar] [CrossRef]
- Merkey, B.V.; Chopp, D.L. The Performance of a Microbial Fuel Cell Depends Strongly on Anode Geometry: A Multidimensional Modeling Study. Bull. Math. Biol. 2012, 74, 834–857. [Google Scholar] [CrossRef]
- Ortiz-Martínez, V.M.; Salar-García, M.J.; de los Ríos, A.P.; Hernández-Fernández, F.J.; Egea, J.A.; Lozano, L.J. Developments in microbial fuel cell modeling. Chem. Eng. J. 2015, 271, 50–60. [Google Scholar] [CrossRef]
- Zhang, X.; Halme, A. Modelling of a Microbial Fuel Cell Process. Biotechnol. Lett. 1995, 17, 809–814. [Google Scholar] [CrossRef]
- Picioreanu, C.; van Loosdrecht, M.C.M.; Curtis, T.P.; Scott, K. Model based evaluation of the effect of pH and electrode geometry on microbial fuel cell performance. Bioelectrochemistry 2010, 78, 8–24. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Choo, Y.F.; Kim, B.H.; Wu, P. Modelling and simulation of two-chamber microbial fuel cell. J. Power Sources 2010, 195, 79–89. [Google Scholar] [CrossRef]
- Pinto, R.P.; Srinivasan, B.; Manuel, M.F.; Tartakovsky, B. A two-population bio-electrochemical model of a microbial fuel cell. Bioresour. Technol. 2010, 101, 5256–5265. [Google Scholar] [CrossRef]
- Oliveira, V.B.; Simões, M.; Melo, L.F.; Pinto, A.M.F.R. A 1D mathematical model for a microbial fuel cell. Energy 2013, 61, 463–471. [Google Scholar] [CrossRef]
- Jayasinghe, N.; Franks, A.; Nevin, K.P.; Mahadevan, R. Metabolic modeling of spatial heterogeneity of biofilms in microbial fuel cells reveals substrate limitations in electrical current generation. Biotechnol. J. 2014, 9, 1350–1361. [Google Scholar] [CrossRef]
- Recio-Garrido, D.; Perrier, M.; Tartakovsky, B. Parameter estimation of a microbial fuel cell process control-oriented model. In Proceedings of the 22nd Mediterranean Conference on Control and Automation, Palermo, Italy, 16–19 June 2014; pp. 918–923. [Google Scholar]
- Recio-Garrido, D.; Perrier, M.; Tartakovsky, B. Modeling, optimization and control of bioelectrochemical systems. Chem. Eng. J. 2016, 289, 180–190. [Google Scholar] [CrossRef]
- Esfandyari, M.; Fanaei, M.A.; Gheshlaghi, R.; Akhavan Mahdavi, M. Mathematical modeling of two-chamber batch microbial fuel cell with pure culture of Shewanella. Chem. Eng. Res. Des. 2017, 117, 34–42. [Google Scholar] [CrossRef]
- Esfandyari, M.; Fanaei, M.A.; Gheshlaghi, R.; Akhavan Mahdavi, M. Dynamic modeling of a continuous two-chamber microbial fuel cell with pure culture of Shewanella. Int. J. Hydrogen Energy 2017, 42, 21198–21202. [Google Scholar] [CrossRef]
- Karamzadeh, M.; Kadivarian, H.; Kadivarian, M.; Kazemi, A. Modeling the influence of substrate concentration, anode electrode surface area and external resistance in a start-up on the performance of microbial fuel cell. Bioresour. Technol. Rep. 2020, 12, 100559. [Google Scholar] [CrossRef]
- Pinto, R.P.; Srinivasan, B.; Escapa, A.; Tartakovsky, B. Multi-population model of a microbial electrolysis cell. Environ. Sci. Technol. 2011, 45, 5039–5046. [Google Scholar] [CrossRef] [PubMed]
- Kato Marcus, A.; Torres, C.I.; Rittmann, B.E. Conduction-Based Modeling of the Biofilm Anode of a Microbial Fuel Cell. Biotechnol. Bioeng. 2007, 98, 1171–1182. [Google Scholar] [CrossRef] [PubMed]
- Sirinutsomboon, B. Modeling of a membraneless single-chamber microbial fuel cell with molasses as an energy source. Int. J. Energy Environ. Eng. 2014, 5, 1–9. [Google Scholar] [CrossRef]
