
Vicarious Social Defeat Increases Conditioned Rewarding Effects of Cocaine and Ethanol Intake in Female Mice


 , ,
, ,
Abstract
1. Introduction
Aims
2. Materials and Methods
2.1. Study Design
2.2. Drugs
2.3. Apparatus and Procedures
2.3.1. Procedure of Vicarious Social Defeat (VSD)
2.3.2. Determination of Plasma Corticosterone (ELISA)
2.3.3. Procedure of the Social Interaction Test (SIT) or Social Withdrawal Ratio
2.3.4. Elevated Plus Maze (EPM)
2.3.5. Splash Test (ST)
2.3.6. Tail Suspension Test (TST)
2.3.7. Conditioned Place Preference (CPP)
2.3.8. Drinking in the Dark (DID)
2.3.9. Oral Ethanol Self-Administration (SA)
2.3.10. Tissue Sampling and Determination of Striatal IL-6
2.4. Statistical Analyses
3. Results
3.1. VSD Female Showed Increased Corticosterone Levels after Vicarious Experience
3.2. VSD Did Not Affect Bodyweight
3.3. VSD Females Did Not Show Social Avoidance or Despair Behaviors
3.4. VSD Females Showed Increased Anxiety and Anhedonia
3.5. VSD Females Showed Increased Conditioned Rewarding Effects of Cocaine
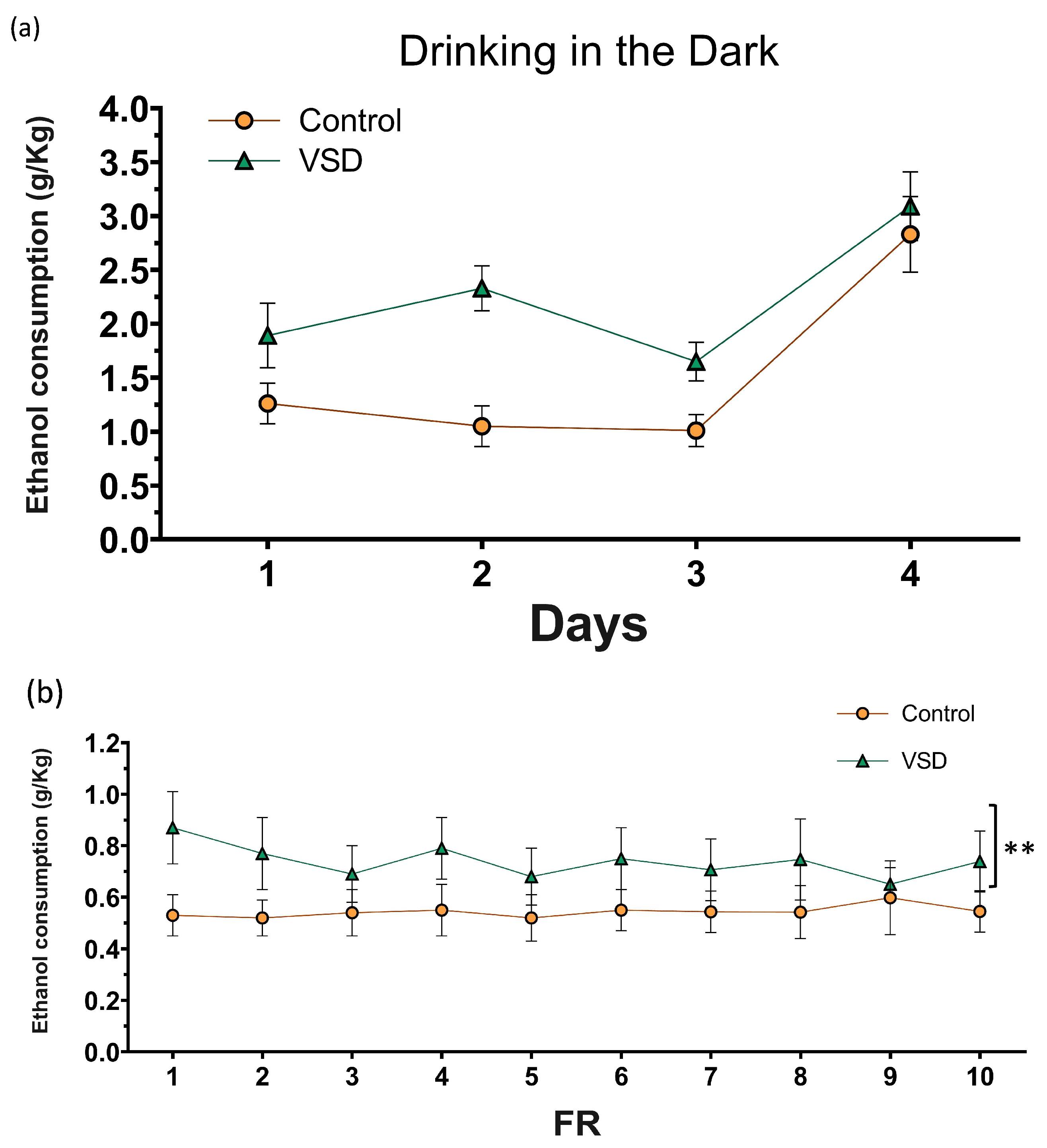
3.6. VSD Females Showed Increased Ethanol Intake
3.7. VSD Did Not Increase Striatal IL-6 Levels in Female Mice
4. Discussion
4.1. VSD Induced an Increase in Corticosterone Levels
4.2. VSD Increased the Conditioned Rewarding Effects of Cocaine
4.3. VSD Increased Ethanol Intake in Stressed Females
4.4. VSD Induce Anxiety-and Anhedonic-like Behaviors
4.5. Limitations and Future Perspectives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Swaab, D.F.; Bao, A.M. Sex differences in stress-related disorders: Major depressive disorder, bipolar disorder, and posttraumatic stress disorder. Handb. Clin. Neurol. 2020, 175, 335–358. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, V.; Heiming, R.S.; Jansen, F.; Lesting, J.; Sachser, N.; Pape, H.C.; Seidenbecher, T. Social defeat: Impact on fear extinction and amygdala- prefrontal cortical theta synchrony in 5-HTT deficient mice. PLoS ONE 2011, 6, e22600. [Google Scholar] [CrossRef]
- Harvey, B.H.; Naciti, C.; Brand, L.; Stein, D.J. Serotonin and stress: Protective or malevolent actions in the biobehavioral response to repeated trauma? Ann. N. Y. Acad. Sci. 2004, 1032, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Amato, J.L.; Bankson, M.G.; Yamamoto, B.K. Prior exposure to chronic stress and MDMA potentiates mesoaccumbens dopamine release mediated by the 5-HT(1B) receptor. Neuropsychopharmacology 2007, 32, 946–954. [Google Scholar] [CrossRef] [PubMed]
- Keeney, A.; Jessop, D.S.; Harbuz, M.S.; Marsden, C.A.; Hogg, S.; Blackburn-Munro, R.E. Differential effects of acute and chronic social defeat stress on hypothalamic- pituitary-adrenal axis function and hippocampal serotonin release in mice. J. Neuroendocrinol. 2006, 18, 330–338. [Google Scholar] [CrossRef]
- Azevedo, H.; Ferreira, M.; Mascarello, A.; Osten, P.; Werneck Guimarães, C.R. The serotonergic and alpha-1 adrenergic receptor modulator ACH-000029 ameliorates anxiety-like behavior in a post-traumatic stress disorder model. Neuropharmacology 2020, 164, 107912. [Google Scholar] [CrossRef]
