
Production of Extracellular Lipase by Bacillus halotolerans from Oil-Contaminated Soil in a Pilot-Scale Submerged Bioreactor


 , ,
, ,  , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Bacterial Strain Isolation and Characterization
2.3. Lipase-Producing Bacterial Strain Screening
2.4. Enzyme Production Media
2.5. Optimization of Lipase Producing Media
2.6. Extraction and Partial Purification of Lipase
2.7. Molecular Identification
2.8. Estimate of Protein and Lipase Activity
2.9. Characterization of Enzyme
2.9.1. Study of pH and Stability on Enzyme Activity
2.9.2. Effect of Temperature on Lipase Activity
2.9.3. Effect of Reaction Time on Lipase Activity
2.9.4. Effect of Metal Ions on Lipase Activity
2.9.5. Effect of Substrate Concentration on Lipase Activity
2.10. Application of Lipase Enzyme in Biodiesel Production
3. Results
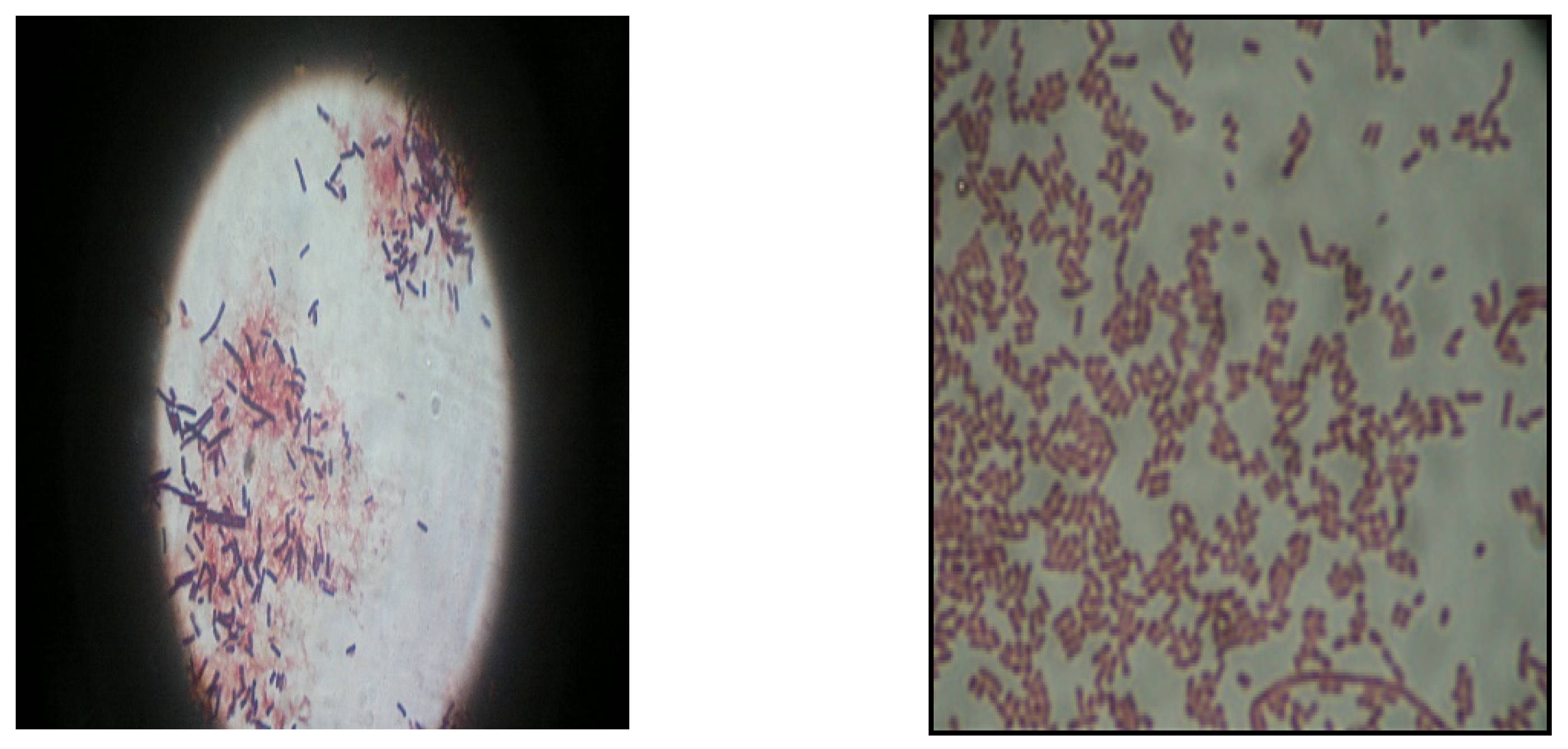
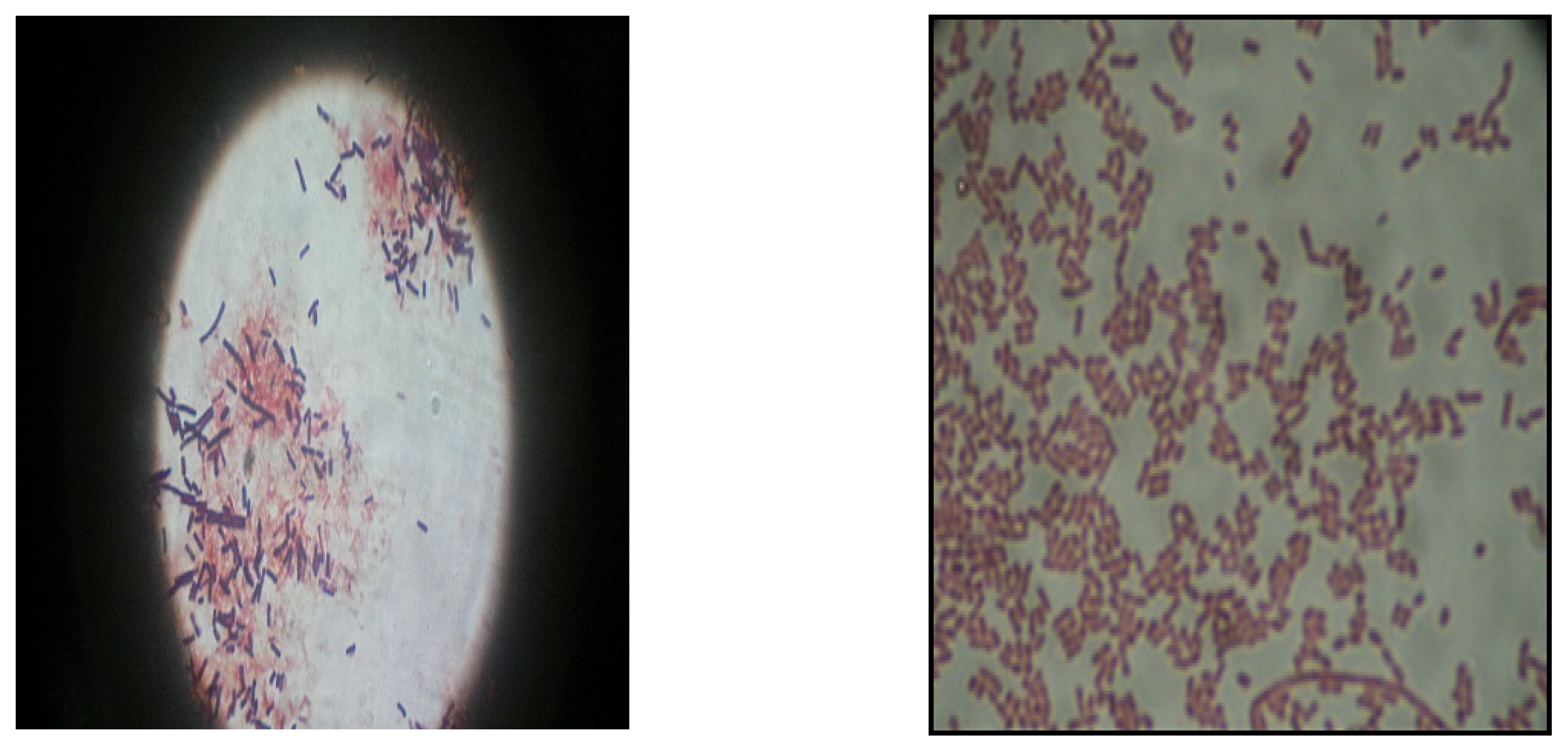
3.1. Bacterial Isolate for Lipase Production
3.2. Phylogenetic Tree Construction by 16S rDNA Gen Sequence
3.3. Determination of Molecular Weight
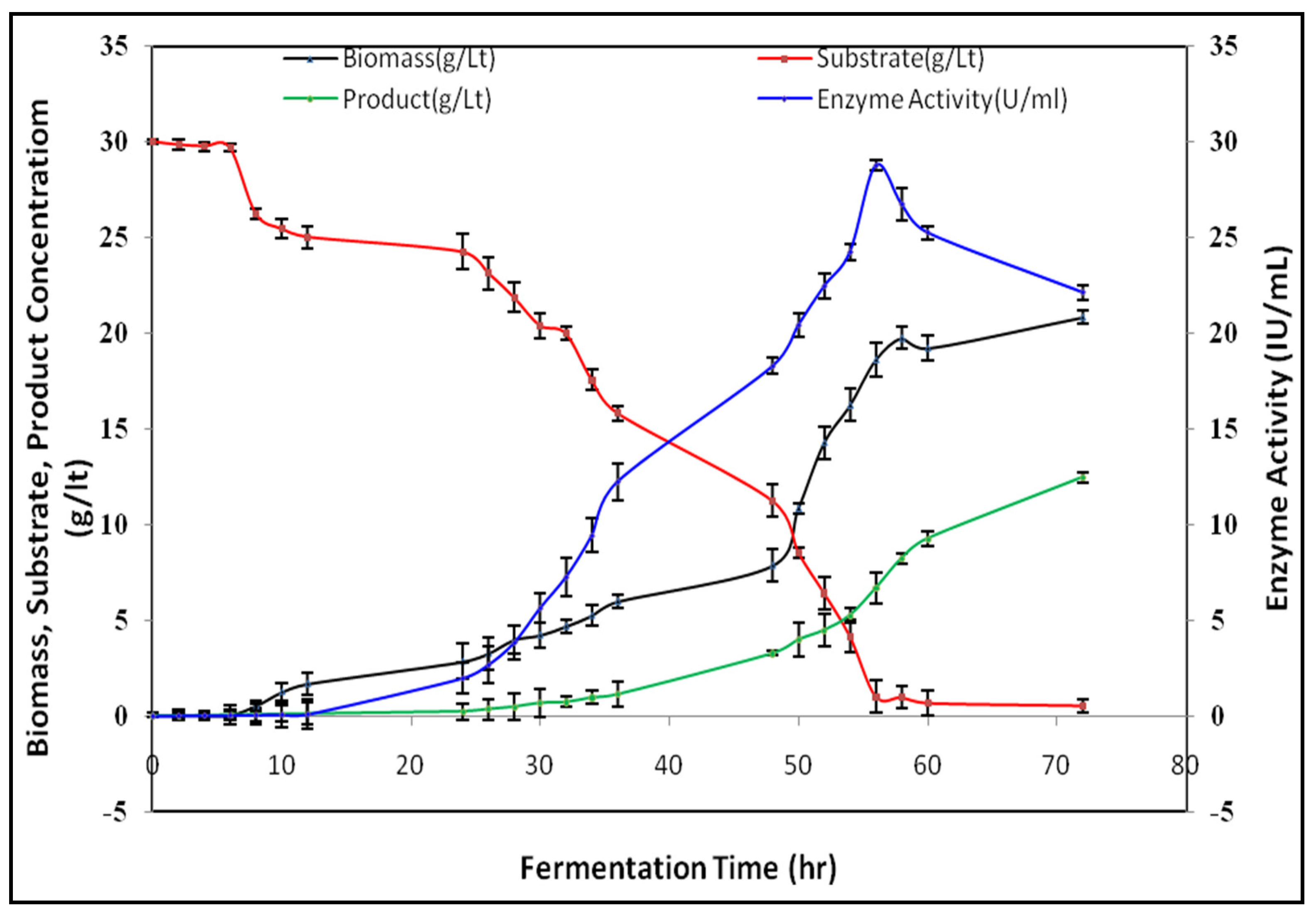
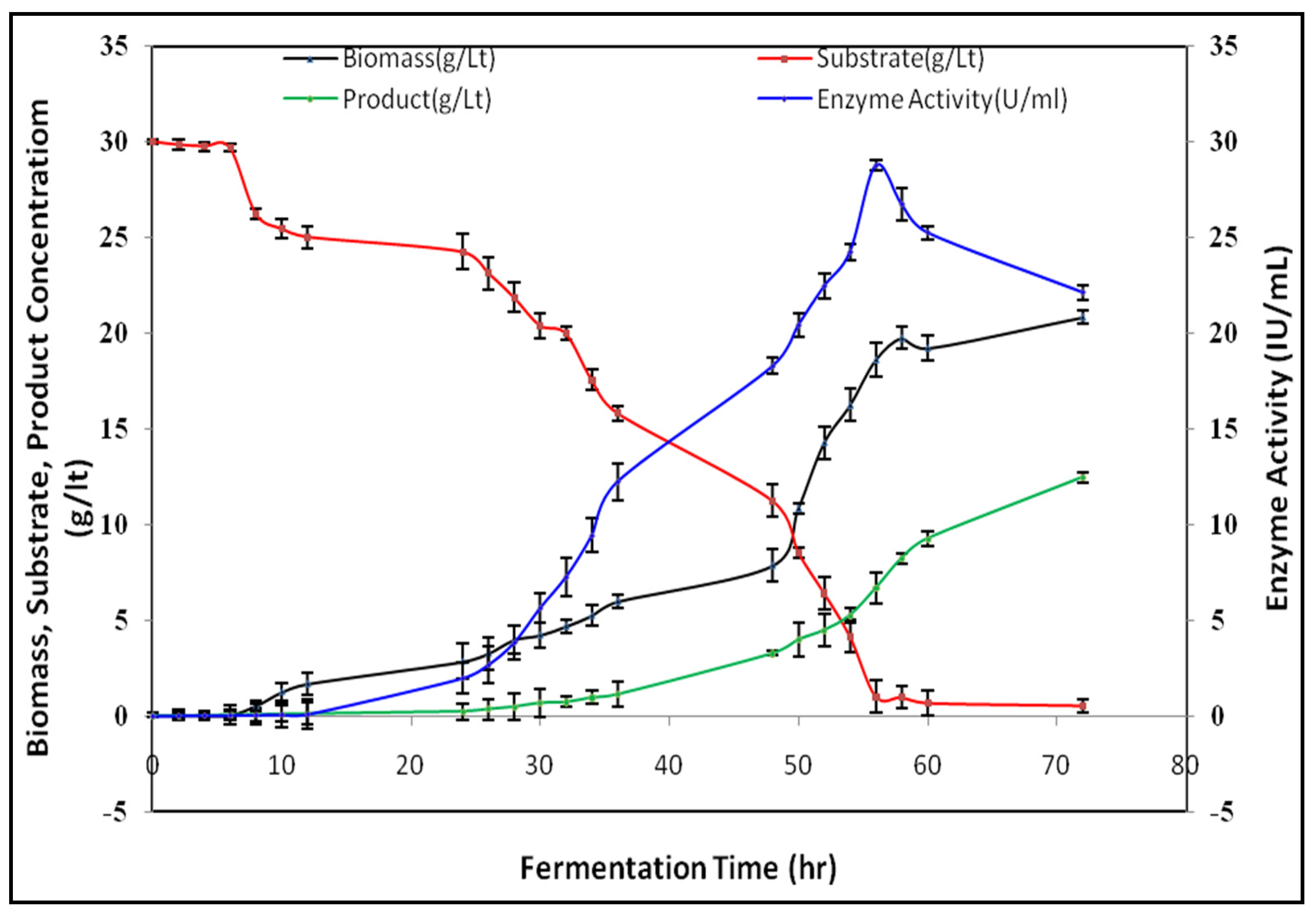
3.4. Batch Submerged Fermentation Studies on Lipase Production
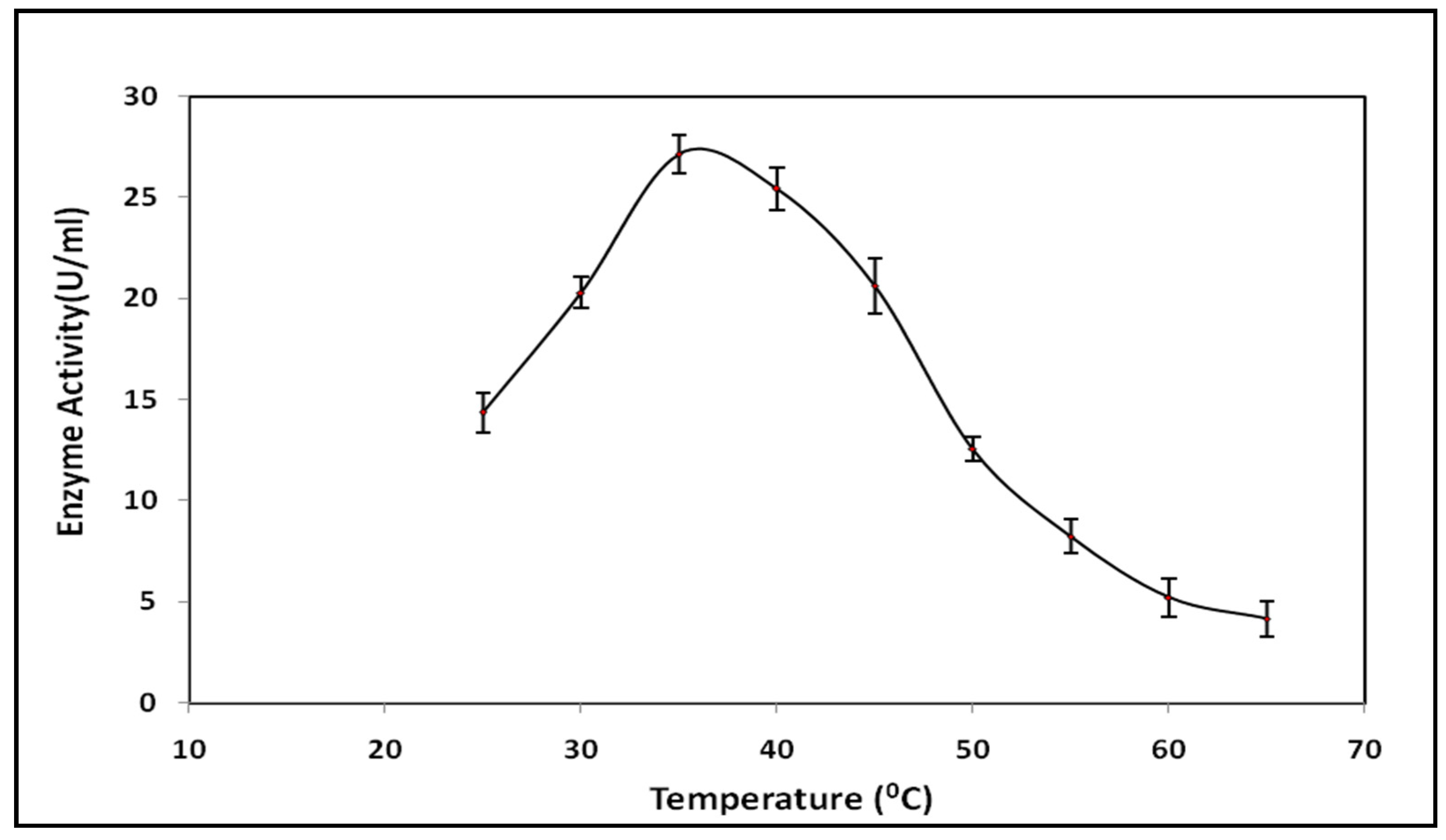
3.5. Optimum Temperature
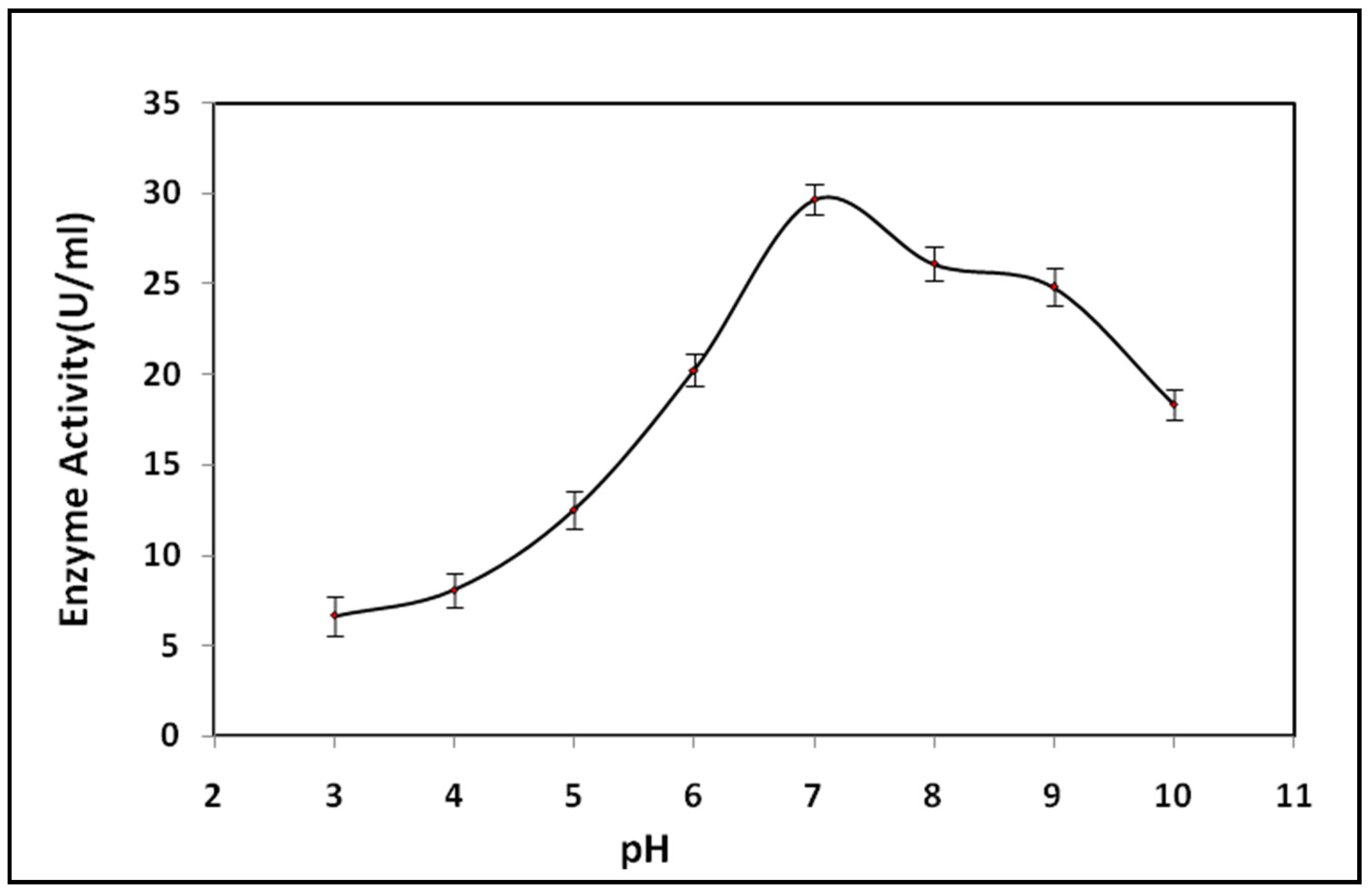
3.6. Optimum pH
3.7. Biodiesel Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghosh, P.K.; Saxena, R.K.; Gupta, R.; Yadav, R.P.; Davidson, S. Microbial Lipases: Production and Applications. Sci. Prog. 1996, 79 Pt 2, 119–157. [Google Scholar] [PubMed]
