Electrophysiological Effects of Extracellular Vesicles Secreted by Cardiosphere-Derived Cells: Unraveling the Antiarrhythmic Properties of Cell Therapies

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
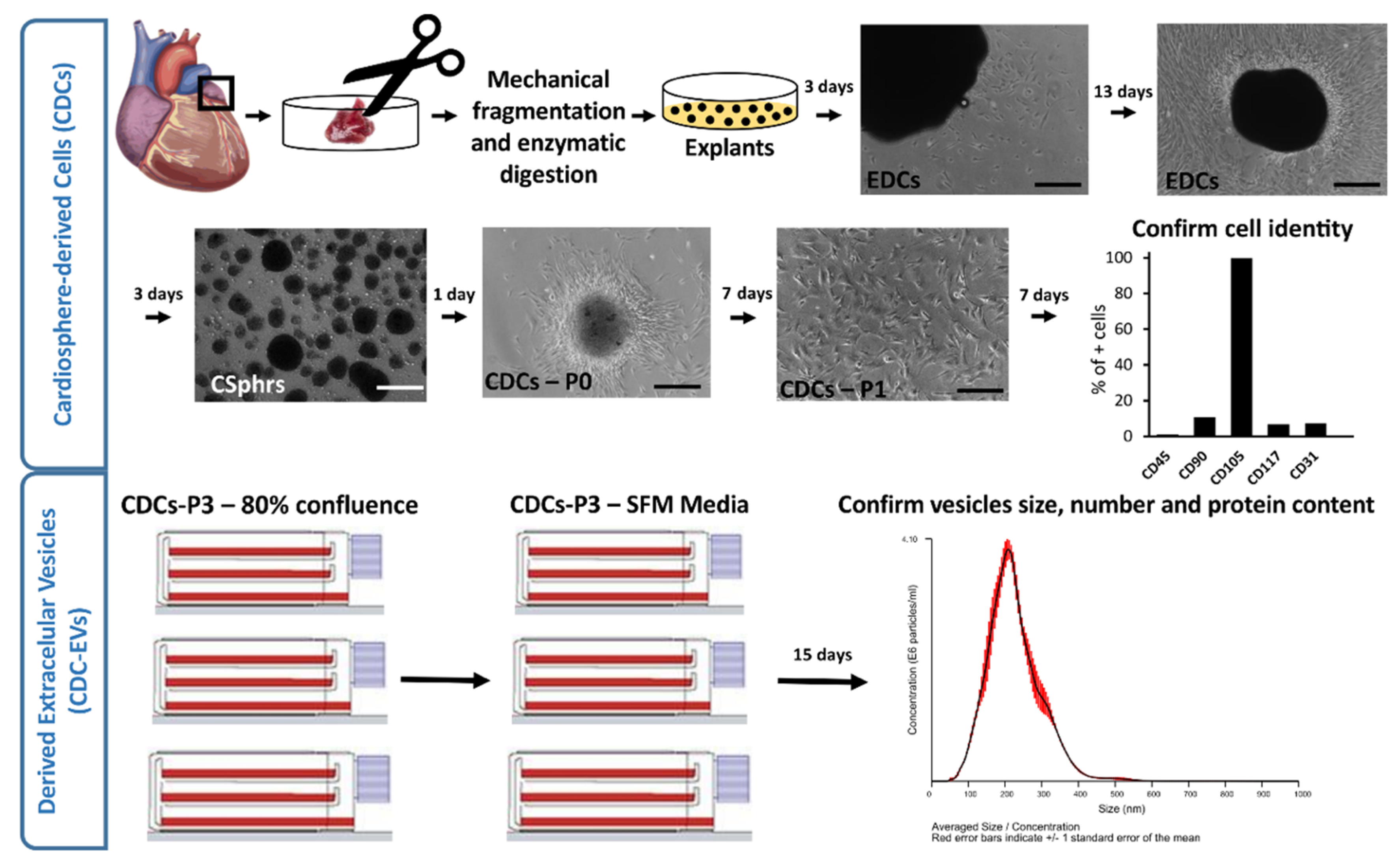
2.1. CDCs and Derived Extracellular Vesicle Isolation
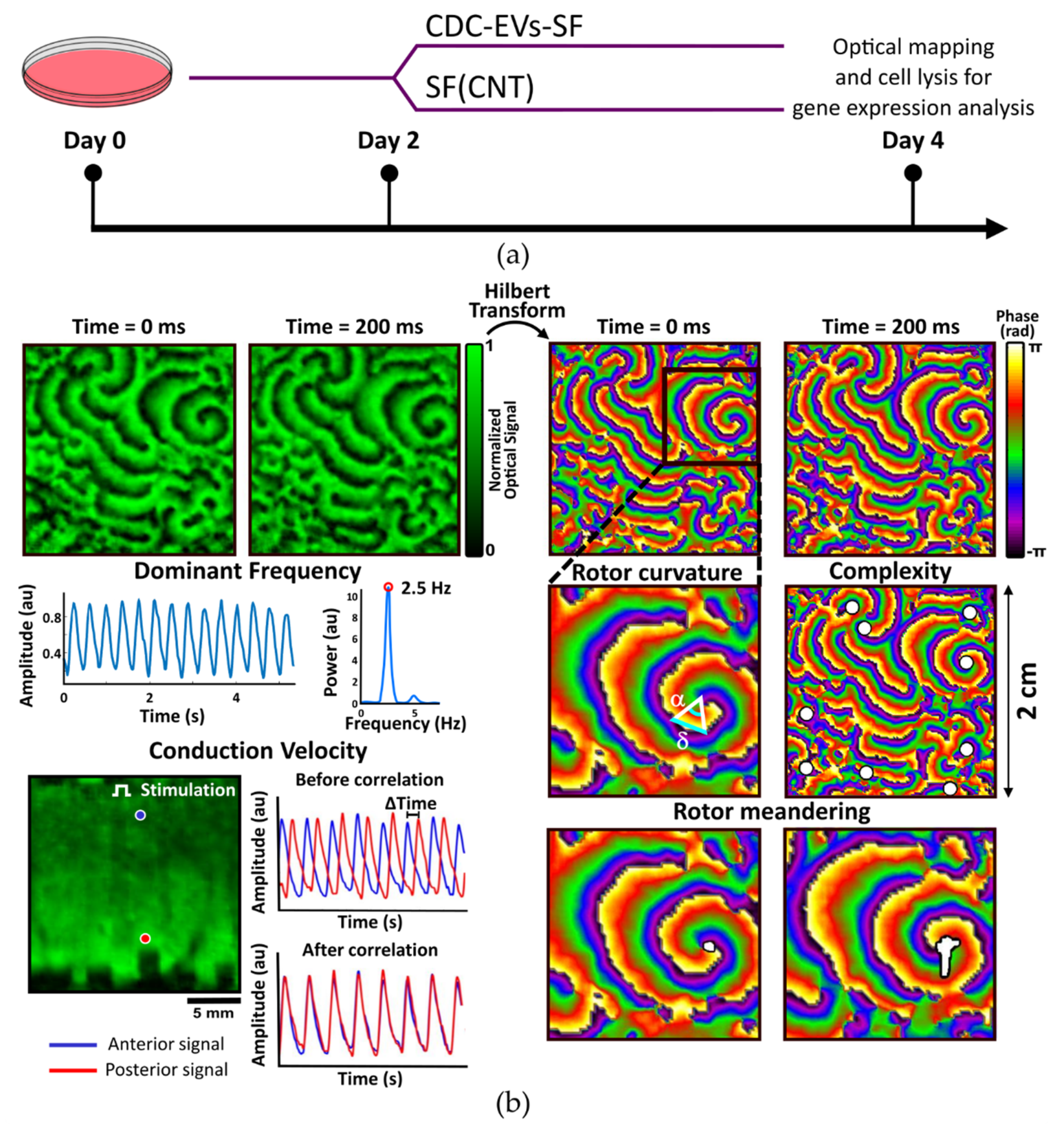
2.2. Experimental Protocol
2.3. Optical Mapping Recordings
2.4. Calcium Image Processing
2.4.1. Spectral Analysis
2.4.2. Phase Singularities and Rotor Dynamics
2.4.3. Conduction Velocity
2.5. Gene Expression Analysis
2.6. Statistical Analysis
3. Results
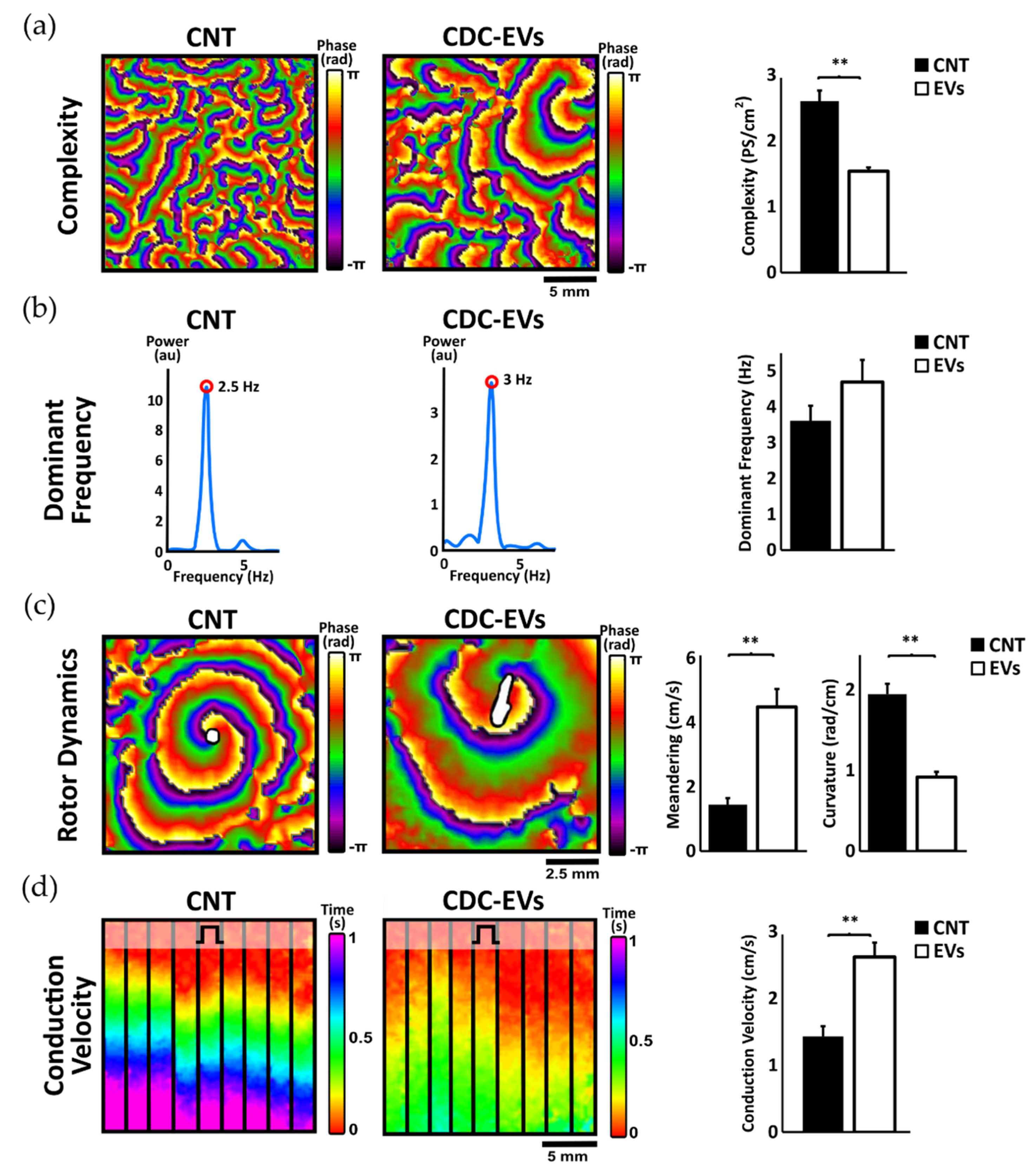
3.1. Effects of Extracellular Vesicles Derived from CDCs on Activation Dominant Frequency, Complexity, and Rotor Dynamics
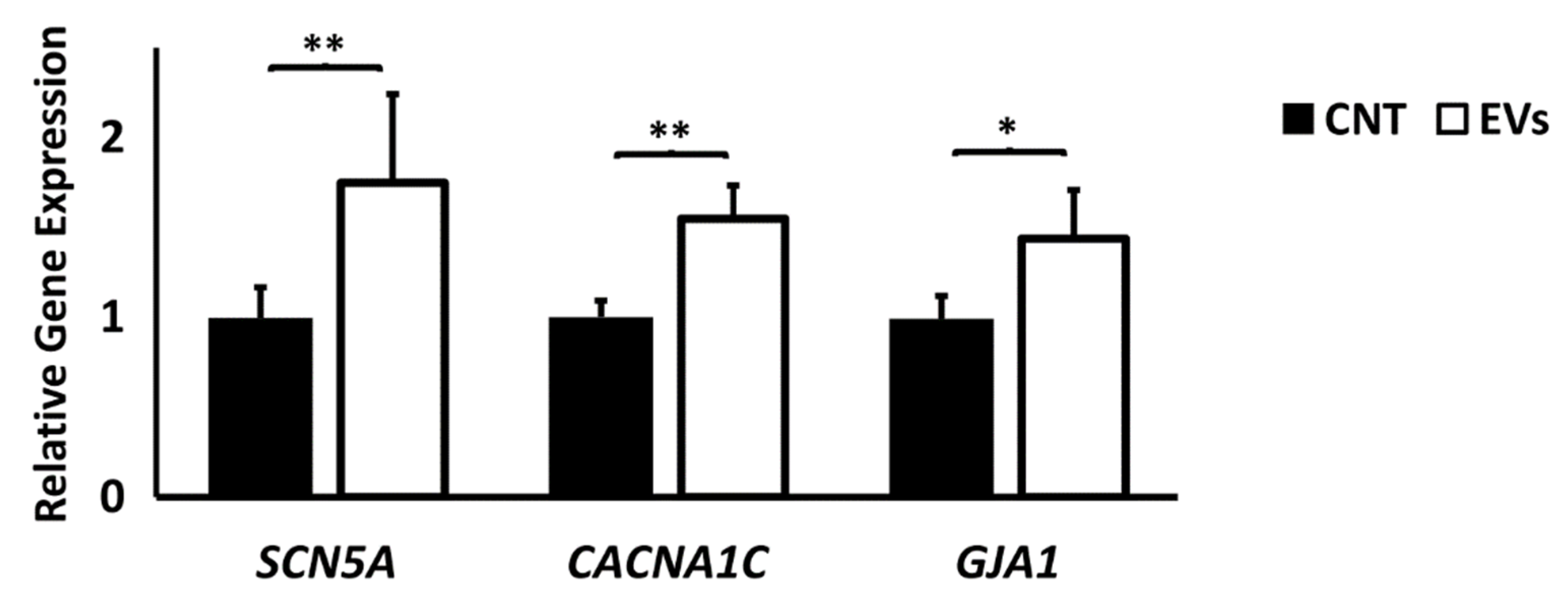
3.2. Extracellular Vesicles Derived from CDCs Increase Conduction Velocity in Arrhythmogenic Substrate and Increase Ion Channel Expression
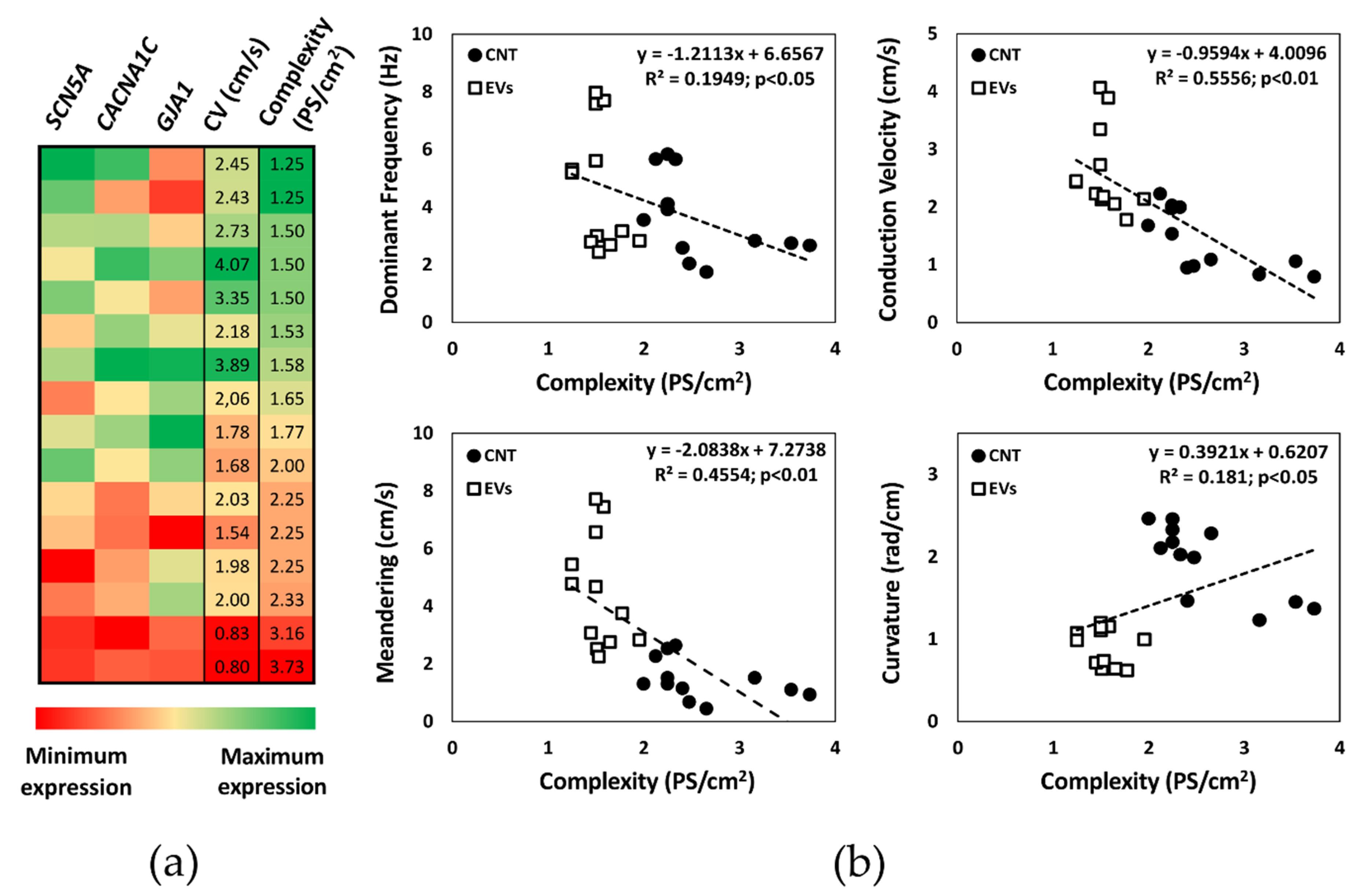
3.3. Activation Complexity Electrophysiological Mechanisms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Antzelevitch, C.; Burashnikov, A. Overview of Basic Mechanisms of Cardiac Arrhythmia. Card. Electrophysiol. Clin. 2011, 3, 23–45. [Google Scholar] [CrossRef] [PubMed]
- Perna, F.; Leo, M. Epidemiology, Classification and Description of Cardiac Arrhythmias. In Sports Cardiology: From Diagnosis to Clinical Management; Fioranelli, M., Frajese, G., Eds.; Springer: Milan, Italy, 2012; pp. 155–177. [Google Scholar]
- Khurshid, S.; Choi, S.H.; Weng, L.-C.; Wang, E.Y.; Trinquart, L.; Benjamin, E.J.; Ellinor, P.T.; Lubitz, S.A. Frequency of Cardiac Rhythm Abnormalities in a Half Million Adults. Circ. Arrhythm. Electrophysiol. 2018, 11, e006273. [Google Scholar] [CrossRef] [PubMed]