- Laumet, G.; Zhou, W.; Dantzer, R.; Edralin, J.D.; Huo, X.; Budac, D.P.; O’Connor, J.C.; Lee, A.W.; Heijnen, C.J.; Kavelaars, A. Upregulation of neuronal kynurenine 3-monooxygenase mediates depression-like behavior in a mouse model of neuropathic pain. Brain. Behav. Immun. 2017, 66, 94–102. [Google Scholar] [CrossRef]
- Wang, J.; Hodes, G.E.; Zhang, H.; Zhang, S.; Zhao, W.; Golden, S.A.; Bi, W.; Menard, C.; Kana, V.; Leboeuf, M.; et al. Epigenetic modulation of inflammation and synaptic plasticity promotes resilience against stress in mice. Nat. Commun. 2018, 9, 477. [Google Scholar] [CrossRef]
- Gordon, H.W. Early environmental stress and biological vulnerability to drug abuse. Psychoneuroendocrinology 2002, 27, 115–126. [Google Scholar] [CrossRef]
- Koob, G.F.; Schulkin, J. Addiction and stress: An allostatic view. Neurosci. Biobehav. Rev. 2019, 106, 245–262. [Google Scholar] [CrossRef]
- Peltier, M.R.; Verplaetse, T.L.; Mineur, Y.S.; Petrakis, I.L.; Cosgrove, K.P.; Picciotto, M.R.; McKee, S.A. Sex differences in stress-related alcohol use. Neurobiol. Stress 2019, 10, 100149. [Google Scholar] [CrossRef]
- Guinle, M.I.B.; Sinha, R. The Role of Stress, Trauma, and Negative Affect in Alcohol Misuse and Alcohol Use Disorder in Women. Alcohol. Res. 2020, 40, 5. [Google Scholar] [CrossRef]
- Brady, K.T.; Dansky, B.S.; Sonne, S.C.; Saladin, M.E. Posttraumatic stress disorder and cocaine dependence. Order of onset. Am. J. Addict. 1998, 7, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Miczek, K.A.; Yap, J.J.; Covington, H.E. Social stress, therapeutics and drug abuse: Preclinical models of escalated and depressed intake. Pharmacol. Ther. 2008, 120, 102–128. [Google Scholar] [CrossRef] [PubMed]
- Russo, S.J.; Nestler, E.J. The brain reward circuitry in mood disorders. Nat. Rev. Neurosci. 2013, 14, 609–625, Erratum in Nat. Rev. Neurosci. 2013, 14, 736. [Google Scholar] [CrossRef]
- Tornatzky, W.; Miczek, K.A. Long-term impairment of autonomic circadian rhythms after brief intermittent social stress. Physiol. Behav. 1993, 53, 983–993. [Google Scholar] [CrossRef]
- Selten, J.P.; van der Ven, E.; Rutten, B.P.; Cantor-Graae, E. The social defeat hypothesis of schizophrenia: An update. Schizophr. Bull. 2013, 39, 1180–1186. [Google Scholar] [CrossRef]
- Berton, O.; McClung, C.A.; Dileone, R.J.; Krishnan, V.; Renthal, W.; Russo, S.J.; Graham, D.; Tsankova, N.M.; Bolanos, C.A.; Rios, M.; et al. Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. Science 2006, 311, 864–868. [Google Scholar] [CrossRef]
- Krishnan, V.; Han, M.H.; Graham, D.L.; Berton, O.; Renthal, W.; Russo, S.J.; Laplant, Q.; Graham, A.; Lutter, M.; Lagace, D.C.; et al. Molecular adaptations underlying susceptibility and resistance to social defeat in brain reward regions. Cell 2007, 131, 391–404. [Google Scholar] [CrossRef]
- Rubinow, D.R.; Schmidt, P.J. Sex differences and the neurobiology of affective disorders. Neuropsychopharmacology 2019, 44, 111–128. [Google Scholar] [CrossRef]
- Harris, A.Z.; Atsak, P.; Bretton, Z.H.; Holt, E.S.; Alam, R.; Morton, M.P.; Abbas, A.I.; Leonardo, E.D.; Bolkan, S.S.; Hen, R.; et al. A Novel Method for Chronic Social Defeat Stress in Female Mice. Neuropsychopharmacology 2018, 43, 1276–1283. [Google Scholar] [CrossRef]
