
The Translational Role of miRNA in Polycystic Ovary Syndrome: From Bench to Bedside—A Systematic Literature Review

 ,
, 
 ,
,  ,
,  ,
,  , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
3. Results
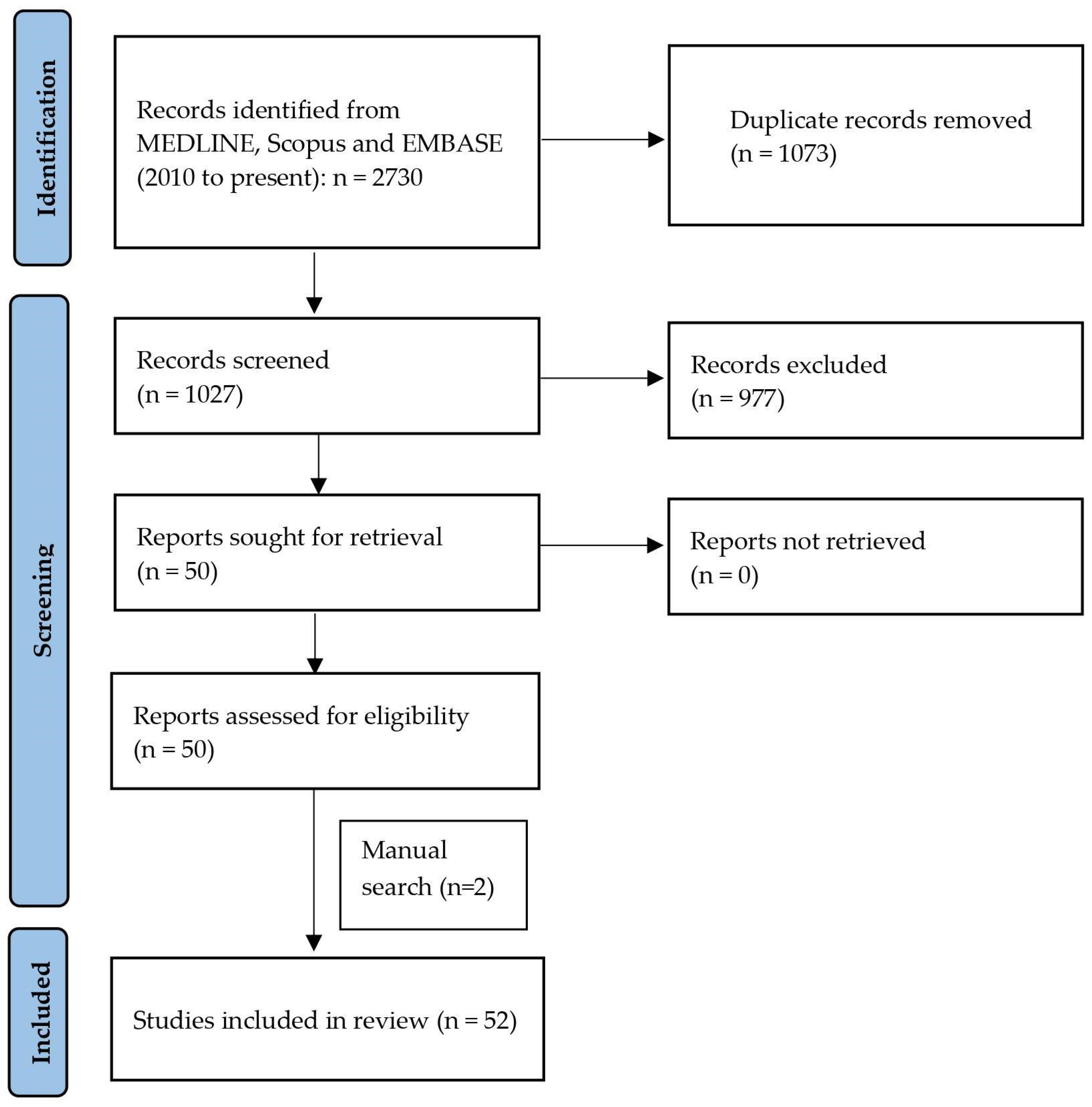
3.1. Literature Search
3.2. Study Characteristics and Quality
4. Discussion
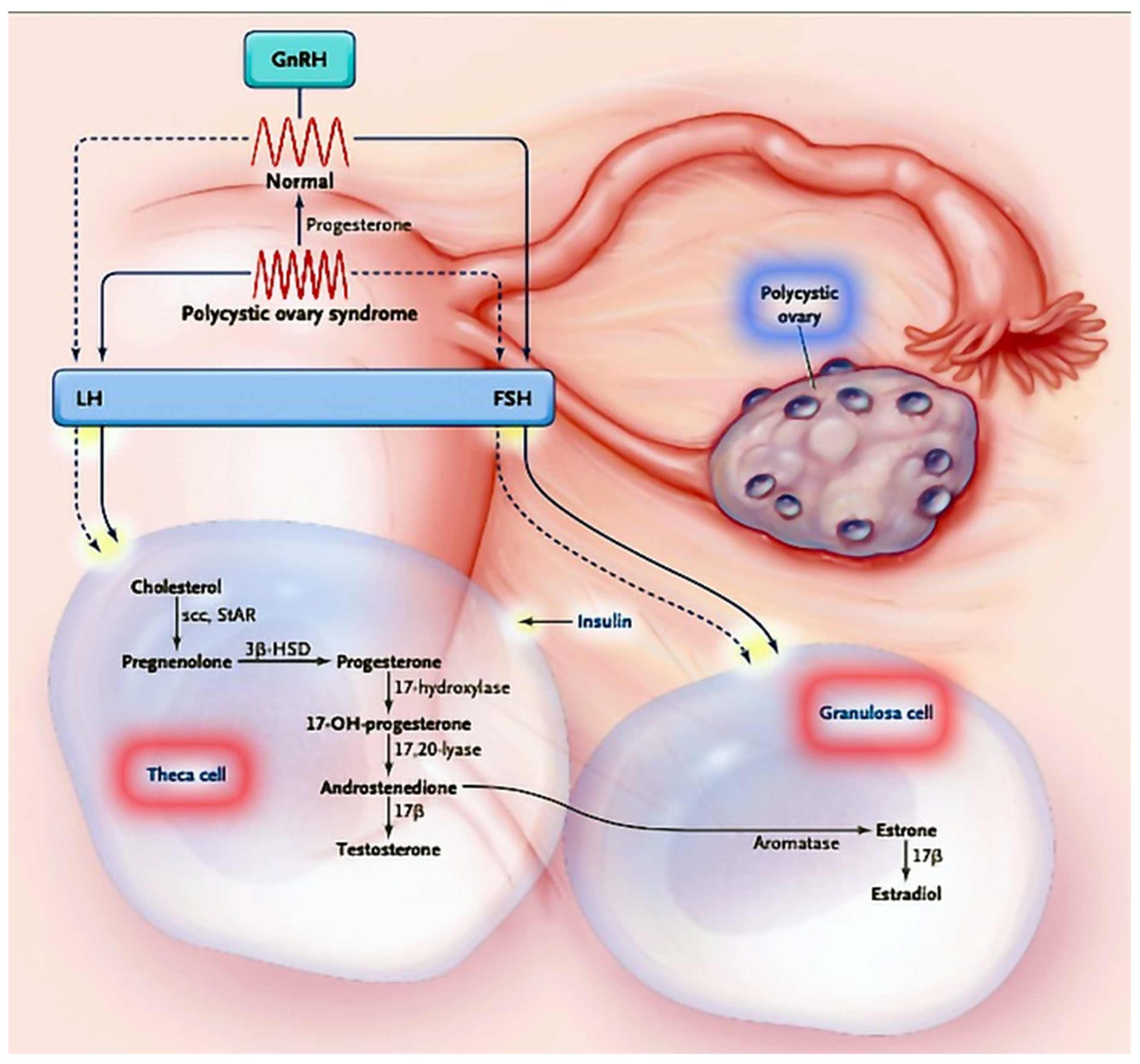
4.1. PCOS: Definition, Clinical Presentation and Heterogeneity
4.2. Metabolic and Fertility Consequences of PCOS
4.3. MicroRNAs in Granulosa Cells
4.4. MicroRNAs in Theca Cells
4.5. MicroRNAs and Fertility in PCOS
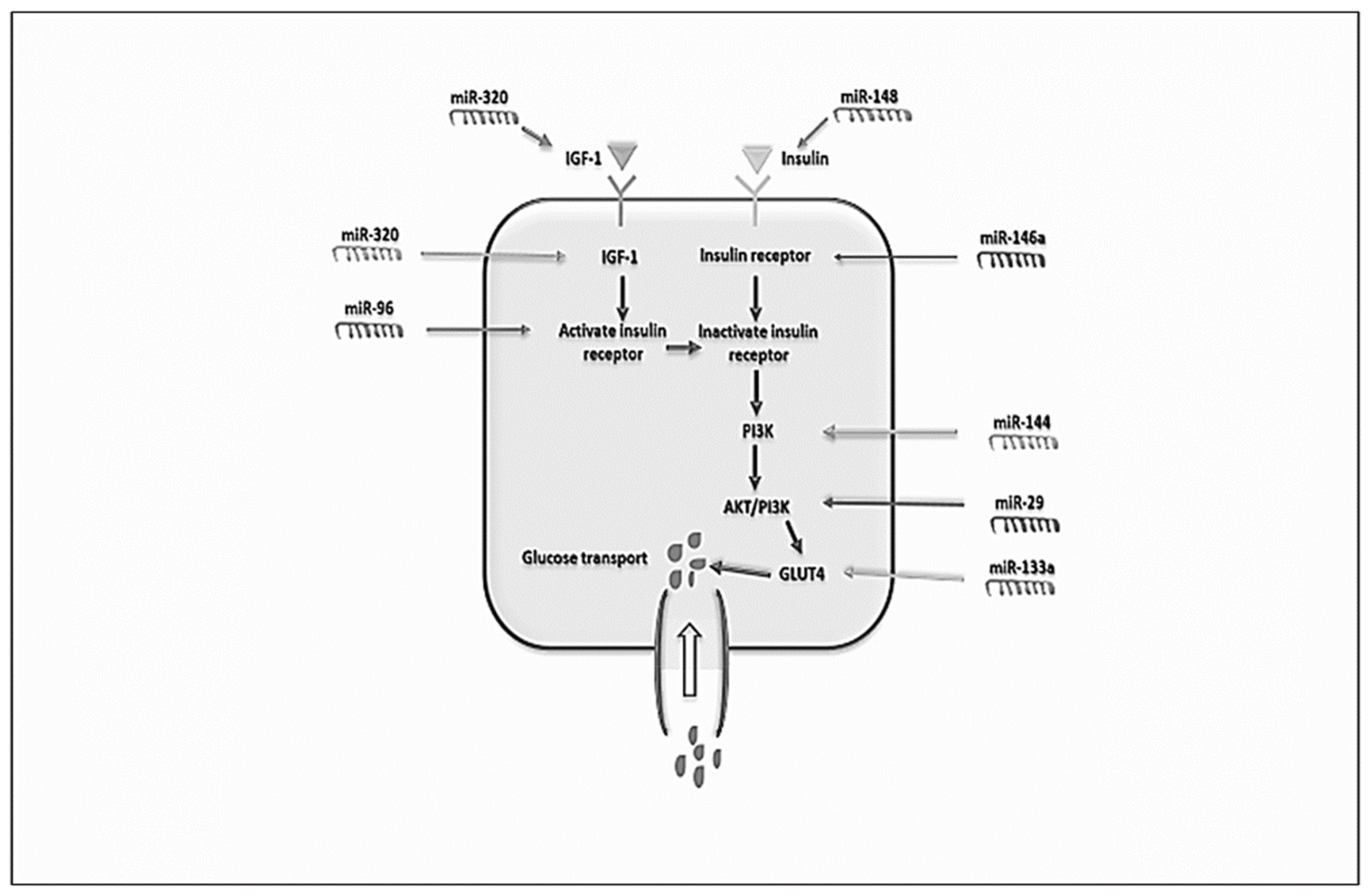
4.6. MicroRNAs, Glucose and Lipid Metabolism in PCOS
4.7. Future Directions: miRNAs as a Potential Therapeutic Target in PCOS and as a Diagnostic Marker for PCOS
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bushati, N.; Cohen, S.M. MicroRNA Functions. Annu. Rev. Cell Dev. Biol. 2007, 23, 175–205. [Google Scholar] [CrossRef] [PubMed]
- Bhaskaran, M.; Mohan, M. MicroRNAs: History, Biogenesis, and Their Evolving Role in Animal Development and Disease. Vet. Pathol. 2014, 51, 759–774. [Google Scholar] [CrossRef] [PubMed]
- Vidigal, J.A.; Ventura, A. The Biological Functions of miRNAs: Lessons from in Vivo Studies. Trends Cell Biol. 2015, 25, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.-H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA Genes Are Transcribed by RNA Polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef]
- Alarcón, C.R.; Lee, H.; Goodarzi, H.; Halberg, N.; Tavazoie, S.F. N6-Methyladenosine Marks Primary microRNAs for Processing. Nature 2015, 519, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; He, J.; Pu, W.; Peng, Y. The Role of Exportin-5 in microRNA Biogenesis and Cancer. Genom. Proteom. Bioinform. 2018, 16, 120–126. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA Biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Yan, H.; Liang, F.-S. MiRNA Inhibition by Proximity-Enabled Dicer Inactivation. Methods 2019, 167, 117–123. [Google Scholar] [CrossRef]
- Michlewski, G.; Cáceres, J.F. Post-Transcriptional Control of miRNA Biogenesis. RNA 2019, 25, 1–16. [Google Scholar] [CrossRef]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef]
- Höck, J.; Meister, G. The Argonaute Protein Family. Genome Biol. 2008, 9, 210. [Google Scholar] [CrossRef] [PubMed]
- Campos-Melo, D.; Droppelmann, C.A.; He, Z.; Volkening, K.; Strong, M.J. Altered microRNA Expression Profile in Amyotrophic Lateral Sclerosis: A Role in the Regulation of NFL MRNA Levels. Mol. Brain 2013, 6, 26. [Google Scholar] [CrossRef]
- Li, J.; Wan, Y.; Guo, Q.; Zou, L.; Zhang, J.; Fang, Y.; Zhang, J.; Zhang, J.; Fu, X.; Liu, H.; et al. Altered microRNA Expression Profile with MiR-146a Upregulation in CD4+ T Cells from Patients with Rheumatoid Arthritis. Arthritis Res. Ther. 2010, 12, R81. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-Y.; Liu, W.; Zhou, Y.-M.; Shen, X.-M.; Wang, Y.-F.; Tang, G.-Y. Altered microRNA Expression Profile with MiR-27b down-Regulation Correlated with Disease Activity of Oral Lichen Planus: MiRNA in OLP. Oral Dis. 2012, 18, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Grunert, M.; Appelt, S.; Dunkel, I.; Berger, F.; Sperling, S.R. Altered microRNA and Target Gene Expression Related to Tetralogy of Fallot. Sci. Rep. 2019, 9, 19063. [Google Scholar] [CrossRef]
- Javadi, M.; Rad, J.S.; Farashah, M.S.G.; Roshangar, L. An Insight on the Role of Altered Function and Expression of Exosomes and microRNAs in Female Reproductive Diseases. Reprod. Sci. 2022, 29, 1395–1407. [Google Scholar] [CrossRef]
- Cianci, A.; Vitale, S.G. D-Chiro-Inositol and PCOS: Between Myth and Reality. The Never-Ending Story. Int. J. Food Sci. Nutr. 2022, 73, 565–570. [Google Scholar] [CrossRef]
- Fulghesu, A.M.; Angioni, S.; Frau, E.; Belosi, C.; Apa, R.; Mioni, R.; Xamin, N.; Capobianco, G.P.; Dessole, S.; Fruzzetti, F.; et al. Ultrasound in Polycystic Ovary Syndrome the Measuring of Ovarian Stroma and Relationship with Circulating Androgens: Results of a Multicentric Study. Hum. Reprod. 2007, 22, 2501–2508. [Google Scholar] [CrossRef]
- Lujan, M.E.; Chizen, D.R.; Pierson, R.A. Diagnostic Criteria for Polycystic Ovary Syndrome: Pitfalls and Controversies. J. Obstet. Gynaecol. Can. 2008, 30, 671–679. [Google Scholar] [CrossRef]
- The Rotterdam ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group. Revised 2003 Consensus on Diagnostic Criteria and Long-Term Health Risks Related to Polycystic Ovary Syndrome (PCOS). Hum. Reprod. 2004, 19, 41–47. [Google Scholar] [CrossRef]
- Azziz, R.; Carmina, E.; Dewailly, D.; Diamanti-Kandarakis, E.; Escobar-Morreale, H.F.; Futterweit, W.; Janssen, O.E.; Legro, R.S.; Norman, R.J.; Taylor, A.E.; et al. Criteria for Defining Polycystic Ovary Syndrome as a Predominantly Hyperandrogenic Syndrome: An Androgen Excess Society Guideline. J. Clin. Endocrinol. Metab. 2006, 91, 4237–4245. [Google Scholar] [CrossRef] [PubMed]
- McCartney, C.R.; Marshall, J.C. Polycystic Ovary Syndrome. N. Engl. J. Med. 2016, 375, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Angioni, S.; Portoghese, E.; Milano, F.; Melis, G.B.; Fulghesu, A.M. Diagnosis of Metabolic Disorders in Women with Polycystic Ovary Syndrome. Obstet. Gynecol. Surv. 2008, 63, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Rojas, J.; Chávez, M.; Olivar, L.; Rojas, M.; Morillo, J.; Mejías, J.; Calvo, M.; Bermúdez, V. Polycystic Ovary Syndrome, Insulin Resistance, and Obesity: Navigating the Pathophysiologic Labyrinth. Int. J. Reprod. Med. 2014, 2014, 719050. [Google Scholar] [CrossRef] [PubMed]
- Fulghesu, A.M.; Piras, C.; Dessì, A.; Succu, C.; Atzori, L.; Pintus, R.; Gentile, C.; Angioni, S.; Fanos, V. Urinary Metabolites Reveal Hyperinsulinemia and Insulin Resistance in Polycystic Ovarian Syndrome (PCOS). Metabolites 2021, 11, 437. [Google Scholar] [CrossRef]
- Fulghesu, A.M.; Angioni, S.; Portoghese, E.; Milano, F.; Batetta, B.; Paoletti, A.M.; Melis, G.B. Failure of the Homeostatic Model Assessment Calculation Score for Detecting Metabolic Deterioration in Young Patients with Polycystic Ovary Syndrome. Fertil. Steril. 2006, 86, 398–404. [Google Scholar] [CrossRef]
- Witchel, S.F.; Oberfield, S.E.; Peña, A.S. Polycystic Ovary Syndrome: Pathophysiology, Presentation, and Treatment with Emphasis on Adolescent Girls. J. Endocr. Soc. 2019, 3, 1545–1573. [Google Scholar] [CrossRef]