- Macle, L.; Nattel, S. Arrhythmias in 2015: Advances in drug, ablation, and device therapy for cardiac arrhythmias. Nat. Rev. Cardiol. 2016, 13, 67–68. [Google Scholar] [CrossRef] [PubMed]
- Gopinathannair, R.; Olshansky, B. Management of tachycardia. F1000Prime Rep. 2015, 7. [Google Scholar] [CrossRef]
- Cho, H.C.; Marbán, E. Biological therapies for cardiac arrhythmias: Can genes and cells replace drugs and devices? Circ. Res. 2010, 106, 674–685. [Google Scholar] [CrossRef]
- King, J.H.; Huang, C.L.-H.; Fraser, J.A. Determinants of myocardial conduction velocity: Implications for arrhythmogenesis. Front. Physiol. 2013, 4. [Google Scholar] [CrossRef]
- Aronis Konstantinos, N.; Berger Ronald, D. Ashikaga Hiroshi Rotors. Circ. Arrhythm. Electrophysiol. 2017, 10, e005634. [Google Scholar]
- Lau, D.H.; Clausen, C.; Sosunov, E.A.; Shlapakova, I.N.; Anyukhovsky, E.P.; Danilo, P.; Rosen, T.S.; Kelly, C.; Duffy, H.S.; Szabolcs, M.J.; et al. Epicardial border zone overexpression of skeletal muscle sodium channel SkM1 normalizes activation, preserves conduction, and suppresses ventricular arrhythmia: An in silico, in vivo, in vitro study. Circulation 2009, 119, 19–27. [Google Scholar] [CrossRef]
- Igarashi, T.; Finet, J.E.; Takeuchi, A.; Fujino, Y.; Strom, M.; Greener, I.D.; Rosenbaum, D.S.; Donahue, J.K. Connexin gene transfer preserves conduction velocity and prevents atrial fibrillation. Circulation 2012, 125, 216–225. [Google Scholar] [CrossRef]
- Golan, D.E.; Tashjian, A.H.; Armstrong, E.J. Principles of Pharmacology: The Pathophysiologic Basis of Drug Therapy; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2011. [Google Scholar]
- Haugan, K.; Miyamoto, T.; Takeishi, Y.; Kubota, I.; Nakayama, J.; Shimojo, H.; Hirose, M. Rotigaptide (ZP123) Improves Atrial Conduction Slowing in Chronic Volume Overload-Induced Dilated Atria. Basic Clin. Pharmacol. Toxicol. 2006, 99, 71–79. [Google Scholar] [CrossRef]
