
Alveolus Lung-on-a-Chip Platform: A Proposal


1
BRECA Health Care SL, 18016 Granada, Spain
2
Research Group on Methodology, Methods, Models and Outcomes of Health and Social Sciences (M3O), University of Vic-Central University of Catalonia, 08500 Vic, Spain
3
Department of Physical Therapy, EUSES University School, University of Barcelona-University of Girona (UB-UdG), 08907 Barcelona, Spain
4
Department of Surgery, School of Veterinary Medicine and Animal Sciences, University of São Paulo, 05508-270 São Paulo, Brazil
5
Facultad de Ciencias Experimentales, Universidad Francisco de Vitoria, 28223 Madrid, Spain
6
Human Movement and Rehabilitation, Post Graduation Program Medical School, Evangelic University of Anápolis-UniEVANGELICA, 75083-515 Anápolis, Brazil
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Chemosensors 2021, 9(9), 248; https://doi.org/10.3390/chemosensors9090248
Submission received: 1 July 2021
/
Revised: 8 August 2021
/
Accepted: 18 August 2021
/
Published: 3 September 2021
(This article belongs to the Section (Bio)chemical Sensing)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Respiratory diseases are top-ranked causes of deaths and disabilities around the world, making new approaches to the treatment necessary. In recent years, lung-on-a-chip platforms have emerged as a potential candidate to replace animal experiments because they can successfully simulate human physiology. In this review, we discuss the main respiratory diseases and their pathophysiology, how to model a lung microenvironment, and how to translate it to clinical applications. Furthermore, we propose a novel alveolus lung-on-a-chip platform, based on all currently available methodologies. This review provides solutions and new ideas to improve the alveolar lung-on-a-chip platform. Finally, we provided evidence that approaches such as 3D printing, organ-a-chip devices and organoids can be used in combination, and some challenges could be overcome.
1. Introduction
To understand the clinical applications of the lung-on-a-chip platform, we will briefly overview the main respiratory diseases and their pathophysiology, the available approaches to lung cell culture, modelling of the lung microenvironment and microfluidic systems.
Respiratory diseases are top-ranked causes of deaths and disabilities, with a high burden on health service costs across the globe. In the European Union, the total cost of respiratory disease accounts to more than €380 billion annually, including the costs of primary health care, hospitalizations, and lost production due to disability [1]. In 2017, the Forum of International Respiratory Societies identified the most common global causes of severe illness and death, the Big Five: chronic obstructive pulmonary disease (COPD), asthma, acute lower respiratory tract infections, tuberculosis and lung cancer [2].
1.1. COPD
COPD is responsible for approximately 6% of deaths, which makes it the third most fatal disease at a global level, with a higher burden in low- and middle-income countries [3]. COPD is typically caused by prolonged exposure to tobacco smoke, air pollution, chemicals, and dust. These factors produce an inflammatory pattern that progressively damages the peripheral lung tissues and causes obstruction of the small airways. As a consequence, decreased blood oxygen levels and increased carbon dioxide levels may result from abnormal gas exchange. Furthermore, narrowing of the airways leads to different degrees of airflow limitation that may impact total lung capacity, exercise capacity and quality of life.
1.2. Asthma
In the group of chronic respiratory diseases, asthma is the most prevalent, affecting 262 million people and killing 461,000 of them in 2019 [4]. Asthma has a multifactorial etiology that can be associated with maternal risk factors during pregnancy (e.g., low fiber diet, cesarean section, environmental exposure to allergens) and/or events that happen in the early life of an individual (e.g., exposure to allergens, use of antibiotics) [5]. These factors trigger Th1- or Th2-mediated immune responses that result in airway hyper responsiveness, smooth muscle dysfunction and airway remodeling [6]. An asthma attack typically involves wheezing, coughing and dyspnea.
1.3. Acute Lower Respiratory Tract Infections
Lower respiratory infections account for more than 4 million deaths annually in the world [2]. The most aggressive ones are pneumonia caused by Streptococcus pneumoniae, and respiratory syncytial virus (RSV) infections, presenting a high incidence of episodes and mortality in children of low- and middle-income countries. Persistent pneumonia and RSV infections are threatening because they can potentially lead to chronic respiratory diseases such as bronchiectasis and asthma, both characterized by airway mucus hypersecretion and difficult management with regular antibiotics [7]. On top of that, climate change also takes a toll on respiratory infections, as variations in temperature and air humidity contribute to increased viral transmission and the need for hospitalization [8].
1.4. Tuberculosis
Although, back in 1993, WHO declared tuberculosis a global emergency, this disease is sadly far from eradication at present. An estimated 10 million people developed tuberculosis in 2019 and 1.4 million died from the disease in that year [9]. Tuberculosis is a communicable disease caused by Mycobacterium tuberculosis, spread through the inhalation of droplets from the cough or sneeze of an infected person. This disease is especially threatening for people living with HIV, as they are more vulnerable to manifestations of tuberculosis symptoms. Despite being curable and preventable, tuberculosis is a very heterogeneous disease that may cause sequelae at different degrees in treated individuals. The most frequent ones include lung cavitation, fibrosis and nodular infiltrates that may lead to persistent lung dysfunction [10].
1.5. Lung Cancer
Among all types of cancer, lung cancer remains a major concern worldwide, with 2.2 million cases registered, being the most fatal one, with 1.8 million deaths in 2020 [11]. Although the pathophysiology of lung cancer is not yet fully understood, it is hypothesized that dysplasia of lung epithelium is triggered by repeated exposure to carcinogens, predominantly cigarette smoke, but also environmental pollutants (e.g., asbestos, arsenic, and chromium) associated with genetic susceptibility [12]. Lung cancer is divided into two major categories: small cell lung carcinoma (SCLC) and non-small cell lung carcinoma (NSCLC). The latter is more prevalent and represents 85% of all cases of lung cancer. Depending on the type of cells involved, NSCLC is further divided into the major categories of adenocarcinoma, squamous- and large-cell carcinoma [12]. Effective treatment approaches are available depending on the type of lung cancer and how far it has spread. However, even after the treatment, individuals may present long-term impairments in several outcomes including lung function, exercise capacity, quality of life, and depressive symptoms [13].
Aside from the Big Five, other respiratory conditions also impact global health. The inhalation of chemicals in the workplace causes a number of interstitial lung diseases including asbestosis, silicosis or coal worker’s pneumoconiosis. Obstructive sleep apnea (OSA) is another common condition, affecting 1–6% of adults [2]. It is characterized by the recurrent narrowing of upper airways, causing breathing interruption and hypoxia during sleep. Other, less common conditions involve alterations in the cardiovascular system, such as pulmonary hypertension, mainly related to left ventricular failure and lung conditions, and pulmonary embolism, a life-threatening condition caused by blockage of a lung artery by a blood clot (thrombus) [1,14].
