
Falling Short: The Contribution of Central Insulin Receptors to Gait Dysregulation in Brain Aging


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Current Therapeutic Approaches
3. Contribution of Peripheral Systems and Special Senses to Gait Dysregulation
3.1. Peripheral Systems
3.2. Special Senses
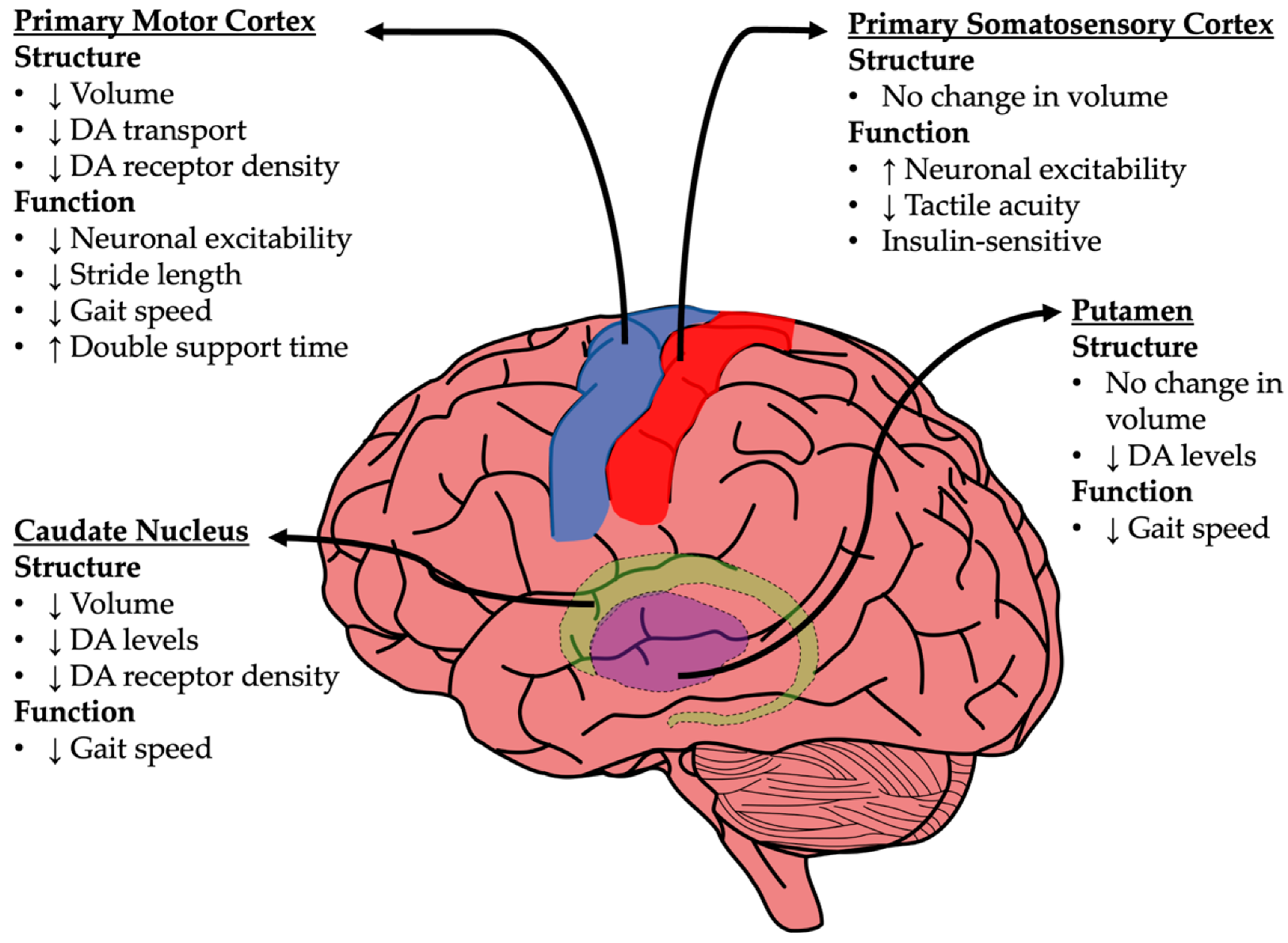
4. Neuroanatomical Changes in Gait Processing Centers
4.1. Basal Ganglia
4.2. Primary Motor Cortex (M1)
4.3. Primary Somatosensory Cortex (S1)
5. The Insulin Receptor as a Potential Novel Therapeutic Target of Gait Dysregulation
5.1. Insulin Receptors in the Brain
5.2. Central Insulin Administration as a Therapeutic Approach
6. Insulin Sensitivity in the Brain: Contradictory Evidence
6.1. Evidence Supporting Brain Insulin Resistance in Aging and AD
6.2. Evidence Supporting Preserved Insulin Sensitivity with Aging
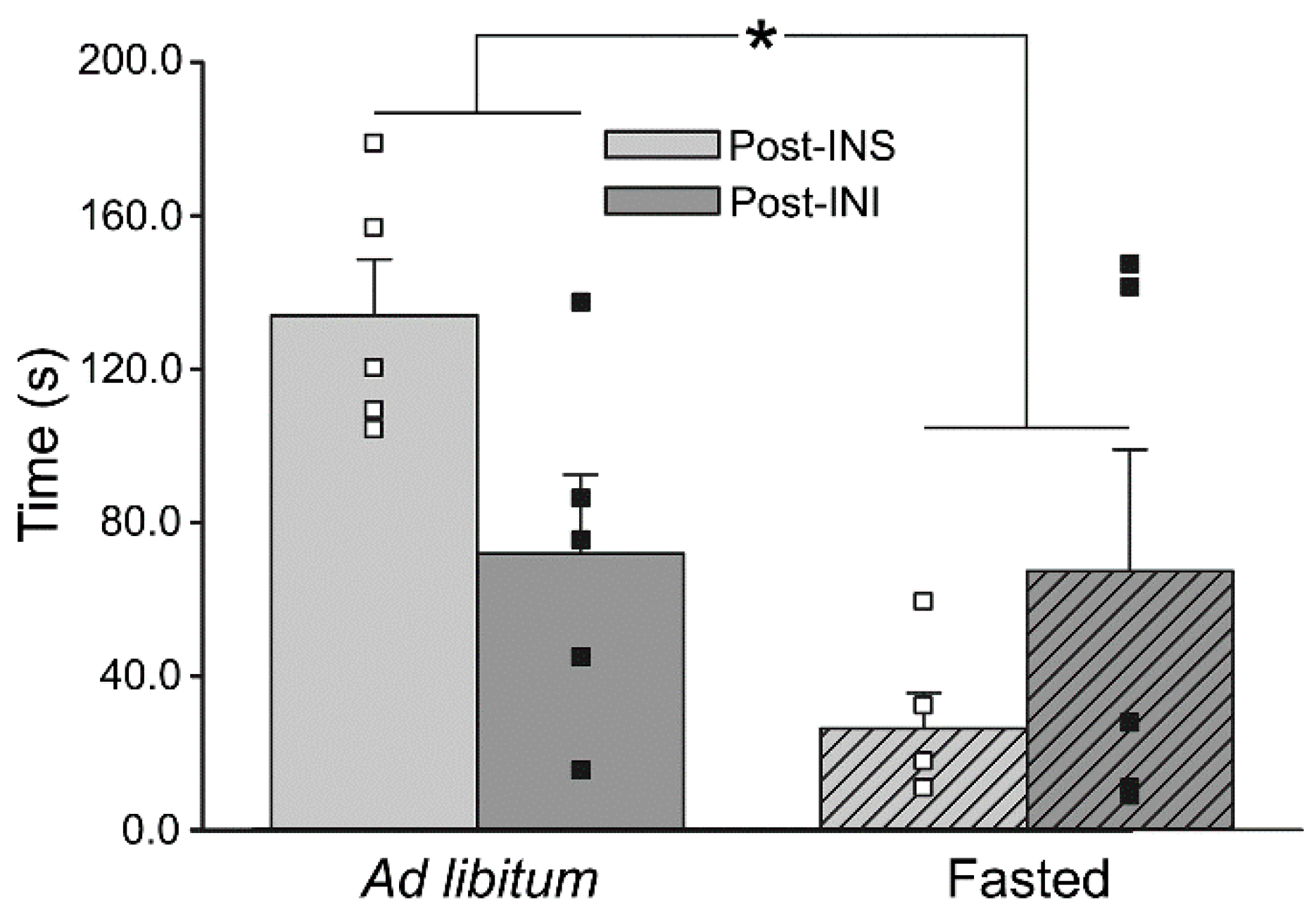
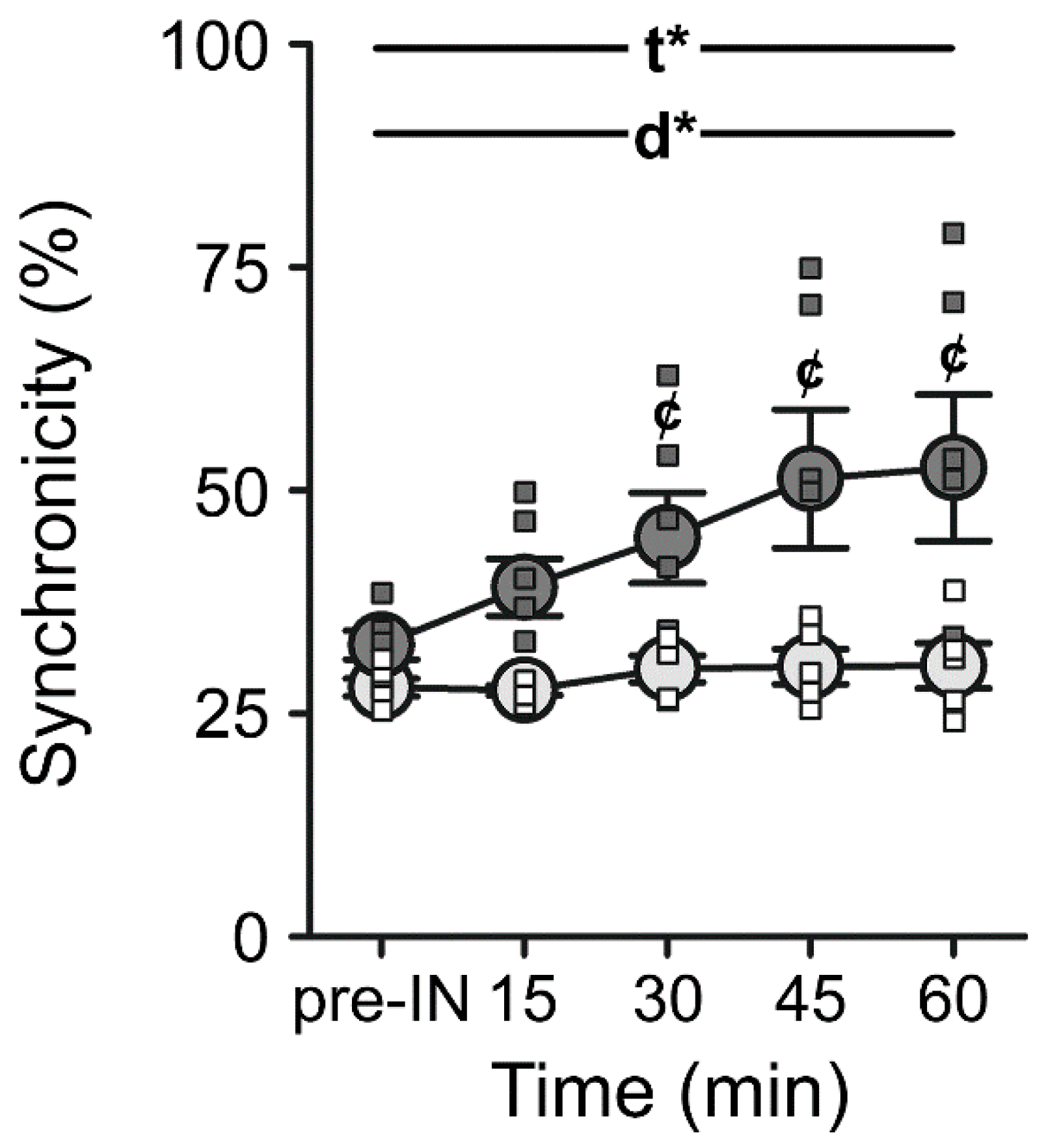
6.3. Could INI Be a Therapeutic Approach for Gait Dysfunction?
7. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bergen, G.; Stevens, M.R.; Burns, E.R. Falls and Fall Injuries Among Adults Aged >/=65 Years-United States, 2014. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 993–998. [Google Scholar] [CrossRef] [PubMed]
- Rubenstein, L.Z.; Josephson, K.R. The epidemiology of falls and syncope. Clin. Geriatr. Med. 2002, 18, 141–158. [Google Scholar] [CrossRef]
- Thapa, P.B.; Gideon, P.; Brockman, K.G.; Fought, R.L.; Ray, W.A. Clinical and biomechanical measures of balance as fall predictors in ambulatory nursing home residents. J. Gerontol. A Biol. Sci. Med. Sci. 1996, 51, M239–M246. [Google Scholar] [CrossRef]
- Tinetti, M.E. Clinical practice. Preventing falls in elderly persons. N. Engl. J. Med. 2003, 348, 42–49. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Public health and aging: Nonfatal injuries among older adults treated in hospital emergency departments-United States, 2001. MMWR Morb. Mortal. Wkly. Rep. 2003, 52, 1019–1022. [Google Scholar]
- Guideline for the prevention of falls in older persons. American Geriatrics Society, British Geriatrics Society, and American Academy of Orthopaedic Surgeons Panel on Falls Prevention. J. Am. Geriatr. Soc. 2001, 49, 664–672. [Google Scholar]
- Lee, H.; Kim, S.H.; Lee, S.C.; Kim, S.; Cho, G.C.; Kim, M.J.; Lee, J.S.; Han, C. Severe Injuries from Low-height Falls in the Elderly Population. J. Korean Med. Sci. 2018, 33, e221. [Google Scholar] [CrossRef]
- Wong, T.H.; Wong, Y.J.; Lau, Z.Y.; Nadkarni, N.; Lim, G.H.; Seow, D.C.C.; Ong, M.E.H.; Tan, K.B.; Nguyen, H.V.; Wong, C.H. Not All Falls Are Equal: Risk Factors for Unplanned Readmission in Older Patients After Moderate and Severe Injury-A National Cohort Study. J. Am. Med. Dir. Assoc. 2019, 20, 201–207.e203. [Google Scholar] [CrossRef] [PubMed]
- Ek, S.; Rizzuto, D.; Fratiglioni, L.; Johnell, K.; Xu, W.; Welmer, A.K. Risk Profiles for Injurious Falls in People Over 60: A Population-Based Cohort Study. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, V.H.; Galet, C.; Lilienthal, M.; Skeete, D.A.; Romanowski, K.S. Association of Clinical Frailty Scores With Hospital Readmission for Falls After Index Admission for Trauma-Related Injury. JAMA Netw. Open 2019, 2, e1912409. [Google Scholar] [CrossRef] [PubMed]
- Ek, S.; Rizzuto, D.; Fratiglioni, L.; Calderon-Larranaga, A.; Johnell, K.; Sjoberg, L.; Xu, W.; Welmer, A.K. Risk Factors for Injurious Falls in Older Adults: The Role of Sex and Length of Follow-Up. J. Am. Geriatr. Soc. 2019, 67, 246–253. [Google Scholar] [CrossRef]
- Aryee, E.; James, S.L.; Hunt, G.M.; Ryder, H.F. Identifying protective and risk factors for injurious falls in patients hospitalized for acute care: A retrospective case-control study. BMC Geriatr. 2017, 17, 260. [Google Scholar] [CrossRef] [PubMed]
- Kusljic, S.; Perera, S.; Manias, E. Age-dependent physiological changes, medicines and sex-influenced types of falls. Exp. Aging Res. 2018, 44, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Close, J.C.; Lord, S.R.; Antonova, E.J.; Martin, M.; Lensberg, B.; Taylor, M.; Hallen, J.; Kelly, A. Older people presenting to the emergency department after a fall: A population with substantial recurrent healthcare use. Emerg. Med. J. 2012, 29, 742–747. [Google Scholar] [CrossRef]
- Gill, T.M.; Murphy, T.E.; Gahbauer, E.A.; Allore, H.G. Association of injurious falls with disability outcomes and nursing home admissions in community-living older persons. Am. J. Epidemiol. 2013, 178, 418–425. [Google Scholar] [CrossRef]
- Rubenstein, L.Z. Falls in older people: Epidemiology, risk factors and strategies for prevention. Age Ageing 2006, 35 (Suppl. 2), ii37–ii41. [Google Scholar] [CrossRef] [PubMed]
- Vieira, E.R.; Palmer, R.C.; Chaves, P.H. Prevention of falls in older people living in the community. BMJ 2016, 353, i1419. [Google Scholar] [CrossRef] [PubMed]
- Sattin, R.W. Falls among older persons: A public health perspective. Annu. Rev. Public Health 1992, 13, 489–508. [Google Scholar] [CrossRef]
- Sattin, R.W.; Lambert Huber, D.A.; DeVito, C.A.; Rodriguez, J.G.; Ros, A.; Bacchelli, S.; Stevens, J.A.; Waxweiler, R.J. The incidence of fall injury events among the elderly in a defined population. Am. J. Epidemiol. 1990, 131, 1028–1037. [Google Scholar] [CrossRef]
- Salkeld, G.; Cumming, R.G.; O’Neill, E.; Thomas, M.; Szonyi, G.; Westbury, C. The cost effectiveness of a home hazard reduction program to reduce falls among older persons. Aust. N. Z. J. Public Health 2000, 24, 265–271. [Google Scholar] [CrossRef]
