Fermentation of Ferulated Arabinoxylan Recovered from the Maize Bioethanol Industry

 , ,
, ,  , , and
, , and
Abstract
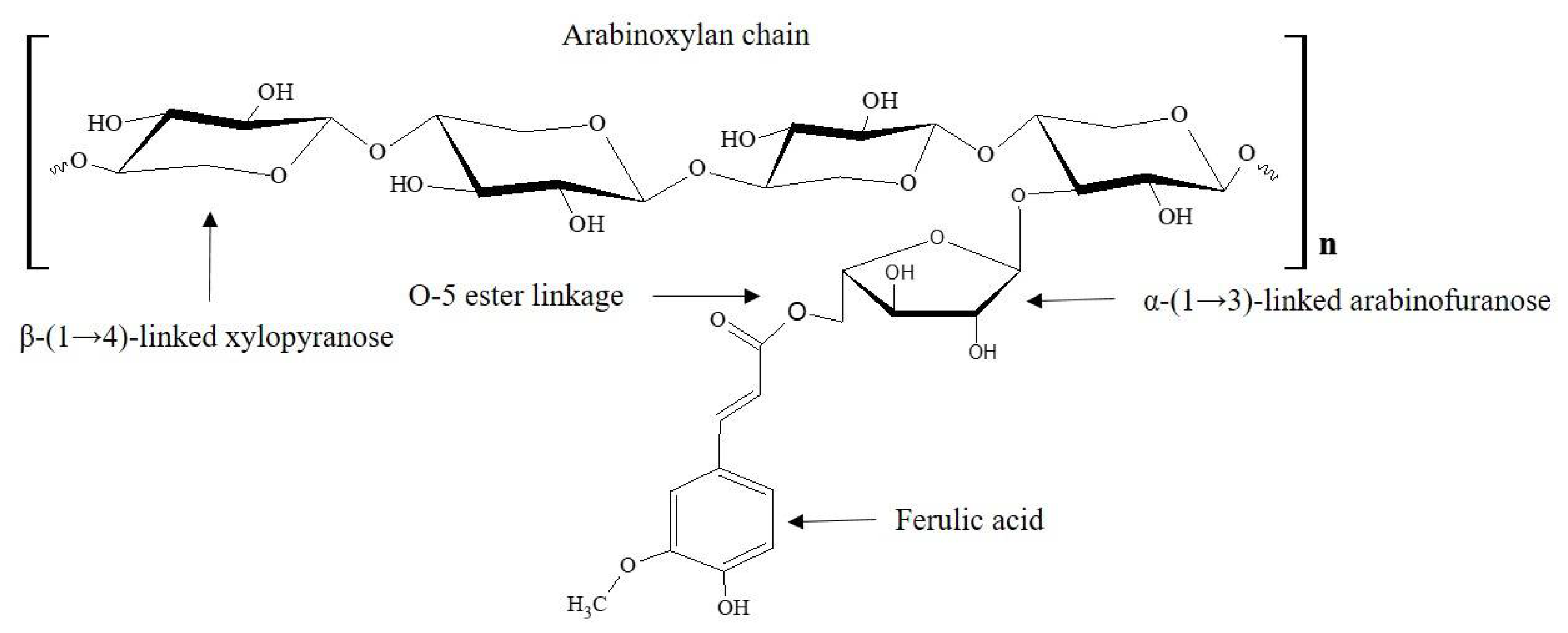
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Organisms and Culture Conditions
2.3. In Vitro Fermentation of AX
2.4. Fourier Transform Infrared (FTIR) Spectroscopy
2.5. Molecular Weight Distribution
2.6. Short-Chain Fatty Acids (SCFA)
2.7. Phenolic Acids
2.8. Antiproliferative Activity Assay
2.8.1. Cell Lines
2.8.2. Determination of Cell Proliferation
2.9. Statistical Analysis
3. Results and Discussion
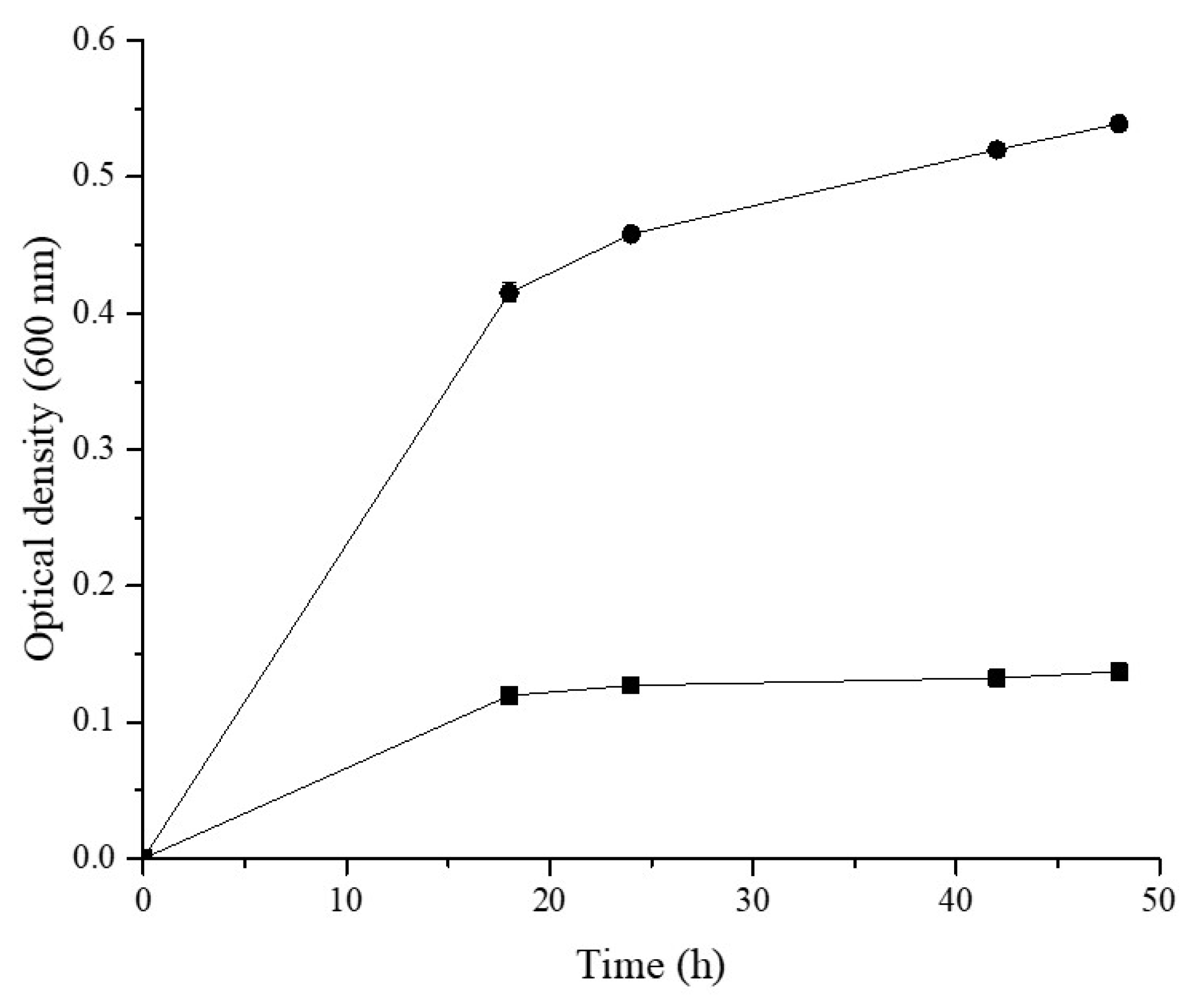
3.1. AX Fermentation
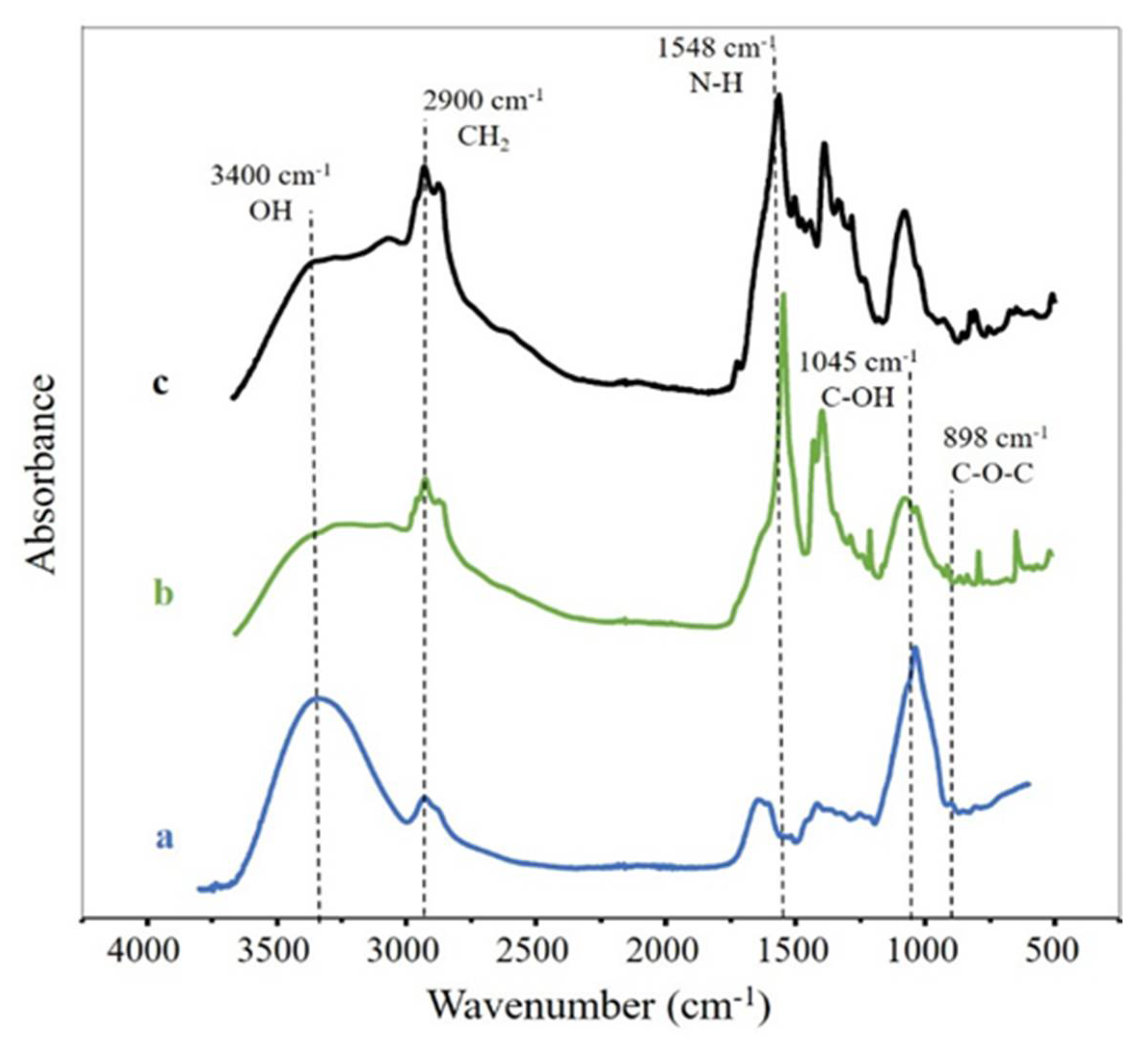
3.2. FTIR Spectroscopy
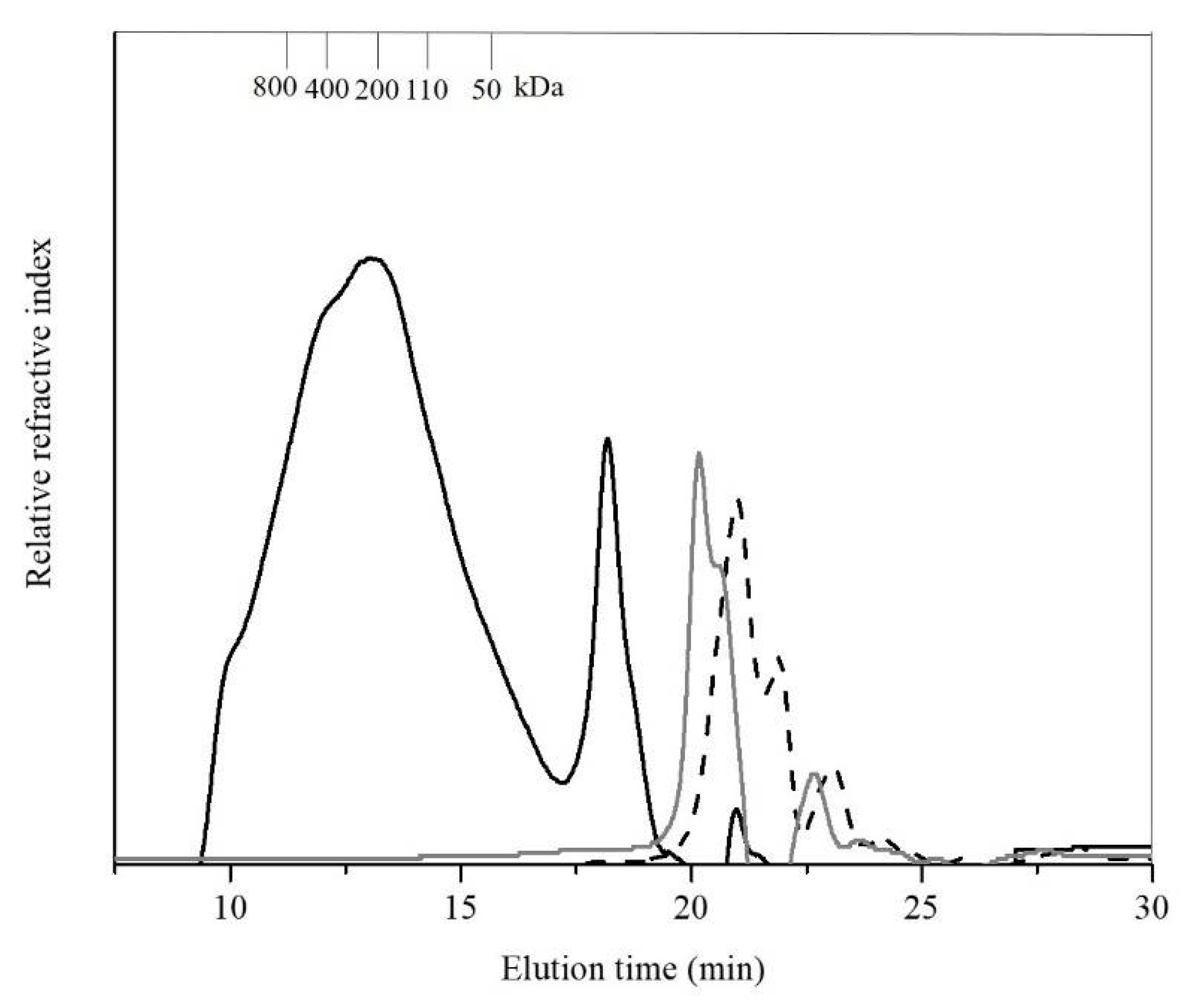
3.3. Molecular Weight Distribution
3.4. Phenolic Acids Contents
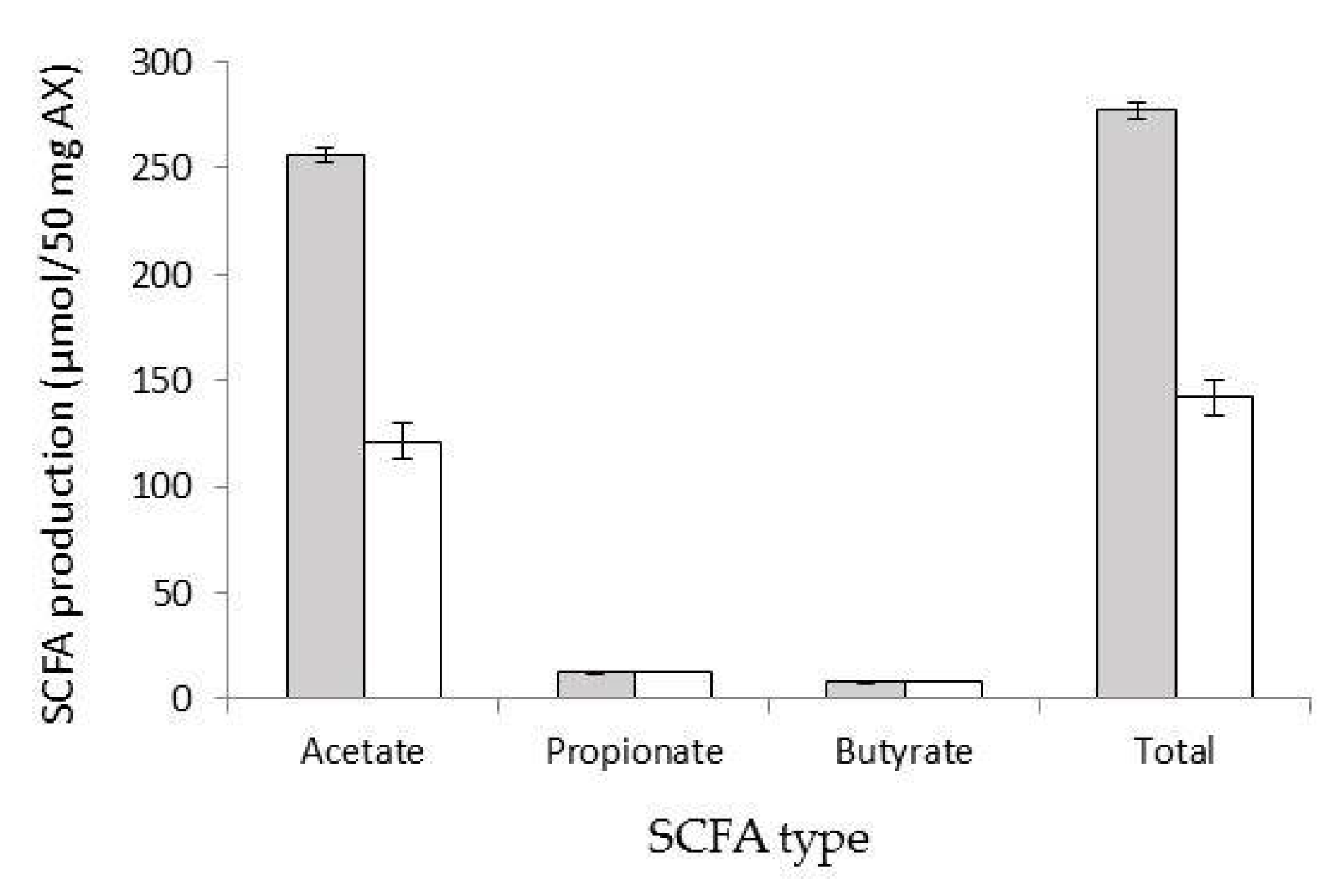
3.5. SCFA Production
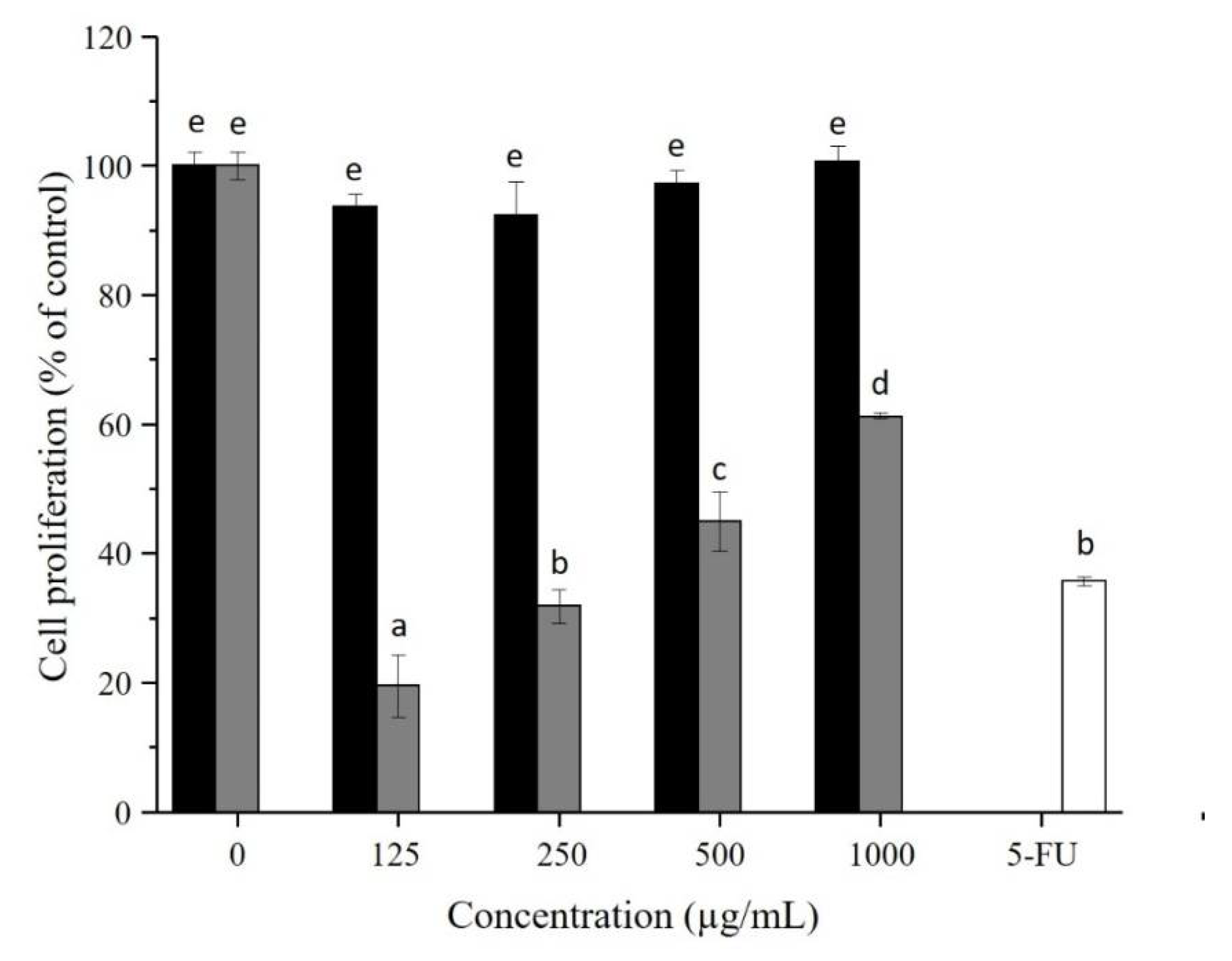
3.6. AX and AX-f Antiproliferative Activity on Caco-2 Colon Cancer Cells
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Izydorczyk, M.S.; Biliaderis, C.G. Arabinoxylans: Techonologically and nutrionally functional plant polysaccharides. In Functional Food Carbohydrates; Biliaderis, C.G., Izydorczyk, M.S., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 249–283. [Google Scholar]
- Smith, M.M.; Hartley, R.D. Occurrence and nature of ferulic acid substitution of cell-wall polysaccharides in graminaceous plants. Carbohydr. Res. 1983, 118, 65–80. [Google Scholar] [CrossRef]
- Chen, Z.; Li, S.; Fu, Y.; Li, C.; Chen, D.; Chen, H. Arabinoxylan structural characteristics, interaction with gut microbiota and potential health functions. J. Funct. Foods 2019, 54, 536–551. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Ohara, T.; Mori, T. Antiproliferative effects of short-chain fatty acids on human colorectal cancer cells via gene expression Inhibition. Anticancer Res. 2019, 39, 4659–4666. [Google Scholar] [CrossRef] [Green Version]
