
Putative Complementary Compounds to Counteract Insulin-Resistance in PCOS Patients

 ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Endocrine Profile of PCOS
3. Metabolic Profile in PCOS
4. Pharmacological Therapies
5. Complementary/Integrative Treatments
5.1. Inositols
5.2. Alpha Lipoic Acid
5.3. Carnitines
5.4. N-acetylcysteine and L-arginine
5.5. Melatonin
5.6. Berberine
6. New Perspectives in Integrative Treatments for PCOS Patients
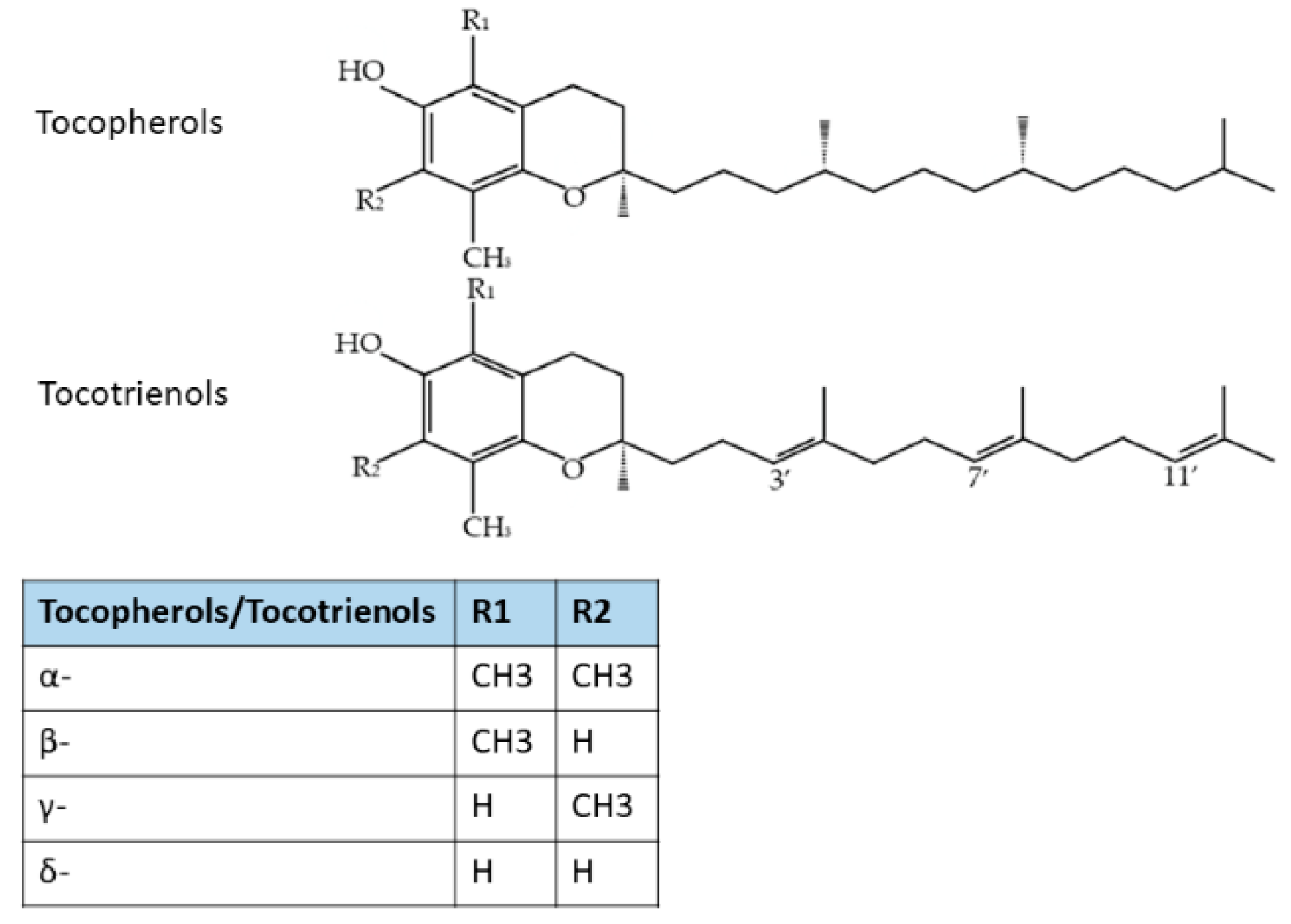
6.1. Tocotrienols
6.2. Decaffeinated Green Coffee
6.3. Gymnema Sylvestre
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Norman, R.J.; Dewailly, D.; Legro, R.S.; Hickey, T.E. Polycystic ovary syndrome. Lancet Lond. Engl. 2007, 370, 685–697. [Google Scholar] [CrossRef]
- Teede, H.J.; Misso, M.L.; Costello, M.F.; Dokras, A.; Laven, J.; Moran, L.; Piltonen, T.; Norman, R.J. Recommendatioon from the international evidence-based guidelin for the assessment and management of polycystic ovary syndrome. Hum. Reprod. 2018, 33, 1602–1618. [Google Scholar] [CrossRef] [PubMed]
- Moller, D.E.; Flier, J.S. Insulin resistance—Mechanisms, syndromes, and implications. N. Engl. J. Med. 1991, 325, 938–948. [Google Scholar] [CrossRef] [PubMed]
- Arslanian, S.A.; Lewy, V.D.; Danadian, K. Glucose intolerance in obese adolescents with polycystic ovary syndrome: Roles of insulin resistance and beta-cell dysfunction and risk of cardiovascular disease. J. Clin. Endocrinol. Metab. 2001, 86, 66–71. [Google Scholar] [CrossRef]
- Cussons, A.J.; Stuckey, B.G.A.; Watts, G.F. Metabolic syndrome and cardiometabolic risk in PCOS. Curr. Diab. Rep. 2007, 7, 66–73. [Google Scholar] [CrossRef]
- Rojas, J.; Chávez-Castillo, M.; Bermúdez, V. The Role of Metformin in Metabolic Disturbances during Pregnancy: Polycystic Ovary Syndrome and Gestational Diabetes Mellitus. Int. J. Reprod. Med. 2014, 2014, 797681. [Google Scholar] [CrossRef]
- Scarfò, G.; Daniele, S.; Fusi, J.; Gesi, M.; Martini, C.; Franzoni, F.; Cela, V.; Artini, P.G. Metabolic and Molecular Mechanisms of Diet and Physical Exercise in the Management of Polycystic Ovarian Syndrome. Biomedicines 2022, 10, 1305. [Google Scholar] [CrossRef]
- Rosenfield, R.L.; Ehrmann, D.A. The Pathogenesis of Polycystic Ovary Syndrome (PCOS): The Hypothesis of PCOS as Functional Ovarian Hyperandrogenism Revisited. Endocr. Rev. 2016, 37, 467–520. [Google Scholar] [CrossRef]
- Crespo, R.P.; Bachega, T.A.S.S.; Mendonça, B.B.; Gomes, L.G. An update of genetic basis of PCOS pathogenesis. Arch. Endocrinol. Metab. 2018, 62, 352–361. [Google Scholar] [CrossRef]
- De Melo, A.S.; Dias, S.V.; Cavalli, R.d.C.; Cardoso, V.C.; Bettiol, H.; Barbieri, M.A.; Ferriani, R.A.; Vieira, C.S. Pathogenesis of polycystic ovary syndrome: Multifactorial assessment from the foetal stage to menopause. Reprod. Camb. Engl. 2015, 150, R11–R24. [Google Scholar] [CrossRef]
- Alessandro, G.; Fedora, A.; Giulia, D.; Alba, M.; Martina, C.; Melania, A.; Tabatha, P.; Veronica, T.; Genazzani, A.R. Polycystic Ovary Syndrome: Considerations about therapeutic strategies choices from fertile life to menopause. In Impact of Polycystic Ovary, Metabolic Syndrome and Obesity on Women Health; ISGE Series; Springer Nature: Berlin, Germany, 2021; Volume 8. [Google Scholar]
- Azziz, R.; Carmina, E.; Dewailly, D.; Diamanti-Kandarakis, E.; Escobar-Morreale, H.F.; Futterweit, W.; Janssen, O.E.; Legro, R.S.; Norman, R.J.; Taylor, A.E.; et al. The Androgen Excess and PCOS Society criteria for the polycystic ovary syndrome: The complete task force report. Fertil. Steril. 2009, 91, 456–488. [Google Scholar] [CrossRef] [PubMed]
- Davinelli, S.; Nicolosi, D.; Di Cesare, C.; Scapagnini, G.; Di Marco, R. Targeting Metabolic Consequences of Insulin Resistance in Polycystic Ovary Syndrome by D-chiro-inositol and Emerging Nutraceuticals: A Focused Review. J. Clin. Med. 2020, 9, 987. [Google Scholar] [CrossRef] [PubMed]
- Samuel, V.T.; Shulman, G.I. Mechanisms for insulin resistance: Common threads and missing links. Cell 2012, 148, 852–871. [Google Scholar] [CrossRef]
- Plouffe, L. Disorders of excessive hair growth in the adolescent. Obstet. Gynecol. Clin. N. Am. 2000, 27, 79–99. [Google Scholar] [CrossRef]
- Zhu, J.-L.; Chen, Z.; Feng, W.-J.; Long, S.-L.; Mo, Z.-C. Sex hormone-binding globulin and polycystic ovary syndrome. Clin. Chim. Acta Int. J. Clin. Chem. 2019, 499, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Prati, A.; Genazzani, A.R.; Genazzani, A.D. Pathogenesis of PCOS: From metabolic and neuroendocrine implications to the choice of the theraputic strategy. In Clinical Management of Infertility; Springer: Berlin/Heidelberg, Germany, 2021; Volume 8, pp. 131–154. [Google Scholar]
- Hill, J.W.; Elmquist, J.K.; Elias, C.F. Hypothalamic pathways linking energy balance and reproduction. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E827–E832. [Google Scholar] [CrossRef] [PubMed]
- Eyvazzadeh, A.D.; Pennington, K.P.; Pop-Busui, R.; Sowers, M.; Zubieta, J.-K.; Smith, Y.R. The role of the endogenous opioid system in polycystic ovary syndrome. Fertil. Steril. 2009, 92, 1–12. [Google Scholar] [CrossRef]
- Genazzani, A.D.; Podfigurna, A.; Szeliga, A.; Meczekalski, B. Kisspeptin in female reproduction: From physiology to pathophysiology. Gynecol. Reprod. Endocrinol. Metab. 2021, 2, 148–155. [Google Scholar]
- Genazzani, A.D. Inositol as putative integrative treatment for PCOS. Reprod. Biomed. Online 2016, 33, 770–780. [Google Scholar] [CrossRef]
- Chen, C.-I.; Hsu, M.-I.; Lin, S.-H.; Chang, Y.-C.I.; Hsu, C.-S.; Tzeng, C.-R. Adiponectin and leptin in overweight/obese and lean women with polycystic ovary syndrome. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2015, 31, 264–268. [Google Scholar] [CrossRef]
- Genazzani, A.D. Inositoli: È tempo di capire per sapere poi come fare. Boll. Ginecol. Endocrinol. 2021, 15, 9–12. [Google Scholar]
- Mohammad, M.B.; Seghinsara, A.M. Polycystic Ovary Syndrome (PCOS), Diagnostic Criteria, and AMH. Asian Pac. J. Cancer Prev. APJCP 2017, 18, 17–21. [Google Scholar] [CrossRef]