- Yang, Y.; Hu, X.; Zhang, Q.; Zou, R. Diabetes mellitus and risk of falls in older adults: A systematic review and meta-analysis. Age Ageing 2016, 45, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Pijpers, E.; Ferreira, I.; de Jongh, R.T.; Deeg, D.J.; Lips, P.; Stehouwer, C.D.; Nieuwenhuijzen Kruseman, A.C. Older individuals with diabetes have an increased risk of recurrent falls: Analysis of potential mediating factors: The Longitudinal Ageing Study Amsterdam. Age Ageing 2012, 41, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Crews, R.T.; Yalla, S.V.; Fleischer, A.E.; Wu, S.C. A growing troubling triad: Diabetes, aging, and falls. J. Aging Res. 2013, 2013, 342650. [Google Scholar] [CrossRef]
- Remelli, F.; Ceresini, M.G.; Trevisan, C.; Noale, M.; Volpato, S. Prevalence and impact of polypharmacy in older patients with type 2 diabetes. Aging Clin. Exp. Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Florence, C.S.; Bergen, G.; Atherly, A.; Burns, E.; Stevens, J.; Drake, C. Medical Costs of Fatal and Nonfatal Falls in Older Adults. J. Am. Geriatr. Soc. 2018, 66, 693–698. [Google Scholar] [CrossRef]
- Youkhana, S.; Dean, C.M.; Wolff, M.; Sherrington, C.; Tiedemann, A. Yoga-based exercise improves balance and mobility in people aged 60 and over: A systematic review and meta-analysis. Age Ageing 2016, 45, 21–29. [Google Scholar] [CrossRef]
- Gillespie, L.D.; Robertson, M.C.; Gillespie, W.J.; Sherrington, C.; Gates, S.; Clemson, L.M.; Lamb, S.E. Interventions for preventing falls in older people living in the community. Cochrane Database Syst. Rev. 2012, 2012, CD007146. [Google Scholar] [CrossRef]
- Sherrington, C.; Fairhall, N.J.; Wallbank, G.K.; Tiedemann, A.; Michaleff, Z.A.; Howard, K.; Clemson, L.; Hopewell, S.; Lamb, S.E. Exercise for preventing falls in older people living in the community. Cochrane Database Syst. Rev. 2019, 1, CD012424. [Google Scholar] [CrossRef]
- Esquivel, M.K. Nutritional Assessment and Intervention to Prevent and Treat Malnutrition for Fall Risk Reduction in Elderly Populations. Am. J. Lifestyle Med. 2018, 12, 107–112. [Google Scholar] [CrossRef]
- Medical Advisory, S. Prevention of falls and fall-related injuries in community-dwelling seniors: An evidence-based analysis. Ont. Health Technol. Assess Ser. 2008, 8, 1–78. [Google Scholar]
- Scragg, R.; Waayer, D.; Stewart, A.W.; Lawes, C.M.M.; Toop, L.; Murphy, J.; Khaw, K.T.; Camargo, C.A., Jr. The Vitamin D Assessment (ViDA) Study: Design of a randomized controlled trial of vitamin D supplementation for the prevention of cardiovascular disease, acute respiratory infection, falls and non-vertebral fractures. J. Steroid. Biochem. Mol. Biol. 2016, 164, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Shahar, D.; Levi, M.; Kurtz, I.; Shany, S.; Zvili, I.; Mualleme, E.; Shahar, A.; Sarid, O.; Melzer, I. Nutritional status in relation to balance and falls in the elderly: A preliminary look at serum folate. Ann. Nutr. Metab. 2009, 54, 59–66. [Google Scholar] [CrossRef]
- Smith, D.A. Documented, Systematic and Individualized Communication With the Attending Physician for Fall Risk Reduction/Injury Mitigation Care Planning. J. Am. Med. Dir. Assoc. 2018, 19, 714–716. [Google Scholar] [CrossRef] [PubMed]
- Patil, R.; Kolu, P.; Raitanen, J.; Valvanne, J.; Kannus, P.; Karinkanta, S.; Sievanen, H.; Uusi-Rasi, K. Cost-effectiveness of vitamin D supplementation and exercise in preventing injurious falls among older home-dwelling women: Findings from an RCT. Osteoporos. Int. 2016, 27, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Henderson, E.J.; Lord, S.R.; Brodie, M.A.; Gaunt, D.M.; Lawrence, A.D.; Close, J.C.; Whone, A.L.; Ben-Shlomo, Y. Rivastigmine for gait stability in patients with Parkinson’s disease (ReSPonD): A randomised, double-blind, placebo-controlled, phase 2 trial. Lancet Neurol. 2016, 15, 249–258. [Google Scholar] [CrossRef]
- Beauchet, O.; Launay, C.P.; Allali, G.; Watfa, G.; Gallouj, K.; Herrmann, F.R.; Annweiler, C. Anti-dementia drugs and changes in gait: A pre-post quasi-experimental pilot study. BMC Neurol. 2013, 13, 184. [Google Scholar] [CrossRef]
- Kapur, S.; Vaughan, C.; Hawkins, J.; Stebbins, G.; Hall, D. Varenicline for the Treatment of Postural and Gait Dysfunction in Parkinson Disease. Neurol. Clin. Pract. 2021, 11, 457–461. [Google Scholar] [CrossRef]
- Lecat, M.; Decavel, P.; Magnin, E.; Lucas, B.; Gremeaux, V.; Sagawa, Y. Multiple Sclerosis and Clinical Gait Analysis before and after Fampridine: A Systematic Review. Eur. Neurol. 2017, 78, 272–286. [Google Scholar] [CrossRef]
- Perez-Ros, P.; Martinez-Arnau, F.M.; Orti-Lucas, R.M.; Tarazona-Santabalbina, F.J. A predictive model of isolated and recurrent falls in functionally independent community-dwelling older adults. Braz. J. Phys. Ther. 2019, 23, 19–26. [Google Scholar] [CrossRef]
- Jiang, Y.; Xia, Q.; Wang, J.; Zhou, P.; Jiang, S.; Diwan, V.K.; Xu, B. Insomnia, Benzodiazepine Use, and Falls among Residents in Long-term Care Facilities. Int. J. Environ. Res. Public Health 2019, 16, 4623. [Google Scholar] [CrossRef]
- Lang, T.; Streeper, T.; Cawthon, P.; Baldwin, K.; Taaffe, D.R.; Harris, T.B. Sarcopenia: Etiology, clinical consequences, intervention, and assessment. Osteoporos. Int. 2010, 21, 543–559. [Google Scholar] [CrossRef] [PubMed]
- Dutta, C. Significance of sarcopenia in the elderly. J. Nutr. 1997, 127, 992S–993S. [Google Scholar] [CrossRef] [PubMed]
