
In Vitro Study of the Multimodal Effect of Na+/K+ ATPase Blocker Ouabain on the Tumor Microenvironment and Malignant Cells

 and
and
Abstract
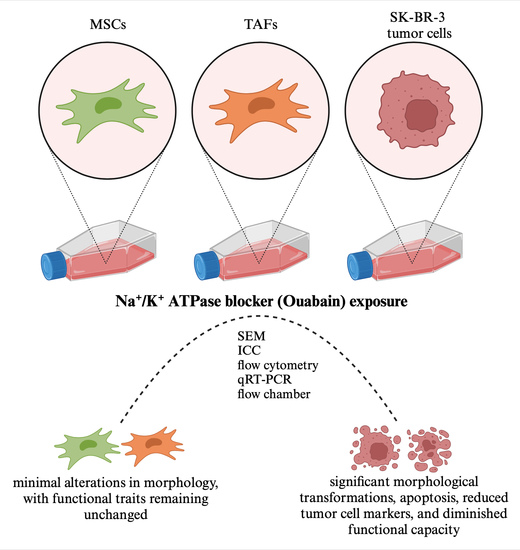
1. Introduction
2. Materials and Methods
2.1. Cell Cultures, Reagents, and Solutions
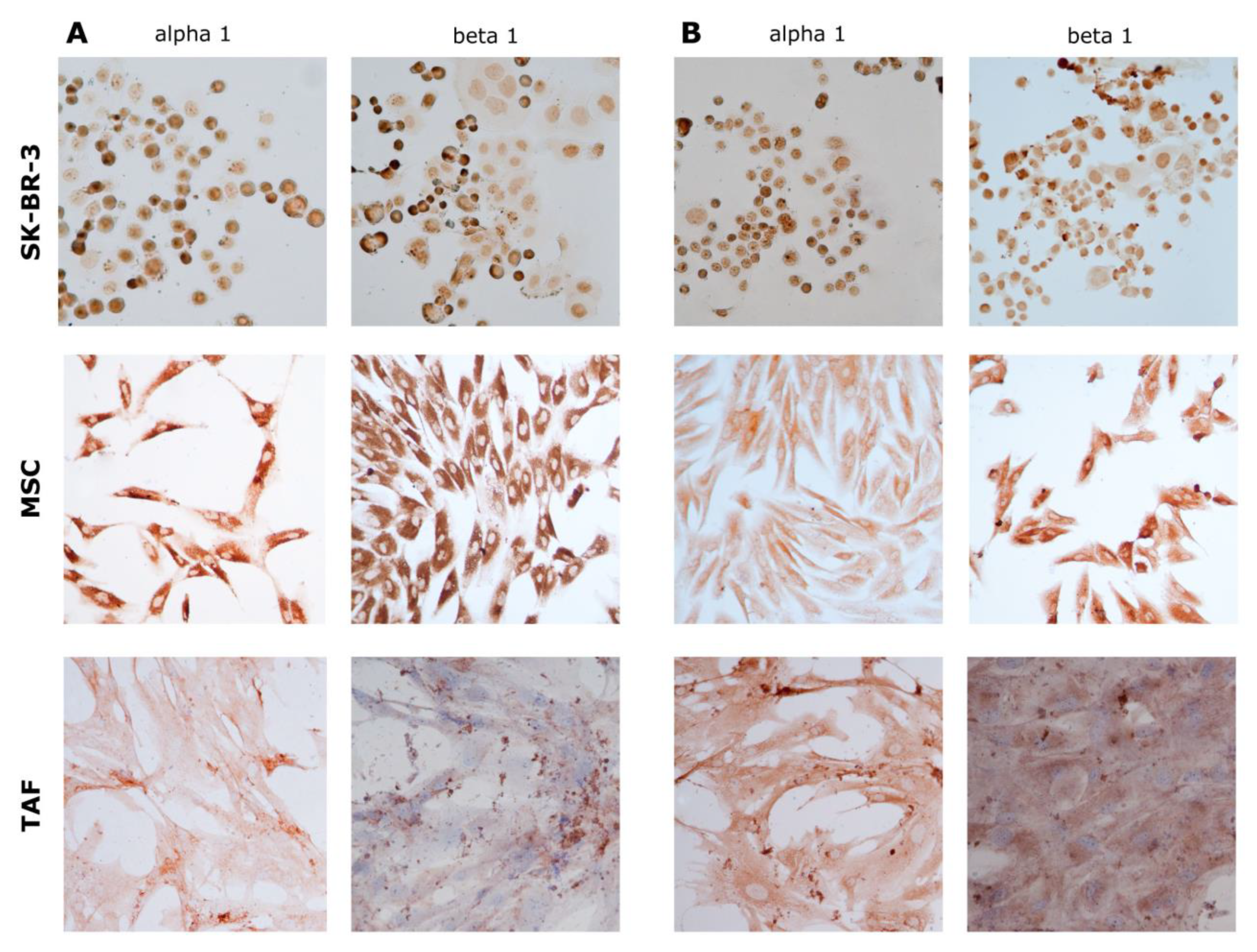
2.2. Immunocytochemistry
2.3. Flow Cytometry
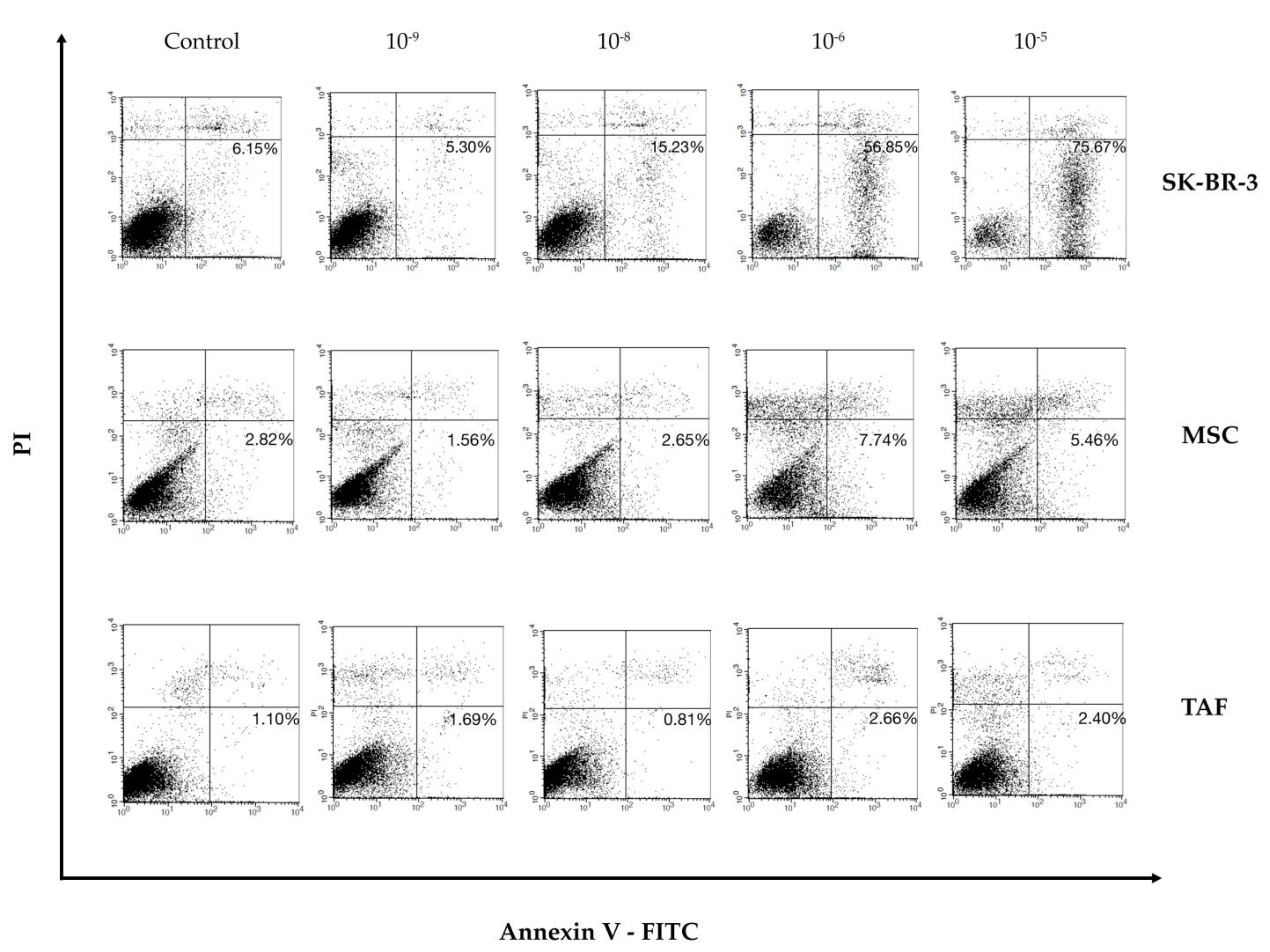
2.4. Annexin V/PI Assay
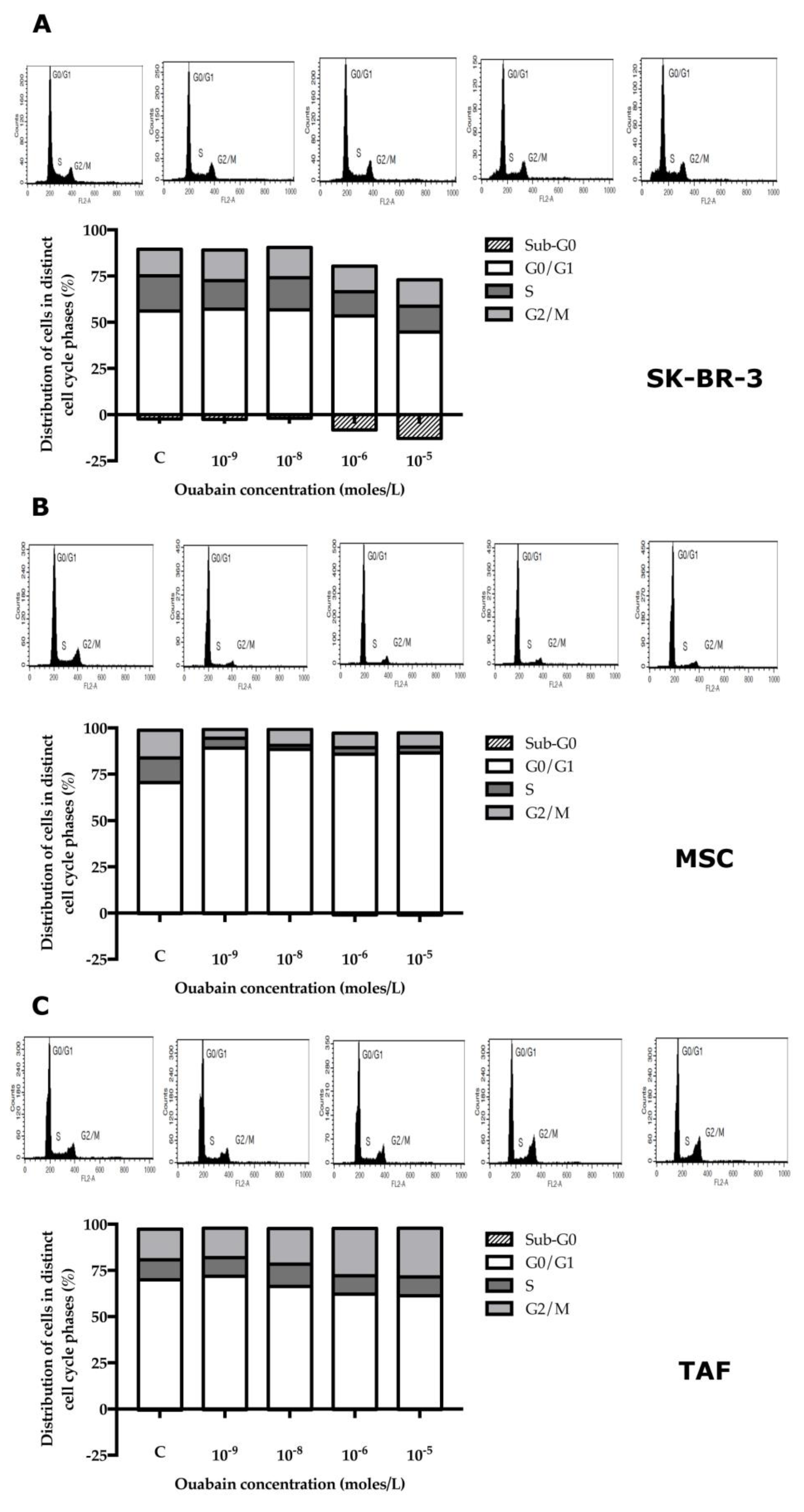
2.5. Cell Cycle Test
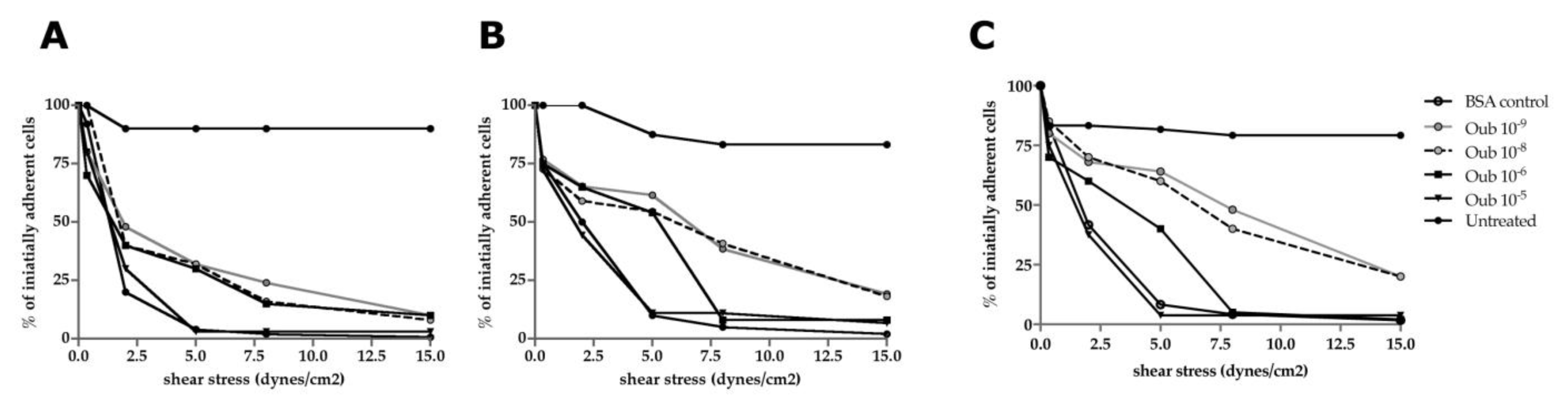
2.6. Flow Chamber Assay
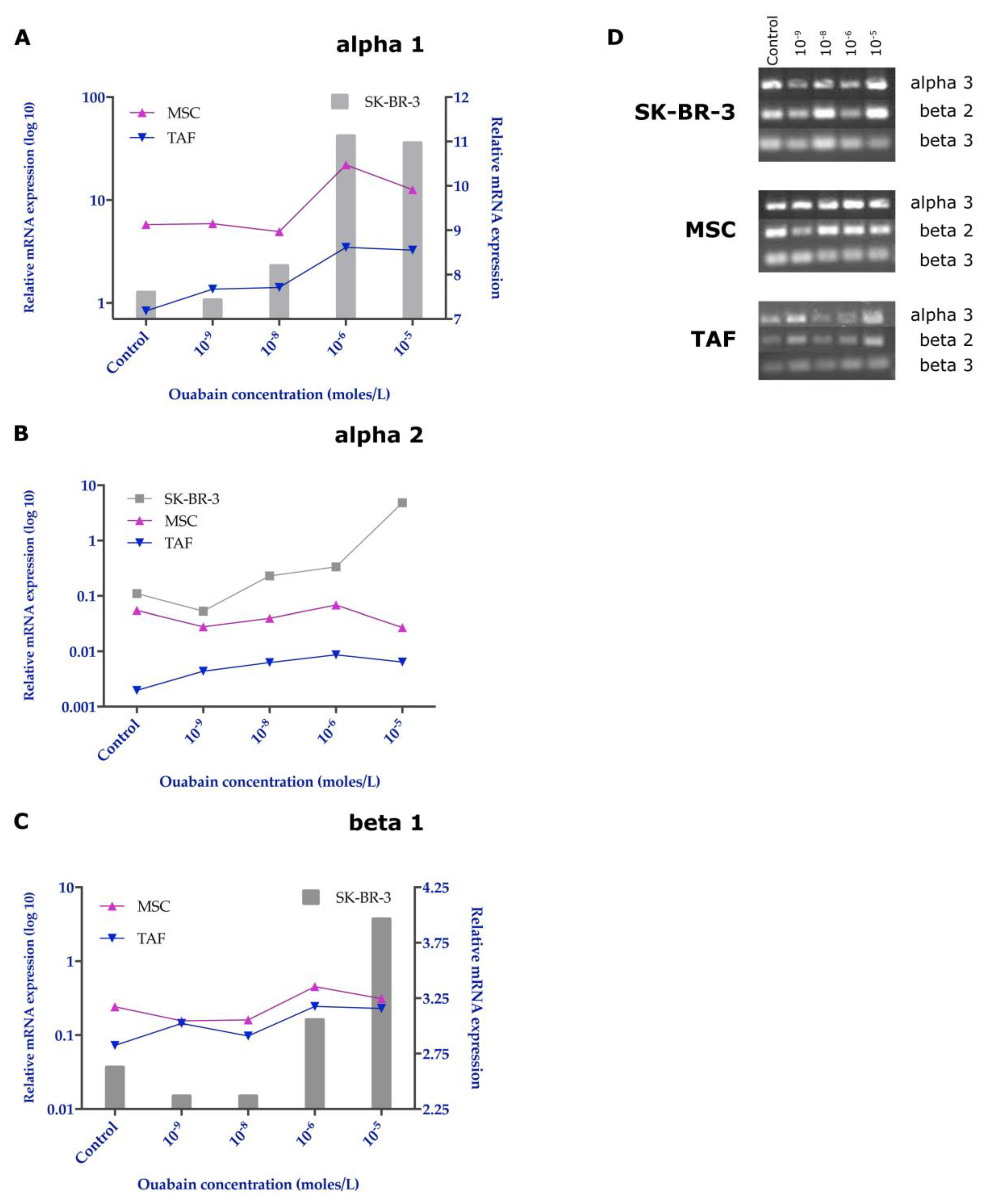
2.7. RNA Extraction and RT-PCR
2.8. Electron Microscopy
2.9. Statistical Analysis
3. Results
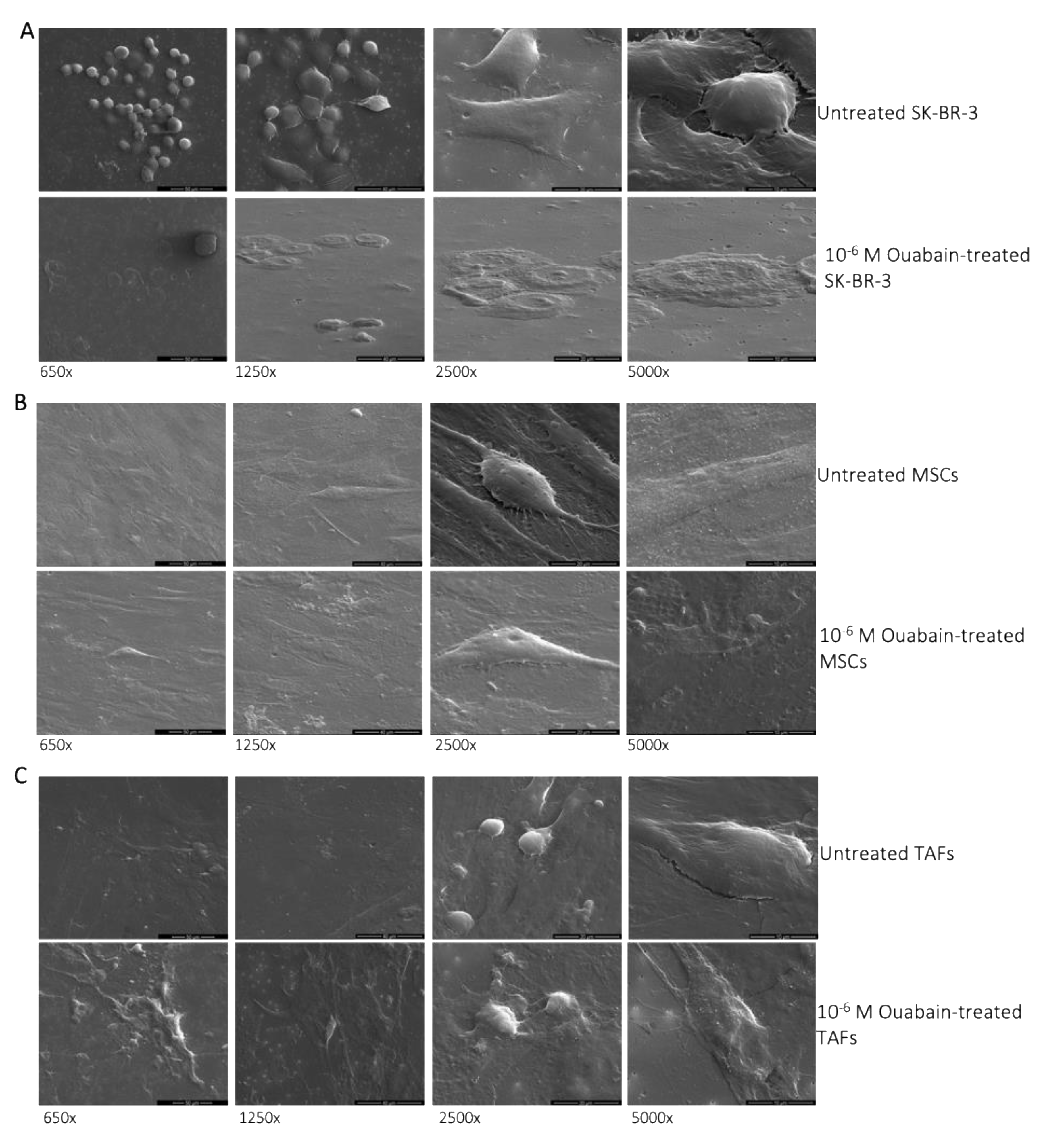
3.1. Morphological Changes of Ouabain-Treated Cells
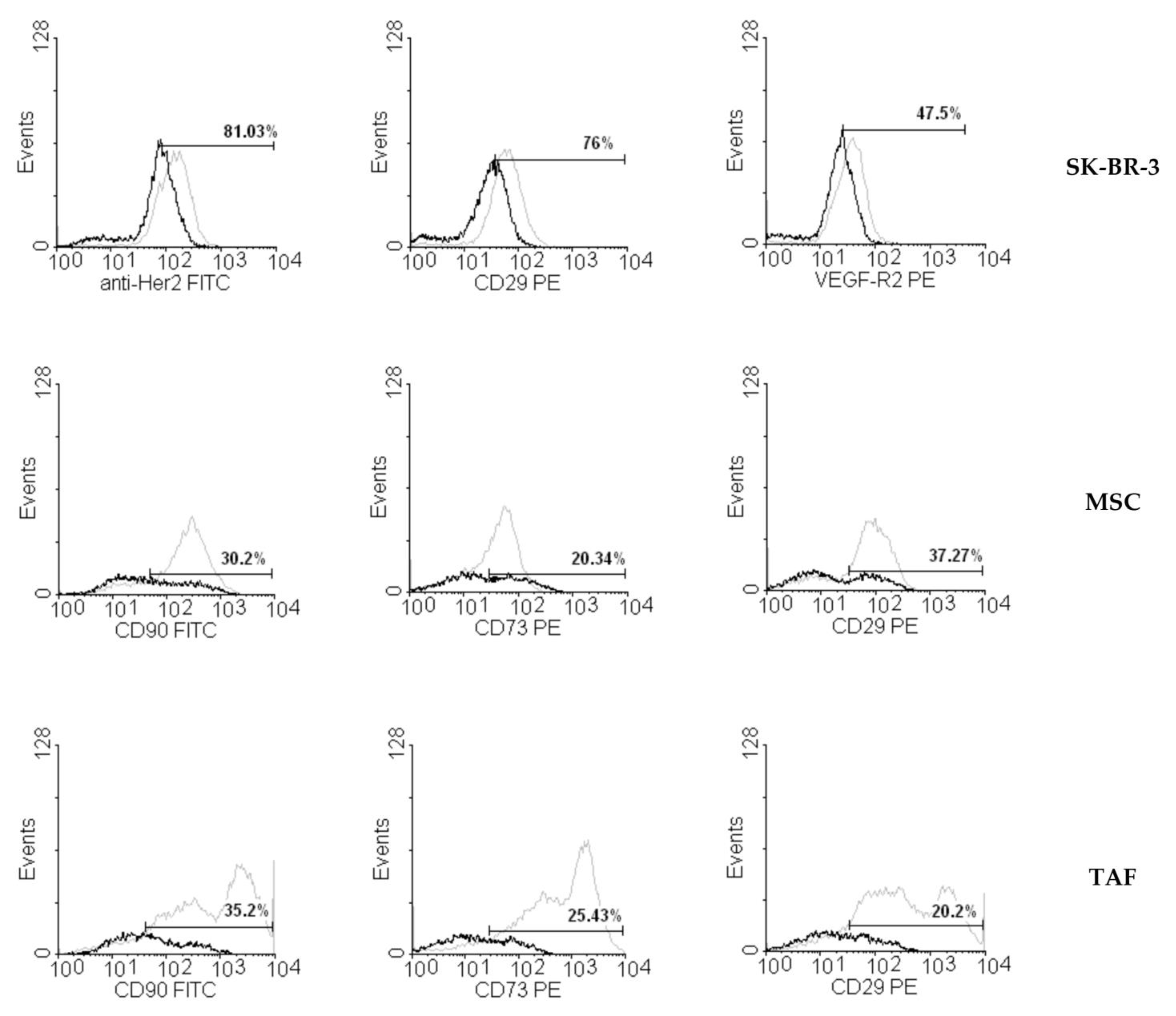
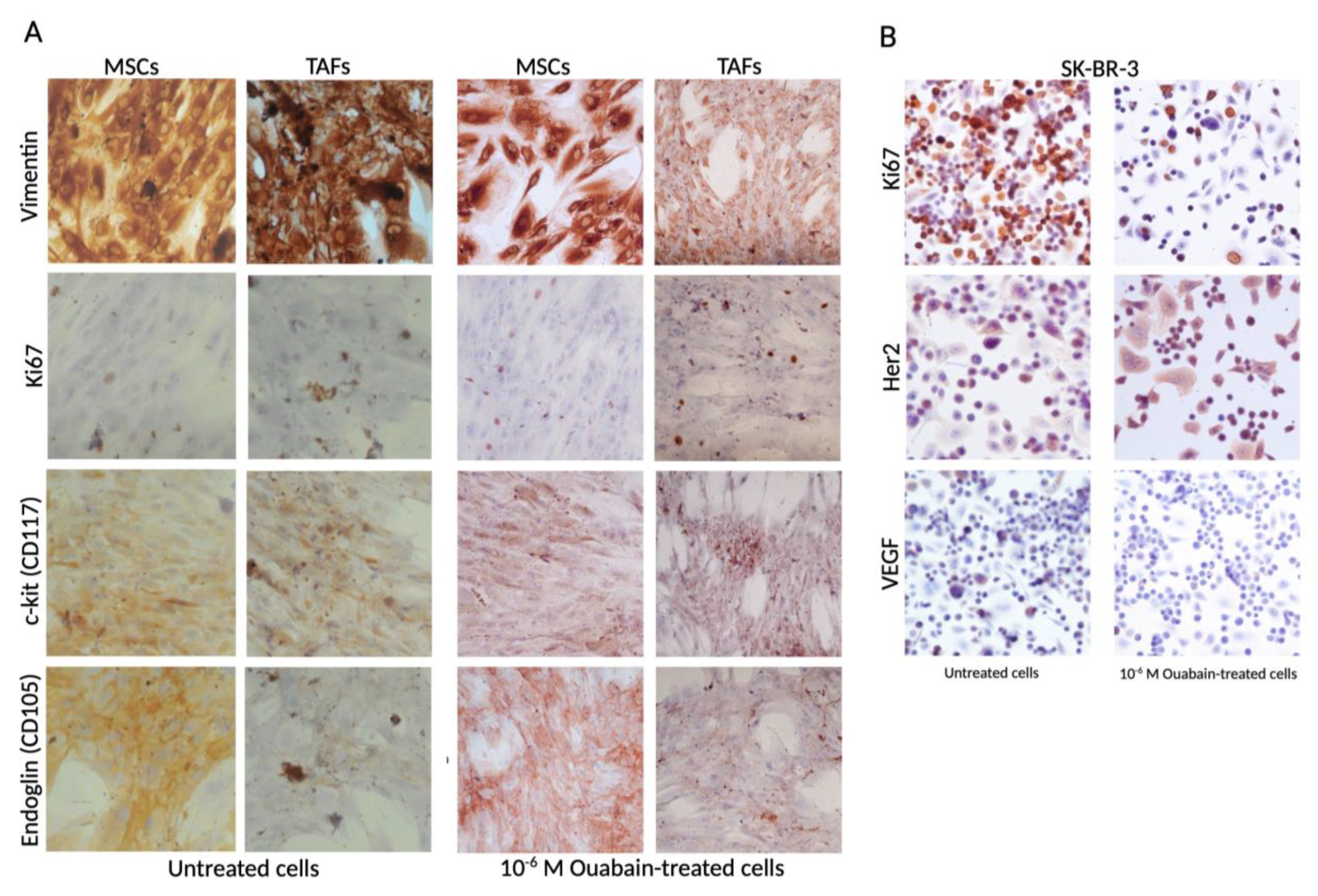
3.2. Ouabain-Induced Immunophenotypic Changes in Tumor, Tumor-Associated Cells, and MSCs
3.3. Effect of Ouabain on Cellular Viability and Cell Cycle
3.4. Functional Studies—Flow Chamber Assay
3.5. Effects of Ouabain on the Expression of the Na+/K+ Pump Subunits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jung, J.; Ryu, S.; Ki, I.A.; Woo, H.A.; Lee, K. Some Biological Consequences of the Inhibition of Na,K-ATPase by Translationally Controlled Tumor Protein (TCTP). Int. J. Mol. Sci. 2018, 19, 1657. [Google Scholar] [CrossRef]
- Bommer, U.A.; Thiele, B.J. The translationally controlled tumour protein (TCTP). Int. J. Biochem. Cell Biol. 2004, 36, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Pirkmajer, S.; Chibalin, A.V. Hormonal regulation of Na+-K+-ATPase from the evolutionary perspective. Curr. Top. Membr. 2019, 83, 315–351. [Google Scholar] [CrossRef] [PubMed]