More recently, in light of the current pandemic of coronavirus disease 2019 (COVID-19), more than 180 million cases and 3.8 million deaths were reported globally by June 2021 [15]. The disease pathophysiology begins with an asymptomatic phase in which individuals are highly contagious. The virus enters the cells and replicates locally in the nasal epithelium, then propagates with the infection of ciliated cells in the conducting airways. The majority of patients do not progress beyond this phase, as the normal immune response is sufficient to contain the spread of infection. However, approximately one-fifth of infected patients will progress with involvement of the lower respiratory tract, presenting more severe symptoms and complications such as acute respiratory distress syndrome (SARS-CoV-2), acute respiratory failure, sepsis, disseminated intravascular coagulation and others [16].
Hence, new approaches to the treatment of respiratory diseases are needed to reduce the length of hospital stay and the number of deaths. In this sense, the lung-on-chip platform appears to serve as a potential alternative to replace animal experiments and, more specifically, simulate human physiology.
2. Lung Cell Culture
The complex physiology of the lung, which could be translated into an engineering system, requires careful selection of the culture method and cell type. Although the two-dimensional (2-D) culture has been extensively used since the early 1900s, this method is not able to reproduce the in vivo cell–cell and cell–extracellular matrix (ECM) interactions [17]. Therefore, for the lung-on-chip platform based on microfluidic chip technology, we need to mimic the lung microenvironment, and the three-dimensional (3-D) culture is indicated.
The lung cell sources can be divided into primary cells, immortalized cell lines and stem cells. There are advantages and disadvantages to all cell sources. Primary cells are extracted directly from patient’s tissue and they include lung fibroblasts [18], small airway epithelial cells [19], bronchial epithelial cells [20] and alveolar type II cells/pneumocytes (AT2) [21]. However, primary cells are highly specialized and lose their proliferation abilities after a while, which is a problem if we desire to keep these cells in a microfluidic device for a prolonged period.
Adenocarcinomic alveolar A549 and NCI-H441 cell lines have commonly been used as a model for alveolar epithelium in different studies and seem to have a good ability to model lung injury and repair [22,23,24,25]. Furthermore, bronchial epithelial cell lines such as BEAS-2B are usually co-cultured with the epithelial cell line A549 to model airway epithelial cell injury in a cigarette smoke model [26], allergic airway inflammation [27], acute respiratory distress syndrome [28] and lung cancer [29]. However, it is unclear how the cell lines maintain the normal physiology of airway cells, which can be a limitation regarding in vitro models and microfluidic devices.
When using stem cells for in vitro models, it is desirable to differentiate them along pulmonary lineages by providing an environment where it is possible to mimic the lung microenvironment. For this reason, we believe that microfluidic devices could be an ideal candidate for this source of cell. Embryonic stem cells (ESCs) have been demonstrated to be able to generate both airway and alveolar cell types in vitro, and could eventually be used in therapeutic and regenerative applications [30]. However, there is a lot of ethical controversy about the use of ESCs and, for this reason, induced pluripotent stem cells (iPSCs) can be used as an alternative resource. Recent data demonstrated that iPSCs can generate type II alveolar cells (iAEC2s) by directed differentiation, and may provide a platform for disease modeling [31]. Unfortunately, deriving mature lung lineages from iPSC precursors is challenging, even with the expertise from researchers regarding the differentiation protocols for producing ectodermal and mesodermal lineages [32]. We must remember that the lung alveolus and airways are composed of multiple epithelial, endothelial and mesenchymal cell types. Therefore, the use of a combination of cells provides physiological relevance and should be considered during applications of regenerative medicine and engineering systems.
Regarding the choice of cell type for culture on the lung-on-chip platform, the specific clinical application should be considered. Asmani and colleagues cultured primary normal human lung fibroblasts (NHLF) or human lung small airway epithelial cells (SAECs) mixed with collagen type-I, and created a fibrotic microtissue system to study idiopathic pulmonary fibrosis [18]. In another study, human airway epithelial cells (Calu-3) were cultured on one side of the microfluidic device and on the other side, to achieve the air–liquid interface (ALI), with human bronchial smooth muscle cells (hBSMCs). Therefore, this model was efficient to study the interaction between the cell matrix in chronic lung diseases [33]. Zhang and colleagues recreated the human alveolar–capillary barrier to evaluate the pulmonary toxicity of nanoparticles [34]. They used a lung-on-a-chip platform in which human alveolar epithelial cells (HPAEpiCs) were cultured on the alveolar side and human umbilical vein endothelial cells (HUVEC) on the capillary side.
3. Modelling Lung Microenvironment
The architecture of an organ is intimately related to its function. The structural design of the lung has been optimized to fulfill its primary function, which is gas exchange. During the branched morphogenesis of the lung, the airway tree is positioned adjacent to the arterial vasculature and ultimately branches into hundreds of millions of alveoli sacs with a diameter ranging between 100 and 200 µm. Given the high number of alveoli sacs and the thin tissue layer that separates air from blood (approx. 500 nm), the lungs offer a large surface area (100 m2) in a relatively small tissue volume (6 L) to support fast gas exchange. This enables the perfused blood to gather oxygen molecules and release carbon dioxide into the atmosphere in a short time [35].
Given the dynamic nature of lung function, lung cells continuously experience various types of mechanical forces, which are known to deeply impact their function and phenotype [36]. Notably, lung cells undergo biophysical stimulation consisting of: (1) cyclic stretching due to expansion and contraction during respiration; (2) shear stress associated with airflow into the lungs and vascular perfusion; (3) local changes in the composition and hence, stiffness and viscoelastic properties of the ECM; (4) local variations in oxygen partial pressure; (5) the presence of an air–liquid interface in the airway epithelium. Although these stimuli are all of a physiological nature, it is worth noting that lung cells are often exposed to non-physiological stimuli as a result of disease or injury. Several examples of this include ventilation-induced lung injury (VILI), stiffening of the ECM in lung cancer or lung fibrosis and changes in oxygenation levels associated with chronic respiratory diseases including COPD and OSA.
The use of animal and 2D in vitro models helped us understand the bases of lung physiology and disease. However, they present many limitations. Studies involving animal models have ethical concerns, are laborious, the interpretation of results is not always straightforward, and they have limited predictability in human clinical trials due to the differences between species. Although conventional 2D models remain the tool of choice for many researchers, they fail to reproduce the architecture, mechanical and functional properties inherent to native tissues, thus hampering the interpolation of results. These types of cultures focus more on the effect of biochemical factors, but it is well known that biophysical factors are equally important in determining cell behavior and need to be considered [37]. Therefore, understanding the mechanisms that underlie lung physiology and disease requires the development of alternative 3D in vitro models to enable the precise application and tight control of the most relevant biochemical and biophysical cues found in the lung cell niche.