- Marques, C.; Oliveira, C.F.; Alves, S.; Chaves, S.; Coutinho, O.; Corte-Real, M.; Preto, A. Acetate-induced apoptosis in colorectal carcinoma cells involves lysosomal membrane permeabilization and cathepsin D release. Cell Death Dis. 2013, 4, e507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glei, M.; Hofmann, T.; Kuster, K.; Hollman, J.; Lindhauer, M.G.; Pool-Zobel, B.L. Both wheat (Triticum aestivum) bran arabinoxylans and gut flora-mediated fermentation products protect human colon cells from genotoxic activities of 4-hydroxynonenal and hydrogen peroxide. J. Agric. Food Chem. 2006, 54, 2088–2095. [Google Scholar] [CrossRef] [PubMed]
- Roy, N.; Narayanankutty, A.; Nazeem, P.A.; Babu, T.D.; Mathew, D. Plant phenolics ferulic acid and p-coumaric acid inhibit colorectal cancer cell proliferation through EGFR down- regulation. Asian Pac. J. Cancer Prev. 2016, 17, 4019–4023. [Google Scholar] [PubMed]
- Janicke, B.; Gunilla, O.; Oredsson, S.M. Differential effects of ferulic acid and p-coumaric acid on S phase distribution and length of S phase in the human colonic cell line Caco-2. J. Agric. Food Chem. 2005, 53, 6658–6665. [Google Scholar] [CrossRef] [PubMed]
- Janicke, B.; Hegardt, C.; Krogh, M.; Önning, G.; Åkesson, B.; Cirenajwis, H.M.; Oredsson, S.M. The antiproliferative effect of dietary fiber phenolic compounds ferulic acid and p-coumaric acid on the cell cycle of Caco-2 cells. Nutr. Cancer 2011, 63, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Rosa, L.S.; Silva, N.J.A.; Soares, N.C.P.; Monteiro, M.C.; Teodoro, A.J. Anticancer properties of phenolic acids in colon cancer—A review. J. Nutr. Food Sci. 2016, 6, 468. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, M.J.; Englyst, H.N.; Macfarlane, S.; Furrie, E.; Macfarlane, G.T.; Mcbain, A.J. Degradation of cross-linked and non-cross-linked arabinoxylans by the intestinal microbiota in children. Appl. Environ. Microbiol. 2003, 69, 6354–6360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, S.A.; Shewry, P.R.; Gibson, G.R.; Sanz, M.L.; Rastall, R.A. In vitro fermentation by human fecal microflora of wheat arabinoxylans. J. Agric. Food Chem. 2007, 55, 4589–4595. [Google Scholar] [CrossRef] [PubMed]
- Pastell, H.; Westermann, P.; Meyer, A.S.; Päivi, T.; Tenkanen, M. In vitro fermentation of arabinoxylan-derived carbohydrates by bifidobacteria and mixed fecal microbiota. J. Agric. Food Chem. 2009, 57, 8598–8606. [Google Scholar] [CrossRef] [PubMed]
- Snelders, J.; Olaerts, H.; Dornez, E.; Van de Wiele, T.; Aura, A.; Vanhaecke, L.; Delcour, J.A.; Courtin, C.M. Structural features and feruloylation modulate the fermentability and evolution of antioxidant properties of arabinoxylanoligosaccharides during in vitro fermentation by human gut derived microbiota. J. Funct. Foods 2014, 10, 1–12. [Google Scholar] [CrossRef]
- Mendez-Encinas, M.A.; Carvajal-Millan, E.; Yadav, M.P.; López-Franco, Y.L.; Rascon-Chu, A.; Lizardi-Mendoza, J.; Brown-Bojorquez, F.; Silva-Campa, E.; Pedroza-Montero, M. Partial removal of protein associated with arabinoxylans: Impact on the viscoelasticity, crosslinking content, and microstructure of the gels formed. J. Appl. Polym. Sci. 2019, 136, 47300. [Google Scholar] [CrossRef]
- van Laere, K.M.J.; Hartemink, R.; Bosveld, M.; Schols, H.A.; Voragen, A.G.J. Fermentation of plant cell wall derived polysaccharides and their corresponding oligosaccharides by intestinal bacteria. J. Agric. Food Chem. 2000, 48, 1644–1652. [Google Scholar] [CrossRef]