- Botelho, A.F.M.; Pierezan, F.; Soto-Blanco, B.; Melo, M.M. A review of cardiac glycosides: Structure, toxicokinetics, clinical signs, diagnosis and antineoplastic potential. Toxicon 2019, 158, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Kumavath, R.; Paul, S.; Pavithran, H.; Paul, M.K.; Ghosh, P.; Barh, D.; Azevedo, V. Emergence of Cardiac Glycosides as Potential Drugs: Current and Future Scope for Cancer Therapeutics. Biomolecules 2021, 11, 1275. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxid. Med. Cell. Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef]
- Fujii, T.; Shimizu, T.; Yamamoto, S.; Funayama, K.; Fujita, K.; Tabuchi, Y.; Ikari, A.; Takeshima, H.; Sakai, H. Crosstalk between Na+,K+-ATPase and a vol-ume-regulated anion channel in membrane microdomains of human cancer cells. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3792–3804. [Google Scholar] [CrossRef]
- Panizza, E.; Zhang, L.; Fontana, J.M.; Hamada, K.; Svensson, D.; Akkuratov, E.E.; Scott, L.; Mikoshiba, K.; Brismar, H.; Lehtiö, J.; et al. Ouabain-regulated phosphoproteome re-veals molecular mechanisms for Na+, K+-ATPase control of cell adhesion, proliferation, and survival. FASEB J. 2019, 33, 10193–10206. [Google Scholar] [CrossRef] [PubMed]
- Askari, A. The sodium pump and digitalis drugs: Dogmas and fallacies. Pharmacol. Res. Perspect. 2019, 7, e00505. [Google Scholar] [CrossRef]