- Cai, G.; Ma, X.; Chen, B.; Huang, Y.; Liu, S.; Yang, H.; Zou, W. MicroRNA-145 Negatively Regulates Cell Proliferation through Targeting IRS1 in Isolated Ovarian Granulosa Cells From Patients with Polycystic Ovary Syndrome. Reprod. Sci. 2017, 24, 902–910. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Heneidi, S.; Lee, J.-M.; Layman, L.C.; Stepp, D.W.; Gamboa, G.M.; Chen, B.-S.; Chazenbalk, G.; Azziz, R. MiRNA-93 Inhibits GLUT4 and Is Overexpressed in Adipose Tissue of Polycystic Ovary Syndrome Patients and Women with Insulin Resistance. Diabetes 2013, 62, 2278–2286. [Google Scholar] [CrossRef]
- Ding, C.-F.; Chen, W.-Q.; Zhu, Y.-T.; Bo, Y.-L.; Hu, H.-M.; Zheng, R.-H. Circulating microRNAs in Patients with Polycystic Ovary Syndrome. Hum. Fertil. 2015, 18, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, S.O.; Reiisi, S.; Parchami Barjui, S. Increased Risk of Polycystic Ovary Syndrome (PCOS) Associated with CC Genotype of MiR-146a Gene Variation. Gynecol. Endocrinol. 2018, 34, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, I.; Nahmias, N.; Novoselsky Persky, M.; Greenfield, C.; Goldman-Wohl, D.; Hurwitz, A.; Haimov-Kochman, R.; Yagel, S.; Imbar, T. Elevated Circulating Micro-Ribonucleic Acid (miRNA)-200b and miRNA-429 Levels in Anovulatory Women. Fertil. Steril. 2017, 107, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Sui, C.; Xun, Y.; Lai, Q.; Jin, L. MiRNA-99a Can Regulate Proliferation and Apoptosis of Human Granulosa Cells via Targeting IGF-1R in Polycystic Ovary Syndrome. J. Assist. Reprod. Genet. 2019, 36, 211–221. [Google Scholar] [CrossRef]
- He, T.; Liu, Y.; Jia, Y.; Wang, H.; Yang, X.; Lu, G.; Liu, H.; Shi, Y. MicroRNA-141 and MicroRNA-200c Are Overexpressed in Granulosa Cells of Polycystic Ovary Syndrome Patients. Front. Med. 2018, 5, 299. [Google Scholar] [CrossRef]
- Hosseini, A.H.; Kohan, L.; Aledavood, A.; Rostami, S. Association of miR-146a Rs2910164 and miR-222 Rs2858060 Polymorphisms with the Risk of Polycystic Ovary Syndrome in Iranian Women: A Case–Control Study. Taiwan. J. Obstet. Gynecol. 2017, 56, 652–656. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, Y.; Xu, S.; Qi, G.; Wu, X. Bioinformatics Identification of microRNAs Involved in Polycystic Ovary Syndrome Based on Microarray Data. Mol. Med. Rep. 2019, 20, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Tang, T.; Zeng, Z.; Wu, J.; Tan, X.; Yan, J. The Expression of Small RNAs in Exosomes of Follicular Fluid Altered in Human Polycystic Ovarian Syndrome. PeerJ 2020, 8, e8640. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Liu, C.; Hao, C.; Tang, Q.; Liu, R.; Lin, S.; Zhang, L.; Yan, W. Identification of Altered microRNAs and mRNAs in the Cumulus Cells of PCOS Patients: MiRNA-509-3p Promotes Oestradiol Secretion by Targeting MAP3K8. Reproduction 2016, 151, 643–655. [Google Scholar] [CrossRef]
- Jiang, L.; Huang, J.; Chen, Y.; Yang, Y.; Li, R.; Li, Y.; Chen, X.; Yang, D. Identification of Several Circulating microRNAs from a Genome-Wide Circulating microRNA Expression Profile as Potential Biomarkers for Impaired Glucose Metabolism in Polycystic Ovarian Syndrome. Endocrine 2016, 53, 280–290. [Google Scholar] [CrossRef]
- Li, Y.; Xiang, Y.; Song, Y.; Wan, L.; Yu, G.; Tan, L. Dysregulated miR-142, -33b and -423 in Granulosa Cells Target TGFBR1 and SMAD7: A Possible Role in Polycystic Ovary Syndrome. Mol. Hum. Reprod. 2019, 25, 638–646. [Google Scholar] [CrossRef]
- Lin, L.; Du, T.; Huang, J.; Huang, L.-L.; Yang, D.-Z. Identification of Differentially Expressed microRNAs in the Ovary of Polycystic Ovary Syndrome with Hyperandrogenism and Insulin Resistance. Chin. Med. J. 2015, 128, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Huang, J.; Li, L.; Chen, Y.; Chen, X.; Zhao, X.; Yang, D. MicroRNA-93 Promotes Ovarian Granulosa Cells Proliferation through Targeting CDKN1A in Polycystic Ovarian Syndrome. J. Clin. Endocrinol. Metab. 2015, 100, E729–E738. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhang, X.; Shi, C.; Lin, J.; Chen, G.; Wu, B.; Wu, L.; Shi, H.; Yuan, Y.; Zhou, W.; et al. Altered microRNAs Expression Profiling in Cumulus Cells from Patients with Polycystic Ovary Syndrome. J. Transl. Med. 2015, 13, 238. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Li, W.; Wu, M.; Cao, S. Ciculating miRNA-21 as a Biomarker Predicts Polycystic Ovary Syndrome (PCOS) in Patients. Clin. Lab. 2015, 61, 1009–1015. [Google Scholar] [CrossRef]
- Long, W.; Zhao, C.; Ji, C.; Ding, H.; Cui, Y.; Guo, X.; Shen, R.; Liu, J. Characterization of Serum microRNAs Profile of PCOS and Identification of Novel Non-Invasive Biomarkers. Cell. Physiol. Biochem. 2014, 33, 1304–1315. [Google Scholar] [CrossRef]
- Luo, H.; Han, Y.; Liu, J.; Zhang, Y. Identification of microRNAs in Granulosa Cells from Patients with Different Levels of Ovarian Reserve Function and the Potential Regulatory Function of MiR-23a in Granulosa Cell Apoptosis. Gene 2019, 686, 250–260. [Google Scholar] [CrossRef]
- Mao, Z.; Fan, L.; Yu, Q.; Luo, S.; Wu, X.; Tang, J.; Kang, G.; Tang, L. Abnormality of Klotho Signaling Is Involved in Polycystic Ovary Syndrome. Reprod. Sci. 2018, 25, 372–383. [Google Scholar] [CrossRef]
- McAllister, J.M.; Han, A.X.; Modi, B.P.; Teves, M.E.; Mavodza, G.R.; Anderson, Z.L.; Shen, T.; Christenson, L.K.; Archer, K.J.; Strauss, J.F. MiRNA Profiling Reveals miRNA-130b-3p Mediates DENND1A Variant 2 Expression and Androgen Biosynthesis. Endocrinology 2019, 160, 1964–1981. [Google Scholar] [CrossRef]
- McCallie, B.; Schoolcraft, W.B.; Katz-Jaffe, M.G. Aberration of Blastocyst microRNA Expression Is Associated with Human Infertility. Fertil. Steril. 2010, 93, 2374–2382. [Google Scholar] [CrossRef]
- Murri, M.; Insenser, M.; Fernández-Durán, E.; San-Millán, J.L.; Escobar-Morreale, H.F. Effects of Polycystic Ovary Syndrome (PCOS), Sex Hormones, and Obesity on Circulating miRNA-21, miRNA-27b, miRNA-103, and miRNA-155 Expression. J. Clin. Endocrinol. Metab. 2013, 98, E1835–E1844. [Google Scholar] [CrossRef]
- Murri, M.; Insenser, M.; Fernández-Durán, E.; San-Millán, J.L.; Luque-Ramírez, M.; Escobar-Morreale, H.F. Non-Targeted Profiling of Circulating microRNAs in Women with Polycystic Ovary Syndrome (PCOS): Effects of Obesity and Sex Hormones. Metabolism 2018, 86, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Naji, M.; Aleyasin, A.; Nekoonam, S.; Arefian, E.; Mahdian, R.; Amidi, F. Differential Expression of MiR-93 and MiR-21 in Granulosa Cells and Follicular Fluid of Polycystic Ovary Syndrome Associating with Different Phenotypes. Sci. Rep. 2017, 7, 14671. [Google Scholar] [CrossRef]