- Schmidt-Dannert, C.; Rúa, M.L.; Schmid, R.D. Two Novel Lipases from Thermophile Bacillus thermocatenulatus: Screening, Purification, Cloning, Overexpression, and Properties. Methods Enzymol. 1997, 284, 194–220. [Google Scholar] [CrossRef]
- Chandra, P.; Enespa; Singh, R.; Arora, P.K. Microbial Lipases and Their Industrial Applications: A Comprehensive Review. Microb. Cell Fact. 2020, 19, 169. [Google Scholar] [CrossRef]
- de Godoy Daiha, K.; Angeli, R.; de Oliveira, S.D.; Almeida, R.V. Are Lipases Still Important Biocatalysts? A Study of Scientific Publications and Patents for Technological Forecasting. PLoS ONE 2015, 10, e0131624. [Google Scholar] [CrossRef]
- Bajpai, P. Application of Enzymes in the Pulp and Paper Industry. Biotechnol. Prog. 1999, 15, 147–157. [Google Scholar] [CrossRef]
- Pandey, A.; Benjamin, S.; Soccol, C.R.; Nigam, P.; Krieger, N.; Soccol, V.T. The Realm of Microbial Lipases in Biotechnology. Biotechnol. Appl. Biochem. 1999, 29, 119–131. [Google Scholar] [PubMed]
- Ertuğrul, S.; Dönmez, G.; Takaç, S. Isolation of Lipase Producing Bacillus sp. from Olive Mill Wastewater and Improving Its Enzyme Activity. J. Hazard. Mater. 2007, 149, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Bhavani, M.; Chowdary, G.V.; David, M.; Archana, G. Screening, Isolation and Biochemical Characterization of Novel Lipase Producing Bacteria from Soil Samples. Int. J. Biol. Eng. 2012, 2, 18–22. [Google Scholar] [CrossRef]
- Ilesanmi, O.I.; Adekunle, A.E.; Omolaiye, J.A.; Olorode, E.M.; Ogunkanmi, A.L. Isolation, Optimization and Molecular Characterization of Lipase Producing Bacteria from Contaminated Soil. Sci. Afr. 2020, 8, e00279. [Google Scholar] [CrossRef]