- Roell, W.; Lewalter, T.; Sasse, P.; Tallini, Y.N.; Choi, B.-R.; Breitbach, M.; Doran, R.; Becher, U.M.; Hwang, S.-M.; Bostani, T.; et al. Engraftment of connexin 43-expressing cells prevents post-infarct arrhythmia. Nature 2007, 450, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Rossello, R.A.; Kohn, D.H. Gap Junction Intercellular Communication: A Review of a Potential Platform to Modulate Craniofacial Tissue Engineering. J. Biomed. Mater. Res. B Appl. Biomater. 2009, 88, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Comtois, P.; Kneller, J.; Nattel, S. Of circles and spirals: Bridging the gap between the leading circle and spiral wave concepts of cardiac reentry. Europace 2005, 7 (Suppl. 2), 10–20. [Google Scholar] [CrossRef] [PubMed]
- Cha, Y.-M.; Friedman, P.A. Mayo Clinic Electrophysiology Manual; Oxford University Press: Oxford, UK, 2013. [Google Scholar]
- Qu, Z.; Xie, F.; Garfinkel, A.; Weiss, J. Origins of Spiral Wave Meander and Breakup in a Two-Dimensional Cardiac Tissue Model. Ann. Biomed. Eng. 2000, 28, 755–771. [Google Scholar] [CrossRef]
- Zipes, D.P.; Jalife, J. Cardiac Electrophysiology: From Cell to Bedside E-Book: Expert Consult; Elsevier Health Sciences: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Lim, H.S.; Hocini, M.; Dubois, R.; Denis, A.; Derval, N.; Zellerhoff, S.; Yamashita, S.; Berte, B.; Mahida, S.; Komatsu, Y.; et al. Complexity and Distribution of Drivers in Relation to Duration of Persistent Atrial Fibrillation. J. Am. Coll. Cardiol. 2017, 69, 1257–1269. [Google Scholar] [CrossRef]
- Albert, C.M.; Stevenson, W.G. The Future of Arrhythmias and Electrophysiology. Circulation 2016, 133, 2687–2696. [Google Scholar] [CrossRef]
- Raymond-Paquin, A.; Andrade, J.; Macle, L. Catheter ablation: An ongoing revolution. J. Thorac. Dis. 2019, 11, S212–S215. [Google Scholar] [CrossRef]