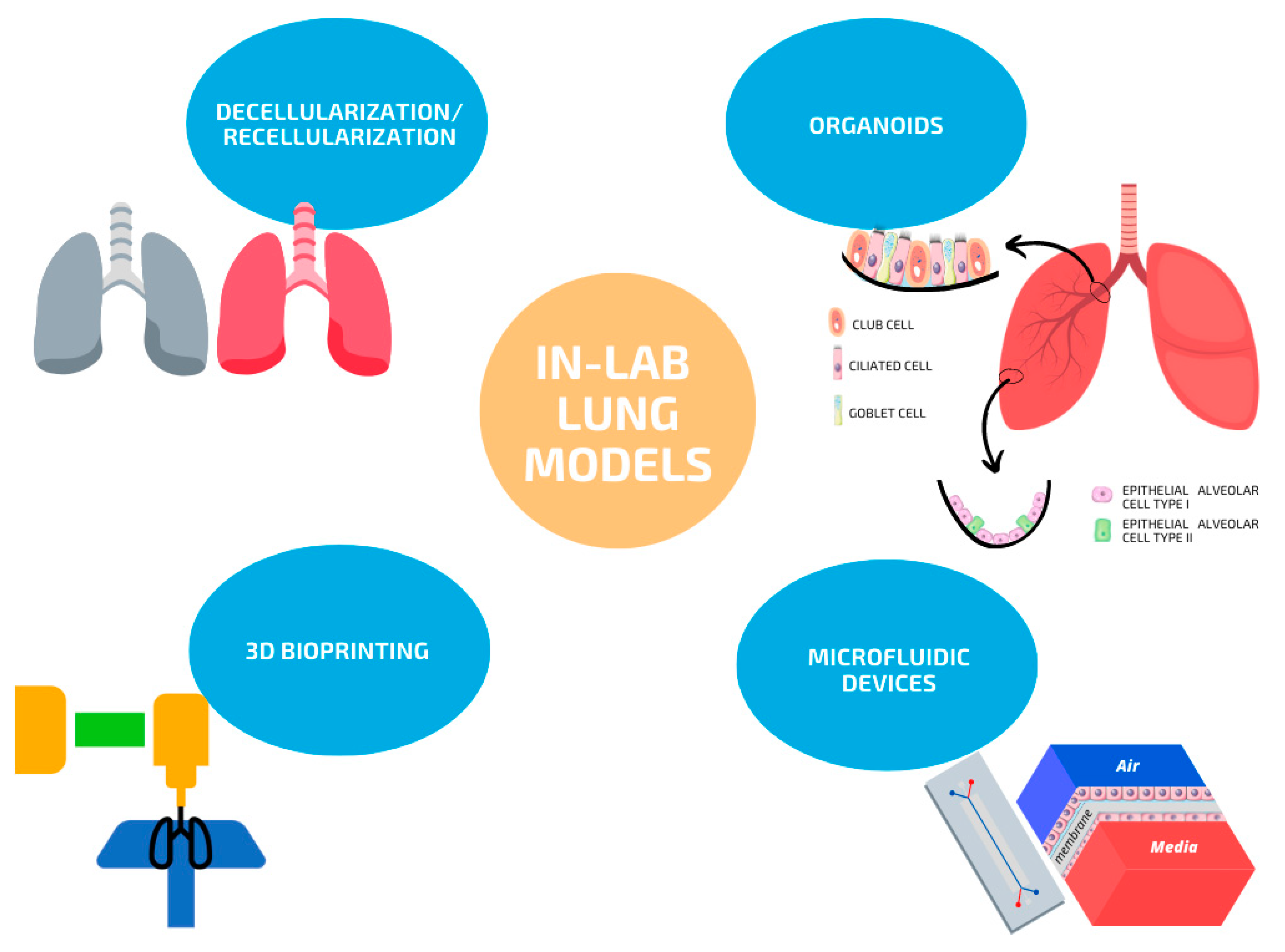
Top-down and bottom-up approaches have been used for the generation of lung models in the laboratory. Top-down strategies refer to lung de- and recellularization with either lung cells, stem cells or their combination, while bottom-up strategies include the generation of organoids, 3D bioprinting and microfluidic lung systems. An overview of the current strategies available for in-lab lung models is depicted in Figure 1.
3.1. Organ Decellularization/Recellularization
The ECM is a non-cellular 3D structure present within all tissues and is essential for life. Every organ in our body has a unique ECM that, in addition to providing physical support for tissue integrity and elasticity, also confers biomechanical cues and regulates diverse cell functions including migration, proliferation and differentiation [38]. At the same time, cells synthesize, degrade, reassemble and chemically modify the components of the ECM. Therefore, its structure is highly dynamic and undergoes continuous controlled remodeling to maintain tissue homeostasis, meaning that the presence and relative abundance of each ECM component may vary over the lifetime according to continuous remodeling. In addition, excessive or uncontrolled ECM remodeling leads to changes in its structure and composition. Such changes directly impact cell behavior and are considered contributors to several pathological conditions such as pulmonary fibrosis and cancer [39].
Given the structural complexity of the lungs and the key role that the ECM plays during lung development and adulthood, organ decellularization has been one of the preferred strategies for the development of biomimetic lung 3D models over the past decade [40]. Through this technique, lung donor cells are removed while the acellular 3D scaffold retaining the biochemical components, mechanical properties and the structural integrity of the native lungs (including the original vasculature) prevails.
Numerous studies on lung decellularization and recellularization for tissue engineering purposes have been carried out by us and other researchers [41,42,43,44]. In this regard, rodent, porcine and human lungs have successfully been decellularized and recellularized, mainly with lung endothelial and epithelial cell lines [41], lung endothelial and epithelial primary cells/progenitors derived from iPSCs or ESCs [45,46,47], or with a combination of different cell types, including bronchial epithelial cells, vascular endothelial cells, lung fibroblasts and mesenchymal stromal cells [42]. A combination of physical, chemical and biological agents is employed for the decellularization of small sections (∼1–3 cm3) [42] and whole lungs [40,41,44]. Cell lysis is usually achieved by means of applying several freezing and thawing cycles, followed by osmotic shocks and subsequent instillation with detergent-based solutions. Some protocols also include a final treatment with DNase to degrade residual DNA fragments once cells have been lysed. Although these works represented a milestone in lung bioengineering with promising future clinical applications, the use of organ decellularization and recellularization has been hampered by some limitations. The main ones are the lack of concurrence on the best method for organ recellularization and the lack of bioreactors allowing for the long-term maintenance of recellularized lungs in the lab. In particular, the task of effective recellularization remains unsolved, mainly due to the lack of consensus about which cell/s type/s and delivery methods would be the most suitable for lung recellularization, together with the need to obtain large cell numbers and adequate cell positioning within entire lobes and whole lungs, which largely depends on the seeding method. On the other hand, investigation into the technology required to sustain the engineered tissue in the long-term is scarce, due to complexity, specialization and costs.