- Chang, Y.M.; Shih, Y.L.; Chen, C.P.; Liu, K.L.; Lee, M.H.; Lee, M.Z.; Hou, H.T.; Huang, H.C.; Lu, H.F.; Peng, S.F.; et al. Ouabain induces apoptotic cell death in human prostate DU 145 cancer cells through DNA damage and TRAIL pathways. Environ. Toxicol. 2019, 34, 1329–1339. [Google Scholar] [CrossRef]
- Khajah, M.A.; Mathew, P.M.; Luqmani, Y.A. Na+/K+ ATPase activity promotes invasion of endocrine resistant breast cancer cells. PLoS ONE 2018, 13, e0193779. [Google Scholar] [CrossRef]
- Xie, Z. Molecular mechanisms of Na/K-ATPase-mediated signal transduction. Ann. N. Y. Acad. Sci. 2003, 986, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Kovacevic, Z.; Peng, Z.; Jin, R.; Wang, P.; Yue, F.; Zheng, M.; Huang, M.L.-H.; Jansson, P.J.; Richardson, V.; et al. The molecular effect of metastasis suppressors on Src signaling and tumorigenesis: New therapeutic targets. Oncotarget 2015, 6, 35522–35541. [Google Scholar] [CrossRef]
- Du, J.; Jiang, L.; Chen, F.; Hu, H.; Zhou, M. Cardiac Glycoside Ouabain Exerts Anticancer Activi-ty via Downregulation of STAT3. Front. Oncol. 2021, 11, 684316. [Google Scholar] [CrossRef] [PubMed]
- Menger, L.; Vacchelli, E.; Adjemian, S.; Martins, I.; Ma, Y.; Shen, S.; Yamazaki, T.; Sukkurwala, A.Q.; Michaud, M.; Mignot, G.; et al. Cardiac glycosides exert anticancer effects by inducing immunogenic cell death. Sci. Transl. Med. 2012, 4, 143ra99. [Google Scholar] [CrossRef] [PubMed]
- Paunescu, V.; Bojin, F.M.; Tatu, C.A.; Gavriliuc, O.I.; Rosca, A.; Gruia, A.T.; Tanasie, G.; Bunu, C.; Crisnic, D.; Gherghiceanu, M.; et al. Tumour-associated fibroblasts and mesenchymal stem cells: More similarities than differences. J. Cell. Mol. Med. 2011, 15, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Aperia, A.; Akkuratov, E.E.; Fontana, J.M.; Brismar, H. Na+-K+-ATPase, a new class of plasma membrane receptors. Am. J. Physiol. Cell Physiol. 