Abstract
The study investigated the effect of adding citrus fruits on the dynamics of reducing sugars during the fermentation of Criollo cocoa beans from Lagunas and Casual (Amazonas, Peru). Both spontaneous fermentations and fermentations supplemented with orange, passion fruit, or pineapple at concentrations of 5% and 10% were conducted over six days in wooden boxes, with samples collected every 24 h. Sucrose, glucose, and fructose contents were quantified using ultra-high-performance liquid chromatography coupled with a refractive index detector (UHPLC-RID), and the data were analyzed via ANOVA and PCA. In samples from Lagunas, initial sucrose levels (69.22 mg/g) decreased sharply during the first two days, whereas in the fermented mass from Casual, the sucrose concentrations were low from the outset (6.3 mg/g). The addition of citrus fruits promoted higher accumulation of glucose and fructose in the intermediate and final stages, reaching maximum concentrations of 76.24 mg/g and 81.06 mg/g, respectively, on day six in Lagunas with 10% fruit supplementation. Multivariate analysis indicated that fruit-treated fermentations exhibited a more active and distinct fermentation profile compared to spontaneous fermentations. These results demonstrate that the controlled addition of citrus fruits represents an innovative, low-cost biotechnological strategy for optimizing cocoa fermentation and may enhance the development of flavor and aroma precursors.
1. Introduction
Criollo cocoa beans (Theobroma cacao L.) belong to a premium cocoa variety, recognized worldwide for their sensory qualities and suitability for organic production [1,2,3]. Although this variety accounts for only 5% of total global cocoa production, its distinct characteristics compared to “common” cocoa beans make it a key ingredient in the manufacture of high-quality chocolate products [4]. The flavor and aroma that characterize this variety are primarily determined by genetic factors. In regions of northeastern Peru, these factors are associated with notable sensory attributes and a significant diversity of native fine-aroma cocoa ecotypes [5]. However, freshly harvested beans do not yet exhibit the characteristic chocolate aroma; the precursors of flavor and aroma are developed during post-harvest processing [6]. The combination of genetic potential and post-harvest fermentation and drying processes shapes the chemical composition of the beans, ultimately determining their organoleptic profiles [1,2].
Fermentation, a critical process for the development of aromatic compounds and the final sensory quality of cocoa, is biologically complex, involving a succession of microorganisms (yeasts → lactic acid bacteria [LAB] → acetic acid bacteria [AAB]) that degrade the sugar-rich mucilage and convert it into ethanol and organic acids, thereby increasing temperature and acidifying the beans [7,8]. The diffusion of these acids throughout the bean mass, together with heat, kills the seed embryo and breaks down cell structures, releasing polyphenols and proteins, which are enzymatically degraded into aromatic compounds, reducing bitterness and astringency [6,9].
The mucilaginous pulp surrounding the cocoa beans, rich in sugars such as sucrose, glucose, and fructose, as well as organic acids (citric acid, pectin, hemicellulose), provides an ideal substrate [10] for the initial fermentation phase, dominated by yeasts of the genera Saccharomyces, Pichia, and Hanseniaspora. During the first few days, these yeasts partially hydrolyze sucrose and release glucose and fructose for metabolism, producing ethanol and carbon dioxide [2,11]. This metabolism generates energy for the yeasts and establishes anaerobic conditions that favor the subsequent growth of lactic acid bacteria, including Lactobacillus, Pediococcus, and Leuconostoc, which consume glucose and other sugars to produce lactic acid. In the later stages, acetic acid bacteria such as Acetobacter and Gluconobacter metabolize the ethanol produced by yeasts into acetic acid [2,12,13]. Simultaneously, the complex sugars in the mucilage undergo acid and enzymatic hydrolysis, temporarily increasing the concentrations of the simple sugars glucose and fructose [14]. However, as fermentation progresses, these reducing sugars are continuously consumed as an energy source and further degraded by microbial enzymes, releasing mono-, di-, and oligosaccharide units [9]. This dynamic of sugar release and consumption drives internal biochemical reactions that transform the composition of the beans, including proteolysis [15] and phenolic compound oxidation [16], ultimately defining the aromatic and nutritional profile.
Fermentation of Criollo cocoa beans occurs in two main stages and is completed within approximately 6–7 days [2,15]. During the first three days, the beans develop acceptable characteristics, while in the subsequent days, the precursors of aroma and the physical properties of the beans are fully established. Throughout the process, a general trend of progressive reduction in free sugars is observed, reflecting the metabolic activity within the beans. Residual sugar concentrations serve as an indicator of fermentation progress [17,18]. In controlled fermentations, after 72 h of complete anaerobic conditions, glucose can be reduced by more than 95% and fructose by approximately 94–96% relative to initial values, leaving residual sugar levels of about 2–5 mg/g at the end of the process [19]. This behavior is similar to the fermentation of other sugar-rich fruits, such as Arabica coffee, where approximately 72% of sucrose, 96% of glucose, and 95% of fructose are consumed after 96 h of fermentation [20]. Microbial metabolites promote continuous polysaccharide hydrolysis, accelerating both the release and subsequent consumption of simple sugars [11]. Sugar consumption is reflected in the decline of soluble solids (°Brix) in the mucilage throughout fermentation, indicating progress of the process [21]. Moreover, the development of the characteristic brown coloration in cocoa beans due to polyphenol oxidation corresponds with near-complete depletion of fermentable sugars, validating its use as an indirect indicator of fermentation degree [6].
Citrus fruits, such as pineapple (Ananas comosus), yellow passion fruit (Passiflora edulis f. flavicarpa), and orange (Citrus sinensis), are tropical fruits cultivated in various regions worldwide. Their taste quality is determined by levels of organic acids and sugars—primarily citric acid, sucrose, glucose, and fructose—which contribute to their characteristic flavors [22,23,24]. The incorporation of citrus fruits, rich in soluble sugars and citric acid, into cocoa fermentation can significantly influence the process dynamics [25] in the substrate availability. During fermentation, microorganisms metabolize carbohydrates, proteins, and phenolic compounds to produce organic acids, alcohols, esters, ketones, and aldehydes, which contribute to the sensory profile [26]. The additional supply of fermentable sugars, organic acids, polyphenols, and aromatic compounds [7,27] provides an extra substrate that stimulates microbial activity, serving as sources of energy and carbon [7,28]. Furthermore, the high concentration of citric acid can lower the pulp pH, favoring the early activity of lactic and acetic acid bacteria and thereby modifying the rate of sugar conversion [6]. In spontaneous fermentations, microbial metabolism relies on the naturally available nutrients, whereas fruit addition increases sugar concentration, influencing the rate of sugar consumption and the proportion of compounds generated that define the final flavor profile [2,26]. These sugars also serve as carbonyl sources for the synthesis of aroma precursors during roasting. Interestingly, although the mechanism is not fully understood, many active compounds, including carbohydrates and volatile molecules, are higher in freshly cut fruit [29], which may temporarily enhance the fermentation process.
Monitoring sugar dynamics is crucial due to their role in the formation of chocolate aroma compounds. During roasting, the residual reducing sugars react with amino acids released during fermentation, generating aldehydes, pyrazines, and other volatile compounds essential for cocoa aroma [12]. Accordingly, the level of fermentable sugars serves as a quantitative biochemical indicator of fermentation progress, particularly under conditions involving the addition of citrus fruits, and reflects the activity of the fermentation process. However, knowledge gaps remain regarding how fruit addition during fermentation alters the availability of fermentable sugars and, consequently, affects the synthesis of aroma and flavor precursors. Previous studies suggest that higher concentrations of simple sugars and organic acids can modulate microbial activity and prolong the release of metabolites associated with aroma development [3,6,26,30]. Therefore, this study aims to evaluate whether the addition of citrus fruits not only modifies sugar dynamics during fermentation but also contributes to enhancing the formation of aromatic and flavor compounds in Criollo cocoa beans.