- Sadraddin, H.; Gaebel, R.; Skorska, A.; Lux, C.A.; Sasse, S.; Ahmad, B.; Vasudevan, P.; Steinhoff, G.; David, R. CD271+ Human Mesenchymal Stem Cells Show Antiarrhythmic Effects in a Novel Murine Infarction Model. Cells 2019, 8, 1474. [Google Scholar] [CrossRef]
- Park, H.; Park, H.; Mun, D.; Kang, J.; Kim, H.; Kim, M.; Cui, S.; Lee, S.-H.; Joung, B. Extracellular Vesicles Derived from Hypoxic Human Mesenchymal Stem Cells Attenuate GSK3β Expression via miRNA-26a in an Ischemia-Reperfusion Injury Model. Yonsei Med. J. 2018, 59, 736–745. [Google Scholar] [CrossRef]
- Mayourian, J.; Cashman, T.J.; Ceholski, D.K.; Johnson, B.V.; Sachs, D.; Kaji, D.A.; Sahoo, S.; Hare, J.M.; Hajjar, R.J.; Sobie, E.A.; et al. Experimental and Computational Insight Into Human Mesenchymal Stem Cell Paracrine Signaling and Heterocellular Coupling Effects on Cardiac Contractility and Arrhythmogenicity. Circ. Res. 2017, 121, 411–423. [Google Scholar] [CrossRef]
- Hwang, H.J.; Chang, W.; Song, B.-W.; Song, H.; Cha, M.-J.; Kim, I.-K.; Lim, S.; Choi, E.J.; Ham, O.; Lee, S.-Y.; et al. Antiarrhythmic potential of mesenchymal stem cell is modulated by hypoxic environment. J. Am. Coll. Cardiol. 2012, 60, 1698–1706. [Google Scholar] [CrossRef] [PubMed]
- Ramireddy, A.; Brodt, C.R.; Mendizabal, A.M.; DiFede, D.L.; Healy, C.; Goyal, V.; Alansari, Y.; Coffey, J.O.; Viles-Gonzalez, J.F.; Heldman, A.W.; et al. Effects of Transendocardial Stem Cell Injection on Ventricular Proarrhythmia in Patients with Ischemic Cardiomyopathy: Results from the POSEIDON and TAC-HFT Trials. Stem Cells Transl. Med. 2017, 6, 1366–1372. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Kilfoil, P.J.; Zhang, R.; Solymani, R.E.; Bresee, C.; Kang, E.M.; Luther, K.; Rogers, R.G.; de Couto, G.; Goldhaber, J.I.; et al. Reverse electrical remodeling in rats with heart failure and preserved ejection fraction. JCI Insight 2018, 3. [Google Scholar] [CrossRef]