- Bula, C.J.; Monod, S.; Hoskovec, C.; Rochat, S. Interventions aiming at balance confidence improvement in older adults: An updated review. Gerontology 2011, 57, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Hewston, P.; Deshpande, N. Fear of Falling and Balance Confidence in Older Adults With Type 2 Diabetes Mellitus: A Scoping Review. Can. J. Diabetes 2018, 42, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Gregg, E.W.; Engelgau, M.M.; Narayan, V. Complications of diabetes in elderly people. BMJ 2002, 325, 916–917. [Google Scholar] [CrossRef]
- Gregg, E.W.; Mangione, C.M.; Cauley, J.A.; Thompson, T.J.; Schwartz, A.V.; Ensrud, K.E.; Nevitt, M.C.; Study of Osteoporotic Fractures Research Group. Diabetes and incidence of functional disability in older women. Diabetes Care 2002, 25, 61–67. [Google Scholar] [CrossRef]
- Schwartz, A.V.; Hillier, T.A.; Sellmeyer, D.E.; Resnick, H.E.; Gregg, E.; Ensrud, K.E.; Schreiner, P.J.; Margolis, K.L.; Cauley, J.A.; Nevitt, M.C.; et al. Older women with diabetes have a higher risk of falls: A prospective study. Diabetes Care 2002, 25, 1749–1754. [Google Scholar] [CrossRef]
- Neri, S.G.R.; Gadelha, A.B.; de David, A.C.; Ferreira, A.P.; Safons, M.P.; Tiedemann, A.; Lima, R.M. The Association Between Body Adiposity Measures, Postural Balance, Fear of Falling, and Fall Risk in Older Community-Dwelling Women. J. Geriatr. Phys. Ther. 2019, 42, E94–E100. [Google Scholar] [CrossRef]
- Kaye, J.A.; Oken, B.S.; Howieson, D.B.; Howieson, J.; Holm, L.A.; Dennison, K. Neurologic evaluation of the optimally healthy oldest old. Arch. Neurol. 1994, 51, 1205–1211. [Google Scholar] [CrossRef]
- Peachey, M.M.; Richardson, J.; Tang, A.V.; Dal-Bello Haas, V.; Gravesande, J. Environmental, behavioural and multicomponent interventions to reduce adults’ sitting time: A systematic review and meta-analysis. Br. J. Sports Med. 2020, 54, 315–325. [Google Scholar] [CrossRef]
- Lord, S.R.; Delbaere, K.; Gandevia, S.C. Use of a physiological profile to document motor impairment in ageing and in clinical groups. J. Physiol. 2016, 594, 4513–4523. [Google Scholar] [CrossRef] [PubMed]
- Anson, E.; Pineault, K.; Bair, W.; Studenski, S.; Agrawal, Y. Reduced vestibular function is associated with longer, slower steps in healthy adults during normal speed walking. Gait Posture 2019, 68, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Chow, M.R.; Ayiotis, A.I.; Schoo, D.P.; Gimmon, Y.; Lane, K.E.; Morris, B.J.; Rahman, M.A.; Valentin, N.S.; Boutros, P.J.; Bowditch, S.P.; et al. Posture, Gait, Quality of Life, and Hearing with a Vestibular Implant. N. Engl. J. Med. 2021, 384, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Matheson, A.J.; Darlington, C.L.; Smith, P.F. Dizziness in the elderly and age-related degeneration of the vestibular system. N. Z. J. Psychol. 1999, 28, 10–16. [Google Scholar]
- Masud, T.; Morris, R.O. Epidemiology of falls. Age Ageing 2001, 30 (Suppl. 4), 3–7. [Google Scholar] [CrossRef]
- Blake, A.J.; Morgan, K.; Bendall, M.J.; Dallosso, H.; Ebrahim, S.B.; Arie, T.H.; Fentem, P.H.; Bassey, E.J. Falls by elderly people at home: Prevalence and associated factors. Age Ageing 1988, 17, 365–372. [Google Scholar] [CrossRef]
- Prudham, D.; Evans, J.G. Factors associated with falls in the elderly: A community study. Age Ageing 1981, 10, 141–146. [Google Scholar] [CrossRef]
- Campbell, A.J.; Reinken, J.; Allan, B.C.; Martinez, G.S. Falls in old age: A study of frequency and related clinical factors. Age Ageing 1981, 10, 264–270. [Google Scholar] [CrossRef]
- Varadaraj, V.; Mihailovic, A.; Ehrenkranz, R.; Lesche, S.; Ramulu, P.Y.; Swenor, B.K. Gait Characteristics of Age-Related Macular Degeneration Patients. Transl. Vis. Sci. Technol. 2017, 6, 14. [Google Scholar] [CrossRef]
- Duggan, E.; Donoghue, O.; Kenny, R.A.; Cronin, H.; Loughman, J.; Finucane, C. Time to Refocus Assessment of Vision in Older Adults? Contrast Sensitivity but Not Visual Acuity Is Associated With Gait in Older Adults. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 1663–1668. [Google Scholar] [CrossRef]
- Bicket, A.K.; Mihailovic, A.; E, J.Y.; Nguyen, A.; Mukherjee, M.R.; Friedman, D.S.; Ramulu, P.Y. Gait in Elderly Glaucoma: Impact of Lighting Conditions, Changes in Lighting, and Fear of Falling. Transl. Vis. Sci. Technol. 2020, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Morrison, J.H.; Baxter, M.G. The ageing cortical synapse: Hallmarks and implications for cognitive decline. Nat. Rev. Neurosci. 2012, 13, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Petreanu, L.; Mao, T.; Sternson, S.M.; Svoboda, K. The subcellular organization of neocortical excitatory connections. Nature 2009, 457, 1142–1145. [Google Scholar] [CrossRef] [PubMed]
- Wimmer, V.C.; Broser, P.J.; Kuner, T.; Bruno, R.M. Experience-induced plasticity of thalamocortical axons in both juveniles and adults. J. Comp. Neurol. 2010, 518, 4629–4648. [Google Scholar] [CrossRef]
- Lacefield, C.O.; Pnevmatikakis, E.A.; Paninski, L.; Bruno, R.M. Reinforcement Learning Recruits Somata and Apical Dendrites across Layers of Primary Sensory Cortex. Cell Rep. 2019, 26, 2000–2008.e2002. [Google Scholar] [CrossRef]
- Cichon, J.; Gan, W.B. Branch-specific dendritic Ca2+ spikes cause persistent synaptic plasticity. Nature 2015, 520, 180–185. [Google Scholar] [CrossRef]
- Clark, B.C.; Woods, A.J.; Clark, L.A.; Criss, C.R.; Shadmehr, R.; Grooms, D.R. The Aging Brain & the Dorsal Basal Ganglia: Implications for Age-Related Limitations of Mobility. Adv. Geriatr. Med. Res. 2019, 1, e190008. [Google Scholar] [CrossRef]
- Phillips, J.G.; Bradshaw, J.L.; Iansek, R.; Chiu, E. Motor functions of the basal ganglia. Psychol. Res. 1993, 55, 175–181. [Google Scholar] [CrossRef]