- Picioreanu, C.; Head, I.M.; Katuri, K.P.; van Loosdrecht, M.C.M.; Scott, K. A computational model for biofilm-based microbial fuel cells. Water Res. 2007, 41, 2921–2940. [Google Scholar] [CrossRef]
- Picioreanu, C.; Katuri, K.P.; Head, I.M.; Van Loosdrecht, M.C.M.; Scott, K. Mathematical model for microbial fuel cells with anodic biofilms and anaerobic digestion. Water Sci. Technol. 2008, 57, 965–971. [Google Scholar] [CrossRef]
- Jafary, T.; Ghoreyshi, A.A.; Najafpour, G.D.; Fatemi, S.; Rahimnejad, M. Investigation on performance of microbial fuel cells based on carbon sources and kinetic models. Int. J. Energy Res. 2013, 37, 1539–1549. [Google Scholar] [CrossRef]
- Tsipa, A.; Koutinas, M.; Pistikopoulos, E.N.; Mantalaris, A. Transcriptional kinetics of the cross-talk between the ortho-cleavage and TOL pathways of toluene biodegradation in Pseudomonas putida mt-2. J. Biotechnol. 2016, 228, 112–123. [Google Scholar] [CrossRef]
- Tsipa, A.; Koutinas, M.; Vernardis, S.; Mantalaris, A. The impact of succinate trace on pWW0 and ortho-cleavage pathway transcription in Pseudomonas putida mt-2 during toluene biodegradation. Bioresour. Technol. 2017, 234, 397–405. [Google Scholar] [CrossRef]
- Koutinas, M.; Kiparissides, A.; Silva-Rocha, R.; Lam, M.C.; Martins dos Santos, V.A.P.; de Lorenzo, V.; Pistikopoulos, E.N.; Mantalaris, A. Linking genes to microbial growth kinetics-An integrated biochemical systems engineering approach. Metab. Eng. 2011, 13, 401–413. [Google Scholar] [CrossRef]
- Tsipa, A.; Pitt, J.A.; Banga, J.R.; Mantalaris, A. A dual-parameter identification approach for data-based predictive modeling of hybrid gene regulatory network-growth kinetics in Pseudomonas putida mt-2. Bioprocess. Biosyst. Eng. 2020, 43, 1671–1688. [Google Scholar] [CrossRef] [PubMed]
- Buldum, G.; Tsipa, A.; Mantalaris, A. Linking Engineered Gene Circuit Kinetic Modeling to Cellulose Biosynthesis Prediction in Escherichia coli: Toward Bioprocessing of Microbial Cell Factories. Ind. Eng. Chem. Res. 2020, 59, 4659–4669. [Google Scholar] [CrossRef]
- Koutinas, M.; Kiparissides, A.; Pistikopoulos, E.N.; Mantalaris, A. Bioprocess systems engineering: Transferring traditional process engineering principles to industrial biotechnology. Comput. Struct. Biotechnol. J. 2012, 3, e201210022. [Google Scholar] [CrossRef]
- Kiparissides, A.; Koutinas, M.; Kontoravdi, C.; Mantalaris, A.; Pistikopoulos, E.N. “Closing the loop” in biological systems modeling-from the in silico to the in vitro. Automatica 2011, 47, 1147–1155. [Google Scholar] [CrossRef]
- Cooke, K.G.; Gay, M.O.; Radachowsky, S.E.; Guzman, J.J.; Chiu, M.A. BackyardNetTM: Distributed Sensor Network Powered by Terrestrial Microbial Fuel Cell. In Proceedings of the Unattended Ground, Sea, and Air Sensor Technologies and Applications XII. International Society for Optics and Photonics, Orlando, FL, USA, 5–8 April 2021. [Google Scholar]
- Prasad, J.; Tripathi, R.K. Scale-up and control the voltage of sediment microbial fuel cell for charging a cell phone. Biosens. Bioelectron. 2021, 172, 112767. [Google Scholar] [CrossRef] [PubMed]
- Pietrelli, A.; Lovecchio, N.; Ferrara, V.; Allard, B. Custom measuring system tailored for MFCs. In Proceedings of the 2019 IEEE 8th International Workshop on Advances in Sensors and Interfaces (IWASI), Otranto, Italy, 13–14 June 2019; pp. 270–273. [Google Scholar]