2. Materials and Methods
2.1. Biological Materials and Reagents
Criollo cocoa pods were collected from the provinces of Bagua and Utcubamba-Cajaruro, Amazonas, Peru (Figure 1). The locations were Casual (5°35′21″ S, 78°33′9″ W; 398 m.s.n.m) and Lagunas (5°45′30.2″ S, 78°18′8.2″ W; 590 m.s.n.m).
Figure 1.
Origins of Criollo cocoa beans in the Amazonas region, Peru.
The citrus fruits selected for this study were Ananas comosus “Piña Gold MD2,” Passiflora edulis f. flavicarpa “Maracuyá amarillo,” and Citrus sinensis (L.) Osbeck “Washington Navel Orange”. These fruits were purchased from local markets and chosen based on their regional abundance and their high reducing sugar and organic acid content, which are relevant for the fermentation process.
Standard sugar solutions, including sucrose, glucose, and fructose, as well as borate buffer and 47 mm nylon filter membranes (pore size 0.20 μm), were obtained from Agilent Technologies (Santa Clara, CA, USA). Other reagents included HPLC-grade petroleum ether and acetonitrile (Sigma Aldrich, St. Louis, MO, USA), 0.45 and 0.22 μm Millex-GP syringe filters (Merck, Darmstadt, Germany), and 1.5 mL glass vials (Wheaton, Millville, NJ, USA).
2.2. Fermentation of Criollo Cocoa Beans and Sampling
The Criollo cocoa beans from both locations were transported to the APROCAM Multiple Services Cooperative, located in the province of Bagua (Car. Bagua-Copallín Km. 4), for post-harvest processing. The samples underwent three experimental fermentation treatments (Figure 2): spontaneous fermentation (SF) and fermentations with the addition of 5% (FF5) and 10% (FF10) citrus fruits, following the procedure described by Castro-Alayo et al. [30] with modifications. All fermentations lasted six days (0–144 h). For each treatment, 35 kg of cocoa beans were placed in wooden boxes and covered with polyethylene sheets to concentrate the temperature, ensuring it did not exceed 50 °C.
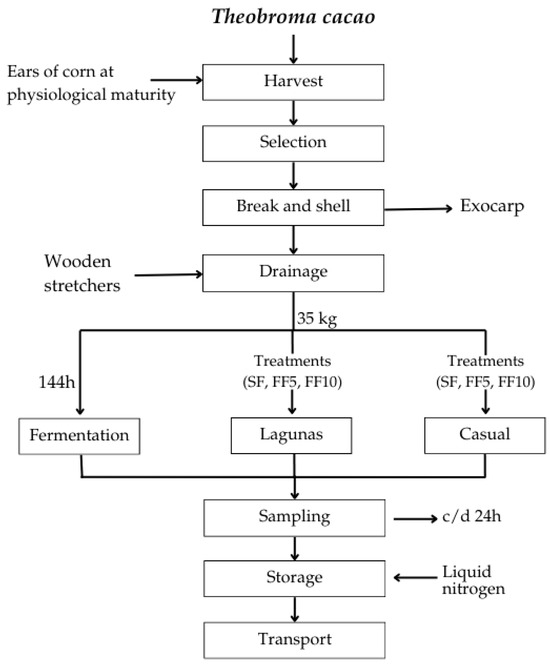
Figure 2.
Flow chart of the experimental fermentation process: spontaneous fermentation (SF); fermentation with 5% citrus fruit addition (FF5); fermentation with 10% citrus fruit addition (FF10).
For the fruit-supplemented treatments, orange, passion fruit, and pineapple (previously sanitized) were cut and added on day two of the fermentation process. Turnings were performed at 48 and 96 h, with an additional turning on the fifth day at 24 h. Fermentations were conducted under ambient temperature and relative humidity conditions typical of the local environment (approximately 31 °C and 56%). At the end of the process (144 h), the beans were removed and allowed to drain, leaving the added fruits in place.
During fermentation, samples were collected every 24 h. Approximately 100 g of beans were aseptically separated from the pulp and placed in sterile 50 mL Falcon tubes for each sampling time (0–144 h). Samples were stored in liquid nitrogen until transfer to the Food Engineering and Post-Harvest Research Laboratory at UNTRM, where they were maintained under ultra-low temperature conditions (Eppendorf, Premium 0410, Hamburg, Germany).
2.3. Sample Treatment
The samples were freeze-dried in 50 mL Falcon tubes at 0.008 bar and −84 °C for 18 h using a freeze dryer (Labconco, model 710.402.010, Kansas City, MO, USA) and subsequently ground to a fine powder, following the procedure described by Balcázar-Zumaeta et al. [15] with modifications. The samples were then defatted by mixing 5 g of powdered material with 90 mL of petroleum ether, shaking for 1 min in a vortex mixer (OHAUS, model PX224/E, Greifensee, Switzerland), and centrifuging at 1000 rpm for 10 min at room temperature in a refrigerated centrifuge (Eppendorf, model 5810R, Hamburg, Germany). The supernatant was discarded, and the defatted samples were stored in a dry environment until further analysis.
2.4. Sugar Analysis
Sugar quantification in the defatted samples was performed following the methods of Alghamdi et al. [31] and Velásquez-Reyes et al. [32] with modifications. Five hundred milligrams of sample were weighed and extracted with 30 mL of ultrapure water, shaken for 5 min, and homogenized in a sonicator (DAIHAN Scientific, WUC-D10H, Wonju, Republic of Korea) at 30 °C for 30 min. The solutions were centrifuged at 10,000 rpm for 10 min at 4 °C. The precipitate was resuspended in 10 mL of water and centrifuged again under the same conditions. The supernatants were combined and centrifuged once more, then filtered sequentially through Whatman No. 40 filter paper, a 0.45 μm nylon filter, and a 0.22 μm membrane filter.
Standards of fructose (2%), glucose (2%), and sucrose (0.5%) were prepared in distilled water, and all solutions were placed in vials for analysis. Sugar separation and quantification were performed using ultra-high-performance liquid chromatography (UHPLC) coupled with a refractive index detector (LC-RID, Agilent technologies, G7162B, Waldbronn, Germany) and OpenLab software version 2.8. Separation was achieved on a Hi-Plex H column (300 × 7.7 mm) maintained at 27 °C, using a mobile phase of distilled water:acetonitrile (10:80 v/v) at a flow rate of 1.5 mL/min with a 10 μL injection volume. Sugars were identified by comparing retention times and chromatograms with those of the standards, and quantified by comparing peak areas with calibration curves generated from the standards: Fructose LOD (0.12) and LOQ (0.38); glucose LOD (0.11) and LOQ (0.35); and sucrose LOD (0.39) and LOQ (1.18).
2.5. Statistical Analysis
The data obtained were subjected to a three-way analysis of variance (ANOVA) to assess the effects of citrus fruit addition and cocoa origin on sugar concentrations, followed by a Tukey multiple comparison test at a 95% confidence level. For the mean values, multivariate analyses were conducted based on the quantified sugars, including cluster analysis (k-means) and principal component analysis (PCA), using RMarkdown software (RStudio, v. 2022.07.2 + 576, Boston, MA, USA).