- Hausdorff, J.M.; Rios, D.A.; Edelberg, H.K. Gait variability and fall risk in community-living older adults: A 1-year prospective study. Arch. Phys. Med. Rehabil. 2001, 82, 1050–1056. [Google Scholar] [CrossRef]
- Peel, N.M. Epidemiology of falls in older age. Can. J. Aging 2011, 30, 7–19. [Google Scholar] [CrossRef]
- Rothman, M.D.; Leo-Summers, L.; Gill, T.M. Prognostic significance of potential frailty criteria. J. Am. Geriatr. Soc. 2008, 56, 2211–2216. [Google Scholar] [CrossRef] [PubMed]
- Albers, M.W.; Gilmore, G.C.; Kaye, J.; Murphy, C.; Wingfield, A.; Bennett, D.A.; Boxer, A.L.; Buchman, A.S.; Cruickshanks, K.J.; Devanand, D.P.; et al. At the interface of sensory and motor dysfunctions and Alzheimer’s disease. Alzheimers Dement. 2015, 11, 70–98. [Google Scholar] [CrossRef] [PubMed]
- Scarmeas, N.; Albert, M.; Brandt, J.; Blacker, D.; Hadjigeorgiou, G.; Papadimitriou, A.; Dubois, B.; Sarazin, M.; Wegesin, D.; Marder, K.; et al. Motor signs predict poor outcomes in Alzheimer disease. Neurology 2005, 64, 1696–1703. [Google Scholar] [CrossRef]
- Carlsson, A.; Winblad, B. Influence of age and time interval between death and autopsy on dopamine and 3-methoxytyramine levels in human basal ganglia. J. Neural. Transm. 1976, 38, 271–276. [Google Scholar] [CrossRef]
- Suhara, T.; Fukuda, H.; Inoue, O.; Itoh, T.; Suzuki, K.; Yamasaki, T.; Tateno, Y. Age-related changes in human D1 dopamine receptors measured by positron emission tomography. Psychopharmacology 1991, 103, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chan, G.L.; Holden, J.E.; Dobko, T.; Mak, E.; Schulzer, M.; Huser, J.M.; Snow, B.J.; Ruth, T.J.; Calne, D.B.; et al. Age-dependent decline of dopamine D1 receptors in human brain: A PET study. Synapse 1998, 30, 56–61. [Google Scholar] [CrossRef]
- Rinne, J.O.; Hietala, J.; Ruotsalainen, U.; Sako, E.; Laihinen, A.; Nagren, K.; Lehikoinen, P.; Oikonen, V.; Syvalahti, E. Decrease in human striatal dopamine D2 receptor density with age: A PET study with [11C]raclopride. J. Cereb. Blood Flow Metab. 1993, 13, 310–314. [Google Scholar] [CrossRef]
- Wong, D.F.; Young, D.; Wilson, P.D.; Meltzer, C.C.; Gjedde, A. Quantification of neuroreceptors in the living human brain: III. D2-like dopamine receptors: Theory, validation, and changes during normal aging. J. Cereb. Blood Flow Metab. 1997, 17, 316–330. [Google Scholar] [CrossRef]
- Dumurgier, J.; Crivello, F.; Mazoyer, B.; Ahmed, I.; Tavernier, B.; Grabli, D.; Francois, C.; Tzourio-Mazoyer, N.; Tzourio, C.; Elbaz, A. MRI atrophy of the caudate nucleus and slower walking speed in the elderly. Neuroimage 2012, 60, 871–878. [Google Scholar] [CrossRef]
- Del Campo, N.; Payoux, P.; Djilali, A.; Delrieu, J.; Hoogendijk, E.O.; Rolland, Y.; Cesari, M.; Weiner, M.W.; Andrieu, S.; Vellas, B.; et al. Relationship of regional brain beta-amyloid to gait speed. Neurology 2016, 86, 36–43. [Google Scholar] [CrossRef]
- Clark, B.C.; Taylor, J.L. Age-related changes in motor cortical properties and voluntary activation of skeletal muscle. Curr. Aging Sci. 2011, 4, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Beauchet, O.; Launay, C.P.; Barden, J.; Liu-Ambrose, T.; Chester, V.L.; Szturm, T.; Grenier, S.; Leonard, G.; Bherer, L.; Annweiler, C.; et al. Association Between Falls and Brain Subvolumes: Results from a Cross-Sectional Analysis in Healthy Older Adults. Brain Topogr. 2017, 30, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Ding, Y.S.; Fowler, J.S.; Wang, G.J.; Logan, J.; Gatley, S.J.; Hitzemann, R.; Smith, G.; Fields, S.D.; Gur, R. Dopamine transporters decrease with age. J. Nucl. Med. 1996, 37, 554–559. [Google Scholar] [PubMed]
- Mesco, E.R.; Joseph, J.A.; Blake, M.J.; Roth, G.S. Loss of D2 receptors during aging is partially due to decreased levels of mRNA. Brain Res. 1991, 545, 355–357. [Google Scholar] [CrossRef]
- Rosano, C.; Aizenstein, H.; Brach, J.; Longenberger, A.; Studenski, S.; Newman, A.B. Special article: Gait measures indicate underlying focal gray matter atrophy in the brain of older adults. J. Gerontol. A Biol. Sci. Med. Sci. 2008, 63, 1380–1388. [Google Scholar] [CrossRef] [PubMed]
- McGinley, M.; Hoffman, R.L.; Russ, D.W.; Thomas, J.S.; Clark, B.C. Older adults exhibit more intracortical inhibition and less intracortical facilitation than young adults. Exp. Gerontol. 2010, 45, 671–678. [Google Scholar] [CrossRef]
- Kossev, A.R.; Schrader, C.; Dauper, J.; Dengler, R.; Rollnik, J.D. Increased intracortical inhibition in middle-aged humans; a study using paired-pulse transcranial magnetic stimulation. Neurosci. Lett. 2002, 333, 83–86. [Google Scholar] [CrossRef]
- Rowe, J.B.; Siebner, H.; Filipovic, S.R.; Cordivari, C.; Gerschlager, W.; Rothwell, J.; Frackowiak, R. Aging is associated with contrasting changes in local and distant cortical connectivity in the human motor system. Neuroimage 2006, 32, 747–760. [Google Scholar] [CrossRef]
- Naccarato, M.; Calautti, C.; Jones, P.S.; Day, D.J.; Carpenter, T.A.; Baron, J.C. Does healthy aging affect the hemispheric activation balance during paced index-to-thumb opposition task? An fMRI study. Neuroimage 2006, 32, 1250–1256. [Google Scholar] [CrossRef]
- Hickmott, P.; Dinse, H. Effects of aging on properties of the local circuit in rat primary somatosensory cortex (S1) in vitro. Cereb. Cortex 2013, 23, 2500–2513. [Google Scholar] [CrossRef]