- Naji, M.; Nekoonam, S.; Aleyasin, A.; Arefian, E.; Mahdian, R.; Azizi, E.; Shabani Nashtaei, M.; Amidi, F. Expression of MiR-15a, MiR-145, and MiR-182 in Granulosa-Lutein Cells, Follicular Fluid, and Serum of Women with Polycystic Ovary Syndrome (PCOS). Arch. Gynecol. Obstet. 2018, 297, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Nanda, D.; Chandrasekaran, S.P.; Ramachandran, V.; Kalaivanan, K.; Carani Venkatraman, A. Evaluation of Serum miRNA-24, miRNA-29a and miRNA-502-3p Expression in PCOS Subjects: Correlation with Biochemical Parameters Related to PCOS and Insulin Resistance. Indian J. Clin. Biochem. 2020, 35, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Rashad, N.M.; Ateya, M.A.-M.; Saraya, Y.S.; Elnagar, W.M.; Helal, K.F.; Lashin, M.E.-B.; Abdelrhman, A.A.; Alil, A.E.; Yousef, M.S. Association of miRNA- 320 Expression Level and Its Target Gene Endothelin-1 with the Susceptibility and Clinical Features of Polycystic Ovary Syndrome. J. Ovarian Res. 2019, 12, 39. [Google Scholar] [CrossRef]
- Roth, L.W.; McCallie, B.; Alvero, R.; Schoolcraft, W.B.; Minjarez, D.; Katz-Jaffe, M.G. Altered microRNA and Gene Expression in the Follicular Fluid of Women with Polycystic Ovary Syndrome. J. Assist. Reprod. Genet. 2014, 31, 355–362. [Google Scholar] [CrossRef]
- Sang, Q.; Yao, Z.; Wang, H.; Feng, R.; Wang, H.; Zhao, X.; Xing, Q.; Jin, L.; He, L.; Wu, L.; et al. Identification of microRNAs in Human Follicular Fluid: Characterization of microRNAs That Govern Steroidogenesis in Vitro and Are Associated with Polycystic Ovary Syndrome in Vivo. J. Clin. Endocrinol. Metab. 2013, 98, 3068–3079. [Google Scholar] [CrossRef]
- Sathyapalan, T.; David, R.; Gooderham, N.J.; Atkin, S.L. Increased Expression of Circulating miRNA-93 in Women with Polycystic Ovary Syndrome May Represent a Novel, Non-Invasive Biomarker for Diagnosis. Sci. Rep. 2015, 5, 16890. [Google Scholar] [CrossRef]
- Scalici, E.; Traver, S.; Mullet, T.; Molinari, N.; Ferrières, A.; Brunet, C.; Belloc, S.; Hamamah, S. Circulating microRNAs in Follicular Fluid, Powerful Tools to Explore in Vitro Fertilization Process. Sci. Rep. 2016, 6, 24976. [Google Scholar] [CrossRef]
- Shi, L.; Liu, S.; Zhao, W.; Shi, J. MiR-483–5p and MiR-486–5p Are down-Regulated in Cumulus Cells of Metaphase II Oocytes from Women with Polycystic Ovary Syndrome. Reprod. BioMed. Online 2015, 31, 565–572. [Google Scholar] [CrossRef]
- Song, J.; Luo, S.; Li, S. MiRNA-592 Is Downregulated and May Target LHCGR in Polycystic Ovary Syndrome Patients. Reprod. Biol. 2015, 15, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Song, D.K.; Sung, Y.-A.; Lee, H. The Role of Serum microRNA-6767-5p as a Biomarker for the Diagnosis of Polycystic Ovary Syndrome. PLoS ONE 2016, 11, e0163756. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Yu, G.; Xiang, Y.; Li, Y.; Wan, L.; Tan, L. Altered MiR-186 and MiR-135a Contribute to Granulosa Cell Dysfunction by Targeting ESR2: A Possible Role in Polycystic Ovary Syndrome. Mol. Cell. Endocrinol. 2019, 494, 110478. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, A.E.; Wissing, M.L.; Englund, A.L.M.; Dalgaard, L.T. MicroRNA Species in Follicular Fluid Associating with Polycystic Ovary Syndrome and Related Intermediary Phenotypes. J. Clin. Endocrinol. Metab. 2016, 101, 1579–1589. [Google Scholar] [CrossRef]
- Sørensen, A.E.; Udesen, P.B.; Maciag, G.; Geiger, J.; Saliani, N.; Januszewski, A.S.; Jiang, G.; Ma, R.C.; Hardikar, A.A.; Wissing, M.L.M.; et al. Hyperandrogenism and Metabolic Syndrome Are Associated with Changes in Serum-Derived microRNAs in Women with Polycystic Ovary Syndrome. Front. Med. 2019, 6, 242. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Sun, J.; Xu, B.; Chrusciel, M.; Gao, J.; Bazert, M.; Stelmaszewska, J.; Xu, Y.; Zhang, H.; Pawelczyk, L.; et al. Functional Characterization of microRNA-27a-3p Expression in Human Polycystic Ovary Syndrome. Endocrinology 2018, 159, 297–309. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, S.; Wang, Y.; Qi, G.; Hou, Y.; Sun, C.; Wu, X. Identification and Potential Value of Candidate microRNAs in Granulosa Cells of Polycystic Ovary Syndrome. THC 2019, 27, 579–587. [Google Scholar] [CrossRef]
- Wu, H.-L.; Heneidi, S.; Chuang, T.-Y.; Diamond, M.P.; Layman, L.C.; Azziz, R.; Chen, Y.-H. The Expression of the miR-25/93/106b Family of Micro-RNAs in the Adipose Tissue of Women with Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2014, 99, E2754–E2761. [Google Scholar] [CrossRef]
- Xiang, Y.; Song, Y.; Li, Y.; Zhao, D.; Ma, L.; Tan, L. MiR-483 Is Down-Regulated in Polycystic Ovarian Syndrome and Inhibits KGN Cell Proliferation via Targeting Insulin-Like Growth Factor 1 (IGF1). Med. Sci. Monit. 2016, 22, 3383–3393. [Google Scholar] [CrossRef]
- Xiong, W.; Lin, Y.; Xu, L.; Tamadon, A.; Zou, S.; Tian, F.; Shao, R.; Li, X.; Feng, Y. Circulatory microRNA 23a and microRNA 23b and Polycystic Ovary Syndrome (PCOS): The Effects of Body Mass Index and Sex Hormones in an Eastern Han Chinese Population. J. Ovarian Res. 2017, 10, 10. [Google Scholar] [CrossRef]
- Xu, B.; Zhang, Y.-W.; Tong, X.-H.; Liu, Y.-S. Characterization of microRNA Profile in Human Cumulus Granulosa Cells: Identification of microRNAs That Regulate Notch Signaling and Are Associated with PCOS. Mol. Cell. Endocrinol. 2015, 404, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Lv, J.; Xu, P.; Gu, L.; Cao, J.; Xu, L.; Xue, K.; Li, Q. Identification of microRNAs and Genes Associated with Hyperandrogenism in the Follicular Fluid of Women with Polycystic Ovary Syndrome. J. Cell. Biochem. 2018, 119, 3913–3921. [Google Scholar] [CrossRef] [PubMed]
- Yao, N.; Yang, B.-Q.; Liu, Y.; Tan, X.-Y.; Lu, C.-L.; Yuan, X.-H.; Ma, X. Follicle-Stimulating Hormone Regulation of microRNA Expression on Progesterone Production in Cultured Rat Granulosa Cells. Endocrine 2010, 38, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Li, M.; Hu, J.; Wang, W.; Gao, M. MiRNA-335-5p Negatively Regulates Granulosa Cell Proliferation via SGK3 in PCOS. Reproduction 2018, 156, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Wang, X.; Yao, G.; Lü, M.; Liang, M.; Sun, Y.; Sun, F. Transactivation of microRNA-320 by microRNA-383 Regulates Granulosa Cell Functions by Targeting E2F1 and SF-1 Proteins. J. Biol. Chem. 2014, 289, 18239–18257. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, H.; Yan, C.; Gao, X.; Ling, X. Deregulation of RUNX2 by MiR-320a Deficiency Impairs Steroidogenesis in Cumulus Granulosa Cells from Polycystic Ovary Syndrome (PCOS) Patients. Biochem. Biophys. Res. Commun. 2017, 482, 1469–1476. [Google Scholar] [CrossRef]