3.2. Organoids
Organoids are stem-cell-derived, self-organized 3D structures, able to recapitulate the primary functions and structure of the tissue of interest. This approach enables the generation of simplified, miniaturized organs, which, when patient-derived, are suitable for research and envisioning personalized medicine. Classic methods for generating organoids involve the use of either pluripotent or tissue-derived stem cells. The generation of organoids from pluripotent stem cells requires a first step for the determination of the germ-layer (endoderm, mesoderm or ectoderm), followed by the sequential addition of specific signaling factors to differentiate into the tissue of interest. On the other hand, organoids derived from adult stem cells require cell isolation from the target tissue, have epithelial origin and, by contrast, do not contain mesenchymal or immune cells unless added separately.
Lung organoids have been developed from human pluripotent stem cells, as well as from epithelial stem/progenitor cell populations within the adult lung. Both basal and secretory club cells from the upper and middle airways and AT2 cells from the lower airways are common choices for their generation [48]. Despite allowing for numerous advances in basic and translational research, they present several limitations which complicate their potential applications in the field of lung development, regenerative medicine, disease modeling, drug safety and efficacy testing. Such limitations are mainly associated with the absence of other lung stroma cell types, such as fibroblasts, which contribute to the generation of ECM and its remodeling, or immune cells, as well as the lack of a vascular network, a poorly defined 3D architecture, etc. Nonetheless, if combined with other approaches such as organ decellularization, 3D bioprinting or organ-on-a-chip devices, some of these drawbacks could be overcome.
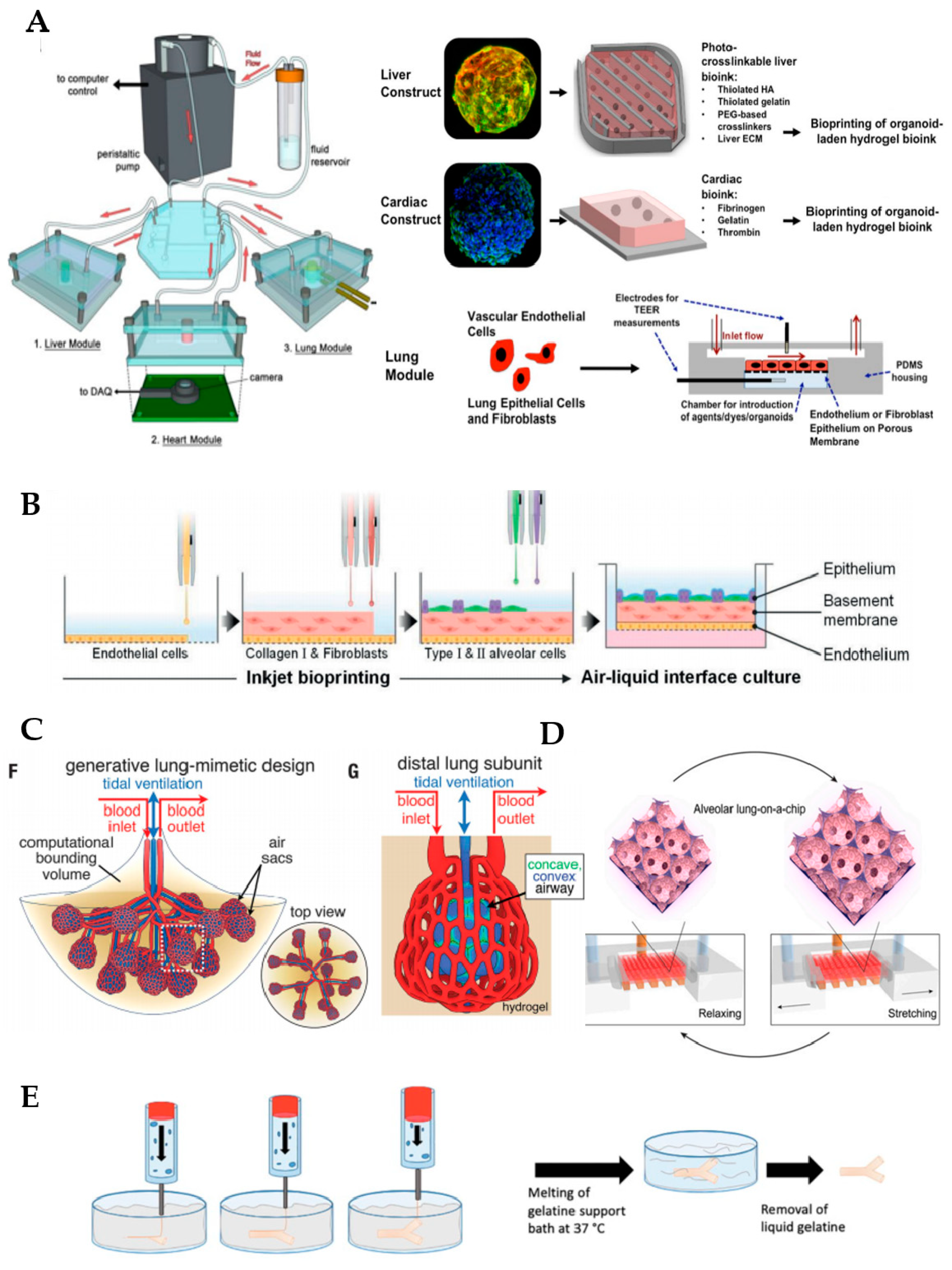
Organoid cultures classically involve the use of Matrigel, a commercially available matrix derived from the Engelbreth–Holm–Swarm mouse tumor, as a supportive material. Nonetheless, this material is afflicted with limitations associated with its ill-defined composition, and lack of reproducibility and tunability, generating some concerns regarding the study of normal regenerative processes. Therefore, hydrogels generated from lung decellularized ECM (dECM) have recently emerged as a potential option for culturing organoids, since they provide the unique environmental cues found in lungs in vivo. A recent comparative study has revealed that lung organoids derived from human bronchial epithelial cells maintain their identity in dECM hydrogels while those cultured in Matrigel show an upregulation of the oncogenic markers and pathways that are known to be aberrant in cancer, thus supporting the initial idea [49]. Other researchers have demonstrated the feasibility of combining the bioprinting of bioengineered organoids with microfluidic systems [50], as we can see in Figure 2. These authors developed a three-tissue organ-on-a-chip closed circulatory perfusion system, composed of liver, heart, and lung, which facilitates the study of inter-organ responses. In this platform, each of the aforementioned tissues is housed in a microfluidic polydimethylsiloxane (PDMS) device fabricated by soft lithography and replica molding. Liver and cardiac modules were created by bioprinting spherical organoids within customized bioinks, while the lung module was formed by the creation of layers of cells (lung fibroblasts, epithelial and endothelial cells) around a porous polyester membrane coated with human lung dECM. Microfluidic devices were connected to a circulatory perfusion system through a central fluid routing breadboard (Figure 2A). The authors first demonstrated the correct performance of each of the individual components of the platform by evaluating their response to a panel of drugs, including bleomycin, propranolol and acetaminophen, amongst others. More importantly, when combined in a single platform, more complex integrated responses were observed, indicating that the functionality of one organoid was deeply influenced by the response of another organoid (for example, bleomycin likely evoked an inflammatory response in lung tissue with the release of inflammatory cytokines with cardiotoxic properties). This and other works in the field highlight the critical relevance of creating combinatory approaches and integrated systems to enhance the predictability of potential drugs and therapeutic in vitro studies.