- Kiran, G.S.; Shanmughapriya, S.; Jayalakshmi, J.; Selvin, J.; Gandhimathi, R.; Sivaramakrishnan, S.; Arunkumar, M.; Thangavelu, T.; Natarajaseenivasan, K. Optimization of Extracellular Psychrophilic Alkaline Lipase Produced by Marine Pseudomonas sp. (MSI057). Bioprocess Biosyst. Eng. 2008, 31, 483–492. [Google Scholar] [CrossRef]
- Kumar, A.; Singh Parihar, S.; Batra, N.K. Enrichment, Isolation and Optimization of Lipase-Producing Staphylococcus sp. from Oil Mill Waste (Oil Cake). J. Exp. Sci. 2012, 3, 26–30. [Google Scholar]
- Alqahtani, Y.S.; More, S.S.; Shaikh, I.A.; KJ, A.; More, V.S.; Niyonzima, F.N.; Muddapur, U.M.; Khan, A.A. Production and Purification of Pectinase from Bacillus subtilis 15A-B92 and Its Biotechnological Applications. Molecules 2022, 27, 4195. [Google Scholar] [CrossRef]
- Mobarak-Qamsari, E.; Kasra-Kermanshahi, R.; Moosavi-Nejad, Z. Isolation and Identification of a Novel, Lipase-Producing Bacterium, Pseudomnas aeruginosa KM110. Iran. J. Microbiol. 2011, 3, 92–98. [Google Scholar]
- Dutta, S.; Ray, L. Production and Characterization of an Alkaline Thermostable Crude Lipase from an Isolated Strain of Bacillus cereus C(7). Appl. Biochem. Biotechnol. 2009, 159, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Demirkan, E.; Aybey Çetinkaya, A.; Abdou, M. Lipase from New Isolate Bacillus Cereus ATA179: Optimization of Production Conditions, Partial Purification, Characterization and Its Potential in the Detergent Industry. Turk. J. Biol. 2021, 45, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Isiaka Adetunji, A.; Olufolahan Olaniran, A. Optimization of Culture Conditions for Enhanced Lipase Production by an Indigenous Bacillus Aryabhattai SE3-PB Using Response Surface Methodology. Biotechnol. Biotechnol. Equip. 2018, 32, 1514–1526. [Google Scholar] [CrossRef]