- Spengler, F.; Godde, B.; Dinse, H.R. Effects of ageing on topographic organization of somatosensory cortex. Neuroreport 1995, 6, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Popescu, I.R.; Le, K.Q.; Ducote, A.L.; Li, J.E.; Leland, A.E.; Mostany, R. Increased intrinsic excitability and decreased synaptic inhibition in aged somatosensory cortex pyramidal neurons. Neurobiol. Aging 2021, 98, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Lenz, M.; Tegenthoff, M.; Kohlhaas, K.; Stude, P.; Hoffken, O.; Gatica Tossi, M.A.; Kalisch, T.; Kowalewski, R.; Dinse, H.R. Increased excitability of somatosensory cortex in aged humans is associated with impaired tactile acuity. J. Neurosci. 2012, 32, 1811–1816. [Google Scholar] [CrossRef]
- Ruitenberg, M.F.L.; Cassady, K.E.; Reuter-Lorenz, P.A.; Tommerdahl, M.; Seidler, R.D. Age-Related Reductions in Tactile and Motor Inhibitory Function Start Early but Are Independent. Front. Aging Neurosci. 2019, 11, 193. [Google Scholar] [CrossRef] [PubMed]
- Hennige, A.M.; Sartorius, T.; Lutz, S.Z.; Tschritter, O.; Preissl, H.; Hopp, S.; Fritsche, A.; Rammensee, H.G.; Ruth, P.; Haring, H.U. Insulin-mediated cortical activity in the slow frequency range is diminished in obese mice and promotes physical inactivity. Diabetologia 2009, 52, 2416–2424. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sartorius, T.; Heni, M.; Tschritter, O.; Preissl, H.; Hopp, S.; Fritsche, A.; Lievertz, P.S.; Gertler, A.; Berthou, F.; Taouis, M.; et al. Leptin affects insulin action in astrocytes and impairs insulin-mediated physical activity. Cell. Physiol. Biochem. 2012, 30, 238–246. [Google Scholar] [CrossRef]
- Sartorius, T.; Hennige, A.M.; Fritsche, A.; Haring, H.U. Sustained Treatment with Insulin Detemir in Mice Alters Brain Activity and Locomotion. PLoS ONE 2016, 11, e0162124. [Google Scholar] [CrossRef]
- Galindo-Mendez, B.; Trevino, J.A.; McGlinchey, R.; Fortier, C.; Lioutas, V.; Novak, P.; Mantzoros, C.S.; Ngo, L.; Novak, V. Memory advancement by intranasal insulin in type 2 diabetes (MemAID) randomized controlled clinical trial: Design, methods and rationale. Contemp. Clin. Trials 2020, 89, 105934. [Google Scholar] [CrossRef]
- Cholerton, B.; Baker, L.D.; Craft, S. Insulin resistance and pathological brain ageing. Diabet. Med. 2011, 28, 1463–1475. [Google Scholar] [CrossRef]
- Frazier, H.N.; Ghoweri, A.O.; Anderson, K.L.; Lin, R.L.; Porter, N.M.; Thibault, O. Broadening the definition of brain insulin resistance in aging and Alzheimer’s disease. Exp. Neurol. 2019, 313, 79–87. [Google Scholar] [CrossRef]
- Tchilian, E.Z.; Zhelezarov, I.E.; Petkov, V.V.; Hadjiivanova, C.I. 125I-insulin binding is decreased in olfactory bulbs of aged rats. Neuropeptides 1990, 17, 193–196. [Google Scholar] [CrossRef]
- Zaia, A.; Piantanelli, L. Insulin receptors in the brain cortex of aging mice. Mech. Ageing Dev. 2000, 113, 227–232. [Google Scholar] [CrossRef]
- Dore, S.; Kar, S.; Rowe, W.; Quirion, R. Distribution and levels of [125I]IGF-I, [125I]IGF-II and [125I]insulin receptor binding sites in the hippocampus of aged memory-unimpaired and -impaired rats. Neuroscience 1997, 80, 1033–1040. [Google Scholar] [CrossRef]
- Hill, J.M.; Lesniak, M.A.; Pert, C.B.; Roth, J. Autoradiographic localization of insulin receptors in rat brain: Prominence in olfactory and limbic areas. Neuroscience 1986, 17, 1127–1138. [Google Scholar] [CrossRef]
- Patel, J.C.; Stouffer, M.A.; Mancini, M.; Nicholson, C.; Carr, K.D.; Rice, M.E. Interactions between insulin and diet on striatal dopamine uptake kinetics in rodent brain slices. Eur. J. Neurosci. 2019, 49, 794–804. [Google Scholar] [CrossRef]
- Cai, W.; Xue, C.; Sakaguchi, M.; Konishi, M.; Shirazian, A.; Ferris, H.A.; Li, M.E.; Yu, R.; Kleinridders, A.; Pothos, E.N.; et al. Insulin regulates astrocyte gliotransmission and modulates behavior. J. Clin. Investig. 2018, 128, 2914–2926. [Google Scholar] [CrossRef]
- Zhao, W.; Chen, H.; Xu, H.; Moore, E.; Meiri, N.; Quon, M.J.; Alkon, D.L. Brain insulin receptors and spatial memory. Correlated changes in gene expression, tyrosine phosphorylation, and signaling molecules in the hippocampus of water maze trained rats. J. Biol. Chem. 1999, 274, 34893–34902. [Google Scholar] [CrossRef]
- Barone, E.; Tramutola, A.; Triani, F.; Calcagnini, S.; Di Domenico, F.; Ripoli, C.; Gaetani, S.; Grassi, C.; Butterfield, D.A.; Cassano, T.; et al. Biliverdin Reductase-A Mediates the Beneficial Effects of Intranasal Insulin in Alzheimer Disease. Mol. Neurobiol. 2019, 56, 2922–2943. [Google Scholar] [CrossRef]
- Anderson, K.L.; Frazier, H.N.; Maimaiti, S.; Bakshi, V.V.; Majeed, Z.R.; Brewer, L.D.; Porter, N.M.; Lin, A.L.; Thibault, O. Impact of Single or Repeated Dose Intranasal Zinc-free Insulin in Young and Aged F344 Rats on Cognition, Signaling, and Brain Metabolism. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 189–197. [Google Scholar] [CrossRef]
- Lo, F.S.; Erzurumlu, R.S. Insulin receptor sensitization restores neocortical excitation/inhibition balance in a mouse model of autism. Mol. Autism. 2018, 9, 13. [Google Scholar] [CrossRef]
- Fine, J.M.; Stroebel, B.M.; Faltesek, K.A.; Terai, K.; Haase, L.; Knutzen, K.E.; Kosyakovsky, J.; Bowe, T.J.; Fuller, A.K.; Frey, W.H.; et al. Intranasal delivery of low-dose insulin ameliorates motor dysfunction and dopaminergic cell death in a 6-OHDA rat model of Parkinson’s Disease. Neurosci. Lett. 2020, 714, 134567. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.; Hallschmid, M.; Schmitz, K.; Schultes, B.; Ratter, F.; Fehm, H.L.; Born, J.; Kern, W. Intranasal insulin improves memory in humans: Superiority of insulin aspart. Neuropsychopharmacology 2007, 32, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Born, J.; Lange, T.; Kern, W.; McGregor, G.P.; Bickel, U.; Fehm, H.L. Sniffing neuropeptides: A transnasal approach to the human brain. Nat. Neurosci. 2002, 5, 514–516. [Google Scholar] [CrossRef] [PubMed]