- Logroño, W.; Guambo, A.; Pérez, M.; Kadier, A.; Recalde, C. A terrestrial single chamber microbial fuel cell-based biosensor for biochemical oxygen demand of synthetic ricewashed wastewater. Sensors 2016, 16, 101. [Google Scholar] [CrossRef] [PubMed]
- Sartori, D.; Brunelli, D. A smart sensor for precision agriculture powered by micro-bial fuel cells. In Proceedings of the IEEE Sensors Applications Symposium (SAS), Catania, Italy, 20–22 April 2016; pp. 1–6. [Google Scholar]
- Rossi, M.; Tosato, P.; Gemma, L.; Torquati, L.; Catania, C.; Camalo, S.; Brunelli, D. Long range wireless sensing powered by plant-microbial fuel cell. In Proceedings of the Design, Automation & Test in Europe Conference & Exhibition (DATE), Lausanne, Switzerland, 27–31 March 2017; pp. 1651–1654. [Google Scholar]
- Song, Y.E.; Boghani, H.C.; Kim, H.S.; Kim, B.G.; Lee, T.; Jeon, B.H.; Premier, G.C.; Kim, J.R. Electricity production by the application of a low voltage DC-DC boost converter to a continuously operating flat-plate microbial fuel cell. Energies 2017, 10, 596. [Google Scholar] [CrossRef]
- Prasad, J.; Tripathi, R.K. A Dc-Dc Boost Converter for Sediment Microbial Fuel Cell Energy Harvesting. In Proceedings of the 2nd IEEE International Conference on Power Electronics, Intelligent Control and Energy Systems (ICPEICES), New Delhi, India, 22–24 October 2018; pp. 712–716. [Google Scholar]
- Kim, T.; Yeo, J.; Yang, Y.; Kang, S.; Paek, Y.; Kwon, J.K.; Jang, J.K. Boosting voltage without electrochemical degradation using energy-harvesting circuits and power management system-coupled multiple microbial fuel cells. J. Power Sources 2019, 410–411, 171–178. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Microorganisms | Substrate [34] | Type of MFC [34] | Compounds Involved in Electron Transfer | Reference |
|---|---|---|---|---|
| Geobacter spp. | ||||
| Geobacter sulfurreducens | acetate, uranyl acetate, butyrate, ethanol | single-, dual-, upflow chamber | monolayer biofilms: c-Cyts (OmcZ, OmcB), alternatively dehydrogenases, quinones, iron-sulfur proteins, and b-Cyts or riboflavin (complex with c-Cyts) thick multilayer biofilms: microbial nanowires = conductive type IV pili (pilA protein monomer units) | [35,36,37,38,39,40,41,42] |
| Geobacter metallireducens | acetate, domestic wastewater | dual chamber | c-Cyts (OmcB in heterogeneous and OmcE in homogeneous electron transfer, respectively) | [43,44] |
| Geobacter anodireducens | acetate, domestic wastewater | single chamber | direct interspecies electron transfer (DIET) | [45] |
| Geobacter sulfurreducens and Geobacter metallireducens | ethanol | single chamber | DIET in the presence of anthraquinone-2,6-disulfonate: conductive pili aggregates | [37,46,47] |
| Geobacter metallireducens and citrate synthase-deficient Geobacter sulfurreducens | ethanol | single chamber | DIET | [48] |
| Geobacter metallireducens and Methanosaeta harundinacea | ethanol, acetate | single chamber | DIET | [33,49] |
| Geobacter metallireducens and Methanosarcina spp. | ethanol, acetate | single chamber | DIET | [33,50] |
| Shewanella spp. | ||||
| Shewanella oneidensis | lactate | single-, mini chamber | riboflavin and flavin mononucleotide = riboflavin-5-phosphate (flavin-c-Cyts complexes) | [51,52,53,54,55] |
| Shewanella oneidensis MR-1 | lactate | single chamber | riboflavin and riboflavin-5-phosphate (complex with decaheme c-Cyts MtrC and OmcA) | [51,52,53,54,56,57] |