2016, 310, C491–C495. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Bai, F.; Chaudhry, M.A.; Pratt, R.; Shapiro, J.I.; Liu, J. The Na/K-ATPase α1 and c-Src form signaling complex under na-tive condition: A crosslinking approach. Sci. Rep. 2020, 10, 6006. [Google Scholar] [CrossRef]
- Yang, X.S.; Xu, Z.W.; Yi, T.L.; Xu, R.C.; Li, J.; Zhang, W.B.; Zhang, S.; Sun, H.T.; Yu, Z.Q.; Xu, H.X.; et al. Ouabain suppresses the growth and migration abilities of glioma U-87MG cells through inhibiting the Akt/mTOR signaling pathway and downregulating the expression of HIF-1α. Mol. Med. Rep. 2018, 17, 5595–5600. [Google Scholar] [CrossRef]
- Xiao, Y.; Meng, C.; Lin, J.; Huang, C.; Zhang, X.; Long, Y.; Huang, Y.; Lin, Y. Ouabain targets the Na+/K+-ATPase α3 isoform to inhibit cancer cell proliferation and induce apoptosis. Oncol. Lett. 2017, 14, 6678–6684. [Google Scholar] [CrossRef]
- Ninsontia, C.; Chanvorachote, P. Ouabain mediates integrin switch in human lung cancer cells. Anticancer Res. 2014, 34, 5495–5502. [Google Scholar] [PubMed]
- de Souza, W.F.; Barbosa, L.A.; Liu, L.; de Araujo, W.M.; De-Freitas-Junior, J.C.M.; Fortunato-Miranda, N.; Fontes, C.F.; Morgado-Díaz, J.A. Ouabain-induced alterations of the apical junctional complex involve α1 and β1 Na,K-ATPase downregulation and ERK1/2 activation in-dependent of caveolae in colorectal cancer cells. J. Membr. Biol. 2014, 247, 23–33. [Google Scholar] [CrossRef]