- Brabazon, F.; Wilson, C.M.; Jaiswal, S.; Reed, J.; Frey, W.H.N.; Byrnes, K.R. Intranasal insulin treatment of an experimental model of moderate traumatic brain injury. J. Cereb. Blood Flow Metab. 2017, 37, 3203–3218. [Google Scholar] [CrossRef] [PubMed]
- Lochhead, J.J.; Thorne, R.G. Intranasal delivery of biologics to the central nervous system. Adv. Drug Deliv. Rev. 2012, 64, 614–628. [Google Scholar] [CrossRef]
- Thorne, R.G.; Emory, C.R.; Ala, T.A.; Frey, W.H., 2nd. Quantitative analysis of the olfactory pathway for drug delivery to the brain. Brain Res. 1995, 692, 278–282. [Google Scholar] [CrossRef]
- de la Monte, S.M. Intranasal insulin therapy for cognitive impairment and neurodegeneration: Current state of the art. Expert Opin. Drug Deliv. 2013, 10, 1699–1709. [Google Scholar] [CrossRef]
- Hanson, L.R.; Frey, W.H., 2nd. Intranasal delivery bypasses the blood-brain barrier to target therapeutic agents to the central nervous system and treat neurodegenerative disease. BMC Neurosci. 2008, 9 (Suppl. 3), S5. [Google Scholar] [CrossRef]
- Maimaiti, S.; Anderson, K.L.; DeMoll, C.; Brewer, L.D.; Rauh, B.A.; Gant, J.C.; Blalock, E.M.; Porter, N.M.; Thibault, O. Intranasal Insulin Improves Age-Related Cognitive Deficits and Reverses Electrophysiological Correlates of Brain Aging. J. Gerontol. A Biol. Sci. Med. Sci. 2016, 71, 30–39. [Google Scholar] [CrossRef]
- Beirami, E.; Oryan, S.; Seyedhosseini Tamijani, S.M.; Ahmadiani, A.; Dargahi, L. Intranasal insulin treatment alleviates methamphetamine induced anxiety-like behavior and neuroinflammation. Neurosci. Lett. 2017, 660, 122–129. [Google Scholar] [CrossRef]
- Adzovic, L.; Lynn, A.E.; D’Angelo, H.M.; Crockett, A.M.; Kaercher, R.M.; Royer, S.E.; Hopp, S.C.; Wenk, G.L. Insulin improves memory and reduces chronic neuroinflammation in the hippocampus of young but not aged brains. J. NeuroInflamm. 2015, 12, 63. [Google Scholar] [CrossRef] [PubMed]
- Mamik, M.K.; Asahchop, E.L.; Chan, W.F.; Zhu, Y.; Branton, W.G.; McKenzie, B.A.; Cohen, E.A.; Power, C. Insulin Treatment Prevents Neuroinflammation and Neuronal Injury with Restored Neurobehavioral Function in Models of HIV/AIDS Neurodegeneration. J. Neurosci. 2016, 36, 10683–10695. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Guo, Z.; Mao, Y.F.; Zheng, T.; Zhang, B. Intranasal Insulin Ameliorates Cerebral Hypometabolism, Neuronal Loss, and Astrogliosis in Streptozotocin-Induced Alzheimer’s Rat Model. Neurotox. Res. 2018, 33, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Rajasekar, N.; Nath, C.; Hanif, K.; Shukla, R. Intranasal insulin improves cerebral blood flow, Nrf-2 expression and BDNF in STZ (ICV)-induced memory impaired rats. Life Sci. 2017, 173, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.F.; Guo, Z.; Zheng, T.; Jiang, Y.; Yan, Y.; Yin, X.; Chen, Y.; Zhang, B. Intranasal insulin alleviates cognitive deficits and amyloid pathology in young adult APPswe/PS1dE9 mice. Aging Cell 2016, 15, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dai, C.L.; Chen, Y.; Iqbal, K.; Liu, F.; Gong, C.X. Intranasal Insulin Prevents Anesthesia-Induced Spatial Learning and Memory Deficit in Mice. Sci. Rep. 2016, 6, 21186. [Google Scholar] [CrossRef]
- Apostolatos, A.; Song, S.; Acosta, S.; Peart, M.; Watson, J.E.; Bickford, P.; Cooper, D.R.; Patel, N.A. Insulin promotes neuronal survival via the alternatively spliced protein kinase CdeltaII isoform. J. Biol. Chem. 2012, 287, 9299–9310. [Google Scholar] [CrossRef]
- Salameh, T.S.; Bullock, K.M.; Hujoel, I.A.; Niehoff, M.L.; Wolden-Hanson, T.; Kim, J.; Morley, J.E.; Farr, S.A.; Banks, W.A. Central Nervous System Delivery of Intranasal Insulin: Mechanisms of Uptake and Effects on Cognition. J. Alzheimers Dis. 2015, 47, 715–728. [Google Scholar] [CrossRef]
- Claxton, A.; Baker, L.D.; Hanson, A.; Trittschuh, E.H.; Cholerton, B.; Morgan, A.; Callaghan, M.; Arbuckle, M.; Behl, C.; Craft, S. Long-acting intranasal insulin detemir improves cognition for adults with mild cognitive impairment or early-stage Alzheimer’s disease dementia. J. Alzheimers Dis. 2015, 44, 897–906. [Google Scholar] [CrossRef]
- Craft, S.; Baker, L.D.; Montine, T.J.; Minoshima, S.; Watson, G.S.; Claxton, A.; Arbuckle, M.; Callaghan, M.; Tsai, E.; Plymate, S.R.; et al. Intranasal insulin therapy for Alzheimer disease and amnestic mild cognitive impairment: A pilot clinical trial. Arch. Neurol. 2012, 69, 29–38. [Google Scholar] [CrossRef]
- Hallschmid, M. Intranasal Insulin for Alzheimer’s Disease. CNS Drugs 2021, 35, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Reger, M.A.; Watson, G.S.; Frey, W.H., 2nd; Baker, L.D.; Cholerton, B.; Keeling, M.L.; Belongia, D.A.; Fishel, M.A.; Plymate, S.R.; Schellenberg, G.D.; et al. Effects of intranasal insulin on cognition in memory-impaired older adults: Modulation by APOE genotype. Neurobiol. Aging 2006, 27, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Reger, M.A.; Watson, G.S.; Green, P.S.; Wilkinson, C.W.; Baker, L.D.; Cholerton, B.; Fishel, M.A.; Plymate, S.R.; Breitner, J.C.; DeGroodt, W.; et al. Intranasal insulin improves cognition and modulates beta-amyloid in early AD. Neurology 2008, 70, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.; Hallschmid, M.; Hatke, A.; Schultes, B.; Fehm, H.L.; Born, J.; Kern, W. Intranasal insulin improves memory in humans. Psychoneuroendocrinology 2004, 29, 1326–1334. [Google Scholar] [CrossRef]
- Benedict, C.; Kern, W.; Schultes, B.; Born, J.; Hallschmid, M. Differential sensitivity of men and women to anorexigenic and memory-improving effects of intranasal insulin. J. Clin. Endocrinol. Metab. 2008, 93, 1339–1344. [Google Scholar] [CrossRef]