- Zhang, H.; Gao, Z.; Zhang, Y.; Wang, H.; Li, Y. MiR-873-5p Regulated LPS-Induced Oxidative Stress via Targeting Heme Oxygenase-1 (HO-1) in KGN Cells. RSC Adv. 2018, 8, 39098–39105. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, X.; Shi, Z.; Zhang, J.; Zhang, J.; Jia, X.; Ling, X. Role of Serum miRNAs in the Prediction of Ovarian Hyperstimulation Syndrome in Polycystic Ovarian Syndrome Patients. Cell. Physiol. Biochem. 2015, 35, 1086–1094. [Google Scholar] [CrossRef]
- Zhong, Z.; Li, F.; Li, Y.; Qin, S.; Wen, C.; Fu, Y.; Xiao, Q. Inhibition of microRNA-19b Promotes Ovarian Granulosa Cell Proliferation by Targeting IGF-1 in Polycystic Ovary Syndrome. Mol. Med. Rep. 2018, 17, 4889–4898. [Google Scholar] [CrossRef]
- Butler, A.E.; Ramachandran, V.; Hayat, S.; Dargham, S.R.; Cunningham, T.K.; Benurwar, M.; Sathyapalan, T.; Najafi-Shoushtari, S.H.; Atkin, S.L. Expression of microRNA in Follicular Fluid in Women with and without PCOS. Sci. Rep. 2019, 9, 16306. [Google Scholar] [CrossRef]
- Luo, Y.; Cui, C.; Han, X.; Wang, Q.; Zhang, C. The Role of miRNAs in Polycystic Ovary Syndrome with Insulin Resistance. J. Assist. Reprod. Genet. 2021, 38, 289–304. [Google Scholar] [CrossRef] [PubMed]
- Gay, S.; Bugeon, J.; Bouchareb, A.; Henry, L.; Delahaye, C.; Legeai, F.; Montfort, J.; Le Cam, A.; Siegel, A.; Bobe, J.; et al. MiR-202 Controls Female Fecundity by Regulating Medaka Oogenesis. PLoS Genet. 2018, 14, e1007593. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Friedman, J.R. MiR-122 Regulates Hepatic Lipid Metabolism and Tumor Suppression. J. Clin. Investig. 2012, 122, 2773–2776. [Google Scholar] [CrossRef] [PubMed]
- Azziz, R.; Adashi, E.Y. Stein and Leventhal: 80 Years On. Am. J. Obstet. Gynecol. 2016, 214, 247.e1–247.e11. [Google Scholar] [CrossRef] [PubMed]
- Clark, N.M.; Podolski, A.J.; Brooks, E.D.; Chizen, D.R.; Pierson, R.A.; Lehotay, D.C.; Lujan, M.E. Prevalence of Polycystic Ovary Syndrome Phenotypes Using Updated Criteria for Polycystic Ovarian Morphology: An Assessment of Over 100 Consecutive Women Self-Reporting Features of Polycystic Ovary Syndrome. Reprod. Sci. 2014, 21, 1034–1043. [Google Scholar] [CrossRef]
- Sanchez-Garrido, M.A.; Tena-Sempere, M. Metabolic Dysfunction in Polycystic Ovary Syndrome: Pathogenic Role of Androgen Excess and Potential Therapeutic Strategies. Mol. Metab. 2020, 35, 100937. [Google Scholar] [CrossRef]
- Rosenfield, R.L.; Ehrmann, D.A. The Pathogenesis of Polycystic Ovary Syndrome (PCOS): The Hypothesis of PCOS as Functional Ovarian Hyperandrogenism Revisited. Endocr. Rev. 2016, 37, 467–520. [Google Scholar] [CrossRef]
- Ehrmann, D.A. Polycystic ovary syndrome. N. Engl. J. Med. 2005, 352, 1223–1236. [Google Scholar] [CrossRef]
- Kim, J.J.; Choi, Y.M. Dyslipidemia in Women with Polycystic Ovary Syndrome. Obstet. Gynecol. Sci. 2013, 56, 137–142. [Google Scholar] [CrossRef]
- Wild, R.A. Dyslipidemia in PCOS. Steroids 2012, 77, 295–299. [Google Scholar] [CrossRef]
- Stadler, J.T.; Marsche, G. Obesity-Related Changes in High-Density Lipoprotein Metabolism and Function. Int. J. Mol. Sci. 2020, 21, 8985. [Google Scholar] [CrossRef] [PubMed]
- Brady, C.; Mousa, S.S.; Mousa, S.A. Polycystic Ovary Syndrome and Its Impact on Women’s Quality of Life: More than Just an Endocrine Disorder. Drug Healthc. Patient Saf. 2009, 1, 9–15. [Google Scholar] [CrossRef]
- Fulghesu, A.M.; Angioni, S.; Belosi, C.; Apa, R.; Selvaggi, L.; Ciampelli, M.; Iuculano, A.; Melis, G.B.; Lanzone, A. Pituitary?Ovarian Response to the Gonadotrophin-Releasing Hormone-Agonist Test in Anovulatory Patients with Polycystic Ovary Syndrome: Predictive Role of Ovarian Stroma. Clin. Endocrinol. 2006, 65, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Kamalanathan, S.; Sahoo, J.P.; Sathyapalan, T. Pregnancy in Polycystic Ovary Syndrome. Indian J. Endocrinol. Metab. 2013, 17, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, H.; Taylor, H.S. Implantation Failure: Molecular Mechanisms and Clinical Treatment. Hum. Reprod. Update 2011, 17, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Ujvari, D.; Jakson, I.; Babayeva, S.; Salamon, D.; Rethi, B.; Gidlöf, S.; Hirschberg, A.L. Dysregulation of In Vitro Decidualization of Human Endometrial Stromal Cells by Insulin via Transcriptional Inhibition of Forkhead Box Protein O1. PLoS ONE 2017, 12, e0171004. [Google Scholar] [CrossRef]
- Park, C.W.; Choi, M.H.; Koong, M.K.; Kim, H.O.; Kim, J.Y.; Yang, K.M. Serum Plasminogen-Activator Inhibitor Type I (PAI-1) Level Is Increased in PCOS Women. Fertil. Steril. 2012, 98, S213–S214. [Google Scholar] [CrossRef]
- Khalaf, W.M.; Akl, S.A.; Ramadan, R.R.; Kamaleldin, M.A.M. Endometrial Thickness and Subendometrial Vascularity in Anovulatory Polycystic Ovarian Syndrome Patients Treated by Metformin. QJM Int. J. Med. 2020, 113, hcaa056.039. [Google Scholar] [CrossRef]
- Geng, X.; Zhao, J.; Huang, J.; Li, S.; Chu, W.; Wang, W.-S.; Chen, Z.-J.; Du, Y. Lnc-MAP3K13-7:1 Inhibits Ovarian GC Proliferation in PCOS via DNMT1 Downregulation-Mediated CDKN1A Promoter Hypomethylation. Mol. Ther. 2021, 29, 1279–1293. [Google Scholar] [CrossRef]
- Otsuka, M.; Zheng, M.; Hayashi, M.; Lee, J.-D.; Yoshino, O.; Lin, S.; Han, J. Impaired microRNA Processing Causes Corpus Luteum Insufficiency and Infertility in Mice. J. Clin. Investig. 2008, 118, 1944–1954. [Google Scholar] [CrossRef]
- Nagaraja, A.K.; Andreu-Vieyra, C.; Franco, H.L.; Ma, L.; Chen, R.; Han, D.Y.; Zhu, H.; Agno, J.E.; Gunaratne, P.H.; DeMayo, F.J.; et al. Deletion of Dicer in Somatic Cells of the Female Reproductive Tract Causes Sterility. Mol. Endocrinol. 2008, 22, 2336–2352. [Google Scholar] [CrossRef]
- Kim, Y.S.; Kim, H.-R.; Kim, H.; Yang, S.C.; Park, M.; Yoon, J.A.; Lim, H.J.; Hong, S.-H.; DeMayo, F.J.; Lydon, J.P.; et al. Deficiency in DGCR8-Dependent Canonical microRNAs Causes Infertility Due to Multiple Abnormalities during Uterine Development in Mice. Sci. Rep. 2016, 6, 20242. [Google Scholar] [CrossRef] [PubMed]
- Tu, J.; Cheung, A.H.-H.; Chan, C.L.-K.; Chan, W.-Y. The Role of microRNAs in Ovarian Granulosa Cells in Health and Disease. Front. Endocrinol. 2019, 10, 174. [Google Scholar] [CrossRef]
- Mu, L.; Sun, X.; Tu, M.; Zhang, D. Non-Coding RNAs in Polycystic Ovary Syndrome: A Systematic Review and Meta-Analysis. Reprod. Biol. Endocrinol. 2021, 19, 10. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liu, Y.; Lv, M.; Xing, Q.; Zhang, Z.; He, X.; Xu, Y.; Wei, Z.; Cao, Y. MiR-323-3p Regulates the Steroidogenesis and Cell Apoptosis in Polycystic Ovary Syndrome (PCOS) by Targeting IGF-1. Gene 2019, 683, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liu, J.; Lei, Q.; Han, H.; Liu, W.; Cunwei, T.; Li, F.; Cao, D. Transcriptome Analysis of the Chicken Follicular Theca Cells with MiR-135a-5p Suppressed. G3 2020, 10, 4071–4081. [Google Scholar] [CrossRef]