- Guo, W.; Wei, B.; Chen, T.; Xu, X.; Ruan, F.; Xiang, M. The Na+/K+ ATPase inhibitor ouabain attenuates stemness and chemo-resistance of osteosarcoma cells. Med. Sci. Monit. 2019, 25, 9426–9434. [Google Scholar] [CrossRef]
- Saito, S.; Ohtsu, M.; Asano, M.; Ishigami, T. Ouabain signaling in oral squamous cell carcinoma cells. J. Oral Sci. 2019, 61, 498–503. [Google Scholar] [CrossRef]
- da Silva, J.M.C.; Azevedo, A.D.N.; Barbosa, R.P.D.S.; Teixeira, M.P.; Vianna, T.A.G.; Fittipaldi, J.; Cabral, V.R.; de Paiva, L.S. Ouabain Decreases Regulatory T Cell Number in Mice by Reducing IL-2 Secretion. Neuroimmunomodulation 2019, 26, 188–197. [Google Scholar] [CrossRef]
- Dang, C.V. MYC on the Path to Cancer. Cell 2012, 149, 22–35. [Google Scholar] [CrossRef]
- Carroll, P.A.; Freie, B.W.; Mathsyaraja, H.; Eisenman, R.N. The MYC transcription factor network: Balancing metabolism, pro-liferation and oncogenesis. Front. Med. 2018, 12, 412–425. [Google Scholar] [CrossRef]
- Dupont, G.; Combettes, L.; Bird, G.S.; Putney, J.W. Calcium oscillations. Cold Spring Harb. Perspect. Biol. 2011, 3, a004226. [Google Scholar] [CrossRef] [PubMed]
- Santivasi, W.L.; Xia, F. Ionizing radiation-induced DNA damage, response, and repair. Antioxid. Redox Signal. 2014, 21, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Hiyoshi, H.; Abdelhady, S.; Segerström, L.; Sveinbjörnsson, B.; Nuriya, M.; Lundgren, T.K.; Desfrere, L.; Miyakawa, A.; Yasui, M.; Kogner, P.; et al. Quiescence and γH2AX in neuroblastoma are regulated by ouabain/Na,K-ATPase. Br. J. Cancer 2012, 106, 1807–1815. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Li, Y.; Su, S.; Wang, N.; Wang, H.; Li, J. Inhibition of cell migration by ouabain in the A549 human lung cancer cell line. Oncol. Lett. 2013, 6, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Pongrakhananon, V.; Chunhacha, P.; Chanvorachote, P. Ouabain Suppresses the Migratory Behavior of Lung Cancer Cells. PLoS ONE 2013, 8, e68623. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Liu, K.; Zhang, Y.; Jiang, M.; Lu, R.; Folts, C.J.; Gao, X.; Noble, M.D.; Zhao, T.; Zhou, Z.; et al. Pharmacological targeting of p38 MAP-Kinase 6 (MAP2K6) inhibits the growth of esophageal adenocarcinoma. Cell. Signal. 2018, 51, 222–232. [Google Scholar] [CrossRef]
- Clausen, M.; Hilbers, F.; Poulsen, H. The structure and function of the Na,K-ATPase isoforms in health and disease. Front. Physiol. 2017, 8, 371. [Google Scholar] [CrossRef]
- Mijatovic, T.; Dufrasne, F.; Kiss, R. Na+/K+-ATPase and cancer. Pharm Pat Anal. 2012, 1, 91–106. [Google Scholar] [CrossRef]
- Ribeiro Franco, P.I.; Rodrigues, A.P.; de Menezes, L.B.; Pacheco Miguel, M. Tumor microenvironment components: Allies of cancer progression. Pathol. Res. Pract. 2020, 216, 152729. [Google Scholar] [CrossRef]
- Vinay, D.S.; Ryan, E.P.; Pawelec, G.; Talib, W.H.; Stagg, J.; Elkord, E.; Lichtor, T.; Decker, W.K.; Whelan, R.L.; Kumara, H.M.C.S.; et al. Immune evasion in cancer: Mechanistic basis and therapeutic strategies. Semin. Cancer Biol. 2015, 35, S185–S198. [Google Scholar] [CrossRef]
- Cassim, S.; Pouyssegur, J. Tumor microenvironment: A metabolic player that shapes the immune response. Int. J. Mol. Sci. 2020, 21, 157. [Google Scholar] [CrossRef]