3. Results and Discussion
3.1. Dynamics of Sucrose, Glucose, and Fructose During the Fermentation of Criollo Cocoa with Citrus Fruit Additions
The dynamics of sugars in cocoa beans begin with the hydrolysis of sucrose, leading to the formation of the reducing sugars glucose and fructose [33]. These transformations occur over approximately six days, corresponding to the period required to achieve an optimal degree of fermentation in Criollo cocoa beans prior to drying. Since reducing sugars (RS) are responsible for the characteristic sensory profile of cocoa and are considered chemical markers of flavor quality [18], variations in sugar content during fermentation have been reported according to cocoa variety and origin [9]. However, it is necessary to investigate these dynamics when additional sugars are introduced, such as those from added citrus fruits (Table 1). Quantification of these sugars is essential to understanding the metabolic behavior of cocoa during fermentation.

Table 1.
Average values of the dynamics of sucrose, glucose, and fructose content (mg/g) in cocoa beans during fermentation from two locations (Casual and Lagunas) under different citrus fruit addition treatments (SF, FF5, and FF10).
Sucrose is the most abundant disaccharide in fresh cocoa mucilage, representing approximately 90% of total sugars in unfermented beans [34,35]. As shown in Table 1, the evolution of sucrose content varied significantly (p < 0.05) depending on the origin of the Criollo cocoa and the fermentation treatments. Sucrose content in cocoa from Casual location, remained relatively stable, showing only minor fluctuations. In contrast, samples from Lagunas exhibited higher sucrose levels after two days of fermentation, with a more pronounced response to treatments with citrus fruit addition, particularly FF10. This increase is attributed to the temporary release of sucrose from the added fruits, which, at higher concentrations, provide readily detectable sugars during the early stages of fermentation [36].
For spontaneous fermentation (SF) at the beginning of the process, samples from Lagunas also displayed a high sucrose concentration of 69.22 mg/g, followed by a rapid decrease within the first 48 h. The initial differences in sucrose content between origins can be attributed to genetic factors, as well as soil and climate conditions affecting carbohydrate accumulation in the mucilage [37,38]. For both origins, sucrose gradually decreased from day 2 until the end of fermentation, reaching levels below 10 mg/g, indicating near-complete depletion. This trend was observed in SF as well as in FF5 and FF10 treatments, where the decrease began on day 3, one day after fruit addition, reflecting the contribution of the supplemental sugars. Similar patterns have been reported by Vizcaino-Almeida et al. [39], in which sucrose concentrations in almonds fermented with fruit pulp declined substantially to 6.54 g/kg. This behavior aligns with the typical stabilization of sucrose during the first 48 h, driven by the activity of anaerobic yeasts and bacteria, which reach maximum populations of 107–108 CFU/g before declining [40]. Key yeast species, including Saccharomyces cerevisiae and Pichia kudriavzevii, hydrolyze sucrose via invertases, which are activated by increased temperature and the diffusion of organic acids into the cocoa bean [11,13,41]. Additionally, sucrose release and consumption rates may vary depending on the composition of the microbial community.
In Table 1, the evolution of glucose content showed significant variations (p < 0.05). For the samples from Lagunas origin, glucose in the SF treatment was initially high on day 0, measuring 69.39 mg/g, but decreased sharply to 18.76 mg/g within 24 h. In contrast, for treatments with citrus fruit addition, a progressive increase was observed starting on day 2, with FF10 reaching a maximum of 76.24 mg/g on day 6—more than a fourfold increase compared to the initial value. Glucose levels remained relatively stable throughout fermentation (Casual location), with only slight increases on day 2 for SF, and no upward trends were observed for any of the fruit-supplemented treatments. Final glucose concentrations ranged from 10 to 30 mg/g, significantly lower than those observed in Lagunas samples. In both locations, glucose levels stabilized from day 3 onward. From a kinetic perspective, glucose concentrations close to or above 200 mg/g can slightly inhibit the growth of Saccharomyces cerevisiae [42], whereas concentrations between 42.9 and 55.4 mg/g have been reported to favor the growth of yeasts and lactic acid bacteria [43]. Similarly, Viesser et al. [13] reported that glucose remains the main reducing sugar after 72–96 h, supporting the growth of yeasts (S. cerevisiae, P. kudriavzevii) and lactic acid bacteria, which can reach maximum populations of 7.8 log CFU/g in fermentations with fruit pulps Vizcaino-Almeida et al. [39], possibly including Lactobacillus fermentum and L. plantarum [41], promoting ethanol and lactic acid formation [1].
The glucose concentrations observed in this study (30–76 mg/g) are well below inhibitory levels, suggesting that the accumulation in FF10 is primarily influenced by microbial interactions and substrate competition rather than direct inhibition. Kouamé et al. [42] reported that the initial proportion of fermentable substrates significantly affects lactic acid bacteria performance, which can be reduced when glucose is consumed competitively with yeasts. The higher initial glucose availability in the fermented mass from cocoa from Lagunas may have produced a different accumulation behavior compared to the one from Casual location, indicating a potentially more diverse microbiota capable of releasing or metabolizing glucose more efficiently [11]. Additionally, the high content of organic acids in the medium—possibly contributed by citrus fruit compounds such as phenolics and organic acids [44]—may favor increased concentrations of reducing sugars, modifying their consumption pathways, as observed in mixed coffee and apple fermentations [14,26]. This could explain why glucose accumulates toward the end of fermentation rather than being consumed immediately, promoting the formation of aromatic aldehydes (e.g., isovaleraldehyde, phenylacetaldehyde) associated with glucose metabolism [22,45].
Fructose content (mg/g) in the evaluated Criollo cocoa samples varied significantly (p < 0.05) according to both origin and treatment (Table 1). In Lagunas samples, fructose increased from day 1 in all treatments, with FF10 reaching the highest concentration of 81.06 mg/g on day 6. SF and FF5 also exhibited significant increases, with slight decreases observed in the final days of fermentation. Observed values ranged from 49.67 to 67.25 mg/g, reflecting the initial proportions of glucose and fructose and the competitive interactions between yeasts and lactic acid bacteria for the substrate [43]. This accumulation may result from a microbial preference for glucose [46,47], the higher contribution of natural fructose from citrus fruits, and partial inhibition of metabolism under acidic conditions [42,47]. In samples from Casual location, a different pattern was observed. Treatments started with similar fructose levels from day 2 onwards, gradually diverging. SF and FF5 remained relatively stable at approximately 38.68 mg/g, while FF10 showed a continuous increase until day 5. Although the addition of citrus fruits led to increases, it did not significantly affect fructose accumulation in this origin, and concentrations converged by the end of fermentation. These results indicate that fructose does not conform to simple kinetic models for reaction order determination [48] and remains available until late fermentation stages, being largely consumed after 120 h [13]. Fructose accumulation may also be influenced by the high citric acid content of the added fruits, which can partially inhibit the activity of lactic acid bacteria, such as Limosilactobacillus fermentum and Lactiplantibacillus plantarum, as well as acetic acid bacteria, including Acetobacter pasteurianus, reducing the rate of glucose and fructose consumption [44,47,49,50]. Accumulation of acetic acid may also contribute to this effect [1]. Similarly, Vizcaino-Almeida et al. [39] reported that while citric acid decreases during fermentation, lactic and acetic acid concentrations increase.