- Lu, T.; Zou, X.; Liu, G.; Deng, M.; Sun, B.; Guo, Y.; Liu, D.; Li, Y. A Preliminary Study on the Characteristics of microRNAs in Ovarian Stroma and Follicles of Chuanzhong Black Goat during Estrus. Genes 2020, 11, 970. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.K.M.; Wood, J.R.; Stewart, D.R.; Ewens, K.; Ankener, W.; Wickenheisser, J.; Nelson-Degrave, V.; Zhang, Z.; Legro, R.S.; Dunaif, A.; et al. Increased Transcription and Increased Messenger Ribonucleic Acid (MRNA) Stability Contribute to Increased GATA6 MRNA Abundance in Polycystic Ovary Syndrome Theca Cells. J. Clin. Endocrinol. Metab. 2005, 90, 6596–6602. [Google Scholar] [CrossRef]
- Yen, H.-W. Selective Alterations in Insulin Receptor Substrates-1, -2 and -4 in Theca but Not Granulosa Cells from Polycystic Ovaries. Mol. Hum. Reprod. 2004, 10, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Xu, P.; Wang, J.; Zhang, C. The Role of miRNA in Polycystic Ovary Syndrome (PCOS). Gene 2019, 706, 91–96. [Google Scholar] [CrossRef]
- Sewer, M.B.; Jagarlapudi, S. Complex Assembly on the Human CYP17 Promoter. Mol. Cell. Endocrinol. 2009, 300, 109–114. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gambioli, R.; Montanino Oliva, M.; Nordio, M.; Chiefari, A.; Puliani, G.; Unfer, V. New Insights into the Activities of D-Chiro-Inositol: A Narrative Review. Biomedicines 2021, 9, 1378. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, A.E.; Udesen, P.B.; Wissing, M.L.; Englund, A.L.M.; Dalgaard, L.T. MicroRNAs Related to Androgen Metabolism and Polycystic Ovary Syndrome. Chem. -Biol. Interact. 2016, 259, 8–16. [Google Scholar] [CrossRef]
- Salas-Huetos, A.; James, E.R.; Aston, K.I.; Jenkins, T.G.; Carrell, D.T.; Yeste, M. The Expression of miRNAs in Human Ovaries, Oocytes, Extracellular Vesicles, and Early Embryos: A Systematic Review. Cells 2019, 8, 1564. [Google Scholar] [CrossRef]
- Khan, H.L.; Bhatti, S.; Abbas, S.; Kaloglu, C.; Isa, A.M.; Younas, H.; Ziders, R.; Khan, Y.L.; Hassan, Z.; Turhan, B.O.; et al. Extracellular microRNAs: Key Players to Explore the Outcomes of in Vitro Fertilization. Reprod. Biol. Endocrinol. 2021, 19, 72. [Google Scholar] [CrossRef]
- Diamanti-Kandarakis, E.; Dunaif, A. Insulin Resistance and the Polycystic Ovary Syndrome Revisited: An Update on Mechanisms and Implications. Endocr. Rev. 2012, 33, 981–1030. [Google Scholar] [CrossRef]
- Reyes-Muñoz, E.; Ortega-González, C.; Martínez-Cruz, N.; Arce-Sánchez, L.; Estrada-Gutierrez, G.; Moran, C.; Sánchez-Serrano, A.P.; Higareda-Sánchez, R.; de la Jara-Díaz, J.F. Association of Obesity and Overweight with the Prevalence of Insulin Resistance, Pre-Diabetes and Clinical–Biochemical Characteristics among Infertile Mexican Women with Polycystic Ovary Syndrome: A Cross-Sectional Study. BMJ Open 2016, 6, e012107. [Google Scholar] [CrossRef]
- Czech, M.P. Mechanisms of Insulin Resistance Related to White, Beige, and Brown Adipocytes. Mol. Metab. 2020, 34, 27–42. [Google Scholar] [CrossRef]
- Abdalla, M.; Deshmukh, H.; Atkin, S.L.; Sathyapalan, T. MiRNAs as a Novel Clinical Biomarker and Therapeutic Targets in Polycystic Ovary Syndrome (PCOS): A Review. Life Sci. 2020, 259, 118174. [Google Scholar] [CrossRef]
- Kurylowicz, A. MicroRNAs in Human Adipose Tissue Physiology and Dysfunction. Cells 2021, 10, 3342. [Google Scholar] [CrossRef]
- Chadt, A.; Al-Hasani, H. Glucose Transporters in Adipose Tissue, Liver, and Skeletal Muscle in Metabolic Health and Disease. Pflug. Arch.—Eur. J. Physiol. 2020, 472, 1273–1298. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Ono, K.; Nishi, H.; Iwanaga, Y.; Nagao, K.; Kinoshita, M.; Kuwabara, Y.; Takanabe, R.; Hasegawa, K.; Kita, T.; et al. MicroRNA-133 Regulates the Expression of GLUT4 by Targeting KLF15 and Is Involved in Metabolic Control in Cardiac Myocytes. Biochem. Biophys. Res. Commun. 2009, 389, 315–320. [Google Scholar] [CrossRef]
- Ciaraldi, T.P.; Aroda, V.; Mudaliar, S.; Chang, R.J.; Henry, R.R. Polycystic Ovary Syndrome Is Associated with Tissue-Specific Differences in Insulin Resistance. J. Clin. Endocrinol. Metab. 2009, 94, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Karolina, D.S.; Armugam, A.; Tavintharan, S.; Wong, M.T.K.; Lim, S.C.; Sum, C.F.; Jeyaseelan, K. MicroRNA 144 Impairs Insulin Signaling by Inhibiting the Expression of Insulin Receptor Substrate 1 in Type 2 Diabetes Mellitus. PLoS ONE 2011, 6, e22839. [Google Scholar] [CrossRef]
- Barber, T.M.; Hanson, P.; Weickert, M.O.; Franks, S. Obesity and Polycystic Ovary Syndrome: Implications for Pathogenesis and Novel Management Strategies. Clin. Med. Insights Reprod. Health 2019, 13, 1179558119874042. [Google Scholar] [CrossRef] [PubMed]
- Zeng, N.; Huang, R.; Li, N.; Jiang, H.; Li, R.; Wang, F.; Chen, W.; Xia, M.; Wang, Q. MiR-451a Attenuates Free Fatty Acids–Mediated Hepatocyte Steatosis by Targeting the Thyroid Hormone Responsive Spot 14 Gene. Mol. Cell. Endocrinol. 2018, 474, 260–271. [Google Scholar] [CrossRef]
- Goedeke, L.; Wagschal, A.; Fernández-Hernando, C.; Näär, A.M. MiRNA Regulation of LDL-Cholesterol Metabolism. Biochim. Biophys. Acta 2016, 1861, 2047–2052. [Google Scholar] [CrossRef]
- Arancio, W.; Calogero Amato, M.; Magliozzo, M.; Pizzolanti, G.; Vesco, R.; Giordano, C. Serum miRNAs in Women Affected by Hyperandrogenic Polycystic Ovary Syndrome: The Potential Role of MiR-155 as a Biomarker for Monitoring the Estroprogestinic Treatment. Gynecol. Endocrinol. 2018, 34, 704–708. [Google Scholar] [CrossRef]
- Bao, B.; Wang, Z.; Ali, S.; Ahmad, A.; Azmi, A.S.; Sarkar, S.H.; Banerjee, S.; Kong, D.; Li, Y.; Thakur, S.; et al. Metformin Inhibits Cell Proliferation, Migration and Invasion by Attenuating CSC Function Mediated by Deregulating miRNAs in Pancreatic Cancer Cells. Cancer Prev. Res. 2012, 5, 355–364. [Google Scholar] [CrossRef]
- Capuani, B.; Pacifici, F.; Della-Morte, D.; Lauro, D. Glucagon Like Peptide 1 and microRNA in Metabolic Diseases: Focusing on GLP1 Action on miRNAs. Front. Endocrinol. 2018, 9, 719. [Google Scholar] [CrossRef]
- Coleman, C.B.; Lightell, D.J.; Moss, S.C.; Bates, M.; Parrino, P.E.; Woods, T.C. Elevation of MiR-221 and -222 in the Internal Mammary Arteries of Diabetic Subjects and Normalization with Metformin. Mol. Cell. Endocrinol. 2013, 374, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Deswal, R.; Dang, A.S. Dissecting the Role of Micro-RNAs as a Diagnostic Marker for Polycystic Ovary Syndrome: A Systematic Review and Meta-Analysis. Fertil. Steril. 2020, 113, 661–669.e2. [Google Scholar] [CrossRef] [PubMed]