| Shewanella oneidensis DSP10 | lactate | mini chamber | riboflavin and flavin mononucleotide | [55] |
| Shewanella loihica PV-4 | lactate, lactic acid, formic acid, cyclodextrin, galactose, arabinose, glucose | single chamber | quinone derivatives and riboflavin or c-Cyts | [56,58] |
| Shewanella sp. MR-4 | lactate | single chamber | riboflavin and riboflavin-5-phosphate | [57] |
| Shewanella putrefaciens | lactate | single chamber | c-Cyts (MtrC and OmcA) | [59] |
| Other Microorganisms | ||||
| Aeromonas hydrophila | acetate | single chamber | c-Cyts | [60] |
| Pseudomonas aeruginosa | glucose | dual chamber | quorum sensing (QS) chemicals: pyocyanin and phenazine-1-carboxamide or type IV pili | [61,62,63,64] |
| Pseudomonas putida, Pseudomonas fluorescens, Pseudomonas aeruginosa and Burkholderia cenocepacia | organic compounds | flow chamber | cyclic diguanosine-5′-monophosphate (c-di-GMP), small RNAs (sRNA) and QS | [64] |
| Clostridium spp. T1,3 | wastewater, glucose, Chlorella vulgaris (microalgae) T2 | single chamber | electrochemically inactive bacteria | [65,66,67,68] |
| Enterococcus gallinarum | glucose | single chamber | DIET | [69,70] |
| Synechocystis spp. | lactate | single chamber | microbial nanowires | [71] |
| Ochrobactrum anthropi | acetate, lactate, propionate, butyrate, glucose, sucrose, cellobiose, glycerol, ethanol | U-tube shaped | DIET | [72] |
| Thermincola ferriacetica | acetate | dual chamber | anthraquinone-2,6-disulfonate | [73] |
| Thermincola potens | acetate | dual chamber | multiheme c-Cyts (MHCs) or anthraquinone-2,6-disulfonate | [74] |
| Geothrix fermentans | acetate, lactate, malate, propionate, components of peptone, yeast extract | dual chamber | riboflavin and a still unknown compound | [75,76] |
| Desulfovibrio alaskensis G20 | lactate, organic acids, formate, short-chain alcohols | single chamber | type I tetraheme cytochrome c3 and transmembrane complexes (QrcA) | [77] |
| Desulfovibrio desulfuricans | lactate | dual chamber | c-Cyts | [78] |
| Lactococcus lactis | glucose | dual chamber | quinones (at least two soluble redox mediators required with the one being 2-amino-3-dicarboxy-1,4-naphthoquinone) | [79] |
| Escherichia coli T3 | glucose | single chamber | through unknown intermediaries [80] | [81] |
| Tissierella Clostridium and Alkaliphilus spp. | yeast extract, acetate, lactate, ethanol, methanol, sucrose | Alkaline Fuel Cell (AFC) | flavin species (indistinguishable from riboflavin) | [82] |
| Klebsiella pneumoniae | glucose | single chamber | 2,6-di-tert-butyl- p-benzoquinone | [83] |
| Rhodopseudomonas palustris DX-1 | acetate, volatile acids, yeast extract, thiosulfate | dual-, micro chamber | c-Cyts | [84,85] |
| Rhodopseudomonas palustris | acetate Arthrospira maxima T2, glycerol | micro- | c-Cyts but unknown intermediaries when Arthrospira maxima as a substrate | [85] |
| Saccharomyces cerevisiae | wastewater | single chamber | through unknown intermediaries [80] | [86] |
| Hansenula anomala | glucose, lactate | dual chamber | DIET | [87] |
| Candida melibiosica | glucose, fructose, sucrose | dual chamber | indirect electron transfer | [88] |
| Lysinibacillus sphaericus VA5 | beef extract | dual chamber | through unknown intermediaries [80] | [89] |
| Citrobacter sp. SX-1 | citrate, acetate, glucose, sucrose, glycerol, lactose | single chamber | through unknown intermediaries [80] | [90] |