- Trainor, B.C.; Pride, M.C.; Villalon Landeros, R.; Knoblauch, N.W.; Takahashi, E.Y.; Silva, A.L.; Crean, K.K. Sex differences in social interaction behavior following social defeat stress in the monogamous California mouse (Peromyscus californicus). PLoS ONE 2011, 6, e17405. [Google Scholar] [CrossRef]
- Greenberg, G.D.; Steinman, M.Q.; Doig, I.E.; Hao, R.; Trainor, B.C. Effects of social defeat on dopamine neurons in the ventral tegmental area in male and female California mice. Eur. J. Neurosci. 2015, 42, 3081–3094. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Chung, J.R.; Zhang, S.; Zhang, H.; Grossman, Y.; Aleyasin, H.; Flanigan, M.E.; Pfau, M.L.; Menard, C.; Dumitriu, D.; et al. Establishment of a repeated social defeat stress model in female mice. Sci. Rep. 2017, 7, 12838. [Google Scholar] [CrossRef]
- Yin, W.; Gallagher, N.R.; Sawicki, C.M.; McKim, D.B.; Godbout, J.P.; Sheridan, J.F. Repeated social defeat in female mice induces anxiety-like behavior associated with enhanced myelopoiesis and increased monocyte accumulation in the brain. Brain. Behav. Immun. 2019, 78, 131–142. [Google Scholar] [CrossRef] [PubMed]
- van Wingen, G.A.; Geuze, E.; Vermetten, E.; Fernandez, G. Perceived threat predicts the neural sequelae of combat stress. Mol. Psychiatry 2011, 16, 664–671. [Google Scholar] [CrossRef]
- Cougle, J.R.; Resnick, H.; Kilpatrick, D.G. Does prior exposure to interpersonal violence increase risk of PTSD following subsequent exposure? Behav. Res. Ther. 2009, 47, 1012–1017. [Google Scholar] [CrossRef]
- Warren, B.L.; Vialou, V.F.; Iñiguez, S.D.; Alcantara, L.F.; Wright, K.N.; Feng, J.; Kennedy, P.J.; Laplant, Q.; Shen, L.; Nestler, E.J.; et al. Neurobiological sequelae of witnessing stressful events in adult mice. Biol. Psychiatry 2013, 73, 7–14. [Google Scholar] [CrossRef]
- Hodes, G.E.; Pfau, M.L.; Leboeuf, M.; Golden, S.A.; Christoffel, D.J.; Bregman, D.; Rebusi, N.; Heshmati, M.; Aleyasin, H.; Warren, B.L.; et al. Individual differences in the peripheral immune system promote resilience versus susceptibility to social stress. Proc. Natl. Acad. Sci. USA 2014, 111, 16136–16141. [Google Scholar] [CrossRef]
- Iñiguez, S.D.; Riggs, L.M.; Nieto, S.J.; Dayrit, G.; Zamora, N.N.; Shawhan, K.L.; Cruz, B.; Warren, B.L. Social defeat stress induces a depression-like phenotype in adolescent male c57BL/6 mice. Stress 2014, 17, 247–255. [Google Scholar] [CrossRef]
- Iñiguez, S.D.; Flores-Ramirez, F.J.; Riggs, L.M.; Alipio, J.B.; Garcia-Carachure, I.; Hernandez, M.A.; Sanchez, D.O.; Lobo, M.K.; Serrano, P.A.; Braren, S.H.; et al. Vicarious Social Defeat Stress Induces Depression-Related Outcomes in Female Mice. Biol. Psychiatry 2018, 83, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Montagud-Romero, S.; Blanco-Gandía, M.C.; Reguilón, M.D.; Ferrer-Pérez, C.; Ballestín, R.; Miñarro, J.; Rodríguez-Arias, M. Social defeat stress: Mechanisms underlying the increase in rewarding effects of drugs of abuse. Eur. J. Neurosci. 2018, 48, 2948–2970. [Google Scholar] [CrossRef] [PubMed]
- Holly, E.N.; DeBold, J.F.; Miczek, K.A. Increased mesocorticolimbic dopamine during acute and repeated social defeat stress: Modulation by corticotropin releasing factor receptors in the ventral tegmental area. Psychopharmacology 2015, 232, 4469–4479. [Google Scholar] [CrossRef] [PubMed]