In all treatments, fructose concentrations were higher than glucose, consistent with previous studies [47,51]. As a ketohexose, fructose is more reactive than glucose in the Maillard reaction, playing a key role in the formation of pyrazines and furfurals during roasting [52,53]. Plant matrix studies indicate that fructose generates higher levels of 2-methylpyrazine, 2,5-dimethylpyrazine, and furfural compared to glucose, supporting its contribution to the desirable sweet, caramelized, and roasted notes in Criollo cocoa [53,54]. Additionally, some cocoa fermentation co-products, such as banana peel, contribute volatile compounds like methyl acetate, along with aldehydes, alcohols, acids, esters, ketones, pyrazines, and terpenes, which enhance fruity, sweet, and ethereal sensory notes [55]. Further studies are warranted to explore these effects in other matrices.
Glucose and fructose represent the main residual reducing sugars during the acetic fermentation phase in FF10 treatments [1,13,18]. From a technological perspective, this phase is considered critical for the synthesis of volatile compounds and the formation of flavor precursors [18,56,57]. These effects may be linked to the natural content of citrus fruits and their influence on the metabolism of various microbial species within the fermentation system [14,57,58]. Although research on Criollo cocoa fermentation has advanced worldwide in recent years [59], the addition of citrus fruits represents an economically viable and accessible biotechnological strategy for small-scale producers. This approach promotes the natural development of microbial communities capable of producing volatile compounds [57], thereby enhancing and differentiating the unique sensory profiles of fine-aroma cocoa beans and their derivatives.
3.2. Quantification of Total Sugars in SF, FF5, and CF10 Criollo Cocoa Beans
The fermentation of Criollo cocoa involves the degradation of sugars present in the pulp, including reducing sugars such as glucose and fructose, which are the main products of sucrose hydrolysis and constitute the primary energy sources utilized by microorganisms during active fermentation [60].
Figure 3a,b shows the evolution of total sugars throughout the fermentation process. In Lagunas samples, SF started with an initial sugar content of 163.55 mg/g, which decreased sharply to 45.89 mg/g on day 1, then increased to reach stable values between 110.65 and 133.78 mg/g until day 5, before declining again to 91.86 mg/g on the final day. Treatments with added fruit also exhibited an initial decrease, followed by a faster recovery, reaching 135.76 mg/g from day 2 and remaining relatively stable. The FF10 treatment exhibited the highest sugar levels during the final days, reaching 164.35 mg/g on day 6. This behavior is associated with the physicochemical environment of the mucilage and its interaction with microbial metabolites, which influence the controlled release of sugars during fermentation Calvo et al. [9]. In contrast, samples from Casual location, the total sugar contents were lower for all treatments, remaining below 70.41 mg/g and displaying greater variability. The SF treatment exhibited a temporary peak on day 2, followed by a gradual decline and stabilization until the final days. Fruit-added treatments showed slight fluctuations, reaching higher values of 71.82 mg/g on day 5. Previous studies have shown that the inclusion of fruit promotes a sustained release of simple sugars due to the fruit’s contribution and the slow degradation of mucilage. Reducing sugar levels may increase during fermentation due to hydrolysis driven by environmental acidification [9,26,39], while in spontaneous fermentation, glucose and fructose levels gradually stabilize over time, supporting the progression of cocoa bean fermentation [11].
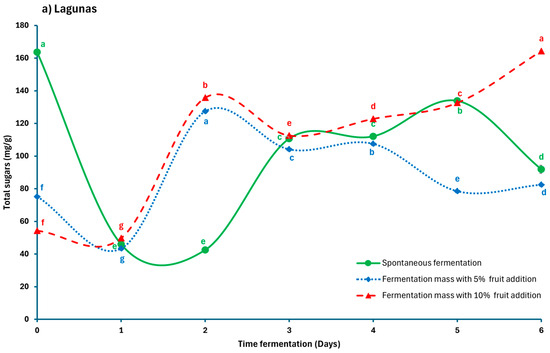
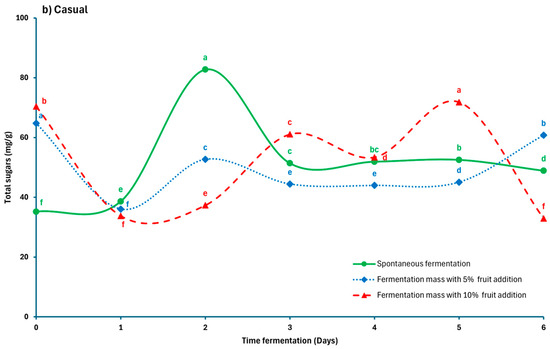
Figure 3.
Total sugar levels during the fermentation of Criollo cocoa beans from two locations: (a) Lagunas; (b) Casual. Different letters in each line (a, b, c, d, e, f, g) indicate significant differences (p < 0.05) between the fermentation times of each process.
3.3. Principal Component Analysis (PCA) in the Formation of Reducing Sugars
Fermentation time was grouped based on the sugar content of the cocoa beans using principal component analysis (PCA), providing a comprehensive overview of the effects of treatment and origin throughout the fermentation process, as reflected in the sugar profile of Criollo cocoa beans (Figure 4 and Figure 5).
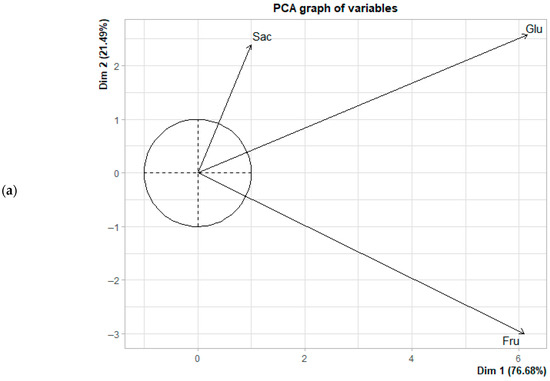
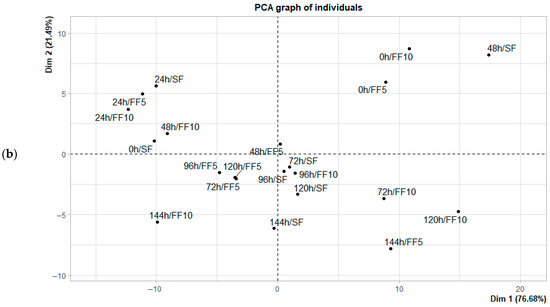
Figure 4.
Principal component analysis (PCA) of the fermentation of Criollo cocoa beans in the Casual origin: (a) Biplot of sugar concentration; (b) Distribution of fermentation times based on each observation in the principal component space.
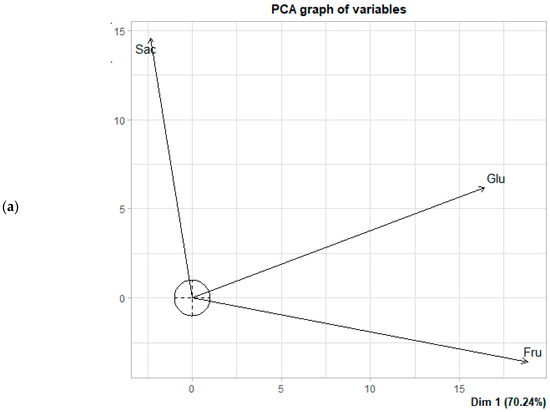
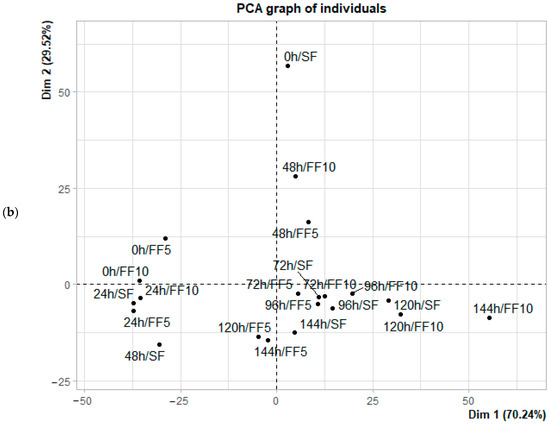
Figure 5.