- Crittenden, R.; Karppinen, S.; Ojanen, S.; Tenkanen, M.; Fagerstrm, R.; Mtt, J.; Saarela, M.; Mattila-Sandholm, T.; Poutanen, K. In vitro fermentation of cereal dietary fibre carbohydrates by probiotic and intestinal bacteria. J. Sci. Food Agric. 2002, 82, 781–789. [Google Scholar] [CrossRef]
- Martínez-López, A.L.; Carvajal-Millan, E.; Micard, V.; Rascón-Chu, A.; Brown-Bojorquez, F.; Sotelo-Cruz, N.; López-Franco, Y.L.; Lizardi-Mendoza, J. In vitro degradation of covalently cross-linked arabinoxylan hydrogels by bifidobacteria. Carbohydr. Polym. 2016, 144, 76–82. [Google Scholar] [CrossRef]
- Zhao, G.; Nyman, M.; Jönsson, J.Å. Rapid determination of short-chain fatty acids in colonic contents and faeces of humans and rats by acidified water-extraction and direct-injection gas chromatography. Biomed. Chromatogr. 2006, 20, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Vansteenkiste, E.; Babot, C.; Rouau, X.; Micard, V. Oxidative gelation of feruloylated arabinoxylan as affected by protein. Influence on protein enzymatic hydrolysis. Food Hydrocoll. 2004, 18, 557–564. [Google Scholar] [CrossRef]
- Rouau, X.; Chaynier, V.; Surget, A.; Gloux, D.; Barron, C.; Meudeu, E.; Montero, J.L.; Criton, M. A dehydrotrimer of ferulic acid from maize bran. Phytochemistry 2003, 63, 899–903. [Google Scholar] [CrossRef]
- Samuelsen, A.B.; Rieder, A.; Grimmer, S.; Michaelsen, T.E.; Knutsen, S.H. Immunomodulatory activity of dietary fiber: Arabinoxylan and mixed-linked beta-glucan isolated from barley show modest activities in vitro. Int. J. Mol. Sci. 2011, 12, 570–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival - application to proliferation and cyto-toxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Hernandez, J.; Goycoolea, F.M.; Quintero, J.; Acosta, A.; Castañeda, M.; Dominguez, Z.; Robles, R.; Vazquez-Moreno, L.; Velazquez, E.F.; Astiazaran, H.; et al. Sonoran propolis: Chemical composition and antiproliferative activity on cancer cell lines. Planta Med. 2007, 73, 1469–1474. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Encinas, M.A.; Carvajal-Millan, E.; Rascón-Chu, A.; Astiazarán-García, H.; Valencia-Rivera, D.E.; Brown-Bojorquez, F.; Alday, E.; Velazquez, C. Arabinoxylan-based particles: In vitro antioxidant and cytotoxicity on a human colon cell line. Medicina 2019, 55, 349. [Google Scholar] [CrossRef] [Green Version]
- Paesani, C.; Salvucci, E.; Moiraghi, M.; Fernadez Canigia, L.; Pérez, G.T. Arabinoxylan from Argentinian whole wheat flour promote the growth of Lactobacillus reuteri and Bifidobacterium breve. Lett. Appl. Microbiol. 2018, 68, 142–148. [Google Scholar] [CrossRef]
- Rivière, A.; Moens, F.; Selak, M.; Maes, D.; Weckx, S.; De Vuyst, L. The ability of bifidobacteria to degrade arabinoxylan oligosaccharide constituents and derived oligosaccharides is strain dependent. Appl. Environ. Microbiol. 2014, 80, 204–217. [Google Scholar] [CrossRef] [Green Version]
- Hromádková, Z.; Smestad, B.; Polovka, M.; Košťálová, Z.; Ebringerová, A. Structural features of two heteroxylan polysaccharide fractions from wheat bran with anti-complementary and antioxidant activities. Carbohydr. Polym. 2013, 93, 22–30. [Google Scholar] [CrossRef]
- Morales-Ortega, A.; Carvajal-Millan, E.; López-Franco, Y.; Rascón-Chu, A.; Lizardi-Mendoza, J.; Torres-Chavez, P.; Campa-Mada, A. Characterization of water extractable arabinoxylans from a spring wheat flour: Rheological properties and microstructure. Molecules 2013, 18, 8417–8428. [Google Scholar] [CrossRef] [Green Version]
- Egüés, I.; Stepan, A.M.; Eceiza, A.; Toriz, G.; Gatenholm, P.; Labidi, J. Corncob arabinoxylan for new materials. Carbohydr. Polym. 2014, 102, 12–20. [Google Scholar] [CrossRef]