- Brunner, Y.F.; Kofoet, A.; Benedict, C.; Freiherr, J. Central insulin administration improves odor-cued reactivation of spatial memory in young men. J. Clin. Endocrinol. Metab. 2015, 100, 212–219. [Google Scholar] [CrossRef]
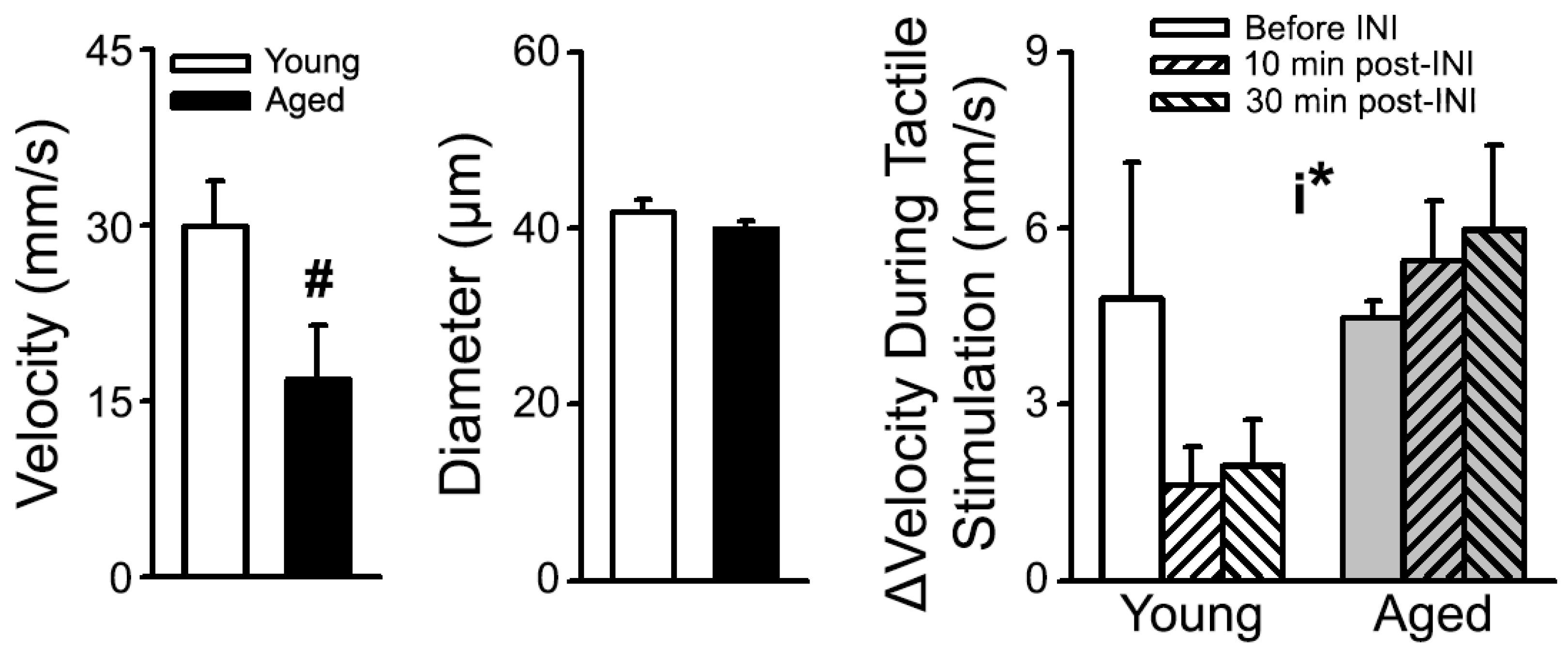
- Lin, R.L.; Frazier, H.N.; Anderson, K.L.; Case, S.L.; Ghoweri, A.O.; Thibault, O. Sensitivity of the S1 neuronal calcium network to insulin and Bay-K 8644 in vivo: Relationship to gait, motivation, and aging processes. Aging cell. 2022, 21, e13661. [Google Scholar] [CrossRef] [PubMed]
- Grichisch, Y.; Cavusoglu, M.; Preissl, H.; Uludag, K.; Hallschmid, M.; Birbaumer, N.; Haring, H.U.; Fritsche, A.; Veit, R. Differential effects of intranasal insulin and caffeine on cerebral blood flow. Hum. Brain Mapp. 2012, 33, 280–287. [Google Scholar] [CrossRef]
- Schilling, T.M.; Ferreira de Sa, D.S.; Westerhausen, R.; Strelzyk, F.; Larra, M.F.; Hallschmid, M.; Savaskan, E.; Oitzl, M.S.; Busch, H.P.; Naumann, E.; et al. Intranasal insulin increases regional cerebral blood flow in the insular cortex in men independently of cortisol manipulation. Hum. Brain Mapp. 2014, 35, 1944–1956. [Google Scholar] [CrossRef]
- Kullmann, S.; Heni, M.; Veit, R.; Scheffler, K.; Machann, J.; Haring, H.U.; Fritsche, A.; Preissl, H. Selective insulin resistance in homeostatic and cognitive control brain areas in overweight and obese adults. Diabetes Care 2015, 38, 1044–1050. [Google Scholar] [CrossRef]
- Wingrove, J.O.; O’Daly, O.; Forbes, B.; Swedrowska, M.; Amiel, S.A.; Zelaya, F.O. Intranasal insulin administration decreases cerebral blood flow in cortico-limbic regions: A neuropharmacological imaging study in normal and overweight males. Diabetes Obes. Metab. 2021, 23, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Novak, V.; Milberg, W.; Hao, Y.; Munshi, M.; Novak, P.; Galica, A.; Manor, B.; Roberson, P.; Craft, S.; Abduljalil, A. Enhancement of vasoreactivity and cognition by intranasal insulin in type 2 diabetes. Diabetes Care 2014, 37, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Akintola, A.A.; van Opstal, A.M.; Westendorp, R.G.; Postmus, I.; van der Grond, J.; van Heemst, D. Effect of intranasally administered insulin on cerebral blood flow and perfusion; a randomized experiment in young and older adults. Aging 2017, 9, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, S. Is sporadic Alzheimer disease the brain type of non-insulin dependent diabetes mellitus? A challenging hypothesis. J. Neural. Transm. 1998, 105, 415–422. [Google Scholar] [CrossRef]
- Frolich, L.; Blum-Degen, D.; Bernstein, H.G.; Engelsberger, S.; Humrich, J.; Laufer, S.; Muschner, D.; Thalheimer, A.; Turk, A.; Hoyer, S.; et al. Brain insulin and insulin receptors in aging and sporadic Alzheimer’s disease. J. Neural. Transm. 1998, 105, 423–438. [Google Scholar] [CrossRef]
- Calvo-Ochoa, E.; Arias, C. Cellular and metabolic alterations in the hippocampus caused by insulin signalling dysfunction and its association with cognitive impairment during aging and Alzheimer’s disease: Studies in animal models. Diabetes Metab. Res. Rev. 2015, 31, 1–13. [Google Scholar] [CrossRef]
- Cholerton, B.; Baker, L.D.; Craft, S. Insulin, cognition, and dementia. Eur. J. Pharmacol. 2013, 719, 170–179. [Google Scholar] [CrossRef]
- Couet, C.; Delarue, J.; Constans, T.; Lamisse, F. Age-related insulin resistance: A review. Horm. Res. 1992, 38, 46–50. [Google Scholar] [CrossRef]
- Gabbouj, S.; Ryhanen, S.; Marttinen, M.; Wittrahm, R.; Takalo, M.; Kemppainen, S.; Martiskainen, H.; Tanila, H.; Haapasalo, A.; Hiltunen, M.; et al. Altered Insulin Signaling in Alzheimer’s Disease Brain-Special Emphasis on PI3K-Akt Pathway. Front. Neurosci. 2019, 13, 629. [Google Scholar] [CrossRef]
- Messier, C.; Teutenberg, K. The role of insulin, insulin growth factor, and insulin-degrading enzyme in brain aging and Alzheimer’s disease. Neural. Plast. 2005, 12, 311–328. [Google Scholar] [CrossRef]
- Chow, H.M.; Shi, M.; Cheng, A.; Gao, Y.; Chen, G.; Song, X.; So, R.W.L.; Zhang, J.; Herrup, K. Age-related hyperinsulinemia leads to insulin resistance in neurons and cell-cycle-induced senescence. Nat. Neurosci. 2019, 22, 1806–1819. [Google Scholar] [CrossRef] [PubMed]
- Talbot, K.; Wang, H.Y.; Kazi, H.; Han, L.Y.; Bakshi, K.P.; Stucky, A.; Fuino, R.L.; Kawaguchi, K.R.; Samoyedny, A.J.; Wilson, R.S.; et al. Demonstrated brain insulin resistance in Alzheimer’s disease patients is associated with IGF-1 resistance, IRS-1 dysregulation, and cognitive decline. J. Clin. Investig. 2012, 122, 1316–1338. [Google Scholar] [CrossRef] [PubMed]
- Denver, P.; English, A.; McClean, P.L. Inflammation, insulin signaling and cognitive function in aged APP/PS1 mice. Brain Behav. Immun. 2018, 70, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Velazquez, R.; Tran, A.; Ishimwe, E.; Denner, L.; Dave, N.; Oddo, S.; Dineley, K.T. Central insulin dysregulation and energy dyshomeostasis in two mouse models of Alzheimer’s disease. Neurobiol. Aging 2017, 58, 1–13. [Google Scholar] [CrossRef]
- Stanley, M.; Macauley, S.L.; Caesar, E.E.; Koscal, L.J.; Moritz, W.; Robinson, G.O.; Roh, J.; Keyser, J.; Jiang, H.; Holtzman, D.M. The Effects of Peripheral and Central High Insulin on Brain Insulin Signaling and Amyloid-beta in Young and Old APP/PS1 Mice. J. Neurosci. 2016, 36, 11704–11715. [Google Scholar] [CrossRef]
- Bomfim, T.R.; Forny-Germano, L.; Sathler, L.B.; Brito-Moreira, J.; Houzel, J.C.; Decker, H.; Silverman, M.A.; Kazi, H.; Melo, H.M.; McClean, P.L.; et al. An anti-diabetes agent protects the mouse brain from defective insulin signaling caused by Alzheimer’s disease- associated Abeta oligomers. J. Clin. Investig. 2012, 122, 1339–1353. [Google Scholar] [CrossRef]