Principal component analysis (PCA) of the fermentation of Criollo cocoa beans from the Lagunas origin: (a) Biplot of sugar concentration; (b) Distribution of fermentation times based on each observation in the principal component space.
Principal component analysis (PCA) indicated that the first two components (PCA1 + PCA2) explained 98.17% (Casual) of the total variance in the data (Figure 4a,b). PC1 was primarily defined by the opposition between glucose and fructose, whereas PC2 was mainly associated with sucrose. From the Lagunas samples, the first two components accounted for 99.76% of the variance in sugar behavior during fermentation (Figure 5a,b). Within the correlation circle, sucrose and fructose exhibited the highest contributions, showing a strong positive correlation with vectors pointing in similar directions. This suggests a direct relationship between these sugars, potentially reflecting the hydrolysis of sucrose to fructose or parallel evolution of their concentrations under fermentation conditions.
Regarding the distribution of observations over time, initial fermentation stages (0–24 h) were associated with sucrose, intermediate stages (48–72 h) occupied central positions in the component space, and final stages (96–144 h) were primarily associated with fructose in Casual, and with fructose along with residual glucose in Lagunas samples. Additionally, treatments with citrus fruits (FF5 and FF10) exhibited a more defined progression and more consistent groupings within the PCA space compared to spontaneous fermentation (SF), indicating a more controlled and predictable sugar evolution throughout the process.
In the early stages of fermentation, sucrose is the predominant sugar and contributes most to the variance, with differences in the sugar profile between treatments and origins not yet significant (p < 0.05). This pattern aligns with previous studies reporting that enzymatic hydrolysis of sucrose leads to a progressive increase in fructose and glucose during the first few days, with variations in proportions influenced by pH, temperature, and the composition of the active microbiota [11,17,33]. Conversely, glucose is positioned nearly perpendicular to sucrose and fructose in the correlation circle for both origins (Figure 4a and Figure 5a), indicating a very low correlation with these sugars and suggesting independent dynamics. This observation is consistent with the findings of Díaz-Muñoz & De Vuyst [46,47], which indicate that glucose is preferentially consumed during the intermediate stages of fermentation, showing a brief initial increase due to hydrolysis but being rapidly metabolized during subsequent stages [13]. However, the differentiating effect becomes evident during the final days of fermentation, particularly in Criollo cocoa beans from Lagunas. Samples from days 5 and 6 in the FF10 treatment were associated with high concentrations of fructose. This finding suggests that the addition of FF10 to Lagunas cocoa beans promotes greater accumulation or release of fructose toward the end of fermentation. A similar, although less pronounced, pattern was observed in spontaneous fermentation (SF) on days 5 and 6, indicating that Criollo cocoa beans inherently possess the capacity to accumulate fructose during the final stages. Therefore, FF10 appears to enhance or accelerate a process that naturally occurs in these beans.
The accumulation of fructose in the last days of fermentation is a key indicator of successful fermentation, as it coincides with the expected development of volatile compounds during roasting [32]. In the samples from the Casual location, SF during the first two days remained near the origin in PCA space and clustered with the initial days of Lagunas, suggesting that the sugar content of the cocoa beans from Casual is lower, which leads to a less marked dynamic over time for all treatments. Overall, both the origin of the beans and the addition of citrus fruits significantly influence the dynamics of fermentable sugars. These reducing sugars, together with peptides and amino acids, undergo the Maillard reaction, contributing to the formation of characteristic chocolate flavor precursors [10,12], with optimal concentrations of reducing sugars occurring approximately concurrently with peak amino acid levels [15,18]. PCA results indicate that glucose and fructose dominate at the end of fermentation, consistent with Vizcaino-Almeida et al. [39], who reported that the addition of fruit increases these reducing sugars, reducing cotyledon acidification and promoting the activity of endogenous enzymes involved in the production of aroma and flavor precursors. Nevertheless, further research is needed, as the type of co-product used during fermentation may influence the resulting volatile profiles [55].
4. Conclusions
Our study allowed us to explore the variation of reducing sugars during the fermentation of Criollo cocoa beans and demonstrated that the addition of citrus fruits significantly modifies fermentation dynamics depending on the origin of the beans. Three distinct phases were observed: an initial stage (0–2 days) characterized by a rapid decrease in sucrose; an intermediate stage (3–5 days) marked by a progressive increase in glucose and fructose influenced by the fruit addition; and a final stage (day 6) dominated by fructose as the main residual sugar. In the samples from Lagunas, the 10% citrus fruit addition (FF10) enhanced these changes, reaching maximum concentrations of glucose (76.24 mg/g) and fructose (81.06 mg/g), indicating a more active fermentation with higher potential for the formation of aromatic precursors.
These results confirm that the controlled addition of citrus fruits can modulate microbial metabolism, differentiating the fermentation profile as a viable biotechnological strategy. Future studies should aim to correlate changes in glucose and fructose dynamics with the actual formation of volatile compounds during roasting and to confirm the impact on final chocolate quality through sensory evaluations. Furthermore, the absence of microbiological data in this study limits the ability to determine which microbial groups dominate each treatment and how fruit addition influences microbial succession. Therefore, the application of metagenomic techniques could clarify the microbial diversity involved in fruit-assisted fermentations and its role in modulating sugar metabolism. Such studies will enable the design of targeted fermentations, optimization of fruit additions, and the establishment of protocols to enhance the expression of superior sensory attributes in Criollo cocoa.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/pr13092834/s1, Table S1: Average values of Sucrose content of cocoa beans during fermentation; Table S2: Average values of Glucose content of cocoa beans during fermentation; and Table S3: Average values of Fructose content of cocoa beans during fermentation.
Author Contributions
Conceptualization: M.L.T.-O., D.M.-M. and E.M.C.-A.; methodology, M.L.T.-O., E.J.C.-D. and A.J.P.-M.; software, A.J.P.-M., D.M.-M. and C.R.B.-Z.; validation, H.A.P.-R., A.J.P.-M. and E.M.C.-A.; formal analysis, M.L.T.-O., E.J.C.-D. and C.R.B.-Z.; investigation, M.L.T.-O., E.J.C.-D., D.M.-M. and E.M.C.-A.; resources, H.A.P.-R., D.M.-M. and C.R.B.-Z.; data curation, M.L.T.-O. and A.J.P.-M.; writing—original draft preparation, M.L.T.-O., E.J.C.-D., D.M.-M. and E.M.C.-A.; writing—review and editing, M.L.T.-O., D.M.-M. and C.R.B.-Z.; visualization, E.J.C.-D. and D.M.-M.; supervision, A.J.P.-M., D.M.-M., E.M.C.-A. and C.R.B.-Z.; project administration, H.A.P.-R., D.M.-M. and C.R.B.-Z.; funding acquisition, D.M.-M. and E.M.C.-A. All authors have read and agreed to the published version of the manuscript.
Funding
The APC was funded by Vicerrectorado de Investigación—Universidad Nacional Toribio Rodríguez de Mendoza de Amazonas.