- Cavalcante-Silva, L.H.A.; Lima, É.A.; Carvalho, D.C.M.; de Sales-Neto, J.M.; Alves, A.K.D.A.; Galvão, J.G.F.M.; Silva, J.S.D.F.D.; Rodrigues-Mascarenhas, S. Much more than a cardiotonic steroid: Modulation of inflammation by ouabain. Front. Physiol. 2017, 8, 895. [Google Scholar] [CrossRef]
- Shen, J.-J.; Zhan, Y.-C.; Li, H.-Y.; Wang, Z. Ouabain impairs cancer metabolism and activates AMPK-Src signaling pathway in human cancer cell lines. Acta Pharmacol. Sin. 2020, 41, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z. Na+/K+-ATPase-mediated signal transduction: From protein interaction to cellular function. Mol. Interv. 2003, 3, 157–168. [Google Scholar] [CrossRef]
- Ye, Q.; Lai, F.; Duan, Q.; Li, Z.; Si, A.; Xie, Z. Expression of mutant α1 Na/K-ATPase defective in conformational transition at-tenuates Src-mediated signal transduction. J. Biol. Chem. 2013, 288, 5803–5814. [Google Scholar] [CrossRef] [PubMed]
- Vagin, O.; Dada, L.A.; Tokhtaeva, E.; Sachs, G. The Na-K-ATPase α1β1 heterodimer as a cell adhesion molecule in epithelia. Am. J. Physiol. Cell Physiol. 2012, 302, C1271–C1281. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Na/K-ATPase alpha 1 | Forward 5′-AAAAACATGGTCCCTCAGCAA-3′ Reverse 5′-CCACAACTTCCTCCGCATTT-3′ | NM_000701.7 (tr. Var 1) 76 bp |
| Na/K-ATPase alpha 2 | Forward 5′-GAATGAGAGGCTCATCAGCATG-3′ Reverse 5′-CAAAGTAGGTGAAGAAGCCACCC-3′ | NM_000702.3 77 bp |
| Na/K-ATPase alpha 3 | Forward 5′-AATGCCTACCTTGAGCTCGG-3′ Reverse 5′-CTCGGGCAGGTAATAATGGC-3′ | NM_152296.3 69 bp |
| Na/K-ATPase alpha 4 | Forward 5′-GATGATCACAAATTAACCTTGGAAGA-3′ Reverse 5′-TTTGCCCTTTGGTGGCTATG-3′ | NM_144699.3 (tr. Var 1) 83 bp |