- Nawani, N.; Kaur, J. Studies on Lipolytic Isoenzymes from a Thermophilic Bacillus sp.: Production, Purification and Biochemical Characterization. Enzym. Microb. Technol. 2007, 40, 881–887. [Google Scholar] [CrossRef]
- Rahman, R.N.Z.R.A.; Baharum, S.N.; Basri, M.; Salleh, A.B. High-Yield Purification of an Organic Solvent-Tolerant Lipase from Pseudomonas sp. Strain S5. Anal. Biochem. 2005, 341, 267–274. [Google Scholar] [CrossRef]
- Sonkar, K.; Singh, D.P. Biochemical Characterization and Thermodynamic Study of Lipase from Psychrotolerant Pseudomonas punonensis. Biocatal. Agric. Biotechnol. 2020, 28, 101686. [Google Scholar] [CrossRef]
- Sugihara, A.; Tani, T.; Tominaga, Y. Purification and Characterization of a Novel Thermostable Lipase from Bacillus sp. J. Biochem. 1991, 109, 211–216. [Google Scholar]
- Ko, W.H.; Wang, I.T.; Ann, P.J. A Simple Method for Detection of Lipolytic Microorganisms in Soils. Soil Biol. Biochem. 2005, 37, 597–599. [Google Scholar] [CrossRef]
- Bharathi, D.; Rajalakshmi, G.; Komathi, S. Optimization and Production of Lipase Enzyme from Bacterial Strains Isolated from Petrol Spilled Soil. J. King Saud Univ. Sci. 2019, 31, 898–901. [Google Scholar] [CrossRef]
- Peng, R.; Lin, J.; Wei, D. Purification and Characterization of an Organic Solvent-Tolerant Lipase from Pseudomonas Aeruginosa CS-2. Appl. Biochem. Biotechnol. 2010, 162, 733–743. [Google Scholar] [CrossRef]