- Urias-Orona, V.; Huerta-Oros, J.; Carvajal-Millán, E.; Lizardi-Mendoza, J.; Rascón-Chu, A.; Gardea, A.A. Component analysis and free radicals scavenging activity of Cicer arietinum L. husk pectin. Molecules 2010, 15, 6948–6955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollet, A.; Van Craeyveld, V.; Van De Wiele, T.; Verstraete, W.; Delcour, J.A.; Courtin, C.M. In vitro fermentation of arabinoxylan oligosaccharides and low molecular mass arabinoxylans with different structural properties from wheat (Triticum aestivum L.) bran and psyllium (Plantago ovata Forsk) seed husk. J. Agric. Food Chem. 2012, 60, 946–954. [Google Scholar] [CrossRef]
- Feng, G.; Flanagan, B.M.; Mikkelsen, D.; Williams, B.A.; Yu, W.; Gilbert, R.G.; Gidley, M.J. Mechanisms of utilisation of arabinoxylans by a porcine faecal inoculum: Competition and co-operation. Sci. Rep. 2018, 8, 4546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Anda-Flores, Y.; Carvajal-Millan, E.; Lizardi-Mendoza, J.; Rascon-Chu, A.; Martínez-López, A.L.; Marquez-Escalante, J.; Brown-Bojorquez, F.; Tanori-Cordova, J. Covalently cross-linked nanoparticles based on ferulated arabinoxylans recovered from a distiller’s dried grains byproduct. Processes 2020, 8, 691. [Google Scholar] [CrossRef]
- Rascon-Chu, A.; Martínez-López, A.L.; Berlanga-Reyes, C.; Carvajal-Millan, E.; Campa-Mada, A.C.; Gardea, A.A.; Orozco-Avitia, A. Arabinoxylans gels as lycopene carriers: In vitro degradation by colonic bacteria. Mater. Res. Soc. Symp. Proc. 2012, 2012, 1487. [Google Scholar] [CrossRef]
- Kaur, A.; Rose, D.J.; Rumpagaporn, P.; Patterson, J.A.; Hamaker, B.R. In vitro batch fecal fermentation comparison of gas and short-chain fatty acid production using “slowly fermentable” dietary fibers. J. Food Sci. 2011, 76, 137–142. [Google Scholar] [CrossRef]
- Macy, J.; Ljungdahl, L.; Gottschalk, G. Pathway of succinate and propionate formation in Bacteroides fragilis. J. Bacteriol. 1978, 134, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef]