- Fleming, R.; Harborne, L.; MacLaughlin, D.T.; Ling, D.; Norman, J.; Sattar, N.; Seifer, D.B. Metformin reduces serum mullerian-inhibiting substance levels in women with polycystic ovary syndrome after protracted treatment. Fertil. Steril. 2005, 83, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Dumesic, D.A.; Oberfield, S.E.; Stener-Victorin, E.; Marshall, J.C.; Laven, J.S.; Legro, R.S. Scientific Statement on the Diagnostic Criteria, Epidemiology, Pathophysiology, and Molecular Genetics of Polycystic Ovary Syndrome. Endocr. Rev. 2015, 36, 487–525. [Google Scholar] [CrossRef] [PubMed]
- Azziz, R.; Carmina, E.; Chen, Z.; Dunaif, A.; Laven, J.S.E.; Legro, R.S.; Lizneva, D.; Natterson-Horowtiz, B.; Teede, H.J.; Yildiz, B.O. Polycystic ovary syndrome. Nat. Rev. Dis. Primer 2016, 2, 16057. [Google Scholar] [CrossRef] [PubMed]
- Ciampelli, M.; Fulghesu, A.M.; Cucinelli, F.; Pavone, V.; Ronsisvalle, E.; Guido, M.; Caruso, A.; Lanzone, A. Impact of insulin and body mass index on metabolic and endocrine variables in polycystic ovary syndrome. Metabolism 1999, 48, 167–172. [Google Scholar] [CrossRef]
- Ciaraldi, T.P.; el-Roeiy, A.; Madar, Z.; Reichart, D.; Olefsky, J.M.; Yen, S.S. Cellular mechanisms of insulin resistance in polycystic ovarian syndrome. J. Clin. Endocrinol. Metab. 1992, 75, 577–583. [Google Scholar] [CrossRef]
- Rosenbaum, D.; Haber, R.S.; Dunaif, A. Insulin resistance in polycystic ovary syndrome: Decreased expression of GLUT-4 glucose transporters in adipocytes. Am. J. Physiol. 1993, 264, E197–E202. [Google Scholar] [CrossRef]
- Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults. Executive Summary of The Third Report of The National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, And Treatment of High Blood Cholesterol In Adults (Adult Treatment Panel III). JAMA 2001, 285, 2486–2497. [Google Scholar] [CrossRef]
- ACOG. Practice Bulletin No. 108: Polycystic ovary syndrome. Obstet. Gynecol. 2009, 114, 936. [Google Scholar] [CrossRef]
- Checa, M.A.; Requena, A.; Salvador, C.; Tur, R.; Callejo, J.; Espinós, J.J.; Fábregues, F.; Herrero, J. Reproductive Endocrinology Interest Group of the Spanish Society of Fertility Insulin-sensitizing agents: Use in pregnancy and as therapy in polycystic ovary syndrome. Hum. Reprod. Update 2005, 11, 375–390. [Google Scholar] [CrossRef] [PubMed]
- Genazzani, A.D.; Prati, A.; Santagni, S.; Ricchieri, F.; Chierchia, E.; Rattighieri, E.; Campedelli, A.; Simoncini, T.; Artini, P.G. Differential insulin response to myo-inositol administration in obese polycystic ovary syndrome patients. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2012, 28, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Madeira, I.R.; Carvalho, C.N.M.; Gazolla, F.M.; de Matos, H.J.; Borges, M.A.; Bordallo, M.A.N. Cut-off point for Homeostatic Model Assessment for Insulin Resistance (HOMA-IR) index established from Receiver Operating Characteristic (ROC) curve in the detection of metabolic syndrome in overweight pre-pubertal children. Arq. Bras. Endocrinol. Metabol. 2008, 52, 1466–1473. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ciampelli, M.; Leoni, F.; Cucinelli, F.; Mancuso, S.; Panunzi, S.; De Gaetano, A.; Lanzone, A. Assessment of insulin sensitivity from measurements in the fasting state and during an oral glucose tolerance test in polycystic ovary syndrome and menopausal patients. J. Clin. Endocrinol. Metab. 2005, 90, 1398–1406. [Google Scholar] [CrossRef] [PubMed]
- Legro, R.S.; Kunselman, A.R.; Dunaif, A. Prevalence and predictors of dyslipidemia in women with polycystic ovary syndrome. Am. J. Med. 2001, 111, 607–613. [Google Scholar] [CrossRef]
- Shi, Y.; Cui, Y.; Sun, X.; Ma, G.; Ma, Z.; Gao, Q.; Chen, Z.-J. Hypertension in women with polycystic ovary syndrome: Prevalence and associated cardiovascular risk factors. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 173, 66–70. [Google Scholar] [CrossRef]
- Muniyappa, R.; Montagnani, M.; Koh, K.K.; Quon, M.J. Cardiovascular actions of insulin. Endocr. Rev. 2007, 28, 463–491. [Google Scholar] [CrossRef]
- Goverde, A.J.; van Koert, A.J.B.; Eijkemans, M.J.; Knauff, E.a.H.; Westerveld, H.E.; Fauser, B.C.J.M.; Broekmans, F.J. Indicators for metabolic disturbances in anovulatory women with polycystic ovary syndrome diagnosed according to the Rotterdam consensus criteria. Hum. Reprod. Oxf. Engl. 2009, 24, 710–717. [Google Scholar] [CrossRef]
- Farhadi-Azar, M.; Behboudi-Gandevani, S.; Rahmati, M.; Mahboobifard, F.; Khalili Pouya, E.; Ramezani Tehrani, F.; Azizi, F. The Prevalence of Polycystic Ovary Syndrome, Its Phenotypes and Cardio-Metabolic Features in a Community Sample of Iranian Population: Tehran Lipid and Glucose Study. Front. Endocrinol. 2022, 13, 825528. [Google Scholar] [CrossRef]
- Utzschneider, K.M.; Kahn, S.E.; Polidori, D.C. Hepatic Insulin Extraction in NAFLD Is Related to Insulin Resistance Rather Than Liver Fat Content. J. Clin. Endocrinol. Metab. 2018, 104, 1855–1865. [Google Scholar] [CrossRef]
- Genazzani, A.D.; Battipaglia, C.; Petrillo, T.; Alberti, C.; Patrizi, B.; Tomatis, V.; Simoncini, T.; Genazzani, A.R. HIE (hepatic insulin extraction) index in overweight/obese. PCOS patients with or without familial diabetes. Gynecol. Reprod. Endocrinol. Metab. 2022, 1, 57–68. [Google Scholar]
- Fosam, A.; Sikder, S.; Abel, B.S.; Tella, S.H.; Walter, M.F.; Mari, A.; Muniyappa, R. Reduced Insulin Clearance and Insulin-Degrading Enzyme Activity Contribute to Hyperinsulinemia in African Americans. J. Clin. Endocrinol. Metab. 2020, 105, dgaa070. [Google Scholar] [CrossRef] [PubMed]
- Leissring, M.A.; González-Casimiro, C.M.; Merino, B.; Suire, C.N.; Perdomo, G. Targeting Insulin-Degrading Enzyme in Insulin Clearance. Int. J. Mol. Sci. 2021, 22, 2235. [Google Scholar] [CrossRef] [PubMed]
- Kiddy, D.S.; Hamilton-Fairley, D.; Bush, A.; Short, F.; Anyaoku, V.; Reed, M.J.; Franks, S. Improvement in endocrine and ovarian function during dietary treatment of obese women with polycystic ovary syndrome. Clin. Endocrinol. 1992, 36, 105–111. [Google Scholar] [CrossRef]
- Schindler, A.E. Non-Contraceptive Benefits of Oral Hormonal Contraceptives. Int. J. Endocrinol. Metab. 2013, 11, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Schindler, A.E. Non-contraceptive use of hormonal contraceptives. Gynecol. Endocrinol. 2008, 24, 235–236. [Google Scholar] [CrossRef]