- Hatzinikolaou, D.G.; Macris, J.B.; Christakopoulos, P.; Kekos, D.; Kolisis, F.N.; Fountoukidis, G. Production and Partial Characterisation of Extracellular Lipase from Aspergillus niger. Biotechnol. Lett. 1996, 18, 547–552. [Google Scholar] [CrossRef]
- Xie, W.; Khosasih, V.; Suwanto, A.; Kim, H.K. Characterization of Lipases from Staphylococcus aureus and Staphylococcus epidermidis Isolated from Human Facial Sebaceous Skin. J. Microbiol. Biotechnol. 2012, 22, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Hasan, F.; Shah, A.A.; Hameed, A. Purification and Characterization of a Mesophilic Lipase from Bacillus subtilis FH5 Stable at High Temperature and PH. Acta Biol. Hung. 2007, 58, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Tran, D.-T.; Yeh, K.-L.; Chen, C.-L.; Chang, J.-S. Enzymatic Transesterification of Microalgal Oil from Chlorella Vulgaris ESP-31 for Biodiesel Synthesis Using Immobilized Burkholderia Lipase. Bioresour. Technol. 2012, 108, 119–127. [Google Scholar] [CrossRef]
- Beller, H.R.; Spormann, A.M.; Sharma, P.K.; Cole, J.R.; Reinhard, M. Isolation and Characterization of a Novel Toluene-Degrading, Sulfate-Reducing Bacterium. Appl. Environ. Microbiol. 1996, 62, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Gupta, N.; Rathi, P. Bacterial Lipases: An Overview of Production, Purification and Biochemical Properties. Appl. Microbiol. Biotechnol. 2004, 64, 763–781. [Google Scholar] [CrossRef]
- Jin, Z.; Zhang, K.; Zhang, L.; Zheng, S.-P.; Han, S.-Y.; Lin, Y. Quantification Analysis of Yeast-Displayed Lipase. Anal. Biochem. 2014, 450, 46–48. [Google Scholar] [CrossRef]