- Radbakhsh, S.; Sathyapalan, T.; Banach, M.; Sahebkar, A. Incretins and microRNAs: Interactions and Physiological Relevance. Pharmacol. Res. 2020, 153, 104662. [Google Scholar] [CrossRef]
- Hoeger, K.M.; Dokras, A.; Piltonen, T. Update on PCOS: Consequences, Challenges, and Guiding Treatment. J. Clin. Endocrinol. Metab. 2021, 106, e1071–e1083. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Zhou, M.; Zhou, G.; Zhu, Q.; Li, W. Effect of Metformin on Adiponectin in PCOS: A Meta-Analysis and a Systematic Review. Eur. J. Obstet. Gynecol. Reprod. Biol. 2021, 267, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Li, S.; Chang, Y.; Fang, C.; Liu, H.; Zhang, X.; Wang, Y. Effect of Metformin Treatment during Pregnancy on Women with PCOS: A Systematic Review and Meta-Analysis. CIM 2016, 39, 120. [Google Scholar] [CrossRef]
- Ali, D.-E.-S.; Shah, M.; Ali, A.; Malik, M.O.; Rehman, F.; Badshah, H.; Ehtesham, E.; Vitale, S.G. Treatment with Metformin and Combination of Metformin Plus Pioglitazone on Serum Levels of IL-6 and IL-8 in Polycystic Ovary Syndrome: A Randomized Clinical Trial. Horm. Metab. Res. 2019, 51, 714–722. [Google Scholar] [CrossRef]
- Noren Hooten, N.; Martin-Montalvo, A.; Dluzen, D.F.; Zhang, Y.; Bernier, M.; Zonderman, A.B.; Becker, K.G.; Gorospe, M.; Cabo, R.; Evans, M.K. Metformin-mediated Increase in DICER1 Regulates microRNA Expression and Cellular Senescence. Aging Cell 2016, 15, 572–581. [Google Scholar] [CrossRef]
- Angioni, S.; Sanna, S.; Magnini, R.; Melis, G.B.; Fulghesu, A.M. The Quantitative Insulin Sensitivity Check Index Is Not Able to Detect Early Metabolic Alterations in Young Patients with Polycystic Ovarian Syndrome. Gynecol. Endocrinol. 2011, 27, 468–474. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Author, Year | Study Type | Sample Size | Age (Years) PCOS/Controls | Main Results Dysregulated miRNAs (Upregulated/Downregulated) | Detected in Cell/Tissue |
|---|---|---|---|---|---|
| Cai et al., 2017 [28] | Case-control study | N = 50 (25 married women with PCOS and 25 controls) | N/A | Downregulated: miR-145 | Granulosa cells |
| Chen et al., 2013 [29] | Case-control study | N = 41 (20 control and 21 PCOS) | 27.46 ± 4.07/ 32.41 ± 6.61 | Upregulated: miR-93, 133 and 223 | Adipose tissue |
| Ding et al., 2015 [30] | Case-control study | Screening cohort (N = 18, 9 PCOS, 9 controls) Verification cohort (N = 18, 9 PCOS, 9 controls) | 27.9 ± 4.3/ 28.7 ± 5.2 | Upregulated: miR-5706, let-7i-3 pm, 4463, 3665 and 638 Downregulated: miR-124-3p, 128, 29a-3p and let-7c | Serum |
| Ebrahimi et al., 2018 [31] | Case-control study | N = 372 (180 with PCOS, 192 controls) | 26.8 ± 5.5/ 27.0 ± 4.38 | Upregulated: miR-146a | Whole blood |
| Eisenberg et al., 2017 [32] | Case-control study | N = 40 (7 normally ovulating, 15 normally ovulating with pure male infertility factor, and 18 with PCOS) | 26.9 ± 4.3/ 26.8 ± 4.7 | Upregulated: miR-200b and 429 | Serum |
| Geng et al., 2019 [33] | Case-control study | N = 30 (15 married women with PCOS and 15 controls) | 27.23 ± 1.83/ 28.53 ± 1.85 | Upregulated: miR-99a | Granulosa cells |
| He et al., 2018 [34] | Case-control study | N = 123 (62 with PCOS, 61 controls) | 28.27 ± 3.10/ 28.71 ± 2.46 | Downregulated: miR-141 and 200c | Granulosa cells |
| Hosseini et al., 2017 [35] | Case-control study | N = 410 (205 with PCOS, 205 controls) | 31.2 ± 5.5/ 28.5 ± 5.0 | Upregulated: miR-146a and 222 | Plasma |
| Hou et al., 2019 [36] | Case-control study | N = 28 (15 with PCOS, 13 controls) | 29.60 ± 0.66/ 29.66 ± 0.82 | Upregulated: miR-3188 and 3135b | Granulosa cells |
| Hu et al., 2020 [37] | Case-control study | N/A | N/A | Upregulated: miR-6087, 199a-5p, 1433p, 483-5p, 200a-3p, and 23b-3p | Follicular fluid |
| Huang et al., 2016 [38] | Case-control study | N = 36 (18 with PCOS, 18 controls) | 32.6 ± 3.1/ 34.6 ± 2.2 | Upregulated: miR-135b-5p, 152, 193a-3p, 194-5p, 196a-5p, 200b-3p, 423-3p, 454-3p, 455-5p, 4659a-3p, 509–3-5p, 509-3p, 513b-5p, 652-5p, 95, 1273e | Cumulus cells |
| Jiang et al., 2016 [39] | Case-control study | N = 175 (65 with PCOS and IGM; 65 with PCOS without IGM; 45 healthy controls) | 27.16 ± 3.56/ 27.98 ± 3.66 | Upregulated: miR-122, 194, and 193b Downregulated: miR-199b-5p | Serum |
| Li et al., 2019 [40] | Case-control study | N = 78 (46 with PCOS, 32 controls) | 29.21 ± 4.78/ 29.43 ± 3.82 | Upregulated: miR-33b and 142 Downregulated: miR-423 | Granulosa cells |
| Lin et al., 2015 [41] | Case-control study | N = 18 (10 with PCOS, 8 controls) | 28.80 ± 3.97/ 32.00 ± 2.16 | Downregulated: miR-19b, 92a, 92b, 141, and 200a | Ovarian theca internal tissues |
| Linlin Jiang, 2015 [42] | Case-control study | N = 24 (16 with PCOS, 8 controls) | 29.69 ± 2.39/ 31.75 ± 4.40 | Upregulated: miR-93, 107 | Granulosa cells |
| Liu et al., 2015 [43] | Case-control study | N = 20 (10 with PCOS, 10 controls) | 27.4 ± 2.6/ 29.4 ± 3.0 | Upregulated: miR-513a-3p, 508-3p, 513b, 514, 509-5p, 513c, 144, 510, 509-3p and 508-5p Downregulated: miR-151-3p, 720, 615-3p, 127-3p, 455-3p, 342-3p and 654-3p | Cumulus cells |
| Liyan Jiang et al., 2015 [44] | Case-control study | N/A | N/A | Upregulated: miR21,222,16,19a,30c, 146a, 24 and 186 | Serum |
| Long et al., 2014 [45] | Multistage restrospective, nested case-control study | N = 136 (68 with PCOS, 68 controls) | 26.6 ± 2.8/ 27.9 ± 3.4 | Upregulated: miR-222, 146a and 30c | Serum |
| Luo et al., 2019 [46] | Case-control study | N = 12 (4 with POR, 4 with PCOS, 4 controls) | PCOS (27.00 ± 3.26) POR (37.00 ± 3.16) Controls (29.00 ± 3.22) | Upregulated: miR-23a | Granulosa cells |
| Mao et al., 2018 [47] | Case-control study | N = 69 (43 with PCOS, 26 controls) | 30.2 ± 2.8/ 31.1 ± 2.1 | Downregulated: miR-126-5p and 29a-5p | Granulosa cells |
| McAllister et al., 2019 [48] | Case-control study | N = 14 (7 with PCOS, 7 controls) | N/A | Upregulated: miR-100-5p, 99b-5p, 1271-5p, 409-5p, 744, 410-3p, 127-3p, 654-5p, 494-3p, 1301-3p, 502-3p, 501-3p and 1293 Downregulated: miR-125a-3p,148b-5p, 195-5p,130b-3p and 4542a-5p | Ovarian theca cells |
| McCallie et al., 2010 [49] | Descriptive study | N = 44 | N/A | Downregulated: miR-let-7a, 19a, 19b, 24, 92, and 93 | Blastocysts |
| Murri et al., 2013 [50] | Case-control study | N = 36 (12 with PCOS, 12 healthy women, 12 men) | 27 ± 4/ 29 ± 3 | Upregulated: miR-21,27b, 103 and 155 | Whole blood |