- Goodman, N.F.; Cobin, R.H.; Futterweit, W.; Glueck, J.S.; Legro, R.S.; Carmina, E. American Association of Clinical Endocrinologists (AACE); American College of Endocrinology (ACE); Androgen Excess and PCOS Society (AES) american association of clinical endocrinologists, american college of endocrinology, and androgen excess and pcos society disease state clinical review: Guide to the best practices in the evaluation and treatment of polycystic ovary syndrome—part 1. Endocr. Pract. Off. J. Am. Coll. Endocrinol. Am. Assoc. Clin. Endocrinol. 2015, 21, 1291–1300. [Google Scholar] [CrossRef]
- Paradisi, R.; Fabbri, R.; Battaglia, C.; Venturoli, S. Ovulatory effects of flutamide in the polycystic ovary syndrome. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2013, 29, 391–395. [Google Scholar] [CrossRef]
- Azziz, R.; Ehrmann, D.; Legro, R.S.; Whitcomb, R.W.; Hanley, R.; Fereshetian, A.G.; O’Keefe, M.; Ghazzi, M.N.; PCOS/Troglitazone Study Group. Troglitazone improves ovulation and hirsutism in the polycystic ovary syndrome: A multicenter, double blind, placebo-controlled trial. J. Clin. Endocrinol. Metab. 2001, 86, 1626–1632. [Google Scholar] [CrossRef]
- Ibáñez, L.; Valls, C.; Ferrer, A.; Marcos, M.V.; Rodriguez-Hierro, F.; de Zegher, F. Sensitization to insulin induces ovulation in nonobese adolescents with anovulatory hyperandrogenism. J. Clin. Endocrinol. Metab. 2001, 86, 3595–3598. [Google Scholar] [CrossRef]
- Nestler, J.E.; Jakubowicz, D.J. Decreases in ovarian cytochrome P450c17 alpha activity and serum free testosterone after reduction of insulin secretion in polycystic ovary syndrome. N. Engl. J. Med. 1996, 335, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Kriplani, A.; Agarwal, N. Effects of metformin on clinical and biochemical parameters in polycystic ovary syndrome. J. Reprod. Med. 2004, 49, 361–367. [Google Scholar] [PubMed]
- Hu, L.; Shen, H.; Wu, Q.F.; Tian, L.; Hu, M.H. Treatment of polycystic ovarian syndrome with insulin resistance by insulin-sensitizer. Clin. Exp. Obstet. Gynecol. 2014, 41, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Morley, L.C.; Tang, T.; Yasmin, E.; Norman, R.J.; Balen, A.H. Insulin-sensitising drugs (metformin, rosiglitazone, pioglitazone, D-chiro-inositol) for women with polycystic ovary syndrome, oligo amenorrhoea and subfertility. Cochrane Database Syst. Rev. 2017, 11, CD003053. [Google Scholar] [CrossRef] [PubMed]
- Thessaloniki ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group. Consensus on infertility treatment related to polycystic ovary syndrome. Fertil. Steril. 2008, 89, 505–522. [Google Scholar] [CrossRef]
- Seow, K.-M.; Juan, C.-C.; Hwang, J.-L.; Ho, L.-T. Laparoscopic surgery in polycystic ovary syndrome: Reproductive and metabolic effects. Semin. Reprod. Med. 2008, 26, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Heimark, D.B.; Nguygen, T.; Nadler, J.L.; Larner, J. Both myo-inositol to chiro-inositol epimerase activities and chiro-inositol to myo-inositol ratios are decreased in tissues of GK type 2 diabetic rats compared to Wistar controls. Biochem. Biophys. Res. Commun. 2002, 293, 1092–1098. [Google Scholar] [CrossRef]
- Büttner, J. Johann Joseph von Scherer (1814-69). The early history of clinical chemistry. J. Clin. Chem. Clin. Biochem. Z. Klin. Chem. Klin. Biochem. 1978, 16, 478–483. [Google Scholar]
- Thomas, R.M.; Nechamen, C.A.; Mazurkiewicz, J.E.; Ulloa-Aguirre, A.; Dias, J.A. The adapter protein APPL1 links FSH receptor to inositol 1,4,5-trisphosphate production and is implicated in intracellular Ca(2+) mobilization. Endocrinology 2011, 152, 1691–1701. [Google Scholar] [CrossRef]
- Unfer, V.; Proietti, S.; Gullo, G.; Porcaro, G.; Carlomagno, G.; Bizzarri, M. Polycystic Ovary Syndrome: Features, Diagnostic Criteria and Treatments. Endocrinol. Metab. Syndr. 2014, 3, 1000136. [Google Scholar] [CrossRef]
- Croze, M.L.; Soulage, C.O. Potential role and therapeutic interests of myo-inositol in metabolic diseases. Biochimie 2013, 95, 1811–1827. [Google Scholar] [CrossRef] [PubMed]
- Kennington, A.; Hill, C.; Craig, J.; Bogardus, C.; Raz, I.; Ortmeyer, H.; Hansen, B.; Romero, G.; Larner, J. Low urinary chiro-inositol excretion in non-insulin-dependent diabetes mellitus. N. Engl. J. Med. 1990, 323, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Baillargeon, J.-P.; Diamanti-Kandarakis, E.; Ostlund, R.E.; Apridonidze, T.; Iuorno, M.J.; Nestler, J.E. Altered D-chiro-inositol urinary clearance in women with polycystic ovary syndrome. Diabetes Care 2006, 29, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Larner, J.; Brautigan, D.L.; Thorner, M.O. D-chiro-inositol glycans in insulin signaling and insulin resistance. Mol. Med. Camb. Mass 2010, 16, 543–552. [Google Scholar] [CrossRef]
- Larner, J.; Craig, J.W. Urinary myo-inositol-to-chiro-inositol ratios and insulin resistance. Diabetes Care 1996, 19, 76–78. [Google Scholar] [CrossRef]
- Genazzani, A.D.; Santagni, S.; Rattighieri, E.; Chierchia, E.; Despini, G.; Marini, G.; Prati, A.; Simoncini, T. Modulatory role of D-chiro-inositol (DCI) on LH and insulin secretion in obese PCOS patients. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2014, 30, 438–443. [Google Scholar] [CrossRef]
- La Marca, A.; Grisendi, V.; Dondi, G.; Sighinolfi, G.; Cianci, A. The menstrual cycle regularization following D-chiro-inositol treatment in PCOS women: A retrospective study. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2015, 31, 52–56. [Google Scholar] [CrossRef]
- Chiu, T.T.Y.; Rogers, M.S.; Law, E.L.K.; Briton-Jones, C.M.; Cheung, L.P.; Haines, C.J. Follicular fluid and serum concentrations of myo-inositol in patients undergoing IVF: Relationship with oocyte quality. Hum. Reprod. Oxf. Engl. 2002, 17, 1591–1596. [Google Scholar] [CrossRef]
- Chiu, T.T.Y.; Rogers, M.S.; Briton-Jones, C.; Haines, C. Effects of myo-inositol on the in-vitro maturation and subsequent development of mouse oocytes. Hum. Reprod. Oxf. Engl. 2003, 18, 408–416. [Google Scholar] [CrossRef]
- Artini, P.G.; Di Berardino, O.M.; Papini, F.; Genazzani, A.D.; Simi, G.; Ruggiero, M.; Cela, V. Endocrine and clinical effects of myo-inositol administration in polycystic ovary syndrome. A randomized study. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2013, 29, 375–379. [Google Scholar] [CrossRef]