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors thank the Laboratorio de la Investigación en Ingeniería de los Alimentos y Poscosecha of the Instituto de Investigación, Innovación y Desarrollo para el Sector Agrario y Agroindustrial (IIDAA), and the Project: “ÓMICAS en cultivos andinos bajo situaciones de estrés abiótico en un contexto del cambio climático en la región Amazonas”, contract N°PE501086005-2023 by Consejo Nacional de Ciencia, Tecnología e Innovación Tecnológica-Concytec, for the facilities provided during the execution of this work.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Becerra, L.D.; Zuluaga, M.; Mayorga, E.Y.; Moreno, F.L.; Ruíz, R.Y.; Escobar, S. Cocoa Seed Transformation under Controlled Process Conditions: Modelling of the Mass Transfer of Organic Acids and Reducing Sugar Formation Analysis. Food Bioprod. Process. 2022, 136, 211–225. [Google Scholar] [CrossRef]
- Castro, E.M.; Idrogo, G.; Siche, R.; Cardenas, F.P. Formation of Aromatic Compounds Precursors during Fermentation of Criollo and Forastero Cocoa. Heliyon 2019, 5, e01157. [Google Scholar] [CrossRef]
- Huamán, A.F.; Torres, M.; Aime, M.C.; Leiva, S.T.; Oliva, S.M.; Díaz, J.R. First Report of Thread Blight Caused by Marasmius tenuissimus on Cacao (Theobroma cacao) in Peru. Plant Dis. 2023, 107, 219. [Google Scholar] [CrossRef]
- Sánchez, V.; Zambrano, J.; Iglesias, C. La Cadena de Valor Del Cacao En América Latina y El Caribe; INIAP: Quito, Ecuador, 2019; ISBN 978-9942-36-465-4. Available online: https://repositorio.iniap.gob.ec/handle/41000/5382 (accessed on 27 August 2025).
- Oliva-Cruz, M.; Goñas, M.; Bobadilla, L.G.; Rubio, K.B.; Escobedo-Ocampo, P.; García Rosero, L.M.; Rojas Briceño, N.B.; Maicelo-Quintana, J.L. Genetic Groups of Fine-Aroma Native Cacao Based on Morphological and Sensory Descriptors in Northeast Peru. Front. Plant Sci. 2022, 13, 896332. [Google Scholar] [CrossRef]
- Ruiz-Santiago, F.L.; Márquez-Rocha, F.J.; García-Alamilla, P.; Carrera-Lanestosa, A.; Ramírez-López, C.; Ocaranza-Sánchez, E.; Jiménez-Rodríguez, D.J. Physicochemical and Biochemical Changes in Cocoa during the Fermentation Step. Fermentation 2024, 10, 405. [Google Scholar] [CrossRef]
- Jagtap, U.B.; Bapat, V.A. Wines from Fruits Other than Grapes: Current Status and Future Prospectus. Food Biosci. 2015, 9, 80–96. [Google Scholar] [CrossRef]
- Subroto, E.; Djali, M.; Indiarto, R.; Lembong, E.; Baiti, N. Microbiological Activity Affects Post-Harvest Quality of Cocoa (Theobroma cacao L.) Beans. Horticulturae 2023, 9, 805. [Google Scholar] [CrossRef]
- Calvo, A.M.; Botina, B.L.; García, M.C.; Cardona, W.A.; Montenegro, A.C.; Criollo, J. Dynamics of Cocoa Fermentation and Its Effect on Quality. Sci. Rep. 2021, 11, 16746. [Google Scholar] [CrossRef]
- Grassia, M.; Salvatori, G.; Roberti, M.; Planeta, D.; Cinquanta, L. Polyphenols, Methylxanthines, Fatty Acids and Minerals in Cocoa Beans and Cocoa Products. Food Meas. 2019, 13, 1721–1728. [Google Scholar] [CrossRef]
- Besançon, L.; Lorn, D.; Kouamé, C.; Grabulos, J.; Lebrun, M.; Fontana, A.; Schorr-Galindo, S.; Boulanger, R.; Strub, C.; Colas de la Noue, A. Influence of Yeast Interactions on the Fermentation Process and Aroma Production in Synthetic Cocoa Pulp vs. Real Mucilage Media. Fermentation 2024, 10, 662. [Google Scholar] [CrossRef]
- Campos, S.d.M.; Martínez-Burgos, W.J.; dos Reis, G.A.; Ocán-Torres, D.Y.; dos Santos Costa, G.; Rosas Vega, F.; Alvarez Badel, B.; Sotelo Coronado, L.; Lima Serra, J.; Soccol, C.R. The Role of Microbial Dynamics, Sensorial Compounds, and Producing Regions in Cocoa Fermentation. Microbiol. Res. 2025, 16, 75. [Google Scholar] [CrossRef]
- Viesser, J.A.; de Melo Pereira, G.V.; de Carvalho Neto, D.P.; Rogez, H.; Góes-Neto, A.; Azevedo, V.; Brenig, B.; Aburjaile, F.; Soccol, C.R. Co-Culturing Fructophilic Lactic Acid Bacteria and Yeast Enhanced Sugar Metabolism and Aroma Formation During Cocoa Beans Fermentation. Int. J. Food Microbiol. 2021, 339, 109015. [Google Scholar] [CrossRef]
- Gutiérrez-Ríos, H.G.; Suárez-Quiroz, M.L.; Hernández-Estrada, Z.J.; Castellanos-Onorio, O.P.; Alonso-Villegas, R.; Rayas-Duarte, P.; Cano-Sarmiento, C.; Figueroa-Hernández, C.Y.; González-Rios, O. Yeasts as Producers of Flavor Precursors During Cocoa Bean Fermentation and Their Relevance as Starter Cultures: A Review. Fermentation 2022, 8, 331. [Google Scholar] [CrossRef]
- Balcázar-Zumaeta, C.R.; Fernández-Romero, E.; Lopes, A.S.; Ferreira, N.R.; Chagas-Júnior, G.C.A.; Yoplac, I.; López-Trigoso, H.A.; Tuesta-Occ, M.L.; Maldonado-Ramirez, I.; Maicelo-Quintana, J.L.; et al. Amino Acid Profile Behavior during the Fermentation of Criollo Cocoa Beans. Food Chem. X 2024, 22, 101486. [Google Scholar] [CrossRef]
- Mayra, O.C.; Manosalvas-Quiroz, L.-A.; Mosquera, N.P.; Samaniego, I. Effect of Fermentation Parameters on the Antioxidant Activity of Ecuadorian Cocoa (Theobroma cacao L.). AIMS Agric. Food 2024, 9, 872–886. [Google Scholar] [CrossRef]
- Guillen-Guerrero, K.M.; de la Rosa-Millan, J. Effects of Fermentation Temperature on the Physicochemical Properties, Bioactive Compounds, and In Vitro Digestive Profile of Cacao (Theobroma cacao) Seeds. Fermentation 2025, 11, 167. [Google Scholar] [CrossRef]
- Rohan, T.A.; Stewart, T. The Precursors of Chocolate Aroma: Production of Reducing Sugars during Fermentation of Cocoa Beans. J. Food Sci. 1967, 32, 399–402. [Google Scholar] [CrossRef]
- Rojas-Rojas, K.; Hernández-Aguirre, C.; Mencía-Guevara, A.; Rojas-Rojas, K.; Hernández-Aguirre, C.; Mencía-Guevara, A. Transformaciones bioquímicas del cacao (Theobroma cacao L.) durante un proceso de fermentación controlada. Agron. Costarric. 2021, 45, 53–65. [Google Scholar] [CrossRef]
- Silva, L.C.F.; Pereira, P.V.R.; Cruz, M.A.D.d.; Costa, G.X.R.; Rocha, R.A.R.; Bertarini, P.L.L.; Amaral, L.R.d.; Gomes, M.S.; Santos, L.D. Enhancing Sensory Quality of Coffee: The Impact of Fermentation Techniques on Coffea arabica cv. Catiguá MG2. Foods 2024, 13, 653. [Google Scholar] [CrossRef]
- Guzmán-Armenteros, T.M.; Ruales, J.; Villacís-Chiriboga, J.; Guerra, L.S. Experimental Prototype of Electromagnetic Emissions for Biotechnological Research: Monitoring Cocoa Bean Fermentation Parameters. Foods 2023, 12, 2539. [Google Scholar] [CrossRef] [PubMed]