3.3. 3D Bioprinting
3D bioprinting is an emerging technology that has attracted considerable interest from the healthcare sector in recent years, especially for its applications in regenerative medicine. By automated additive manufacturing of cells and biomaterials (bioinks) of different nature, this technique enables the customized fabrication of relatively complex 3D structures in a reproducible manner that can potentially be used as lung tissue models. Fabrication methodologies include extrusion-based, inkjet-based, laser-assisted bioprinting and stereolithography (SLA) [51]. Extrusion-based is the most widely employed due to its simplicity, affordability, flexibility and scalability. In this technique, the application of pressure allows for the dispensing of a continuous filament of material from the syringe to the stage. Depending on the dispensing mechanism employed, this is categorized into pneumatic, piston-driven and screw-driven dispensing. Despite the previously mentioned advantages, this technique presents some limitations, such as a lower resolution and generation of higher shear stress, when compared with others. On the other hand, inkjet-based bioprinting is a non-contact printing technique that uses a drop-on-demand technology, allowing for a highly accurate deposition of very small droplets of material, which results in the generation of 3D structures with higher resolutions. When using laser-assisted bioprinting, the bioink is projected from a film to the depositing stage by using a laser beam as a driving force to trigger the droplet release; the laser evaporates the bioink, creating an expanding bubble with the subsequent jet formation and the final deposition of the droplet onto the substrate. This method is the most widely used for the high-resolution patterning of bioinks. Similar to laser-assisted bioprinting, SLA is a nozzle-free technique that uses digital light to cure the liquid bioink in a layer-by-layer fashion.
In the respiratory system, most of the progress has been restricted to conducting airways [52,53] due to the complexity of recreating alveolar sacs. These and other studies showed that the generation of 3D structures with the desired organization and composition via 3D bioprinting can be feasible. However, before bioprinting of complete tissues and organs suitable for transplantation becomes a reality, there are many challenges that need to be faced.
As mentioned previously, one of the most remarkable challenges in lung bioengineering is the generation of a vascular network closely adjacent to the alveoli. By using 3D bioprinting technology, Kang et al. (2021) fabricated a three-layered alveolar barrier with a total thickness of approximately 10 µm in a transwell insert [54]. The bottom layer consists of lung endothelial cells, the middle layer of a collagen I hydrogel containing fibroblasts and the top layer contains type I and II alveolar epithelial cells deposited by automated high-resolution drop-on-demand inkjet printing (Figure 2B). The authors demonstrated that this approach can recapitulate the structure, morphology and function of the lung tissue, and could potentially be used as disease models to study influenza infection, amongst others. Grigoryan et al. (2019) formed photopolymerizable PEGDA-based hydrogels containing intricate and functional vascular architectures through the addition of biocompatible food additives, including tartrazine, which work as photoabsorbers for projection stereolithography [55]. By using this breakthrough technique, authors were able to reproduce the lung multivascular network surrounding alveolar sacs and demonstrated its functionality: the tissue resists the pressure generated during pulsatile breathing (i.e., cycles of intake and outflow of air simulating pressures and frequencies in human lungs) and blood flow (Figure 2C). More intriguingly, blood cells can be reoxygenated as they flow through the vascular network surrounding the air sac, thus demonstrating that the main function of the lungs—gas exchange—is completely fulfilled. This important development solves the longstanding questions about how to vascularize bioprinted tissues, which is fundamental to their maturation in the lab and in transplantation. In addition, the technique can be extrapolated to the biofabrication of other organs, including the liver.
Another challenge is finding a consensus on the most suitable bioink for applications in lung bioengineering. The predominant components of lung ECM in adulthood are collagens (mostly types I, III and IV), elastin, proteoglycans and glycoproteins [56]. However, the majority of natural polymers used in bioprinting, which include alginate, collagen or fibrin do not present adequate rheological properties for 3D bioprinting when used alone and are not able to provide the complexity found in the native organ, which is essential for accomplishing cell functions. Building hybrid biomaterials from decellularized lung ECM combined with synthetic or other natural materials is a strategy that has attracted the attention of researchers since it solves the current issues associated with the bioprinting of either natural or synthetic materials alone.
De Santis et al. (2021) made significant advances in this regard by improving both the properties of the bioink and the bioprinting methodology employed in the generation of human small airways composed of the regionally specified primary human airway epithelial progenitor and smooth muscle cells [57]. By using the Freeform Reversible Embedding of Suspended Hydrogels (FRESH) method, which uses thermo-sensitive gel baths or sacrificial materials such as gelatin, collagen or pluronic as supportive materials that degrade after bioprinting the structure of interest, they were able to generate branched small airways with a good resolution that remain patent for up to one month in vitro (Figure 2E). In addition, the bioink of choice consisted of an alginate reinforced with ECM from human decellularized lungs, which keeps the natural components and the biological inductive properties of native tissue (low immunogenicity, proangiogenic properties, etc.) while conferring proper rheological and gelation properties for the 3D bioprinting process. In collaboration with other authors, the group developed a hybrid hydrogel where a clickable decellularized lung ECM crosslinker was incorporated into poly(ethylene glycol)-α-methacrylate PEGαMA in order to reproduce controllable spatiotemporal changes in its stiffness, with applications in diseases such as lung fibrosis [58].
Despite the current advances, once the 3D structure is bioprinted, the main challenge is to maintain the bioprinted tissue in a proper microenvironment to support its maturation, thus enabling the performance of basic and translational research. At this step, the use of bioreactors to support bioprinted lung tissue maturation through biomimicry is essential, but they must be custom-designed for each bioprinting application. The design, fabrication and performance of the bioreactors required to that goal is quite complex. To our knowledge, there are no commercial bioreactors that are able to provide the growing tissue with the physiological stimulation of the lung (including mechanical stretch and gas and fluid perfusion). However, the use of miniaturized bioreactors to maintain small constructs and organ-on-a-chip systems has been extended along the academic sector and in the industry.
4. Microfluidic Lung Systems
Microfluidic lung systems constitute an effective and affordable small-scale alternative, where single cultures, co-cultures and even 3D cultures can be maintained under tightly controlled conditions. These models allow for the performance of more realistic experiments where specific microenvironmental parameters such as oxygen tension, shear stress and mechanical stretch can be precisely and individually controlled and are compatible with live cell imaging. Therefore, these platforms have the potential to provide new insights into normal lung function and disease, as well as to more accurately predict the effectiveness and safety of new treatments under a microenvironment biomimicking the lung [61].