- Newman, E.L.; Leonard, M.Z.; Arena, D.T.; de Almeida, R.M.M.; Miczek, K.A. Social defeat stress and escalation of cocaine and alcohol consumption: Focus on CRF. Neurobiol. Stress 2018, 9, 151–165. [Google Scholar] [CrossRef]
- Ferrer-Pérez, C.; Castro-Zavala, A.; Luján, M.Á.; Filarowska, J.; Ballestín, R.; Miñarro, J.; Valverde, O.; Rodríguez-Arias, M. Oxytocin prevents the increase of cocaine-related responses produced by social defeat. Neuropharmacology 2019, 146, 50–64. [Google Scholar] [CrossRef]
- Hwa, L.S.; Holly, E.N.; DeBold, J.F.; Miczek, K.A. Social stress-escalated intermittent alcohol drinking: Modulation by CRF-R1 in the ventral tegmental area and accumbal dopamine in mice. Psychopharmacology 2016, 233, 681–690. [Google Scholar] [CrossRef]
- Ferrer-Pérez, C.; Reguilón, M.D.; Manzanedo, C.; Miñarro, J.; Rodríguez-Arias, M. Social Housing Conditions Modulate the Long-Lasting Increase in Cocaine Reward Induced by Intermittent Social Defeat. Front. Behav. Neurosci. 2019, 13, 148. [Google Scholar] [CrossRef]
- Rodriguez-Arias, M.; Navarrete, F.; Blanco-Gandia, M.C.; Arenas, M.C.; Bartoll-Andrés, A.; Aguilar, M.A.; Rubio, G.; Miñarro, J.; Manzanares, J. Social defeat in adolescent mice increases vulnerability to alcohol consumption. Addict. Biol. 2016, 21, 87–97. [Google Scholar] [CrossRef]
- Newman, E.L.; Albrechet-Souza, L.; Andrew, P.M.; Auld, J.G.; Burk, K.C.; Hwa, L.S.; Zhang, E.Y.; DeBold, J.F.; Miczek, K.A. Persistent escalation of alcohol consumption by mice exposed to brief episodes of social defeat stress: Suppression by CRF-R1 antagonism. Psychopharmacology 2018, 235, 1807–1820. [Google Scholar] [CrossRef]
- Giménez-Gómez, P.; Ballestín, R.; Gil de Biedma-Elduayen, L.; Vidal, R.; Ferrer-Pérez, C.; Reguilón, M.D.; O’Shea, E.; Miñarro, J.; Colado, M.I.; Rodríguez-Arias, M. Decreased kynurenine pathway potentiate resilience to social defeat effect on cocaine reward. Neuropharmacology 2021, 197, 108753. [Google Scholar] [CrossRef]
- Reguilón, M.D.; Ballestín, R.; Miñarro, J.; Rodríguez-Arias, M. Resilience to social defeat stress in adolescent male mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2022, 119, 110591. [Google Scholar] [CrossRef] [PubMed]
- Newman, E.L.; Covington, H.E., 3rd; Leonard, M.Z.; Burk, K.; Miczek, K.A. Hypoactive Thalamic Crh+ Cells in a Female Mouse Model of Alcohol Drinking After Social Trauma. Biol. Psychiatry 2021, 90, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Arias, M.; Miñarro, J.; Aguilar, M.A.; Pinazo, J.; Simón, V.M. Effects of risperidone and SCH 23390 on isolation-induced aggression in male mice. Eur. Neuropsychopharmacol. 1998, 8, 95–103. [Google Scholar] [CrossRef]
- Golden, S.A.; Covington, H.E.; Berton, O.; Russo, S.J. A standardized protocol for repeated social defeat stress in mice. Nat. Protoc. 2011, 6, 1183–1191. [Google Scholar] [CrossRef] [PubMed]
- Hodes, G.E.; Pfau, M.L.; Purushothaman, I.; Ahn, H.F.; Golden, S.A.; Christoffel, D.J.; Magida, J.; Brancato, A.; Takahashi, A.; Flanigan, M.E.; et al. Sex Differences in Nucleus Accumbens Transcriptome Profiles Associated with Susceptibility versus Resilience to Subchronic Variable Stress. J. Neurosci. 2015, 35, 16362–16376. [Google Scholar] [CrossRef]