- Balcázar-Zumaeta, C.R.; Maicelo-Quintana, J.L.; Salón-Llanos, G.; Barrena, M.; Muñoz-Astecker, L.D.; Cayo-Colca, I.S.; Torrejón-Valqui, L.; Castro-Alayo, E.M. A Novel Technique Using Confocal Raman Spectroscopy Coupled with PLS-DA to Identify the Types of Sugar in Three Tropical Fruits. Appl. Sci. 2024, 14, 8476. [Google Scholar] [CrossRef]
- Hu, L.; Yang, C.; Zhang, L.; Feng, J.; Xi, W. Effect of Light-Emitting Diodes and Ultraviolet Irradiation on the Soluble Sugar, Organic Acid, and Carotenoid Content of Postharvest Sweet Oranges (Citrus sinensis (L.) Osbeck). Molecules 2019, 24, 3440. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wei, X.; Ali, M.M.; Rizwan, H.M.; Li, B.; Li, H.; Jia, K.; Yang, X.; Ma, S.; Li, S.; et al. Changes in the Content of Organic Acids and Expression Analysis of Citric Acid Accumulation-Related Genes during Fruit Development of Yellow (Passiflora edulis f. Flavicarpa) and Purple (Passiflora edulis f. Edulis) Passion Fruits. Int. J. Mol. Sci. 2021, 22, 5765. [Google Scholar] [CrossRef] [PubMed]
- Purbaningrum, K.; Hidayat, C.; Witasari, L.D.; Utami, T. Flavor Precursors and Volatile Compounds Improvement of Unfermented Cocoa Beans by Hydrolysis Using Bromelain. Foods 2023, 12, 820. [Google Scholar] [CrossRef]
- He, Z.; Cao, L.; Zhao, L.; Zhou, Y.; Gong, J.; Zhu, C.; Tan, C. Effects of the Mixed Fermentation of Apples and Yeast on the Sensory Evaluation, Physicochemical Composition and Flavor of Coffee Beans. Food Biosci. 2025, 68, 106337. [Google Scholar] [CrossRef]
- Garrido, J.; Borges, F. Wine and Grape Polyphenols—A Chemical Perspective. Food Res. Int. 2013, 54, 1844–1858. [Google Scholar] [CrossRef]
- Kim, I.J.; Park, S.; Kyoung, H.; Song, M.; Kim, S.R. Microbial Valorization of Fruit Processing Waste: Opportunities, Challenges, and Strategies. Curr. Opin. Food Sci. 2024, 56, 101147. [Google Scholar] [CrossRef]
- Zhou, D.; Liu, Q.; Wu, C.; Li, T.; Tu, K. Characterization of Soluble Sugars, Glycosidically Bound and Free Volatiles in Fresh-Cut Pineapple Stored at Different Temperature. Food Biosci. 2021, 43, 101329. [Google Scholar] [CrossRef]
- Castro-Alayo, E.M.; Torrejón-Valqui, L.; Medina-Mendoza, M.; Cayo-Colca, I.S.; Cárdenas-Toro, F.P. Kinetics Crystallization and Polymorphism of Cocoa Butter throughout the Spontaneous Fermentation Process. Foods 2022, 11, 1769. [Google Scholar] [CrossRef]
- Alghamdi, B.A.; Alshumrani, E.S.; Saeed, M.S.B.; Rawas, G.M.; Alharthi, N.T.; Baeshen, M.N.; Helmi, N.M.; Alam, M.Z.; Suhail, M. Analysis of Sugar Composition and Pesticides Using HPLC and GC–MS Techniques in Honey Samples Collected from Saudi Arabian Markets. Saudi J. Biol. Sci. 2020, 27, 3720–3726. [Google Scholar] [CrossRef] [PubMed]
- Velásquez-Reyes, D.; Rodrí guez-Campos, J.; Avendaño-Arrazate, C.; Gschaedler, A.; Alcázar-Valle, M.; Lugo-Cervantes, E. Forastero and Criollo Cocoa Beans, Differences on the Profile of Volatile and Non-Volatile Compounds in the Process from Fermentation to Liquor. Heliyon 2023, 9, e15129. [Google Scholar] [CrossRef]
- Chagas Junior, G.C.A.; Ferreira, N.R.; Gloria, M.B.A.; Martins, L.H.D.S.; Lopes, A.S. Chemical Implications and Time Reduction of On-Farm Cocoa Fermentation by Saccharomyces cerevisiae and Pichia kudriavzevii. Food Chem. 2021, 338, 127834. [Google Scholar] [CrossRef]
- Afoakwa, E.O. Chocolate Science and Technology, 1st ed.; Wiley: Hoboken, NJ, USA, 2016; ISBN 978-1-118-91378-9. [Google Scholar] [CrossRef]
- Cartas, J.; Alvarenga, N.; Partidário, A.; Lageiro, M.; Roseiro, C.; Gonçalves, H.; Leitão, A.E.; Ribeiro, C.M.; Dias, J. Influence of Geographical Origin in the Physical and Bioactive Parameters of Single Origin Dark Chocolate. Eur. Food Res. Technol. 2024, 250, 2569–2580. [Google Scholar] [CrossRef]
- Peña González, M.A.; Ortiz Urgiles, J.P.; Santander Pérez, F.A.; Lazo Vélez, M.A.; Caroca Cáceres, R.S. Physicochemical Changes during Controlled Laboratory Fermentation of Cocoa (CCN-51) with the Inclusion of Fruits and on-Farm Inoculation. Braz. J. Food Technol. 2023, 26, e2023013. [Google Scholar] [CrossRef]
- Febrianto, N.A.; Wang, S.; Zhu, F. Chemical and Biological Properties of Cocoa Beans Affected by Processing: A Review. Crit. Rev. Food Sci. Nutr. 2022, 62, 8403–8434. [Google Scholar] [CrossRef] [PubMed]
- Megías-Pérez, R.; Grimbs, S.; D’Souza, R.N.; Bernaert, H.; Kuhnert, N. Profiling, Quantification and Classification of Cocoa Beans Based on Chemometric Analysis of Carbohydrates Using Hydrophilic Interaction Liquid Chromatography Coupled to Mass Spectrometry. Food Chem. 2018, 258, 284–294. [Google Scholar] [CrossRef]
- Vizcaino-Almeida, C.R.; Guajardo-Flores, D.; Caroca-Cáceres, R.; Serna-Saldívar, S.O.; Briones-García, M.; Lazo-Vélez, M.A. Non-Conventional Fermentation at Laboratory Scale of Cocoa Beans: Using Probiotic Microorganisms and Substitution of Mucilage by Fruit Pulps. Int. J. Food Sci. Technol. 2022, 57, 4307–4315. [Google Scholar] [CrossRef]
- Ho, V.T.T.; Fleet, G.H.; Zhao, J. Unravelling the Contribution of Lactic Acid Bacteria and Acetic Acid Bacteria to Cocoa Fermentation Using Inoculated Organisms. Int. J. Food Microbiol. 2018, 279, 43–56. [Google Scholar] [CrossRef]
- Moreira, I.M.D.V.; Miguel, M.G.D.C.P.; Duarte, W.F.; Dias, D.R.; Schwan, R.F. Microbial Succession and the Dynamics of Metabolites and Sugars during the Fermentation of Three Different Cocoa (Theobroma cacao L.) Hybrids. Food Res. Int. 2013, 54, 9–17. [Google Scholar] [CrossRef]
- Kouamé, C.; Loiseau, G.; Grabulos, J.; Boulanger, R.; Mestres, C. Development of a Model for the Alcoholic Fermentation of Cocoa Beans by a Saccharomyces Cerevisiae Strain. Int. J. Food Microbiol. 2021, 337, 108917. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Zambrano, M.; Grimbs, S.; Ullrich, M.S.; Hütt, M.-T. A Mathematical Model of Cocoa Bean Fermentation. R. Soc. Open Sci. 2018, 5, 180964. [Google Scholar] [CrossRef]
- Ruiz-Rodríguez, L.G.; Mendoza, L.M.; Van-Nieuwenhove, C.P.; Pescuma, M.; Mozzi, F.B. Fermentación de Jugos y Bebidas a Base de Frutas; Instituto Danone: Barcelona, Spain, 2020; ISBN 978-987-25312-2-5. Available online: https://ri.conicet.gov.ar/handle/11336/120385 (accessed on 27 August 2025).