To date, research efforts have mainly been conducted to recreate alveoli, bronchi and bronchioles. The first lung-on-a-chip device was fabricated by Hugh and coauthors (2010), who recreated the blood–air barrier [62]. Through the use of soft lithography, they generated a PDMS device consisting of two microchannels, separated by a 10 µm thick, porous and stretchable membrane coated with proteins of the ECM. The upper side of the membrane was seeded with human alveolar epithelial cells, while the lower side was seeded with lung capillary endothelial cells. By depleting the cell culture media of the upper channel and maintaining a continuous media flow in the lower channel (vascular), researchers generated an air–liquid interface similar to the alveolar–capillary barrier. The simulation of physiological breathing was also feasible by cyclically stretching cells through the application of vacuum, which deforms the elastic membrane to which cells are adhered. Using this model, Hugh et al. studied lung physiological processes such as the differentiation potential of epithelial progenitor cells, carried out toxicological studies and modeled the pulmonary edema and thrombosis [63].
Other microfluidic lung systems modeling the alveolar region were set up in other studies, using a similar working principle [59,64]. In some of these models, the flat and synthetic porous membrane was replaced by a biological membrane containing proteins of the ECM and relevant 3D shapes, thus conferring a more physiologically relevant 3D microarchitecture and enhancing their biological performance [59,64]. Although PDMS has been the synthetic material of choice for the fabrication of most lung-on-a-chip devices and their membranes, it presents a high absorption of small molecules such as hydrophobic drugs, thus hindering the widespread use of PDMS-based microfluidic devices for drug screening. Some PDMS-treatments have been proposed to overcome this problem, as well as alternatives consisting of the use of hydrogels. Huang et al. (2021) generated a 3D porous gelatin methacryloyl (GelMa) hydrogel, presenting an alveolar sac-like 3D structure (Figure 2D) [59]. Amongst the advantages of using GelMa instead of synthetic membranes, the stiffness (6.23 ± 0.64 kPa) is within the range reported for native lungs [65]. In addition, GelMa is not biologically inert, since it contains arginine–glycine–aspartic acid sequences that promote cell attachment, and matrix metalloproteinase-responsive peptide motifs, which facilitate its remodeling and promote processes such as cell migration and proliferation. Besides all these features conferred by GelMa, a mechanical stimulation simulating breathing is also feasible with this model. Multiscale analyses revealed that opal structures resembling alveoli sacs were more adequate than planar membranes made by GelMa or PDMS in maintaining the functions of primary human alveolar epithelial cells. Zamprodogno et al. (2021) created a thin, elastic, biological membrane by drop-casting a collagen–elastin solution onto a thin gold mesh with a poresize of 260 µm that supports the array of 40 alveoli [66]. This membrane showed a good performance, enabling the recreation of the air–liquid interface and application of mechanical stretch to reproduce respiratory motions. Therefore, such microfluidic devices are considered a second generation of organ-on-a-chips, offering a higher level of analogy to whole-organ systems through the mimicry of biological barriers.
Several groups have created lung airway-on-a-chip, aiming to model lung physiology and diseases that primarily affect the airways, such as asthma or COPD [66,67,68,69]. Sellgren et al. (2014) created a biomimetic microfluidic device composed of three individually accessible and vertically stacked compartments, separated from each other by a synthetic nanoporous membrane [67]. From top to bottom, the compartments contained either tracheobronchial epithelial, lung fibroblasts or endothelial primary cells, cultured on an air–liquid interface. Similarly, Humayun et al. (2018) designed a three-compartment device where a suspended hydrogel composed of a mixture of collagen I and Matrigel was injected in the middle compartment to act as a barrier between airway epithelial and smooth muscle cells, thus avoiding the use of synthetic porous membranes to recreate the lamina propria [33]. The accommodation of air–liquid culture conditions normally resulted in the formation of a pseudostratified, differentiated bronchiolar epithelium, confirming the good performance of the models. Some of these models also reproduced respiratory symptoms including asthmatic inflammation and were used as platforms to test drug efficacy under a microenvironment that biomimics the pathology [68,69].
Lung-on-a-chip models providing mechanical stimuli, aiming to study mechanical injury caused by a cyclic stretch in VILI or by movement of liquid plugs mimicking those diseases in which pulmonary surfactant production is compromised (asthma, pneumonia, cystic fibrosis, etc.), have been developed [70,71]. Regarding VILI, Tas et al. (2021) recently developed a lung-on-a-chip model using 3D printable moulds and a commercial nanofibrous poly(caprolactone) membrane. The thin (∼20 µm) membrane was cast between two PDMS channels fabricated from 3D printed molds [70]. Proof-of-concept experiments using murine lung epithelial cells showed the good performance of the model, which recreates the air–liquid interface while providing mechanical stretch at relevant magnitudes (25%) without the need to employ complex biofabrication techniques. On the other hand, Nonaka et al. (2020) designed a computer-controlled PDMS chip to expose rat lung mesenchymal stem cells (MSCs) cultured in lung ECM to realistic biochemical and stiffness substrate cues while applying cyclical stretch simulating ventilation (20% amplitude, 12 cycles/min), aiming to determine whether biophysical preconditioning could potentiate the immunomodulatory properties of MSC [72]. Dynamic stretch was generated by the application of cyclical positive pressure underneath a flexible membrane where 3D cultures were attached. After biophysical preconditioning for 7 days, MSCs were applied through femoral venous injection to treat mild VILI in Sprague Dawley rats. Amongst the other results, only preconditioned MSCs were found to induce a significant recovery in elastance. Although this is the first study to demonstrate that biophysically preconditioned MSCs were more effective than non-preconditioned MSCs in reducing mild VILI in a rodent model, more detailed studies highlighting the importance of biophysical stimuli to enhance translational research are required.