- Kamenov, Z.; Kolarov, G.; Gateva, A.; Carlomagno, G.; Genazzani, A.D. Ovulation induction with myo-inositol alone and in combination with clomiphene citrate in polycystic ovarian syndrome patients with insulin resistance. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2015, 31, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Wdowiak, A. Myoinositol Improves Embryo Development in PCOS Patients Undergoing ICSI. Int. J. Endocrinol. 2016, 2016, 6273298. [Google Scholar] [CrossRef] [PubMed]
- Vartanyan, E.V.; Tsaturova, K.A.; Devyatova, E.A.; Mikhaylyukova, A.S.; Levin, V.A.; Petuhova, N.L.; Markin, A.V.; Steptsova, E.M. Improvement in quality of oocytes in polycystic ovarian syndrome in programs of in vitro fertilization. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2017, 33, 8–11. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Emekçi Özay, Ö.; Özay, A.C.; Çağlıyan, E.; Okyay, R.E.; Gülekli, B. Myo-inositol administration positively effects ovulation induction and intrauterine insemination in patients with polycystic ovary syndrome: A prospective, controlled, randomized trial. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2017, 33, 524–528. [Google Scholar] [CrossRef]
- Dinicola, S.; Unfer, V.; Facchinetti, F.; Soulage, C.O.; Greene, N.D.; Bizzarri, M.; Laganà, A.S.; Chan, S.-Y.; Bevilacqua, A.; Pkhaladze, L.; et al. Inositols: From Established Knowledge to Novel Approaches. Int. J. Mol. Sci. 2021, 22, 10575. [Google Scholar] [CrossRef]
- Laganà, A.S.; Vitagliano, A.; Noventa, M.; Ambrosini, G.; D’Anna, R. Myo-inositol supplementation reduces the amount of gonadotropins and length of ovarian stimulation in women undergoing IVF: A systematic review and meta-analysis of randomized controlled trials. Arch. Gynecol. Obstet. 2018, 298, 675–684. [Google Scholar] [CrossRef]
- Zheng, X.; Lin, D.; Zhang, Y.; Lin, Y.; Song, J.; Li, S.; Sun, Y. Inositol supplement improves clinical pregnancy rate in infertile women undergoing ovulation induction for ICSI or IVF-ET. Medicine 2017, 96, e8842. [Google Scholar] [CrossRef]
- Unfer, V.; Carlomagno, G.; Dante, G.; Facchinetti, F. Effects of myo-inositol in women with PCOS: A systematic review of randomized controlled trials. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2012, 28, 509–515. [Google Scholar] [CrossRef]
- Carlomagno, G.; Unfer, V.; Roseff, S. The D-chiro-inositol paradox in the ovary. Fertil. Steril. 2011, 95, 2515–2516. [Google Scholar] [CrossRef]
- Unfer, V.; Carlomagno, G.; Papaleo, E.; Vailati, S.; Candiani, M.; Baillargeon, J.-P. Hyperinsulinemia Alters Myoinositol to d-chiroinositol Ratio in the Follicular Fluid of Patients With PCOS. Reprod. Sci. Thousand Oaks Calif 2014, 21, 854–858. [Google Scholar] [CrossRef]
- Heimark, D.; McAllister, J.; Larner, J. Decreased myo-inositol to chiro-inositol (M/C) ratios and increased M/C epimerase activity in PCOS theca cells demonstrate increased insulin sensitivity compared to controls. Endocr. J. 2014, 61, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Sacchi, S.; Marinaro, F.; Tondelli, D.; Lui, J.; Xella, S.; Marsella, T.; Tagliasacchi, D.; Argento, C.; Tirelli, A.; Giulini, S.; et al. Modulation of gonadotrophin induced steroidogenic enzymes in granulosa cells by d-chiroinositol. Reprod. Biol. Endocrinol. RBE 2016, 14, 52. [Google Scholar] [CrossRef] [PubMed]
- Gambioli, R.; Forte, G.; Aragona, C.; Bevilacqua, A.; Bizzarri, M.; Unfer, V. The use of D-chiro-Inositol in clinical practice. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Monastra, G.; Vucenik, I.; Harrath, A.H.; Alwasel, S.H.; Kamenov, Z.A.; Laganà, A.S.; Monti, N.; Fedeli, V.; Bizzarri, M. PCOS and Inositols: Controversial Results and Necessary Clarifications. Basic Differences Between D-Chiro and Myo-Inositol. Front. Endocrinol. 2021, 12, 660381. [Google Scholar] [CrossRef] [PubMed]
- Cheang, K.I.; Baillargeon, J.-P.; Essah, P.A.; Ostlund, R.E.; Apridonize, T.; Islam, L.; Nestler, J.E. Insulin-stimulated release of D-chiro-inositol-containing inositolphosphoglycan mediator correlates with insulin sensitivity in women with polycystic ovary syndrome. Metabolism 2008, 57, 1390–1397. [Google Scholar] [CrossRef]
- Ravanos, K.; Monastra, G.; Pavlidou, T.; Goudakou, M.; Prapas, N. Can high levels of D-chiro-inositol in follicular fluid exert detrimental effects on blastocyst quality? Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 5491–5498. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Dragotto, J.; Giuliani, A.; Bizzarri, M. Myo-inositol and D-chiro-inositol (40:1) reverse histological and functional features of polycystic ovary syndrome in a mouse model. J. Cell. Physiol. 2019, 234, 9387–9398. [Google Scholar] [CrossRef]
- Nordio, M.; Basciani, S.; Camajani, E. The 40:1 myo-inositol/D-chiro-inositol plasma ratio is able to restore ovulation in PCOS patients: Comparison with other ratios. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5512–5521. [Google Scholar] [CrossRef]
- Roseff, S.; Montenegro, M. Inositol Treatment for PCOS Should Be Science-Based and Not Arbitrary. Int. J. Endocrinol. 2020, 2020, 6461254. [Google Scholar] [CrossRef]
- Reed, L.J. From lipoic acid to multi-enzyme complexes. Protein Sci. Publ. Protein Soc. 1998, 7, 220–224. [Google Scholar] [CrossRef]
- Padmalayam, I.; Hasham, S.; Saxena, U.; Pillarisetti, S. Lipoic acid synthase (LASY): A novel role in inflammation, mitochondrial function, and insulin resistance. Diabetes 2009, 58, 600–608. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Packer, L.; Witt, E.H.; Tritschler, H.J. alpha-Lipoic acid as a biological antioxidant. Free Radic. Biol. Med. 1995, 19, 227–250. [Google Scholar] [CrossRef]
- Golbidi, S.; Badran, M.; Laher, I. Diabetes and alpha lipoic Acid. Front. Pharmacol. 2011, 2, 69. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.B.; Negrato, C.A. Alpha-lipoic acid as a pleiotropic compound with potential therapeutic use in diabetes and other chronic diseases. Diabetol. Metab. Syndr. 2014, 6, 80. [Google Scholar] [CrossRef] [PubMed]
- Scaramuzza, A.; Giani, E.; Redaelli, F.; Ungheri, S.; Macedoni, M.; Giudici, V.; Bosetti, A.; Ferrari, M.; Zuccotti, G.V. Alpha-Lipoic Acid and Antioxidant Diet Help to Improve Endothelial Dysfunction in Adolescents with Type 1 Diabetes: A Pilot Trial. J. Diabetes Res. 2015, 2015, 474561. [Google Scholar] [CrossRef]