| Raoultella electrica sp. | glucose | dual chamber | quinones (Q-8 major, also Q-9 and Q-10) | [91] |
| Ochrobactrum sp. 575 | xylose | dual chamber | through unknown intermediaries [80] | [92] |
| Cellulomonas spp. | cellulose | dual-, single chamber | DIET | [93,94] |
| Gluconobacter oxydans | ethanol, glucose | dual-, single chamber | c-Cyts or DIET or microbial nanowires | [95,96,97] |
| Gluconobacter thailandicus | glucose | current production still unexplored | membrane-bound dehydrogenases with pyrroloquinoline quinone (PQQ) and NAD(P)+-dependent enzymes | [98] |
| Models | Model Approach | Substrate | Microbial Growth Kinetic Models Used | Microbial Culture | Electrode Modeled | |
|---|---|---|---|---|---|---|
| 1 | Zhang and Halme [109] | ODE | - | Monod | Pure culture | anode |
| 2 | Picioreanu et al. [110] | ODE and PDE | acetate | Monod | Pure culture | anode |
| 3 | Zeng et al. [111] | ODE | acetate, solution of glucose and glutamic acid | Monod | Pure culture | anode and cathode |
| 4 | Pinto et al. [112] | ODE | acetate | Double Monod | Dual species | anode |
| 5 | Pinto et al. [120] | ODE | synthetic wastewater | Double Monod | Dual species | anode |
| 6 | Kato Marcus et al. [121] | ODE and PDE | glucose | Nernst-Monod | Dual species | anode |
| 7 | Oliveira et al. [113] | ODE | acetate | Monod | Pure culture | anode and cathode |
| 8 | Sirinutsomboon [122] | ODE | molasses | Monod + Nernst-Monod | Pure culture | anode and cathode |
| 9 | Jayasinghe et al. [114] | PDE | ammonium | Nernst-Monod | Pure culture | anode |
| 10 | Merkey and Chopp [107] | acetate | Nernst-Monod | Dual species | anode | |
| 11 | Picioreanu et al. [123] | ODE and PDE | acetate | Double Monod | Multiple species | anode |
| 12 | Picioreanu et al. [124] | ODE and PDE | acetate | Double Monod | Multiple species | anode |
| 16 | Recio-Garrido et al. [115] | ODE | acetate | Double Monod | Dual species | anode |
| 17 | Esfandyari et al. [117] | ODE | lactate | Monod, Backman and Tessier | Pure culture | anode and cathode |
| 18 | Esfandyari et al. [118] | ODE | lactate | Nernst-Monod | Pure culture | anode and cathode |
| Gadkari et al. [7] | ODE | - | Multiplicative Monod | Dual species | anode | |
| Karamzadeh et al. [119] | ODE and PDE | dairy wastewater | Nernst-Monod | Dual species | anode |
| Type of Models | Specific Growth Rate | Ref. | |
|---|---|---|---|
| Typical model | Monod | [105] | |
| Inhibition models | Haldane | [1] | |
| Aiba et al. | [125] | ||
| Tessier | [125] | ||
| Edwards | [6] | ||
| Luong | [6] | ||
| Hans-Levenspiel | [6] | ||
| Moser | [8] | ||
| Blackman | [117] | ||
| redox potential with the Monod kinetics | Monod–Nerst | [121] | |
| Multiplicative Monod | [123] | ||
| r: output voltage which can be obtained (mV), power density (mW/m2), current density (mA/m2) or substrate degradation rate (kg/m2 d) at each substrate concentration. rmax: maximum output voltage which can be obtained (mV), maximum power density (mW/m2), maximum current density (mA/m2) or maximum substrate degradation rate (kg/m2 d) among all the range of substrate concentration. S: substrate concentration (g/L). Ks: dissociation constant for substrate-enzyme binding (g/L) or half saturation coefficient. Ki: dissociation constant for inhibitory substrate-enzyme interactions (g/L). KIE: Edwards inhibition coefficient (g/L). Sm: critical inhibitory concentration above which the reaction stops (g/L). n