It is well known that oxygen levels have a direct impact on processes such as cell proliferation, cell migration and stem cell differentiation. The prediction of oxygen levels within the microfluidic lung systems is of vital importance to create adequate oxygen tension to study cellular behavior under realistic conditions. The most common oxygen-sensitive indicators employed to measure oxygen levels in microfluidic devices are optical fibers and oxygen-sensitive compounds including ruthenium- and metalloporphyrin-based molecules. A comprehensive review of these and other oxygen sensor types for applications in microfluidic systems was published by Grist et al. (2010) [73]. In previous works, we and coworkers fabricated simple devices that allowed for the fast diffusion of gases to the cell culture through thin elastic and gas-permeable PDMS membranes [74,75,76]. When connected to commercial gas blenders or to two gas reservoirs providing different oxygen concentrations (i.e., 20% and 1% O2), through an automated two-way solenoid valve, cells can undergo cyclic changes in PO2 in short periods of time (approx. 6 s), as determined by an oxygen-sensitive ruthenium dye and optical fiber. In addition, some of these models allowed for the application of cyclic stretch at a frequency and amplitude simulating lung breathing [72,74]. Although these systems did not recreate the complex 3D architecture of the alveolar barrier or incorporate shear stress through microfluidics, because they were out of the scope, their simple fabrication, low cost and ability to apply several biophysical cues to cultured cells made them attractive tools. These models have a special relevance for the study of the molecular mechanisms underlying disorders such as OSA and lung cancer, where changes in oxygen tension play a pivotal role in determining their fate and new treatment approaches. In this line, the increased expression of inflammatory molecules such as prostaglandin E2 has been observed in macrophages in response to intermittent hypoxia (OSA model), as well as their increased migration towards hypoxic cancer cells, likely due to an increased release of cytokines with chemoattractant properties by the latter [75,76].
Moreover, Benam et al. (2016) described a model of COPD where the living human bronchiolar epithelium from patients was connected to a device that “breathes”cigarette smoke in and out of the small airway-on-a-chip. This smoking human small airway chip allowed for identification of the COPD-specific molecular signatures, ciliary micropathologies, and epithelial responses to smoke generated by whole cigarettes and electronic cigarettes [77]. Furthermore, this provided a tool for studying airway pathophysiology and biological responses in COPD patients.
To mimic the disease environment, Asmani et al. (2018) considered the effect of tissue stiffening on the progression of lung fibrosis. They developed a microfluidic chip technology, where membranous human lung microtissues were created to model key biomechanical events occurring during lung fibrogenesis [18]. In this sense, the tissue stiffening has to be considered in the lung cancer as well. Yang et al. (2016), developed a lung-on-a-chip microdevice to test anti-cancer drugs [78]. On this platform, they created a membrane made by poly(lactic-co-glycolic acid) (PLGA) nanofiber with a controlled stiffness, which simulated the alveolar respiratory and allowed for the invasion of cancer cells. In those studies, it was clear that all features of the disease must be considered and can be mimicked on a microfluidic chip platform.
Recently, the combination of different approaches has been attracting the attention of biomedical scientists, physicists and engineers. The integration of organoids and 3D bioprinting with organ-a-chip technology attempts to gather the strengths of each of these strategies and increase their versatility. As an example, both organoid-on-a-chip and organ-on-a-chip fabrication by 3D bioprinting have promising applications in pharmacology because (1) either organoids or 3D bioprinted structures are more similar to actual organs, making them more suitable for target identification and future validation and (2) organs-on-a-chip are more controllable and reproducible devices, which are best suited for drug efficacy and safety screenings [52,79].