- Mendez-Encinas, M.; Carvajal-Millan, E.; Rascon-Chu, A.; Astiazaran-Garcia, H.; Valencia-Rivera, D. Ferulated arabinoxylans and their gels: Functional properties and potential application as antioxidant and anticancer agent. Oxid. Med. Cell. Longev. 2018, 2018, 22. [Google Scholar] [CrossRef] [Green Version]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; de los Reyes-Gavilán, C.G.; Salazar, N. Intestinal short chain fatty acids and their link with diet and human health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [Green Version]
- McNabney, S.M.; Henagan, T.M. Short chain fatty acids in the colon and peripheral tissues: A focus on butyrate, colon cancer, obesity and insulin resistance. Nutrients 2017, 9, 1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; Vuyst, L. De Bifidobacteria and butyrate-producing colon bacteria: Importance and strategies for their stimulation in the human gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Smith, C.; Li, W.; Ashworth, J. Characterization of nitric oxide modulatory activities of alkaline-extracted and enzymatic-modified arabinoxylans from corn bran in cultured human monocytes. J. Agric. Food Chem. 2016, 64, 8128–82137. [Google Scholar] [CrossRef]
- Niño-Medina, G.; Carvajal-Millán, E.; Lizardi, J.; Rascon-Chu, A.; Marquez-Escalante, J.; Gardea, A.; Martinez-Lopez, A.L.; Guerrero, V. Maize processing waste water arabinoxylans: Gelling capability and cross-linking content. Food Chem. 2009, 115, 1286–1290. [Google Scholar] [CrossRef]
- Arun, K.B.; Madhavan, A.; Reshmitha, T.R.; Thomas, S.; Nisha, P. Short chain fatty acids enriched fermentation metabolites of soluble dietary fibre from Musa paradisiaca drives HT29 colon cancer cells to apoptosis. PLoS ONE 2019, 14, e0216604. [Google Scholar]