- Craft, S.; Asthana, S.; Newcomer, J.W.; Wilkinson, C.W.; Matos, I.T.; Baker, L.D.; Cherrier, M.; Lofgreen, C.; Latendresse, S.; Petrova, A.; et al. Enhancement of memory in Alzheimer disease with insulin and somatostatin, but not glucose. Arch. Gen. Psychiatry 1999, 56, 1135–1140. [Google Scholar] [CrossRef]
- Reger, M.A.; Watson, G.S.; Green, P.S.; Baker, L.D.; Cholerton, B.; Fishel, M.A.; Plymate, S.R.; Cherrier, M.M.; Schellenberg, G.D.; Frey, W.H., 2nd; et al. Intranasal insulin administration dose-dependently modulates verbal memory and plasma amyloid-beta in memory-impaired older adults. J. Alzheimers Dis. 2008, 13, 323–331. [Google Scholar] [CrossRef]
- Pancani, T.; Anderson, K.L.; Brewer, L.D.; Kadish, I.; DeMoll, C.; Landfield, P.W.; Blalock, E.M.; Porter, N.M.; Thibault, O. Effect of high-fat diet on metabolic indices, cognition, and neuronal physiology in aging F344 rats. Neurobiol. Aging 2013, 34, 1977–1987. [Google Scholar] [CrossRef]
- Sartorius, T.; Peter, A.; Heni, M.; Maetzler, W.; Fritsche, A.; Haring, H.U.; Hennige, A.M. The brain response to peripheral insulin declines with age: A contribution of the blood-brain barrier? PLoS ONE 2015, 10, e0126804. [Google Scholar] [CrossRef]
- Banks, W.A.; Jaspan, J.B.; Huang, W.; Kastin, A.J. Transport of insulin across the blood-brain barrier: Saturability at euglycemic doses of insulin. Peptides 1997, 18, 1423–1429. [Google Scholar] [CrossRef]
- Banks, W.A.; Owen, J.B.; Erickson, M.A. Insulin in the brain: There and back again. Pharmacol. Ther. 2012, 136, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Baskin, D.G.; Porte, D., Jr.; Guest, K.; Dorsa, D.M. Regional concentrations of insulin in the rat brain. Endocrinology 1983, 112, 898–903. [Google Scholar] [CrossRef] [PubMed]
- Baskin, D.G.; Brewitt, B.; Davidson, D.A.; Corp, E.; Paquette, T.; Figlewicz, D.P.; Lewellen, T.K.; Graham, M.K.; Woods, S.G.; Dorsa, D.M. Quantitative autoradiographic evidence for insulin receptors in the choroid plexus of the rat brain. Diabetes 1986, 35, 246–249. [Google Scholar] [CrossRef][Green Version]
- Schwartz, M.W.; Figlewicz, D.F.; Kahn, S.E.; Baskin, D.G.; Greenwood, M.R.; Porte, D., Jr. Insulin binding to brain capillaries is reduced in genetically obese, hyperinsulinemic Zucker rats. Peptides 1990, 11, 467–472. [Google Scholar] [CrossRef]
- Sasaki, N.; Fukatsu, R.; Tsuzuki, K.; Hayashi, Y.; Yoshida, T.; Fujii, N.; Koike, T.; Wakayama, I.; Yanagihara, R.; Garruto, R.; et al. Advanced glycation end products in Alzheimer’s disease and other neurodegenerative diseases. Am. J. Pathol. 1998, 153, 1149–1155. [Google Scholar] [CrossRef]
- Heni, M.; Schopfer, P.; Peter, A.; Sartorius, T.; Fritsche, A.; Synofzik, M.; Haring, H.U.; Maetzler, W.; Hennige, A.M. Evidence for altered transport of insulin across the blood-brain barrier in insulin-resistant humans. Acta Diabetol. 2014, 51, 679–681. [Google Scholar] [CrossRef]
- Thienel, M.; Wilhelm, I.; Benedict, C.; Born, J.; Hallschmid, M. Intranasal insulin decreases circulating cortisol concentrations during early sleep in elderly humans. Neurobiol. Aging 2017, 54, 170–174. [Google Scholar] [CrossRef]
- Grillo, C.A.; Piroli, G.G.; Kaigler, K.F.; Wilson, S.P.; Wilson, M.A.; Reagan, L.P. Downregulation of hypothalamic insulin receptor expression elicits depressive-like behaviors in rats. Behav. Brain Res. 2011, 222, 230–235. [Google Scholar] [CrossRef]
- Hallschmid, M.; Benedict, C.; Schultes, B.; Born, J.; Kern, W. Obese men respond to cognitive but not to catabolic brain insulin signaling. Int. J. Obes. 2008, 32, 275–282. [Google Scholar] [CrossRef]
- Shih, A.Y.; Driscoll, J.D.; Drew, P.J.; Nishimura, N.; Schaffer, C.B.; Kleinfeld, D. Two-photon microscopy as a tool to study blood flow and neurovascular coupling in the rodent brain. J. Cereb. Blood Flow Metab. 2012, 32, 1277–1309. [Google Scholar] [CrossRef] [PubMed]
- McDowell, K.P.; Berthiaume, A.A.; Tieu, T.; Hartmann, D.A.; Shih, A.Y. VasoMetrics: Unbiased spatiotemporal analysis of microvascular diameter in multi-photon imaging applications. Quant. Imaging Med. Surg. 2021, 11, 969–982. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.-L.; Thibault, O. Measures of S1 CBF at Rest and in Response to Tactile Stimulation Following INI Delivery; University of Kentucky College of Medicine: Lexington, KY, USA, 2022; Unpublished work. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Case, S.L.; Frazier, H.N.; Anderson, K.L.; Lin, R.-L.; Thibault, O. Falling Short: The Contribution of Central Insulin Receptors to Gait Dysregulation in Brain Aging. Biomedicines 2022, 10, 1923. https://doi.org/10.3390/biomedicines10081923
Case SL, Frazier HN, Anderson KL, Lin R-L, Thibault O. Falling Short: The Contribution of Central Insulin Receptors to Gait Dysregulation in Brain Aging. Biomedicines. 2022; 10(8):1923. https://doi.org/10.3390/biomedicines10081923
Chicago/Turabian StyleCase, Sami L., Hilaree N. Frazier, Katie L. Anderson, Ruei-Lung Lin, and Olivier Thibault. 2022. "Falling Short: The Contribution of Central Insulin Receptors to Gait Dysregulation in Brain Aging" Biomedicines 10, no. 8: 1923. https://doi.org/10.3390/biomedicines10081923
APA StyleCase, S. L., Frazier, H. N., Anderson, K. L., Lin, R.-L., & Thibault, O. (2022). Falling Short: The Contribution of Central Insulin Receptors to Gait Dysregulation in Brain Aging. Biomedicines, 10(8), 1923. https://doi.org/10.3390/biomedicines10081923