- Cho, J.H.; Kilfoil, P.J.; Solymani, R.; Luther, K.; Rogers, R.G.; de Couto, G.; Zhang, R.; Sanchez, L.; Goldhaber, J.I.; Marban, E.; et al. Abstract 15421: Anti-Arrhythmic Effects of Heart-Derived Cell Therapy in a Rat Model of Heart Failure With Preserved Ejection Fraction. Circulation 2017, 136, A15421. [Google Scholar] [CrossRef] [PubMed]
- Askar, S.F.A.; Ramkisoensing, A.A.; Atsma, D.E.; Schalij, M.J.; de Vries, A.A.F.; Pijnappels, D.A. Engraftment patterns of human adult mesenchymal stem cells expose electrotonic and paracrine proarrhythmic mechanisms in myocardial cell cultures. Circ. Arrhythm. Electrophysiol. 2013, 6, 380–391. [Google Scholar] [CrossRef][Green Version]
- Eun, L.Y.; Song, H.; Choi, E.; Lee, T.G.; Moon, D.W.; Hwang, D.; Byun, K.H.; Sul, J.H.; Hwang, K.C. Implanted bone marrow-derived mesenchymal stem cells fail to metabolically stabilize or recover electromechanical function in infarcted hearts. Tissue Cell 2011, 43, 238–245. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef]
- Marbán, E. The Secret Life of Exosomes: What Bees Can Teach Us About Next-Generation Therapeutics. J. Am. Coll. Cardiol. 2018, 71, 193–200. [Google Scholar] [CrossRef]
- Ibrahim, A.G.-E.; Cheng, K.; Marbán, E. Exosomes as Critical Agents of Cardiac Regeneration Triggered by Cell Therapy. Stem Cell Rep. 2014, 2, 606–619. [Google Scholar] [CrossRef]
- Chen, B.; Li, Q.; Zhao, B.; Wang, Y. Stem Cell-Derived Extracellular Vesicles as a Novel Potential Therapeutic Tool for Tissue Repair. Stem Cells Transl. Med. 2017, 6, 1753–1758. [Google Scholar] [CrossRef]
- Makkar, R.R.; Smith, R.R.; Cheng, K.; Malliaras, K.; Thomson, L.E.; Berman, D.; Czer, L.S.; Marbán, L.; Mendizabal, A.; Johnston, P.V.; et al. Intracoronary cardiosphere-derived cells for heart regeneration after myocardial infarction (CADUCEUS): A prospective, randomised phase 1 trial. Lancet 2012, 379, 895–904. [Google Scholar] [CrossRef]