- Wang, T.; Gong, X.; Jiang, R.; Li, H.; Du, W.; Kuang, G. Ferulic acid inhibits proliferation and promotes apoptosis via blockage of PI3K/Akt pathway in osteosarcoma cell. Am. J. Transl. Res. 2016, 8, 968–980. [Google Scholar]
- Longley, D.B.; Harkin, D.P.; Johnston, P.G. 5-Fluorouracil: Mechanisms of action and clinical strategies. Nat. Rev. Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Esterified | |||
| Sample | FA | di-FA | tri-FA |
| µg/mg Sample (Dry Weight) | |||
| AX | 5.45 ± 0.09 | 0.35 ± 0.07 | 0.03 ± 0.00 |
| AX-f | 2.07 ± 0.03 | 0.03 ± 0.01 | nd |
| Control | nd | nd | nd |
| Non-Esterified | |||
| Sample | FA | di-FA | tri-FA |
| µg/mg Sample (Dry Weight) | |||
| AX | Nd | nd | nd |
| AX-f | 1.14 ± 0.06 | nd | nd |
| Control | nd | nd | nd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendez-Encinas, M.A.; Valencia-Rivera, D.E.; Carvajal-Millan, E.; Astiazaran-Garcia, H.; Micard, V.; Rascón-Chu, A. Fermentation of Ferulated Arabinoxylan Recovered from the Maize Bioethanol Industry. Processes 2021, 9, 165. https://doi.org/10.3390/pr9010165
Mendez-Encinas MA, Valencia-Rivera DE, Carvajal-Millan E, Astiazaran-Garcia H, Micard V, Rascón-Chu A. Fermentation of Ferulated Arabinoxylan Recovered from the Maize Bioethanol Industry. Processes. 2021; 9(1):165. https://doi.org/10.3390/pr9010165
Chicago/Turabian StyleMendez-Encinas, Mayra A., Dora E. Valencia-Rivera, Elizabeth Carvajal-Millan, Humberto Astiazaran-Garcia, Valérie Micard, and Agustín Rascón-Chu. 2021. "Fermentation of Ferulated Arabinoxylan Recovered from the Maize Bioethanol Industry" Processes 9, no. 1: 165. https://doi.org/10.3390/pr9010165
APA StyleMendez-Encinas, M. A., Valencia-Rivera, D. E., Carvajal-Millan, E., Astiazaran-Garcia, H., Micard, V., & Rascón-Chu, A. (2021). Fermentation of Ferulated Arabinoxylan Recovered from the Maize Bioethanol Industry. Processes, 9(1), 165. https://doi.org/10.3390/pr9010165