- Hu, J.; Cai, W.; Wang, C.; Du, X.; Lin, J.; Cai, J. Purification and Characterization of Alkaline Lipase Production by Pseudomonas aeruginosa HFE733 and Application for Biodegradation in Food Wastewater Treatment. Biotechnol. Biotechnol. Equip. 2018, 32, 583–590. [Google Scholar] [CrossRef]
- Iftikhar, T.; Niaz, M.; Zia, M.A.; Haq, I.U. Production of Extracellular Lipases by Rhizopus oligosporus in a Stirred Fermentor. Braz. J. Microbiol. 2010, 41, 1124–1132. [Google Scholar] [CrossRef]
- Dharmsthiti, S.; Pratuangdejkul, J.; Theeragool, G.; Luchai, S. Lipase Activity and Gene Cloning of Acinetobacter Calcoacetlcus LP009. J. Gen. Appl. Microbiol. 1998, 44, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Igibah, E.C.; Agashua, L.O.; Sadiq, A.A. Hydro-Geochemical Features and Groundwater Attribute Evaluation in North-Central Abuja, Nigeria. Sci. Afr. 2020, 8, e00324. [Google Scholar] [CrossRef]
- Ghori, M.I.; Iqbal, M.J.; Hameed, A. Characterization of a Novel Lipase from Bacillus sp. Isolated from Tannery Wastes. Braz. J. Microbiol. 2011, 42, 22–29. [Google Scholar] [CrossRef]
- Lee, D.-W.; Koh, Y.-S.; Kim, K.-J.; Kim, B.-C.; Choi, H.-J.; Kim, D.-S.; Suhartono, M.T.; Pyun, Y.-R. Isolation and Characterization of a Thermophilic Lipase from Bacillus thermoleovorans ID-1. FEMS Microbiol. Lett. 1999, 179, 393–400. [Google Scholar] [CrossRef]
- Lee, S.Y.; Rhee, J.S. Production and Partial Purification of a Lipase from Pseudomonas putida 3SK. Enzym. Microb. Technol. 1993, 15, 617–623. [Google Scholar] [CrossRef]