| Murri et al., 2018 [51] | Case-control study | N = 35 (12 with PCOS, 11 healthy women, 12 men) | 27 ± 4/ 28 ± 3 | Upregulated: miR-34c-5p and 548d-3p Downregulated: miR-26a-5p, 30c-5p, 107 and 199a-3p | Serum |
| Naji et al., 2017 [52] | Case-control study | N = 66 (19 with normoandrogenic PCOS, 22 with hyperandrogenic PCOS, 25 controls) | 28.89 ± 1.07/ 28.24 ± 0.82 | Upregulated in granulosa cells: miR-93 Downregulated in follicular fluid: miR-93 and 21 | Follicular fluid, granulosa cells |
| Naji et al., 2018 [53] | Case-control study | N = 41 (20 with PCOS, 21 controls) | 29.25 ± 0.84/ 28.42 ± 0.91 | Upregulated in follicular fluid: miR-182 Downregulated in granulosa-lutein cells: miR-145 and 182 | Serum, granulosa-lutein cells, follicular fluid |
| Nanda et al., 2020 [54] | Case-control study | N = 40 (20 with PCOS, 20 controls) | 28.35 ± 7.45/ 25.15 ± 4.12 | Upregulated: miR-122, 194, and 193b Downregulated: miR-199b-5p | Serum |
| Rashad et al., 2019 [55] | Case-control study | N = 100 (60 with PCOS, 40 controls) | N/A | Downregulated: miR-320 | Serum |
| Roth et al., 2014 [56] | Case-control study | N/A | 33.1 ± 4.4/ 27.1 ± 3.6 | Upregulated: miR-32, 34c, 135a, 18b, and 9 | Follicular fluid |
| Sang et al., 2013 [57] | Case-control study | N = 44 (22 with PCOS, 22 controls) | 29.09 ± 0.70/ 30.83 ± 0.90 | Downregulated: miR-132 and 320 | Follicular fluid |
| Sathyapalan et al., 2015 [58] | Case-control study | N = 49 (25 with PCOS, 24 controls) | 32.1 ± 9.0/ 32.2 ± 7.7 | Upregulated: miR-93 and 223 | Plasma |
| Scalici et al., 2016 [59] | Case-control study | N = 121 (30 with PCOS, 91 controls) | Mean age for cohort 33.7 ± 4.5 | Upregulated: miR-30a Downregulated: miR-140 and let-7b | Follicular fluid |
| Shi et al., 2015 [60] | Case-control study | N = 48 (24 with PCOS, 24 controls) | 28.3 ± 3.3/ 28.5 ± 3.6 | Downregulated: miR-483–5p and 486–5p | Cumulus cells |
| Song et al., 2015 [61] | Case-control study | N = 134 (67 with PCOS, 67 controls) | 26.7 ± 2.7/ 27.6 ± 3.3 | Downregulated: miR-592,124-3p, 128, 29-3p, 16, 106b, 19a, 24, 186, let-7c and 1228 | Serum |
| Song et al., 2016 [62] | Case-control study | N = 42 (21 with PCOS, 21 controls) with preceded pilot study (N = 17) | 23 ± 4/ 24 ± 6 | Downregulated: miR-4522, 324-3p, and 6767-5p | Serum |
| Song et al., 2019 [63] | Case-control study | N = 83 (63 with PCOS, 20 controls) | 28.21 ± 2.78/ 27.43 ± 3.62 | Upregulated: miR-186 and 135a | Granulosa cells |
| Sørensen et al., 2016 [64] | Case-control study | N = 70 49 PCOS women (19 of which were hyperandrogenic and 30 normo-androgenic) and 21 healthy matched women | 28.1 ± 4.3/ 27.8 ± 3.8 | Upregulated: miR-518f-3p, Downregulated: miR-24-3p, -29a, -151-3p, and -574-3p | Follicular fluid |
| Sørensen et al., 2016 [65] | Case-control study | N = 62 42 PCOS patients and 20 Controls | 27.0 ± 7.5/ 27.0 ± 6.3 | Upregulated: miR-485-3p, miR-1290, and miR-7-1-3p Downregulated: miR-21-3p, miR-139-3p, miR -572, miR-361-5p, miR-143-3p, miR-345-5p, miR-1276, and miR-22-5p | Follicular fluid |
| Wang et al., 2018 [66] | Case-control study | N = 30 17 PCOS patients and 13 controls | 28.7 ± 0.7/ 30.0 ± 0.7 | Upregulated: miR-27a-3p | Granulosa cells |
| Wang et al., 2019 [67] | Case-control study | N = 45 24 PCOS patients and 21 controls | 28.7 ± 0.8/ 29.6 ± 1.0 | Upregulated: miR-3188 and 3135b | Granulosa cells |
| Wu et al., 2014 [68] | Case-control study | N = 31 16 women with PCOS (8 with and 8 without IR) and 15 non-PCOS (9 with and 6 without IR). | 32.33 ± 5.03/ 25.49 ± 5.56 | Upregulated: miR-93, and 25 | Adipose tissue |
| Xiang et al., 2016 [69] | Case-control study | N = 40 (20 with PCOS, 20 controls) | 27.3 ± 2.5/ 28.2 ± 3.7 | Downregulated: miR-483 | Ovarian cortex |
| Xiong et al., 2017 [70] | Case-control study | N = 48 (18 with PCOS, 30 controls) | N/A | Downregulated: miR-23a and 23b | Serum |
| Xu et al., 2015 [71] | Case-control study | N = 41 (21 with PCOS, 20 controls) | N/A | Upregulated: miR-423-3p, 3651, 3653, 151b, 1273 g-3p, 590-5p, 3648, 7845-5p, 27a-5p, 1275, 483-3p, 7-5p, 483-5p, 10a-5p, 184, 619-5p, 513b-5p, 1307-5p, 4516, 1307-3p, 514b-5p Downregulated: miR-3529-3p, 7974, 3065-5p, 214-3p, 200a-3p, 203a, 4732-5p, 423-5p, 3184-5p, 548n, 221-3p, 149-5p, 1298-5p, 193a-3p, 365a-3p, 219a-1-3p, 550b-2-5p, 144-5p, 660-5p, 548e-3p, 652-3p, 222-3p,506-5p, 193a-5p, 210-5p, 365b-5p, 330-3p, 223-3p, 186-5p, 185-5p, 92b-3p, 199b-3p, 766-5p, 15b-3p, 339-5p, 3960, 766-3p, let-7a-3p | Cumulus granulosa cells |
| Xue et al., 2018 [72] | Case-control study | N = 6 (3 with PCOS, 3 controls) | N/A | Upregulated: miR-200a-3p, 10b-3p, 200b-3p,29c-3p, 99a-3p and 125a-5p Downregulated: miR-105-3p | Follicular fluid |
| Yao et al., 2018 [73] | Prospective, observational study | Female Sprague–Dawley rats (23 days old) | N/A | Downregulated: miR-335-5p | Follicular fluid |
| Yao et al., 2018 [74] | Case-control study | N = 106 (55 with PCOS, 51 controls) | 28.13 ± 0.41/ 27.37 ± 0.46 | Downregulated: miR-335-5p | Follicular fluid |
| Yin et al., 2014 [75] | Prospective, observational study | Mice ovaries | N/A | Upregulated: miR-320 and miR-383 | Follicular fluid, granulosa cells |
| Zhang et al., 2017 [76] | Case-control study | N = 33 (21 with PCOS, 12 controls) | N/A | Downregulated: miR-320a | Cumulus cells |
| Zhang et al., 2018 [77] | Case-control study | N = 40 (20 with PCOS, 20 controls) | N/A | Upregulated: miR-873-5p | Follicular fluid |
| Zhao et al., 2015 [78] | Multistage retrospective nested case-control study | N = 384 patients with PCOS; 30 with developed OHSS after IVF/ICSI and 70 controls | 27.93 ± 3.84/ 27.70 ± 3.44 | Upregulated: miR-146a, 30c and 191 Downregulated: miR-16, 223, 212, 451 and 92a | Serum |
| Zhong et al., 2018 [79] | Case-control study | N = 28 (18 with PCOS, 10 controls) | N/A | Downregulated: miR-19b | Granulosa cells, ovarian cortex |
| miRNA | Species | Reported Biological Role | |
|---|---|---|---|
| miR-9 | Human | Inhibits testosterone release [56] | |
| miR-18b | Human | Inhibits testosterone and estradiol release [56] Promotes progesterone release [56] | |
| miR-20a | Human | Postively correlated with hyperandrogenism (increase of either total testosterone, free testosterone, DHEAS or androstenedione) [38] | |
| miR-21 | Human Rat | Anti-apoptotic properties; involved in oocyte maturation [53] | |
| miR-24 | Human | Inverse correlation with insulin, LH, testosterone and the LH:FSH ratio [44] | |
| miR-27b | Human | Positively correlated with testosterone; increased expression in PCOS [50] | |