and m: empirical constants. μmax: maximum specific growth rate. q: specific rate of electron donor utilization (mmol mg/VS d). vs.: volatile solids, a measure of biomass. qmax: maximum specific rate of electron donor utilization (mmol mg/VS d). φa: volumetric fraction of active biomass (dimensionless). Sd: electron donor concentration (mmol/cm3). KSd: half-max specific rate of electron donor concentration (mmol/cm3). F: Faraday constant (96,485 coulomb per mol-e−). R: ideal gas constant (8.3145 J/mol K);. T: temperature (298.15 K). ƞ: (Eanode − EKA), where Eanode is the potential of the anodic electron acceptor, EKA is the anodic acceptor potential for the half max-rate. SMox: concentration of the oxidized mediator (M). KSMox: half saturation coefficient of the mediator. | |||
| Low Power Commercial Transceiver Comparison | ||||
|---|---|---|---|---|
| Device [Protocol] | ActiveTx (I) (mA) | Supply Range (V) | Frequency (GHz) | Sleep Mode (I) (µA) |
| Nordic nrF24LE1 [802.11] | 11.1 | 1.9 to 3.6 | 2.4 | 0.550 |
| CC2533 [802.15.4] | 28.5 | 2 to 3.6 | 2.4 | 1 |
| CC1310 [802.15.4 g] | 13.7 | 1.8 to 3.8 | 0.686 | 7 |
| RN313C [802.11/Wi-Fi] | 210 | 2 to 3.3 | 2.4 | 4 |
| Enocean STM312 [802.15.4] | 100 | 2.1 to 5 | 0.868 | 4 |
| ATMEL ATA8510 [802.3] | 9.4 | 1.9 to 3.6 | 0.868 | 0.600 |
| MRF24J40MA [802.15.4] | 23 | 2.4 to 3.6 | 2.4 | 2 |
| XbeePRO S2B [802.15.4 ZigBee] | 233 | 3.1 to 3.46 | 2.4 | 4 |
| Energy Harvesting DC/DC Converter Comparison | |||||
|---|---|---|---|---|---|
| Converter | Start-Up VIN (mV) | VOUT (V) | MPPT/FOCV | Efficiency | IQ (µA) |
| LTC 3108 | 20 | 2.35/3.3/4.1/5 | No | 0.6 | 6 |
| LTC 3105 | 250 | 1.6 to 5.25 | FOCV | 0.6 | 24 |
| ECT 310 | 20 to 500 | 3 to 5 | No | 0.3 | N/A |
| S-882Z | 300 | 1.4 to 2.4 | No | 0.2 | N/A |
| MAX1722 | 400 | 1.8 to 5.5 | No | 0.95 | 0.300 |
| BQ25504 | 330 | 1.8 to 5 | MPPT | 0.8 | 0.330 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsipa, A.; Varnava, C.K.; Grenni, P.; Ferrara, V.; Pietrelli, A. Bio-Electrochemical System Depollution Capabilities and Monitoring Applications: Models, Applicability, Advanced Bio-Based Concept for Predicting Pollutant Degradation and Microbial Growth Kinetics via Gene Regulation Modelling. Processes 2021, 9, 1038. https://doi.org/10.3390/pr9061038
Tsipa A, Varnava CK, Grenni P, Ferrara V, Pietrelli A. Bio-Electrochemical System Depollution Capabilities and Monitoring Applications: Models, Applicability, Advanced Bio-Based Concept for Predicting Pollutant Degradation and Microbial Growth Kinetics via Gene Regulation Modelling. Processes. 2021; 9(6):1038. https://doi.org/10.3390/pr9061038
Chicago/Turabian StyleTsipa, Argyro, Constantina K. Varnava, Paola Grenni, Vincenzo Ferrara, and Andrea Pietrelli. 2021. "Bio-Electrochemical System Depollution Capabilities and Monitoring Applications: Models, Applicability, Advanced Bio-Based Concept for Predicting Pollutant Degradation and Microbial Growth Kinetics via Gene Regulation Modelling" Processes 9, no. 6: 1038. https://doi.org/10.3390/pr9061038
APA StyleTsipa, A., Varnava, C. K., Grenni, P., Ferrara, V., & Pietrelli, A. (2021). Bio-Electrochemical System Depollution Capabilities and Monitoring Applications: Models, Applicability, Advanced Bio-Based Concept for Predicting Pollutant Degradation and Microbial Growth Kinetics via Gene Regulation Modelling. Processes, 9(6), 1038. https://doi.org/10.3390/pr9061038