- Tran, D.-T.; Chen, C.-L.; Chang, J.-S. Effect of Solvents and Oil Content on Direct Transesterification of Wet Oil-Bearing Microalgal Biomass of Chlorella vulgaris ESP-31 for Biodiesel Synthesis Using Immobilized Lipase as the Biocatalyst. Bioresour. Technol. 2013, 135, 213–221. [Google Scholar] [CrossRef]
- Yagar, H.; Balkan, U. Entrapment of Laurel Lipase in Chitosan Hydrogel Beads. Artif. Cells Nanomed. Biotechnol. 2017, 45, 864–870. [Google Scholar] [CrossRef]
- Tripathi, R.; Singh, J.; Bharti, R.K.; Thakur, I.S. Isolation, Purification and Characterization of Lipase from Microbacterium sp. and Its Application in Biodiesel Production. Energy Procedia 2014, 54, 518–529. [Google Scholar] [CrossRef]
- Borkar, P.S.; Bodade, R.G.; Rao, S.R.; Khobragade, C.N. Purification and Characterization of Extracellular Lipase from a New Strain: Pseudomonas aeruginosa SRT 9. Braz. J. Microbiol. 2009, 40, 358–366. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain No | Gram Nature | Shape | Mobility | Endo Spore Staining | Catalase Test | Casein Test | Zone of Hydrolysis (Diameter in mm) |
|---|---|---|---|---|---|---|---|
| VSH01 | + | Long rod | − | − | + | − | 10.5 ± 0.1 |
| VSH02 | + | Short rod | + | + | − | − | 7.2 ± 0.9 |
| VSH03 | + | Short rod | + | + | + | − | 12.1 ± 0.1 |
| VSH04 | − | Long rod | + | − | + | + | 12.1 ± 0.5 |
| VSH05 | − | Short rod | + | + | + | _ | 6.2 ± 0.3 |
| VSH06 | + | Streptococcus | − | − | − | − | 6.1 ± 0.6 |
| VSH07 | + | Staphylococcus | − | − | + | − | 10.2 ± 0.3 |
| VSH08 | + | Short rod | + | + | + | − | 18.9 ± 0.6 |
| VSH09 | + | Short rod | + | + | + | + | 22.4 ± 0.1 |