| miR-29a | Human | Inveresely associated with LH and insulin; positively correlated with DHEA and androstendione [54,80] | |
| miR-30c | Human Rat | Increased expression after FSH exposure (positive association) [44] | |
| miR-34b-3p | Human | Negative association with with hyperandrogenism (either total testosterone, free testosterone, DHEAS or androstenedione) [51] | |
| miR-103 | Human | Promotes progesterone release and inhibits estradiol release [50] | |
| miR-107 | Human | Increases testosterone release [51] | |
| miR-132 | Human Rat | Increases estradiol secretion [57] Reduces progesterone and testosterone release [57] | |
| miR-135a | Human | Reduces progesterone and testosterone release [63,81] | |
| miR-139 | Human | Negative association with with hyperandrogenism (either total testosterone, free testosterone, DHEAS or androstenedione) [65] | |
| miR-146a | Human | Reduces progesterone, estradiol and testosterone release [31] | |
| miR-151 | Human | Postively associated with total testosterone and free testosterone [43] | |
| miR-155 | Human | Inhibits testosterone release [50] | |
| miR-222 | Human | Increases estradiol secretion [35] | |
| miR-224 | Human Mouse | Induces granulosa cells proliferation [82] Increases estradiol release [82] | |
| miR-320 | Human | Increases testosterone release [75,76] | |
| miR-361 | Human | Positive correlation with SHBG levels [65] | |
| miR-383 | Human | Increases estradiol secretion [75] | |
| miR-433 | Human | Positive correlation with SHBG levels [37] | |
| miR-518 | Human | Positively correlated with total and free testosterone, DHEA-S and androstendione [64] | |
| miR-1225 | Human | Negatively associated with serum insulin and HOMA-IR [83] | |
| miR-1290 | Human | Postively correlated with hyperandrogenism (increase of either total testosterone, free testosterone, DHEAS or androstenedione) [65] | |
| Study | Selection | Comparability | Outcome | Overall |
|---|---|---|---|---|
| Cai et al., 2017 [28] | *** | ** | ** | 7 |
| Chen et al., 2013 [29] | *** | ** | *** | 8 |
| Ding et al., 2015 [30] | *** | *** | *** | 9 |
| Ebrahimi et al., 2018 [31] | *** | * | ** | 6 |
| Eisenberg et al., 2017 [32] | *** | ** | *** | 8 |
| Geng et al., 2019 [33] | *** | ** | ** | 7 |
| He et al., 2018 [34] | *** | *** | ** | 8 |
| Hosseini et al., 2017 [35] | ** | ** | ** | 6 |
| Hou et al., 2019 [36] | *** | ** | ** | 7 |
| Hu et al., 2020 [37] | *** | * | *** | 7 |
| Huang et al., 2016 [38] | *** | ** | *** | 8 |
| Jiang et al., 2016 [39] | *** | *** | *** | 9 |
| Li et al., 2019 [40] | *** | ** | ** | 7 |
| Lin et al., 2015 [41] | *** | *** | ** | 8 |
| Linlin Jiang, 2015 [42] | ** | ** | *** | 7 |
| Liu et al., 2015 [43] | *** | ** | *** | 8 |
| Liyan Jiang et al., 2015 [44] | ** | ** | ** | 6 |
| Long et al., 2014 [45] | *** | *** | *** | 9 |
| Luo et al., 2019 [46] | ** | ** | *** | 7 |
| Mao et al., 2018 [47] | *** | *** | ** | 8 |
| McAllister et al., 2019 [48] | ** | ** | *** | 7 |
| McCallie et al., 2010 [49] | ** | ** | *** | 7 |
| Murri et al., 2013 [50] | *** | *** | ** | 8 |
| Murri et al., 2018 [51] | *** | *** | *** | 9 |
| Naji et al., 2017 [52] | *** | *** | *** | 9 |
| Naji et al., 2018 [53] | *** | *** | *** | 9 |
| Nanda et al., 2020 [54] | ** | *** | ** | 7 |
| Rashad et al., 2019 [55] | ** | ** | ** | 6 |
| Roth et al., 2014 [56] | ** | ** | *** | 7 |
| Sang et al., 2013 [57] | *** | *** | ** | 8 |
| Sathyapalan et al., 2015 [58] | *** | *** | ** | 8 |
| Scalici et al., 2016 [59] | *** | *** | *** | 9 |
| Shi et al., 2015 [60] | *** | ** | ** | 7 |
| Song et al., 2015 [61] | *** | *** | ** | 8 |
| Song et al., 2016 [62] | *** | *** | ** | 8 |
| Song et al., 2019 [63] | *** | *** | ** | 8 |
| Sørensen et al., 2016 [64] | *** | *** | *** | 9 |
| Sørensen et al., 2016 [65] | *** | *** | *** | 9 |
| Wang et al., 2018 [66] | *** | *** | ** | 8 |
| Wang et al., 2019 [67] | *** | *** | ** | 8 |
| Wu et al., 2014 [68] | *** | *** | ** | 8 |
| Xiang et al., 2016 [69] | *** | ** | ** | 7 |
| Xiong et al., 2017 [70] | *** | ** | ** | 7 |
| Xu et al., 2015 [71] | *** | ** | *** | 8 |
| Xue et al., 2018 [72] | ** | ** | ** | 6 |
| Yao et al., 2018 [73] | ** | ** | ** | 6 |
| Yao et al., 2018 [74] | *** | ** | ** | 7 |
| Yin et al., 2014 [75] | ** | *** | ** | 7 |
| Zhang et al., 2017 [76] | ** | ** | *** | 7 |
| Zhang et al., 2018 [77] | ** | ** | *** | 7 |
| Zhao et al., 2015 [78] | *** | *** | *** | 9 |
| Zhong et al., 2018 [79] | ** | ** | *** | 7 |
| Study | miRNAs as Diagnostic Biomarkers | miRNAs as Therapeutic Targets |
|---|---|---|
| Ali et al. [129] | N/A | miR-26a |
| Capuani et al. [130] | N/A | miR132, miR-212, miR-338, miR-758, miR34a, miR-21, miR-200b, miR-200c |
| Coleman et al. [131] | N/A | miR-221 and miR-222 |
| Deswal et al. [132] | miR-29a-5p and miR-320 | N/A |
| Mu et al. [104] | miR-93 and miR-320 | N/A |
| Radbakhsh et al. [133] | N/A | miR-33, miR-155-5p, miR-197, miR-6356, miR-1197-3p, miR-875-5P and miR-6763 |
| Sathyapalan et al. [58] | miR-93 | N/A |
| Scalici et al. [59] | miR-let-7b, miR-30a and miR-140 | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vitale, S.G.; Fulghesu, A.M.; Mikuš, M.; Watrowski, R.; D’Alterio, M.N.; Lin, L.-T.; Shah, M.; Reyes-Muñoz, E.; Sathyapalan, T.; Angioni, S. The Translational Role of miRNA in Polycystic Ovary Syndrome: From Bench to Bedside—A Systematic Literature Review. Biomedicines 2022, 10, 1816. https://doi.org/10.3390/biomedicines10081816
Vitale SG, Fulghesu AM, Mikuš M, Watrowski R, D’Alterio MN, Lin L-T, Shah M, Reyes-Muñoz E, Sathyapalan T, Angioni S. The Translational Role of miRNA in Polycystic Ovary Syndrome: From Bench to Bedside—A Systematic Literature Review. Biomedicines. 2022; 10(8):1816. https://doi.org/10.3390/biomedicines10081816
Chicago/Turabian StyleVitale, Salvatore Giovanni, Anna Maria Fulghesu, Mislav Mikuš, Rafał Watrowski, Maurizio Nicola D’Alterio, Li-Te Lin, Mohsin Shah, Enrique Reyes-Muñoz, Thozhukat Sathyapalan, and Stefano Angioni. 2022. "The Translational Role of miRNA in Polycystic Ovary Syndrome: From Bench to Bedside—A Systematic Literature Review" Biomedicines 10, no. 8: 1816. https://doi.org/10.3390/biomedicines10081816
APA StyleVitale, S. G., Fulghesu, A. M., Mikuš, M., Watrowski, R., D’Alterio, M. N., Lin, L.-T., Shah, M., Reyes-Muñoz, E., Sathyapalan, T., & Angioni, S. (2022). The Translational Role of miRNA in Polycystic Ovary Syndrome: From Bench to Bedside—A Systematic Literature Review. Biomedicines, 10(8), 1816. https://doi.org/10.3390/biomedicines10081816