- Hardie, D.G.; Hawley, S.A. AMP-activated protein kinase: The energy charge hypothesis revisited. BioEssays News Rev. Mol. Cell. Dev. Biol. 2001, 23, 1112–1119. [Google Scholar] [CrossRef] [PubMed]
- Konrad, D.; Somwar, R.; Sweeney, G.; Yaworsky, K.; Hayashi, M.; Ramlal, T.; Klip, A. The antihyperglycemic drug alpha-lipoic acid stimulates glucose uptake via both GLUT4 translocation and GLUT4 activation: Potential role of p38 mitogen-activated protein kinase in GLUT4 activation. Diabetes 2001, 50, 1464–1471. [Google Scholar] [CrossRef]
- Lee, W.J.; Song, K.-H.; Koh, E.H.; Won, J.C.; Kim, H.S.; Park, H.-S.; Kim, M.-S.; Kim, S.-W.; Lee, K.-U.; Park, J.-Y. Alpha-lipoic acid increases insulin sensitivity by activating AMPK in skeletal muscle. Biochem. Biophys. Res. Commun. 2005, 332, 885–891. [Google Scholar] [CrossRef]
- Evans, J.L.; Goldfine, I.D.; Maddux, B.A.; Grodsky, G.M. Oxidative stress and stress-activated signaling pathways: A unifying hypothesis of type 2 diabetes. Endocr. Rev. 2002, 23, 599–622. [Google Scholar] [CrossRef]
- Jacob, S.; Ruus, P.; Hermann, R.; Tritschler, H.J.; Maerker, E.; Renn, W.; Augustin, H.J.; Dietze, G.J.; Rett, K. Oral administration of RAC-alpha-lipoic acid modulates insulin sensitivity in patients with type-2 diabetes mellitus: A placebo-controlled pilot trial. Free Radic. Biol. Med. 1999, 27, 309–314. [Google Scholar] [CrossRef]
- Jacob, S.; Henriksen, E.J.; Schiemann, A.L.; Simon, I.; Clancy, D.E.; Tritschler, H.J.; Jung, W.I.; Augustin, H.J.; Dietze, G.J. Enhancement of glucose disposal in patients with type 2 diabetes by alpha-lipoic acid. Arzneimittelforschung 1995, 45, 872–874. [Google Scholar] [PubMed]
- González, F.; Rote, N.S.; Minium, J.; Kirwan, J.P. Reactive oxygen species-induced oxidative stress in the development of insulin resistance and hyperandrogenism in polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2006, 91, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Genazzani, A.D.; Shefer, K.; Della Casa, D.; Prati, A.; Napolitano, A.; Manzo, A.; Despini, G.; Simoncini, T. Modulatory effects of alpha-lipoic acid (ALA) administration on insulin sensitivity in obese PCOS patients. J. Endocrinol. Investig. 2018, 41, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, W.; Liu, Y.; Li, Y.; Gao, L.; Zhao, J. Alpha-lipoic acid attenuates insulin resistance and improves glucose metabolism in high fat diet-fed mice. Acta Pharmacol. Sin. 2014, 35, 1285–1292. [Google Scholar] [CrossRef] [PubMed]
- Macut, D.; Božić-Antić, I.; Bjekić-Macut, J.; Tziomalos, K. Management of endocrine disease: Polycystic ovary syndrome and nonalcoholic fatty liver disease. Eur. J. Endocrinol. 2017, 177, R145–R158. [Google Scholar] [CrossRef]
- Genazzani, A. Effects of a Combination of Alpha Lipoic Acid and Myo-Inositol on Insulin Dynamics in Overweight/Obese Patients with PCOS. Endocrinol. Metab. Syndr. 2014, 3, 3. [Google Scholar] [CrossRef]
- De Cicco, S.; Immediata, V.; Romualdi, D.; Policola, C.; Tropea, A.; Di Florio, C.; Tagliaferri, V.; Scarinci, E.; Della Casa, S.; Lanzone, A.; et al. Myoinositol combined with alpha-lipoic acid may improve the clinical and endocrine features of polycystic ovary syndrome through an insulin-independent action. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2017, 33, 698–701. [Google Scholar] [CrossRef]
- Genazzani, A.D.; Prati, A.; Simoncini, T.; Napolitano, A. Modulatory role of D-chiro-inositol and alpha lipoic acid combination on hormonal and metabolic parameters of overweight/obese PCOS patients. Eur. Gynecol. Obstet. 2019, 1, 29–33. [Google Scholar]
- Cianci, A.; Panella, M.; Fichera, M.; Falduzzi, C.; Bartolo, M.; Caruso, S. d-chiro-Inositol and alpha lipoic acid treatment of metabolic and menses disorders in women with PCOS. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2015, 31, 483–486. [Google Scholar] [CrossRef]
- Genazzani, A. Expert’s opinion: Integrative treatment with inositols and lipoic acid for insulin resistance of PCOS. Gynecol. Reprod. Endocrinol. Metab. 2020, 3, 146–157. [Google Scholar]
- Laganà, A.S.; Monti, N.; Fedeli, V.; Gullo, G.; Bizzarri, M. Does Alpha-lipoic acid improve effects on polycystic ovary syndrome? Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Genazzani, A. Inositols: Reflections on how to choose the appropriate one for PCOS. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2020, 36, 1045–1046. [Google Scholar] [CrossRef] [PubMed]
- Pekala, J.; Patkowska-Sokoła, B.; Bodkowski, R.; Jamroz, D.; Nowakowski, P.; Lochyński, S.; Librowski, T. L-carnitine--metabolic functions and meaning in humans life. Curr. Drug Metab. 2011, 12, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Vogt, C.; Georgi, A.; Werner, G. Enantiomeric separation of D/L-carnitine using HPLC and CZE after derivatization. Chromatographia 1995, 40, 287–295. [Google Scholar] [CrossRef]
- Chalmers, R.A.; Roe, C.R.; Stacey, T.E.; Hoppel, C.L. Urinary excretion of l-carnitine and acylcarnitines by patients with disorders of organic acid metabolism: Evidence for secondary insufficiency of l-carnitine. Pediatr. Res. 1984, 18, 1325–1328. [Google Scholar] [CrossRef]
- Seline, K.-G.; Johein, H. The determination of l-carnitine in several food samples. Food Chem. 2007, 105, 793–804. [Google Scholar] [CrossRef]
- Lopaschuk, G.D. Carnitine and Myocardial Glucose Metabolism. In Carnitine Today; De Simone, C., Famularo, G., Eds.; Molecular Biology Intelligence Unit; Springer US: Boston, MA, USA, 1997; pp. 71–93. ISBN 978-1-4615-6005-0. [Google Scholar]
- Kerbey, A.L.; Vary, T.C.; Randle, P.J. Molecular mechanisms regulating myocardial glucose oxidation. Basic Res. Cardiol. 1985, 80 (Suppl. 2), 93–96. [Google Scholar]
- Agarwal, A.; Sengupta, P.; Durairajanayagam, D. Role of L-carnitine in female infertility. Reprod. Biol. Endocrinol. RBE 2018, 16, 5. [Google Scholar] [CrossRef]
- Petrillo, T.; Battipaglia, C.; Virmani, M.A.; Genazzani, A.R.; Genazzani, A.D. Neuroendocrine Effects of Carnitines on Reproductive Impairments. Int. J. Mol. Sci. 2021, 22, 10781. [Google Scholar] [CrossRef]