| VSH10 | − | Staphylococcus | − | + | + | − | 14.3 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahnashi, M.H.; Hombalimath, V.S.; Shaikh, I.A.; Muddapur, U.M.; Desai, S.V.; Achappa, S.; El-Sherbiny, M.; Ghoneim, M.M.; Jefri, O.A.; Alshahrani, M.M.; et al. Production of Extracellular Lipase by Bacillus halotolerans from Oil-Contaminated Soil in a Pilot-Scale Submerged Bioreactor. Processes 2022, 10, 1548. https://doi.org/10.3390/pr10081548
Mahnashi MH, Hombalimath VS, Shaikh IA, Muddapur UM, Desai SV, Achappa S, El-Sherbiny M, Ghoneim MM, Jefri OA, Alshahrani MM, et al. Production of Extracellular Lipase by Bacillus halotolerans from Oil-Contaminated Soil in a Pilot-Scale Submerged Bioreactor. Processes. 2022; 10(8):1548. https://doi.org/10.3390/pr10081548
Chicago/Turabian StyleMahnashi, Mater H., Veeranna S. Hombalimath, Ibrahim Ahmed Shaikh, Uday M. Muddapur, Shivalingsarj V. Desai, Sharanappa Achappa, Mohamed El-Sherbiny, Mohammed M. Ghoneim, Ohoud A. Jefri, Mohammed Merae Alshahrani, and et al. 2022. "Production of Extracellular Lipase by Bacillus halotolerans from Oil-Contaminated Soil in a Pilot-Scale Submerged Bioreactor" Processes 10, no. 8: 1548. https://doi.org/10.3390/pr10081548
APA StyleMahnashi, M. H., Hombalimath, V. S., Shaikh, I. A., Muddapur, U. M., Desai, S. V., Achappa, S., El-Sherbiny, M., Ghoneim, M. M., Jefri, O. A., Alshahrani, M. M., Alhujaily, M., Al Ali, A., Mannasaheb, B. A., Al Awadh, A. A., & Khan, A. A. (2022). Production of Extracellular Lipase by Bacillus halotolerans from Oil-Contaminated Soil in a Pilot-Scale Submerged Bioreactor. Processes, 10(8), 1548. https://doi.org/10.3390/pr10081548