- Planchez, B.; Surget, A.; Belzung, C. Animal models of major depression: Drawbacks and challenges. J. Neural. Transm. (Vienna) 2019, 126, 1383–1408. [Google Scholar] [CrossRef]
- Can, A.; Dao, D.T.; Terrillion, C.E.; Piantadosi, S.C.; Bhat, S.; Gould, T.D. The tail suspension test. J. Vis. Exp. 2012, 59, e3769. [Google Scholar] [CrossRef]
- Rhodes, J.S.; Best, K.; Belknap, J.K.; Finn, D.A.; Crabbe, J.C. Evaluation of a simple model of ethanol drinking to intoxication in C57BL/6J mice. Physiol. Behav. 2005, 84, 53–63. [Google Scholar] [CrossRef]
- Navarrete, F.; Rubio, G.; Manzanares, J. Effects of naltrexone plus topiramate on ethanol self-administration and tyrosine hydroxylase gene expression changes. Addict. Biol. 2014, 19, 862–873. [Google Scholar] [CrossRef]
- Heffner, T.G.; Hartman, J.A.; Seiden, L.S. A rapid method for the regional dissection of the rat brain. Pharmacol. Biochem. Behav. 1980, 13, 453–456. [Google Scholar] [CrossRef]
- Solomon, M.B. Evaluating social defeat as a model for psychopathology in adult female rodents. J. Neurosci. Res. 2017, 95, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Carnevali, L.; Montano, N.; Tobaldini, E.; Thayer, J.F.; Sgoifo, A. The contagion of social defeat stress: Insights from rodent studies. Neurosci. Biobehav. Rev. 2020, 111, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Meyza, K.Z.; Bartal, I.B.; Monfils, M.H.; Panksepp, J.B.; Knapska, E. The roots of empathy: Through the lens of rodent models. Neurosci. Biobehav. Rev. 2017, 76 Pt B, 216–234. [Google Scholar] [CrossRef]
- Crick, N.R.; Grotpeter, J.K. Relational aggression, gender, and social-psychological adjustment. Child. Dev. 1995, 66, 710–722. [Google Scholar] [CrossRef]
- Montagud-Romero, S.; Nuñez, C.; Blanco-Gandia, M.C.; Martínez-Laorden, E.; Aguilar, M.A.; Navarro-Zaragoza, J.; Almela, P.; Milanés, M.V.; Laorden, M.L.; Miñarro, J.; et al. Repeated social defeat and the rewarding effects of cocaine in adult and adolescent mice: Dopamine transcription factors, proBDNF signaling pathways, and the TrkB receptor in the mesolimbic system. Psychopharmacology 2017, 234, 2063–2075. [Google Scholar] [CrossRef]
- Fox, H.C.; Sinha, R. Sex differences in drug-related stress-system changes: Implications for treatment in substance-abusing women. Harv. Rev. Psychiatry 2009, 17, 103–119. [Google Scholar] [CrossRef]
- Back, S.E.; Brady, K.T.; Jackson, J.L.; Salstrom, S.; Zinzow, H. Gender differences in stress reactivity among cocaine-dependent individuals. Psychopharmacology 2005, 180, 169–176. [Google Scholar] [CrossRef]
- Fox, H.C.; Hong, K.A.; Siedlarz, K.M.; Sinha, R. Enhanced sensitivity to stress and drug/alcohol craving in abstinent cocaine dependent individuals compared to social drinkers. Neuropsychopharmacology 2008, 33, 796–805. [Google Scholar] [CrossRef]
- McKay, J.R.; Rutherford, M.J.; Cacciola, J.S.; Kabasakalian-McKay, R.; Alterman, A.I. Gender differences in the relapse experiences of cocaine patients. J. Nerv. Ment. Dis. 1996, 184, 616–622. [Google Scholar] [CrossRef]