- Malaguarnera, M.; Vacante, M.; Avitabile, T.; Malaguarnera, M.; Cammalleri, L.; Motta, M. L-Carnitine supplementation reduces oxidized LDL cholesterol in patients with diabetes. Am. J. Clin. Nutr. 2009, 89, 71–76. [Google Scholar] [CrossRef]
- Genazzani, A.D.; Prati, A.; Genazzani, A.R.; Battipaglia, C.; Simoncini, T.; Szeliga, A.; Podfigurna, A.; Meczekalski, B. Synergistic effects of the integrative administration of acetyl-L-carnitine, L-carnitine, L-arginine and N-acetyl-cysteine on metabolic dynamics and on hepatic insulin extraction in overweight/obese patients with PCOS. Gynecol. Reprod. Endocrinol. Metab 2020, 1, 56–63. [Google Scholar]
- Fenkci, S.M.; Fenkci, V.; Oztekin, O.; Rota, S.; Karagenc, N. Serum total L-carnitine levels in non-obese women with polycystic ovary syndrome. Hum. Reprod. Oxf. Engl. 2008, 23, 1602–1606. [Google Scholar] [CrossRef] [PubMed]
- Molfino, A.; Cascino, A.; Conte, C.; Ramaccini, C.; Rossi Fanelli, F.; Laviano, A. Caloric restriction and L-carnitine administration improves insulin sensitivity in patients with impaired glucose metabolism. JPEN J. Parenter. Enteral Nutr. 2010, 34, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Petersen, K.F.; Befroy, D.; Dufour, S.; Dziura, J.; Ariyan, C.; Rothman, D.L.; DiPietro, L.; Cline, G.W.; Shulman, G.I. Mitochondrial dysfunction in the elderly: Possible role in insulin resistance. Science 2003, 300, 1140–1142. [Google Scholar] [CrossRef] [PubMed]
- Manco, M.; Calvani, M.; Mingrone, G. Effects of dietary fatty acids on insulin sensitivity and secretion. Diabetes Obes. Metab. 2004, 6, 402–413. [Google Scholar] [CrossRef]
- Samimi, M.; Jamilian, M.; Ebrahimi, F.A.; Rahimi, M.; Tajbakhsh, B.; Asemi, Z. Oral carnitine supplementation reduces body weight and insulin resistance in women with polycystic ovary syndrome: A randomized, double-blind, placebo-controlled trial. Clin. Endocrinol. Oxf. 2016, 84, 851–857. [Google Scholar] [CrossRef]
- Oka, S.; Kamata, H.; Kamata, K.; Yagisawa, H.; Hirata, H. N-acetylcysteine suppresses TNF-induced NF-kappaB activation through inhibition of IkappaB kinases. FEBS Lett. 2000, 472, 196–202. [Google Scholar] [CrossRef]
- Ammon, H.P.; Müller, P.H.; Eggstein, M.; Wintermantel, C.; Aigner, B.; Safayhi, H.; Stützle, M.; Renn, W. Increase in glucose consumption by acetylcysteine during hyperglycemic clamp. A study with healthy volunteers. Arzneimittelforschung 1992, 42, 642–645. [Google Scholar]
- Fulghesu, A.M.; Ciampelli, M.; Muzj, G.; Belosi, C.; Selvaggi, L.; Ayala, G.F.; Lanzone, A. N-acetyl-cysteine treatment improves insulin sensitivity in women with polycystic ovary syndrome. Fertil. Steril. 2002, 77, 1128–1135. [Google Scholar] [CrossRef]
- Nácul, A.P.; Andrade, C.D.; Schwarz, P.; de Bittencourt, P.I.H.; Spritzer, P.M. Nitric oxide and fibrinogen in polycystic ovary syndrome: Associations with insulin resistance and obesity. Eur. J. Obstet. Gynecol. Reprod. Biol. 2007, 133, 191–196. [Google Scholar] [CrossRef]
- Masha, A.; Manieri, C.; Dinatale, S.; Bruno, G.A.; Ghigo, E.; Martina, V. Prolonged treatment with N-acetylcysteine and L-arginine restores gonadal function in patients with polycystic ovary syndrome. J. Endocrinol. Investig. 2009, 32, 870–872. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, J.K.; Waqar, A.; Jain, A.; Joseph, C.; Srivastava, K.; Ochuba, O.; Alkayyali, T.; Ruo, S.W.; Poudel, S. Oxidative Stress in Polycystic Ovarian Syndrome and the Effect of Antioxidant N-Acetylcysteine on Ovulation and Pregnancy Rate. Cureus 2021, 13, e17887. [Google Scholar] [CrossRef]
- Krishna, M.B.; Joseph, A.; Thomas, P.L.; Dsilva, B.; Pillai, S.M.; Laloraya, M. Impaired Arginine Metabolism Coupled to a Defective Redox Conduit Contributes to Low Plasma Nitric Oxide in Polycystic Ovary Syndrome. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 43, 1880–1892. [Google Scholar] [CrossRef] [PubMed]
- Yousefi Rad, E.; Nazarian, B.; Saboori, S.; Falahi, E.; Hekmatdoost, A. Effects of l-arginine supplementation on glycemic profile: Evidence from a systematic review and meta-analysis of clinical trials. J. Integr. Med. 2020, 18, 284–291. [Google Scholar] [CrossRef]
- Goradel, N.H.; Asghari, M.H.; Moloudizargari, M.; Negahdari, B.; Haghi-Aminjan, H.; Abdollahi, M. Melatonin as an angiogenesis inhibitor to combat cancer: Mechanistic evidence. Toxicol. Appl. Pharmacol. 2017, 335, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.; Jain, M.; Haldar, C.; Singh, T.B.; Jain, S. Melatonin and its correlation with testosterone in polycystic ovarian syndrome. J. Hum. Reprod. Sci. 2013, 6, 253–258. [Google Scholar] [CrossRef]
- Li, H.; Liu, M.; Zhang, C. Women with polycystic ovary syndrome (PCOS) have reduced melatonin concentrations in their follicles and have mild sleep disturbances. BMC Womens Health 2022, 22, 79. [Google Scholar] [CrossRef]
- Tamura, H.; Nakamura, Y.; Korkmaz, A.; Manchester, L.C.; Tan, D.-X.; Sugino, N.; Reiter, R.J. Melatonin and the ovary: Physiological and pathophysiological implications. Fertil. Steril. 2009, 92, 328–343. [Google Scholar] [CrossRef]
- Pacchiarotti, A.; Carlomagno, G.; Antonini, G.; Pacchiarotti, A. Effect of myo-inositol and melatonin versus myo-inositol, in a randomized controlled trial, for improving in vitro fertilization of patients with polycystic ovarian syndrome. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2016, 32, 69–73. [Google Scholar] [CrossRef]
- Mojaverrostami, S.; Asghari, N.; Khamisabadi, M.; Heidari Khoei, H. The role of melatonin in polycystic ovary syndrome: A review. Int. J. Reprod. Biomed. 2019, 17, 865–882. [Google Scholar] [CrossRef]
- Tagliaferri, V.; Romualdi, D.; Scarinci, E.; Cicco, S.D.; Florio, C.D.; Immediata, V.; Tropea, A.; Santarsiero, C.M.; Lanzone, A.; Apa, R. Melatonin Treatment May Be Able to Restore Menstrual Cyclicity in Women With PCOS: A Pilot Study. Reprod. Sci. Thousand Oaks Calif. 2018, 25, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Pai, S.A.; Majumdar, A.S. Protective effects of melatonin against metabolic and reproductive disturbances in polycystic ovary syndrome in rats. J. Pharm. Pharmacol. 2014, 66, 1710–1721. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Shi, H.; Liu, Y.; Zhao, S.; Zhao, H. Applications of Melatonin in Female Reproduction in the Context of Oxidative Stress. Oxid. Med. Cell. Longev. 2021, 2021, 6668365. [Google Scholar] [CrossRef]