- Wiercigroch, E.; Szafraniec, E.; Czamara, K.; Pacia, M.Z.; Majzner, K.; Kochan, K.; Kaczor, A.; Baranska, M.; Malek, K. Raman and Infrared Spectroscopy of Carbohydrates: A Review. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2017, 185, 317–335. [Google Scholar] [CrossRef]
- Díaz-Muñoz, C.; De Vuyst, L. Functional Yeast Starter Cultures for Cocoa Fermentation. J. Appl. Microbiol. 2022, 133, 39–66. [Google Scholar] [CrossRef]
- Sari, A.B.T.; Fahrurrozi; Marwati, T.; Djaafar, T.F.; Hatmi, R.U.; Purwaningsih; Wanita, Y.P.; Lisdiyanti, P.; Perwitasari, U.; Juanssilfero, A.B.; et al. Chemical Composition and Sensory Profiles of Fermented Cocoa Beans Obtained from Various Regions of Indonesia. Int. J. Food Sci. 2023, 2023, 5639081. [Google Scholar] [CrossRef]
- Megias-Perez, R.; Moreno-Zambrano, M.; Behrends, B.; Corno, M.; Kuhnert, N. Monitoring the Changes in Low Molecular Weight Carbohydrates in Cocoa Beans During Spontaneous Fermentation: A Chemometric and Kinetic Approach. Food Res. Int. 2020, 128, 108865. [Google Scholar] [CrossRef]
- Ghisolfi, R.; Bandini, F.; Vaccari, F.; Bellotti, G.; Bortolini, C.; Patrone, V.; Puglisi, E.; Morelli, L. Bacterial and Fungal Communities Are Specifically Modulated by the Cocoa Bean Fermentation Method. Foods 2023, 12, 2024. [Google Scholar] [CrossRef]
- Tigrero-Vaca, J.; Maridueña-Zavala, M.G.; Liao, H.-L.; Prado-Lince, M.; Zambrano-Vera, C.S.; Monserrate-Maggi, B.; Cevallos-Cevallos, J.M. Microbial Diversity and Contribution to the Formation of Volatile Compounds during Fine-Flavor Cacao Bean Fermentation. Foods 2022, 11, 915. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-Y.; Chen, K.-F.; Changchien, L.-L.; Chen, K.-C.; Peng, R.Y. Volatile Variation of Theobroma cacao Malvaceae L. Beans Cultivated in Taiwan Affected by Processing via Fermentation and Roasting. Molecules 2022, 27, 3058. [Google Scholar] [CrossRef]
- Fayek, N.M.; Xiao, J.; Farag, M.A. A Multifunctional Study of Naturally Occurring Pyrazines in Biological Systems; Formation Mechanisms, Metabolism, Food Applications and Functional Properties. Crit. Rev. Food Sci. Nutr. 2023, 63, 5322–5338. [Google Scholar] [CrossRef] [PubMed]
- Laemont, J.; Barringer, S. Effect of pH, Reducing Sugars, and Protein on Roasted Sunflower Seed Aroma Volatiles. Foods 2023, 12, 4155. [Google Scholar] [CrossRef] [PubMed]
- Quintana-Fuentes, L.F.; García-Jerez, A.; Rodríguez-Negrette, A.C.; Hoyos-Merlano, N.T.; Alvis-Bermúdez, A. Impact of Spontaneous Fermentation on the Physicochemical and Sensory Qualities of Cacao. Fermentation 2025, 11, 377. [Google Scholar] [CrossRef]
- Tonin, I.P.; Baqueta, M.R.; Hantao, L.W.; Marini, F.; de Souza Silveira, P.T.; Martins, M.O.P.; Efraim, P. Volatiles from Cocoa Bean to Nibs, Liquor, and Chocolate: An Integrated Chemometric Analysis of Fermentation Treatments with Pulp Reduction, Malt Residues, and Banana Peel. Food Chem. 2025, 487, 144774. [Google Scholar] [CrossRef]
- Barišić, V.; Kopjar, M.; Jozinović, A.; Flanjak, I.; Ačkar, Đ.; Miličević, B.; Šubarić, D.; Jokić, S.; Babić, J. The Chemistry behind Chocolate Production. Molecules 2019, 24, 3163. [Google Scholar] [CrossRef] [PubMed]
- Rottiers, H.; Tzompa Sosa, D.A.; De Winne, A.; Ruales, J.; De Clippeleer, J.; De Leersnyder, I.; De Wever, J.; Everaert, H.; Messens, K.; Dewettinck, K. Dynamics of Volatile Compounds and Flavor Precursors During Spontaneous Fermentation of Fine Flavor Trinitario Cocoa Beans. Eur. Food Res. Technol. 2019, 245, 1917–1937. [Google Scholar] [CrossRef]
- Chacón, C.Y.; Mori, P.L.; Chavez, S.G. Antioxidantes y Polifenoles Totales de Chocolate Negro Con Incorporación de Cacao (Theobroma cacao L.) Crudo. Rev. Investig. Altoandin. 2021, 23, 266–273. [Google Scholar] [CrossRef]
- Nascimento, L.L.; Pereira, M.S.; de Almeida, L.S.; da Silveira Ferreira, L.; de Moura Pita, B.L.; de Souza, C.O.; Ribeiro, C.D.F.; Fricks, A.T. Innovation in Cocoa Fermentation: Evidence from Patent Documents and Scientific Articles. Fermentation 2024, 10, 251. [Google Scholar] [CrossRef]
- Santander Muñoz, M.; Rodríguez Cortina, J.; Vaillant, F.E.; Escobar Parra, S. An Overview of the Physical and Biochemical Transformation of Cocoa Seeds to Beans and to Chocolate: Flavor Formation. Crit. Rev. Food Sci. Nutr. 2020, 60, 1593–1613. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).