| Na/K-ATPase beta 1 | Forward 5′-TCAGTGAATTTAAGCCCACATATCA-3′ Reverse 5′-CTTCTGGATCTGAGGAATCTGTGTT-3′ | NM_001677.3 74 bp |
| Na/K-ATPase beta 2 | Forward 5′-CCAGCATGTTCAGAAGCTCAAC-3′ Reverse 5′-GCGGCAGACATCATTCTTTTG-3′ | NM_001678.3 79 bp |
| Na/K-ATPase beta 3 | Forward 5′-CTGGCCGAGTGGAAGCTC-3′ Reverse 5′-GGTGCGCCCCAGGAA-3′ | NM_001679.2 60 bp |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harich, O.-O.; Gavriliuc, O.-I.; Ordodi, V.-L.; Tirziu, A.; Paunescu, V.; Panaitescu, C.; Bojin, M.-F. In Vitro Study of the Multimodal Effect of Na+/K+ ATPase Blocker Ouabain on the Tumor Microenvironment and Malignant Cells. Biomedicines 2023, 11, 2205. https://doi.org/10.3390/biomedicines11082205
Harich O-O, Gavriliuc O-I, Ordodi V-L, Tirziu A, Paunescu V, Panaitescu C, Bojin M-F. In Vitro Study of the Multimodal Effect of Na+/K+ ATPase Blocker Ouabain on the Tumor Microenvironment and Malignant Cells. Biomedicines. 2023; 11(8):2205. https://doi.org/10.3390/biomedicines11082205
Chicago/Turabian StyleHarich, Octavia-Oana, Oana-Isabella Gavriliuc, Valentin-Laurentiu Ordodi, Alexandru Tirziu, Virgil Paunescu, Carmen Panaitescu, and Maria-Florina Bojin. 2023. "In Vitro Study of the Multimodal Effect of Na+/K+ ATPase Blocker Ouabain on the Tumor Microenvironment and Malignant Cells" Biomedicines 11, no. 8: 2205. https://doi.org/10.3390/biomedicines11082205
APA StyleHarich, O.-O., Gavriliuc, O.-I., Ordodi, V.-L., Tirziu, A., Paunescu, V., Panaitescu, C., & Bojin, M.-F. (2023). In Vitro Study of the Multimodal Effect of Na+/K+ ATPase Blocker Ouabain on the Tumor Microenvironment and Malignant Cells. Biomedicines, 11(8), 2205. https://doi.org/10.3390/biomedicines11082205