- Ferrer-Pérez, C.; Reguilón, M.D.; Miñarro, J.; Rodríguez-Arias, M. Endogenous oxytocin is essential for the buffering effects of pair housing against the increase in cocaine reward induced by social stress. Physiol. Behav. 2020, 221, 112913. [Google Scholar] [CrossRef]
- Holly, E.N.; Shimamoto, A.; Debold, J.F.; Miczek, K.A. Sex differences in behavioral and neural cross-sensitization and escalated cocaine taking as a result of episodic social defeat stress in rats. Psychopharmacology 2012, 224, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Shimamoto, A.; Holly, E.N.; Boyson, C.O.; DeBold, J.F.; Miczek, K.A. Individual differences in anhedonic and accumbal dopamine responses to chronic social stress and their link to cocaine self-administration in female rats. Psychopharmacology 2015, 232, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.M.; Zhou, X.; Cunningham, A.M.; Ramakrishnan, A.; Cates, H.M.; Lardner, C.K.; Peña, C.J.; Bagot, R.C.; Issler, O.; Van der Zee, Y.; et al. Crystallin Mu in Medial Amygdala Mediates the Effect of Social Experience on Cocaine Seeking in Males but Not in Females. Biol. Psychiatry 2022, 92, 895–906. [Google Scholar] [CrossRef]
- Caldwell, E.E.; Riccio, D.C. Alcohol self-administration in rats: Modulation by temporal parameters related to repeated mild social defeat stress. Alcohol 2010, 44, 265–274. [Google Scholar] [CrossRef]
- González-Portilla, M.; Montagud-Romero, S.; Navarrete, F.; Gasparyan, A.; Manzanares, J.; Miñarro, J.; Rodríguez-Arias, M. Pairing Binge Drinking and a High-Fat Diet in Adolescence Modulates the Inflammatory Effects of Subsequent Alcohol Consumption in Mice. Int. J. Mol. Sci. 2021, 22, 5279. [Google Scholar] [CrossRef]
- Albrechet-Souza, L.; Viola, T.W.; Grassi-Oliveira, R.; Miczek, K.A.; de Almeida, R.M.M. Corticotropin Releasing Factor in the Bed Nucleus of the Stria Terminalis in Socially Defeated and Non-stressed Mice with a History of Chronic Alcohol Intake. Front. Pharmacol. 2017, 8, 762. [Google Scholar] [CrossRef] [PubMed]
- Macedo, G.C.; Morita, G.M.; Domingues, L.P.; Favoretto, C.A.; Suchecki, D.; Quadros, I.M.H. Consequences of continuous social defeat stress on anxiety- and depressive-like behaviors and ethanol reward in mice. Horm. Behav. 2018, 97, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Dalla, C.; Antoniou, K.; Drossopoulou, G.; Xagoraris, M.; Kokras, N.; Sfikakis, A.; Papadopoulou-Daifoti, Z. Chronic mild stress impact: Are females more vulnerable? Neuroscience 2005, 135, 703–714. [Google Scholar] [CrossRef]
- Haller, J.; Baranyi, J.; Bakos, N.; Halász, J. Social instability in female rats: Effects on anxiety and buspirone efficacy. Psychopharmacology 2004, 174, 197–202. [Google Scholar] [CrossRef]
- Isingrini, E.; Camus, V.; Le Guisquet, A.M.; Pingaud, M.; Devers, S.; Belzung, C. Association between repeated unpredictable chronic mild stress (UCMS) procedures with a high fat diet: A model of fluoxetine resistance in mice. PLoS ONE 2010, 5, e10404. [Google Scholar] [CrossRef]
- Willner, P. Chronic mild stress (CMS) revisited: Consistency and behavioural-neurobiological concordance in the effects of CMS. Neuropsychobiology 2005, 52, 90–110. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Luo, Y.; Wang, H.; Kuang, S.; Liang, G.; Yang, Y.; Mai, S.; Yang, J. Re-evaluation of the interrelationships among the behavioral tests in rats exposed to chronic unpredictable mild stress. PLoS ONE 2017, 12, e0185129. [Google Scholar] [CrossRef] [PubMed]