- Jamilian, M.; Foroozanfard, F.; Mirhosseini, N.; Kavossian, E.; Aghadavod, E.; Bahmani, F.; Ostadmohammadi, V.; Kia, M.; Eftekhar, T.; Ayati, E.; et al. Effects of Melatonin Supplementation on Hormonal, Inflammatory, Genetic, and Oxidative Stress Parameters in Women With Polycystic Ovary Syndrome. Front. Endocrinol. 2019, 10, 273. [Google Scholar] [CrossRef] [PubMed]
- Shabani, A.; Foroozanfard, F.; Kavossian, E.; Aghadavod, E.; Ostadmohammadi, V.; Reiter, R.J.; Eftekhar, T.; Asemi, Z. Effects of melatonin administration on mental health parameters, metabolic and genetic profiles in women with polycystic ovary syndrome: A randomized, double-blind, placebo-controlled trial. J. Affect. Disord. 2019, 250, 51–56. [Google Scholar] [CrossRef]
- Birdsall, T. Berberine: Therapeutic potential of an alkaloid found in several medicinal plants. Altern. Med. Rev. 1997, 2, 94–103. [Google Scholar]
- Kumar, A.; Ekavali, N.; Chopra, K.; Mukherjee, M.; Pottabathini, R.; Dhull, D.K. Current knowledge and pharmacological profile of berberine: An update. Eur. J. Pharmacol. 2015, 761, 288–297. [Google Scholar] [CrossRef]
- Zhang, Y.; Cui, Y.-L.; Gao, L.-N.; Jiang, H.-L. Effects of β-cyclodextrin on the intestinal absorption of berberine hydrochloride, a P-glycoprotein substrate. Int. J. Biol. Macromol. 2013, 59, 363–371. [Google Scholar] [CrossRef]
- Wang, K.; Feng, X.; Chai, L.; Cao, S.; Qiu, F. The metabolism of berberine and its contribution to the pharmacological effects. Drug Metab. Rev. 2017, 49, 139–157. [Google Scholar] [CrossRef]
- Kuang, H.; Duan, Y.; Li, D.; Xu, Y.; Ai, W.; Li, W.; Wang, Y.; Liu, S.; Li, M.; Liu, X.; et al. The role of serum inflammatory cytokines and berberine in the insulin signaling pathway among women with polycystic ovary syndrome. PLoS ONE 2020, 15, e0235404. [Google Scholar] [CrossRef]
- Lee, Y.S.; Kim, W.S.; Kim, K.H.; Yoon, M.J.; Cho, H.J.; Shen, Y.; Ye, J.-M.; Lee, C.H.; Oh, W.K.; Kim, C.T.; et al. Berberine, a natural plant product, activates AMP-activated protein kinase with beneficial metabolic effects in diabetic and insulin-resistant states. Diabetes 2006, 55, 2256–2264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Liu, X.; Zhuang, L.; Liu, X.; Zhao, H.; Shan, Y.; Liu, Z.; Li, F.; Wang, Y.; Fang, J. Berberine decreases insulin resistance in a PCOS rats by improving GLUT4: Dual regulation of the PI3K/AKT and MAPK pathways. Regul. Toxicol. Pharmacol. RTP 2020, 110, 104544. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Sun, Z.; Zhang, Y.; Liu, B.; Guan, Y.; Lu, M. The use of berberine for women with polycystic ovary syndrome undergoing IVF treatment. Clin. Endocrinol. 2014, 80, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Zhao, H.; Wang, A.; Sui, M.; Liang, K.; Deng, H.; Ma, Y.; Zhang, Y.; Zhang, H.; Guan, Y. A clinical study on the short-term effect of berberine in comparison to metformin on the metabolic characteristics of women with polycystic ovary syndrome. Eur. J. Endocrinol. 2012, 166, 99–105. [Google Scholar] [CrossRef]
- Li, L.; Li, C.; Pan, P.; Chen, X.; Wu, X.; Ng, E.H.Y.; Yang, D. A Single Arm Pilot Study of Effects of Berberine on the Menstrual Pattern, Ovulation Rate, Hormonal and Metabolic Profiles in Anovulatory Chinese Women with Polycystic Ovary Syndrome. PLoS ONE 2015, 10, e0144072. [Google Scholar] [CrossRef]
- Brusq, J.-M.; Ancellin, N.; Grondin, P.; Guillard, R.; Martin, S.; Saintillan, Y.; Issandou, M. Inhibition of lipid synthesis through activation of AMP kinase: An additional mechanism for the hypolipidemic effects of berberine. J. Lipid Res. 2006, 47, 1281–1288. [Google Scholar] [CrossRef]
- Kong, W.; Wei, J.; Abidi, P.; Lin, M.; Inaba, S.; Li, C.; Wang, Y.; Wang, Z.; Si, S.; Pan, H.; et al. Berberine is a novel cholesterol-lowering drug working through a unique mechanism distinct from statins. Nat. Med. 2004, 10, 1344–1351. [Google Scholar] [CrossRef]
- Birringer, M.; Siems, K.; Maxones, A.; Frank, J.; Lorkowski, S. Natural 6-hydroxy-chromanols and -chromenols: Structural diversity, biosynthetic pathways and health implications. RSC Adv. 2018, 8, 4803–4841. [Google Scholar] [CrossRef]
- Szewczyk, K.; Chojnacka, A.; Górnicka, M. Tocopherols and Tocotrienols-Bioactive Dietary Compounds; What Is Certain, What Is Doubt? Int. J. Mol. Sci. 2021, 22, 6222. [Google Scholar] [CrossRef]
- Zhao, L.; Fang, X.; Marshall, M.R.; Chung, S. Regulation of Obesity and Metabolic Complications by Gamma and Delta Tocotrienols. Molecules 2016, 21, 344. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.-Y.; Suhaimi, F.H.; Ahmad, F.; Ima-Nirwana, S. Vitamin E As a Potential Interventional Treatment for Metabolic Syndrome: Evidence from Animal and Human Studies. Front. Pharmacol. 2017, 8, 444. [Google Scholar] [CrossRef] [PubMed]
- Vafa, M.; Haghighat, N.; Moslehi, N.; Eghtesadi, S.; Heydari, I. Effect of Tocotrienols enriched canola oil on glycemic control and oxidative status in patients with type 2 diabetes mellitus: A randomized double-blind placebo-controlled clinical trial. J. Res. Med. Sci. Off. J. Isfahan Univ. Med. Sci. 2015, 20, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Vitamin E in Neuroprotection Study (VENUS) Investigators; Hor, C.P.; Fung, W.Y.; Ang, H.A.; Lim, S.C.; Kam, L.Y.; Sim, S.-W.; Lim, L.H.; Choon, W.Y.; Wong, J.W.; et al. Efficacy of Oral Mixed Tocotrienols in Diabetic Peripheral Neuropathy: A Randomized Clinical Trial. JAMA Neurol. 2018, 75, 444–452. [Google Scholar] [CrossRef]
- Wong, W.-Y.; Ward, L.C.; Fong, C.W.; Yap, W.N.; Brown, L. Anti-inflammatory γ- and δ-tocotrienols improve cardiovascular, liver and metabolic function in diet-induced obese rats. Eur. J. Nutr. 2017, 56, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Kamisah, Y.; Mohamed, N.; Muhammad, N.; Masbah, N.; Fahami, N.A.M.; Mohamed, I.N.; Shuid, A.N.; Saad, Q.M.; Abdullah, A.; et al. Potential Role of Tocotrienols on Non-Communicable Diseases: A Review of Current Evidence. Nutrients 2020, 12, 259. [Google Scholar] [CrossRef]
- Parker, R.A.; Pearce, B.C.; Clark, R.W.; Gordon, D.A.; Wright, J.J. Tocotrienols regulate cholesterol production in mammalian cells by post-transcriptional suppression of 3-hydroxy-3-methylglutaryl-coenzyme A reductase. J. Biol. Chem. 1993, 268, 11230–11238. [Google Scholar] [CrossRef]