- Grigorian-Shamagian, L.; Liu, W.; Fereydooni, S.; Middleton, R.C.; Valle, J.; Cho, J.H.; Marbán, E. Cardiac and systemic rejuvenation after cardiosphere-derived cell therapy in senescent rats. Eur. Heart J. 2017, 38, 2957–2967. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.R.; Barile, L.; Cho, H.C.; Leppo, M.K.; Hare, J.M.; Messina, E.; Giacomello, A.; Abraham, M.R.; Marbán, E. Regenerative potential of cardiosphere-derived cells expanded from percutaneous endomyocardial biopsy specimens. Circulation 2007, 115, 896–908. [Google Scholar] [CrossRef] [PubMed]
- Claycomb, W.C.; Lanson, N.A.; Stallworth, B.S.; Egeland, D.B.; Delcarpio, J.B.; Bahinski, A.; Izzo, N.J. HL-1 cells: A cardiac muscle cell line that contracts and retains phenotypic characteristics of the adult cardiomyocyte. Proc. Natl. Acad. Sci. USA 1998, 95, 2979–2984. [Google Scholar] [CrossRef] [PubMed]
- Climent, A.M.; Guillem, M.S.; Fuentes, L.; Lee, P.; Bollensdorff, C.; Fernández-Santos, M.E.; Suárez-Sancho, S.; Sanz-Ruiz, R.; Sánchez, P.L.; Atienza, F.; et al. Role of atrial tissue remodeling on rotor dynamics: An in vitro study. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1964–H1973. [Google Scholar] [CrossRef]
- Del-Canto, I.; Gómez-Cid, L.; Hernández Romero, I.; Guillem, M.S.; Fernández-Santos, E.; Atienza, F.; Such, L.; Fernández-Avilés, F.; Chorro, F.J.; Climent, A.M. Ranolazine-mediated Attenuation of Mechanoelectric Feedback in Atrial Myocyte Monolayers. Front. Physiol. 2020, 11, 922. [Google Scholar]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef]
- Van Gorp, P.R.R.; Trines, S.A.; Pijnappels, D.A.; de Vries, A.A.F. Multicellular In vitro Models of Cardiac Arrhythmias: Focus on Atrial Fibrillation. Front. Cardiovasc. Med. 2020, 7. [Google Scholar] [CrossRef]