- Baptista-de-Souza, D.; Rodrigues Tavares, L.R.; Canto-de-Souza, L.; Nunes-de-Souza, R.L.; Canto-de-Souza, A. Behavioral, hormonal, and neural alterations induced by social contagion for pain in mice. Neuropharmacology 2022, 203, 108878. [Google Scholar] [CrossRef] [PubMed]
- Von Frijtag, J.C.; Reijmers, L.G.; Van der Harst, J.E.; Leus, I.E.; Van den Bos, R.; Spruijt, B.M. Defeat followed by individual housing results in long-term impaired reward- and cognition-related behaviours in rats. Behav. Brain Res. 2000, 117, 137–146. [Google Scholar] [CrossRef]
- Rygula, R.; Abumaria, N.; Flügge, G.; Fuchs, E.; Rüther, E.; Havemann-Reinecke, U. Anhedonia and motivational deficits in rats: Impact of chronic social stress. Behav. Brain Res. 2005, 162, 127–134. [Google Scholar] [CrossRef]
- Lagace, D.C.; Donovan, M.H.; DeCarolis, N.A.; Farnbauch, L.A.; Malhotra, S.; Berton, O.; Nestler, E.J.; Krishnan, V.; Eisch, A.J. Adult hippocampal neurogenesis is functionally important for stress-induced social avoidance. Proc. Natl. Acad. Sci. USA 2010, 107, 4436–4441. [Google Scholar] [CrossRef]
- Mineur, Y.S.; Belzung, C.; Crusio, W.E. Effects of unpredictable chronic mild stress on anxiety and depression-like behavior in mice. Behav. Brain Res. 2006, 175, 43–50. [Google Scholar] [CrossRef]
- Lu, Q.; Xiang, H.; Zhu, H.; Chen, Y.; Lu, X.; Huang, C. Intranasal lipopolysaccharide administration prevents chronic stress-induced depression- and anxiety-like behaviors in mice. Neuropharmacology 2021, 200, 108816. [Google Scholar] [CrossRef]
- Lu, J.; Huang, C.; Lu, Q.; Lu, X. Therapeutic and Prophylactic Effects of Amphotericin B Liposomes on Chronic Social Defeat Stress-Induced Behavioral Abnormalities in Mice. Front. Pharmacol. 2022, 13, 918177. [Google Scholar] [CrossRef]
- Calpe-López, C.; Martínez-Caballero, M.A.; García-Pardo, M.P.; Aguilar, M.A. Intermittent voluntary wheel running promotes resilience to the negative consequences of repeated social defeat in mice. Physiol. Behav. 2022, 254, 113916. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Avgustinovich, D.F.; Kudryavtseva, N.N.; Murphy, D.L. BDNF in anxiety and depression. Science 2006, 312, 1598–1599. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.C.; Cao, X.; Das, M.; Zhu, X.H.; Gao, T.M. Behavioral animal models of depression. Neurosci. Bull. 2010, 26, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Baptista-de-Souza, D.; Nunciato, A.C.; Pereira, B.C.; Fachinni, G.; Zaniboni, C.R.; Canto-de-Souza, A. Mice undergoing neuropathic pain induce anxiogenic-like effects and hypernociception in cagemates. Behav. Pharmacol. 2015, 26, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Newman, E.L.; Covington, H.E., 3rd; Suh, J.; Bicakci, M.B.; Ressler, K.J.; DeBold, J.F.; Miczek, K.A. Fighting Females: Neural and Behavioral Consequences of Social Defeat Stress in Female Mice. Biol. Psychiatry 2019, 86, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Lukas, M.; Neumann, I.D. Social preference and maternal defeat-induced social avoidance in virgin female rats: Sex differences in involvement of brain oxytocin and vasopressin. J. Neurosci. Methods 2014, 234, 101–107. [Google Scholar] [CrossRef]