- Torabi, S.; Yeganehjoo, H.; Shen, C.-L.; Mo, H. Peroxisome proliferator-activated receptor γ down-regulation mediates the inhibitory effect of d-δ-tocotrienol on the differentiation of murine 3T3-F442A preadipocytes. Nutr. Res. N. Y. N. 2016, 36, 1345–1352. [Google Scholar] [CrossRef]
- Pang, K.-L.; Chin, K.-Y. The Role of Tocotrienol in Protecting Against Metabolic Diseases. Molecules 2019, 24, 923. [Google Scholar] [CrossRef]
- Meganathan, P.; Fu, J.-Y. Biological Properties of Tocotrienols: Evidence in Human Studies. Int. J. Mol. Sci. 2016, 17, 1682. [Google Scholar] [CrossRef]
- Suleman, F.; Khan, D.A.; Pervez, M.A.; Aamir, M. Effects of delta-tocotrienol supplementation on glycaemic control in individuals with prediabetes: A randomized controlled study. JPMA J. Pak. Med. Assoc. 2022, 72, 4–7. [Google Scholar] [CrossRef]
- Mahjabeen, W.; Khan, D.A.; Mirza, S.A.; Pervez, M.A. Effects of delta-tocotrienol supplementation on Glycemic Control, oxidative stress, inflammatory biomarkers and miRNA expression in type 2 diabetes mellitus: A randomized control trial. Phytother. Res. PTR 2021, 35, 3968–3976. [Google Scholar] [CrossRef] [PubMed]
- Pervez, M.A.; Khan, D.A.; Slehria, A.U.R.; Ijaz, A. Delta-tocotrienol supplementation improves biochemical markers of hepatocellular injury and steatosis in patients with nonalcoholic fatty liver disease: A randomized, placebo-controlled trial. Complement. Ther. Med. 2020, 52, 102494. [Google Scholar] [CrossRef] [PubMed]
- Vitaglione, P.; Morisco, F.; Mazzone, G.; Amoruso, D.C.; Ribecco, M.T.; Romano, A.; Fogliano, V.; Caporaso, N.; D’Argenio, G. Coffee reduces liver damage in a rat model of steatohepatitis: The underlying mechanisms and the role of polyphenols and melanoidins. Hepatology 2010, 52, 1652–1661. [Google Scholar] [CrossRef] [PubMed]
- Natella, F.; Scaccini, C. Role of coffee in modulation of diabetes risk. Nutr. Rev. 2012, 70, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Baspinar, B.; Eskici, G.; Ozcelik, A.O. How coffee affects metabolic syndrome and its components. Food Funct. 2017, 8, 2089–2101. [Google Scholar] [CrossRef] [PubMed]
- Cossiga, V.; Lembo, V.; Guarino, M.; Tuccillo, C.; Morando, F.; Pontillo, G.; Fiorentino, A.; Caporaso, N.; Morisco, F. Berberis aristata, Elaeis guineensis and Coffea canephora Extracts Modulate the Insulin Receptor Expression and Improve Hepatic Steatosis in NAFLD Patients: A Pilot Clinical Trial. Nutrients 2019, 11, 3070. [Google Scholar] [CrossRef] [PubMed]
- Cossiga, V.; Lembo, V.; Nigro, C.; Mirra, P.; Miele, C.; D’Argenio, V.; Leone, A.; Mazzone, G.; Veneruso, I.; Guido, M.; et al. The Combination of Berberine, Tocotrienols and Coffee Extracts Improves Metabolic Profile and Liver Steatosis by the Modulation of Gut Microbiota and Hepatic miR-122 and miR-34a Expression in Mice. Nutrients 2021, 13, 1281. [Google Scholar] [CrossRef]
- Roshan, H.; Nikpayam, O.; Sedaghat, M.; Sohrab, G. Effects of green coffee extract supplementation on anthropometric indices, glycaemic control, blood pressure, lipid profile, insulin resistance and appetite in patients with the metabolic syndrome: A randomised clinical trial. Br. J. Nutr. 2018, 119, 250–258. [Google Scholar] [CrossRef]
- Kanetkar, P.; Singhal, R.; Kamat, M. Gymnema sylvestre: A Memoir. J. Clin. Biochem. Nutr. 2007, 41, 77–81. [Google Scholar] [CrossRef]
- Sinsheimer, J.E.; Subba Rao, G. Constituents from Gymnem sylvestre Leaves VI: Acylated Genins of the Gymnemic Acids-Isolation and Preliminary Characterization. J. Pharm. Sci. 1970, 59, 629–632. [Google Scholar] [CrossRef]
- Bhansali, S.; Shafiq, N.; Pandhi, P.; Singh, A.P.; Singh, I.; Singh, P.K.; Sharma, S.; Malhotra, S. Effect of a deacyl gymnemic acid on glucose homeostasis & metabolic parameters in a rat model of metabolic syndrome. Indian J. Med. Res. 2013, 137, 1174–1179. [Google Scholar] [PubMed]
- El Shafey, A.A.M.; El-Ezabi, M.M.; Seliem, M.M.E.; Ouda, H.H.M.; Ibrahim, D.S. Effect of Gymnema sylvestre R. Br. leaves extract on certain physiological parameters of diabetic rats. J. King Saud Univ.—Sci. 2013, 25, 135–141. [Google Scholar] [CrossRef]
- Zuñiga, L.Y.; González-Ortiz, M.; Martínez-Abundis, E. Effect of Gymnema sylvestre Administration on Metabolic Syndrome, Insulin Sensitivity, and Insulin Secretion. J. Med. Food 2017, 20, 750–754. [Google Scholar] [CrossRef]
- Martínez-Abundis, E.; Méndez-Del Villar, M.; Pérez-Rubio, K.G.; Zuñiga, L.Y.; Cortez-Navarrete, M.; Ramírez-Rodriguez, A.; González-Ortiz, M. Novel nutraceutic therapies for the treatment of metabolic syndrome. World J. Diabetes 2016, 7, 142–152. [Google Scholar] [CrossRef]
- Tiwari, P.; Mishra, B.N.; Sangwan, N.S. Phytochemical and pharmacological properties of Gymnema sylvestre: An important medicinal plant. BioMed Res. Int. 2014, 2014, 830285. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrillo, T.; Semprini, E.; Tomatis, V.; Arnesano, M.; Ambrosetti, F.; Battipaglia, C.; Sponzilli, A.; Ricciardiello, F.; Genazzani, A.R.; Genazzani, A.D. Putative Complementary Compounds to Counteract Insulin-Resistance in PCOS Patients. Biomedicines 2022, 10, 1924. https://doi.org/10.3390/biomedicines10081924
Petrillo T, Semprini E, Tomatis V, Arnesano M, Ambrosetti F, Battipaglia C, Sponzilli A, Ricciardiello F, Genazzani AR, Genazzani AD. Putative Complementary Compounds to Counteract Insulin-Resistance in PCOS Patients. Biomedicines. 2022; 10(8):1924. https://doi.org/10.3390/biomedicines10081924
Chicago/Turabian StylePetrillo, Tabatha, Elisa Semprini, Veronica Tomatis, Melania Arnesano, Fedora Ambrosetti, Christian Battipaglia, Alessandra Sponzilli, Francesco Ricciardiello, Andrea R. Genazzani, and Alessandro D. Genazzani. 2022. "Putative Complementary Compounds to Counteract Insulin-Resistance in PCOS Patients" Biomedicines 10, no. 8: 1924. https://doi.org/10.3390/biomedicines10081924
APA StylePetrillo, T., Semprini, E., Tomatis, V., Arnesano, M., Ambrosetti, F., Battipaglia, C., Sponzilli, A., Ricciardiello, F., Genazzani, A. R., & Genazzani, A. D. (2022). Putative Complementary Compounds to Counteract Insulin-Resistance in PCOS Patients. Biomedicines, 10(8), 1924. https://doi.org/10.3390/biomedicines10081924