- Brundel, B.J.J.M.; Kampinga, H.H.; Henning, R.H. Calpain inhibition prevents pacing-induced cellular remodeling in a HL-1 myocyte model for atrial fibrillation. Cardiovasc. Res. 2004, 62, 521–528. [Google Scholar] [CrossRef]
- Hong, J.; Choi, J.H.; Kim, T.Y.; Lee, K.-J. Spiral reentry waves in confluent layer of HL-1 cardiomyocyte cell lines. Biochem. Biophys. Res. Commun. 2008, 377, 1269–1273. [Google Scholar] [CrossRef]
- Mace, L.C.; Yermalitskaya, L.V.; Yi, Y.; Yang, Z.; Murray, K.T. Transcriptional Remodeling of Rapidly Stimulated HL-1 Atrial Myocytes Exhibits Concordance with Human Atrial Fibrillation. J. Mol. Cell Cardiol. 2009, 47, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-T.; Chiang, F.-T.; Chen, W.-P.; Hwang, J.-J.; Tseng, C.-D.; Wu, C.-K.; Yu, C.-C.; Wang, Y.-C.; Lai, L.-P.; Lin, J.-L. Angiotensin II induces complex fractionated electrogram in a cultured atrial myocyte monolayer mediated by calcium and sodium-calcium exchanger. Cell Calcium 2011, 49, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-T.; Chiang, F.-T.; Tseng, C.-D.; Yu, C.-C.; Wang, Y.-C.; Lai, L.-P.; Hwang, J.-J.; Lin, J.-L. Mechanical stretch of atrial myocyte monolayer decreases sarcoplasmic reticulum calcium adenosine triphosphatase expression and increases susceptibility to repolarization alternans. J. Am. Coll. Cardiol. 2011, 58, 2106–2115. [Google Scholar] [CrossRef] [PubMed]
- Dias, P.; Desplantez, T.; El-Harasis, M.A.; Chowdhury, R.A.; Ullrich, N.D.; Cabestrero de Diego, A.; Peters, N.S.; Severs, N.J.; MacLeod, K.T.; Dupont, E. Characterisation of connexin expression and electrophysiological properties in stable clones of the HL-1 myocyte cell line. PLoS ONE 2014, 9, e90266. [Google Scholar] [CrossRef]