- Kim, J.; Pokharel, K.; Sandali, M.; Kim, C.S. Establishment of the Mouse Model of Social Avoidance Induced by Female-Directed Female Aggression. Chronic Stress (Thousand Oaks) 2022, 6, 24705470221129288. [Google Scholar] [CrossRef]
- Taylor, S.E.; Klein, L.C.; Lewis, B.P.; Gruenewald, T.L.; Gurung, R.A.; Updegraff, J.A. Biobehavioral responses to stress in females: Tend-and-befriend, not fight-or-flight. Psychol. Rev. 2000, 107, 411–429. [Google Scholar] [CrossRef]
- Finnell, J.E.; Lombard, C.M.; Padi, A.R.; Moffitt, C.M.; Wilson, L.B.; Wood, C.S.; Wood, S.K. Physical versus psychological social stress in male rats reveals distinct cardiovascular, inflammatory and behavioral consequences. PLoS ONE 2017, 12, e0172868. [Google Scholar]
- Bali, V.; Simmons, S.C.; Manning, C.E.; Doyle, M.A.; Rodriguez, M.; Stark, A.R.; Ayala Rosario, S.N.; Robison, A.J.; Mazei-Robison, M.S. Characterization of proinflammatory markers in the ventral tegmental area across mouse models of chronic stress. Neuroscience 2021, 461, 11–22. [Google Scholar] [CrossRef]
- Patki, G.; Salvi, A.; Liu, H.; Salim, S. Witnessing traumatic events and post-traumatic stress disorder: Insights from an animal model. Neurosci. Lett. 2015, 600, 28–32. [Google Scholar] [CrossRef]
- Warren, B.L.; Sial, O.K.; Alcantara, L.F.; Greenwood, M.A.; Brewer, J.S.; Rozofsky, J.P.; Parise, E.M.; Bolaños-Guzmán, C.A. Altered gene expression and spine density in nucleus accumbens of adolescent and adult male mice exposed to emotional and physical stress. Dev. Neurosci. 2014, 36, 250–260. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | VSD | |
|---|---|---|
| Time OA | 85 ± 13 | 49 ± 12 * |
| % Time OA | 43 ± 5 | 25 ± 6 * |
| Time center | 112 ± 12 | 115 ± 6 |
| Entries OA | 22 ± 3 | 17 ± 2 |
| % Entries OA | 51 ± 6 | 39 ± 4 |
| Total Entries | 43 ± 3 | 42 ± 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ródenas-González, F.; Arenas, M.C.; Blanco-Gandía, M.C.; Manzanedo, C.; Rodríguez-Arias, M. Vicarious Social Defeat Increases Conditioned Rewarding Effects of Cocaine and Ethanol Intake in Female Mice. Biomedicines 2023, 11, 502. https://doi.org/10.3390/biomedicines11020502
Ródenas-González F, Arenas MC, Blanco-Gandía MC, Manzanedo C, Rodríguez-Arias M. Vicarious Social Defeat Increases Conditioned Rewarding Effects of Cocaine and Ethanol Intake in Female Mice. Biomedicines. 2023; 11(2):502. https://doi.org/10.3390/biomedicines11020502
Chicago/Turabian StyleRódenas-González, Francisco, María Carmen Arenas, María Carmen Blanco-Gandía, Carmen Manzanedo, and Marta Rodríguez-Arias. 2023. "Vicarious Social Defeat Increases Conditioned Rewarding Effects of Cocaine and Ethanol Intake in Female Mice" Biomedicines 11, no. 2: 502. https://doi.org/10.3390/biomedicines11020502
APA StyleRódenas-González, F., Arenas, M. C., Blanco-Gandía, M. C., Manzanedo, C., & Rodríguez-Arias, M. (2023). Vicarious Social Defeat Increases Conditioned Rewarding Effects of Cocaine and Ethanol Intake in Female Mice. Biomedicines, 11(2), 502. https://doi.org/10.3390/biomedicines11020502