- Houston, C.; Tzortzis, K.N.; Roney, C.; Saglietto, A.; Pitcher, D.S.; Cantwell, C.D.; Chowdhury, R.A.; Ng, F.S.; Peters, N.S.; Dupont, E. Characterisation of re-entrant circuit (or rotational activity) in vitro using the HL1-6 myocyte cell line. J. Mol. Cell. Cardiol. 2018, 119, 155–164. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Cid, L.; Moro-López, M.; de la Nava, A.S.; Hernández-Romero, I.; Fernández, A.I.; Suárez-Sancho, S.; Atienza, F.; Grigorian-Shamagian, L.; Fernández-Avilés, F. Electrophysiological Effects of Extracellular Vesicles Secreted by Cardiosphere-Derived Cells: Unraveling the Antiarrhythmic Properties of Cell Therapies. Processes 2020, 8, 924. https://doi.org/10.3390/pr8080924
Gómez-Cid L, Moro-López M, de la Nava AS, Hernández-Romero I, Fernández AI, Suárez-Sancho S, Atienza F, Grigorian-Shamagian L, Fernández-Avilés F. Electrophysiological Effects of Extracellular Vesicles Secreted by Cardiosphere-Derived Cells: Unraveling the Antiarrhythmic Properties of Cell Therapies. Processes. 2020; 8(8):924. https://doi.org/10.3390/pr8080924
Chicago/Turabian StyleGómez-Cid, Lidia, Marina Moro-López, Ana S. de la Nava, Ismael Hernández-Romero, Ana I. Fernández, Susana Suárez-Sancho, Felipe Atienza, Lilian Grigorian-Shamagian, and Francisco Fernández-Avilés. 2020. "Electrophysiological Effects of Extracellular Vesicles Secreted by Cardiosphere-Derived Cells: Unraveling the Antiarrhythmic Properties of Cell Therapies" Processes 8, no. 8: 924. https://doi.org/10.3390/pr8080924
APA StyleGómez-Cid, L., Moro-López, M., de la Nava, A. S., Hernández-Romero, I., Fernández, A. I., Suárez-Sancho, S., Atienza, F., Grigorian-Shamagian, L., & Fernández-Avilés, F. (2020). Electrophysiological Effects of Extracellular Vesicles Secreted by Cardiosphere-Derived Cells: Unraveling the Antiarrhythmic Properties of Cell Therapies. Processes, 8(8), 924. https://doi.org/10.3390/pr8080924