5. Future Applications
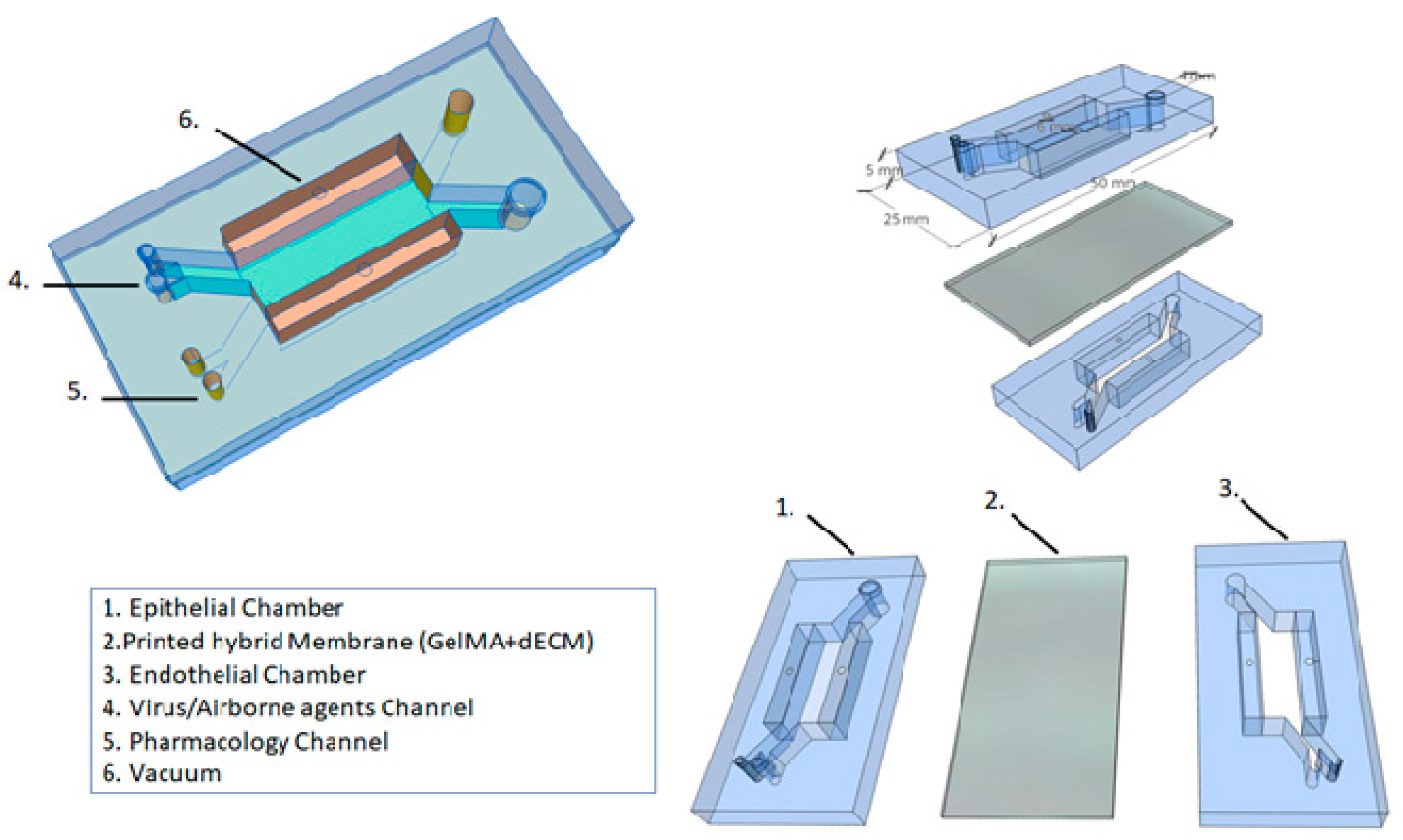
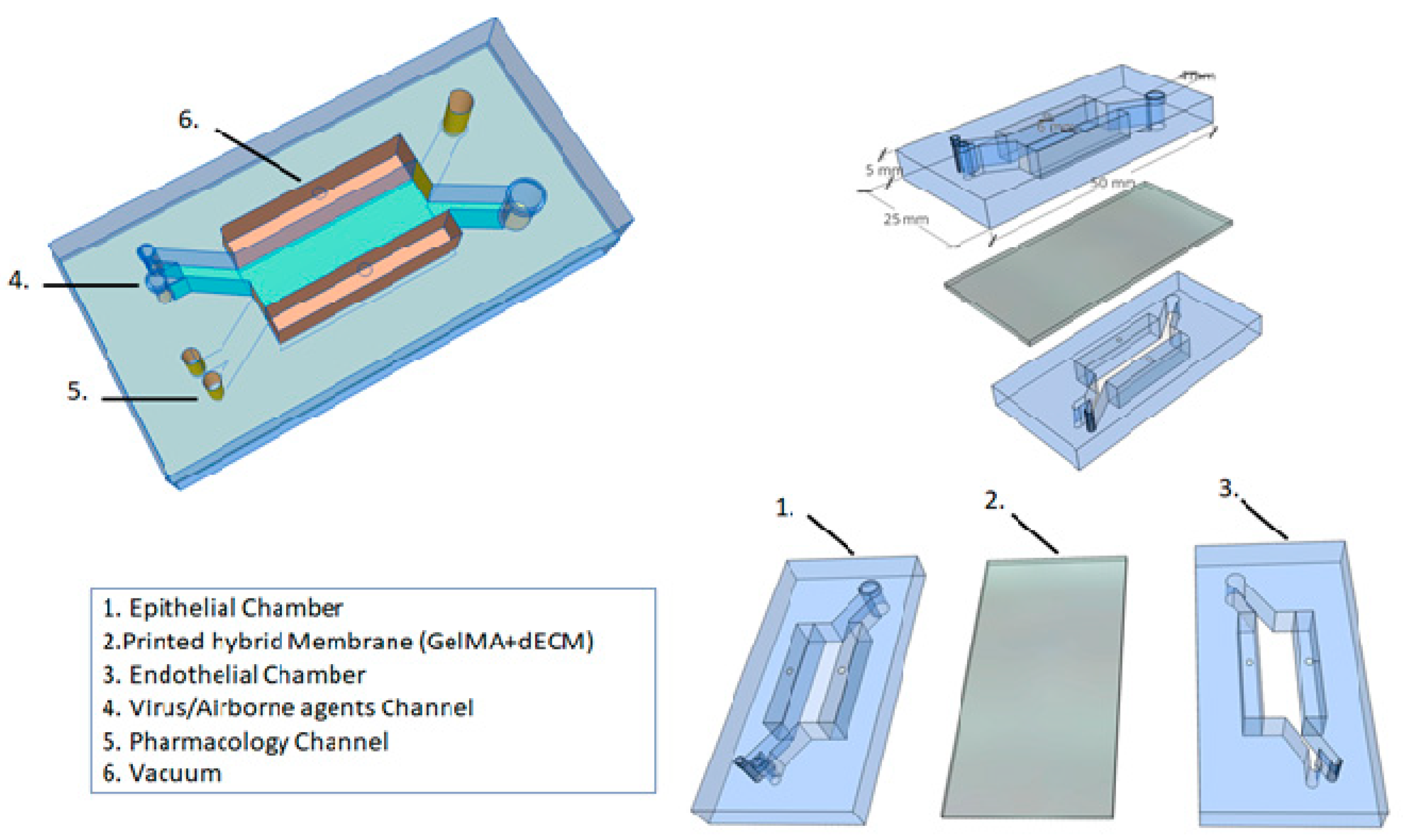
Based on these different approaches to the development of microfluidic platforms, and considering the future clinical applications, we propose a novel alveolar lung-on-a-chip platform (Figure 3). This microfluidic device has a combination approach, considering the best technologies in the field of regenerative medicine. We believe that it is possible to create a device in which a viral infection (e.g., SARS-CoV-2) can be induced, or the respiratory epithelium can be exposed to airborne environment agents (e.g., tobacco smoke, pollutants). Additionally, in this model, a pharmacological treatment could be applied.
Regarding the structure, our device can be generated by soft lithography using PDMS, and separated by two aligned upper and lower microchannels (epithelial and endothelial) to simulate the alveolar–capillary barrier. The device contains two lateral vacuum chambers to mimic the physiological respiratory cycle. Each of those chambers will have a tube to allow for control of the flow of the culture medium, oxygen levels, percentage of CO2, and percentage of mechanical strain applied by vacuum to the side chambers on the hybrid membrane.
5.1. Printed Hybrid Membrane
Several authors use the porous and stretchable membrane coated with ECM proteins to model a 3D environment for the cells in a lung-on-a-chip device. In our device, we propose a bioprinting technology associated with a dECM. We previously discussed the role of dECM in cell functions and remodeling processes. However, to shape the membrane, which will recreate the alveolar–capillary barrier, the GelMa hydrogel will be printed together with dECM. In this way, it is possible to recreate human alveolar sacs with advantages for the cells, stiffness and biological cues. The hybrid membrane can be printed with 10 µm thickness, and guarantee porosity and stretchability during strain in the respiratory cycle.
5.2. Alveolar Lung-on-a-Chip Culture
In the proposed device, we aim to solve some problems faced by cell cultures in microfluidic technology. For this purpose, organoids will be employed. It is clear that organoids and organ-on-a-chip platforms have the same goal: to recreate the complexity of human organs in vitro [74]. The organoids (hPSCs differentiate into iAEC2s) will be cultured on top of the hybrid membrane (epithelial channel) and lung microvascular endothelial cells (HMVEC-L) will be cultured on the opposite side of the same membrane (endothelial channel). The alveolar lung-on-a-chip will be exposed to cyclic strain (10% strain, 0.2 Hz) to mimic the physiological breathing and continuous flow (60 μL/h) by media through the top and bottom channels for 21 days. With the cell culture in the device, it is possible to induce a viral infection or to expose the alveolus to airborne environment agents through the inlet channel (virus/airborne agents channel), which provides direct access to the epithelial channel. Considering the COVID-19 pandemic, a model of SARS-CoV-2 infection could be applied to understand the disease mechanism and explore treatment options. In this device, there is a drug channel that gives direct access to the endothelial channel, which allows for some pharmacological tests to be run. Furthermore, this device will also enable morphological analysis, epithelial barrier evaluation and 24 h cell tracking.
We hope this platform allows for the creation of a translational medicine, where we can recreate and treat lung diseases. Here, we can provide a precise medicine, where the patient can receive individualized treatment. It is possible to develop studies based on the extraction of cells from patients and more realistic modeling of lung diseases.
6. Conclusions
It is well known that a lot of work and effort is required to develop and improve the organ-on-a-chip methodology. However, this review provides solutions and new ideas to improve the alveolar lung-on-a-chip platform. We provided evidence that approaches such as 3D printing, organ-a-chip devices and organoids can be used in combination, and some challenges could be overcome.
Author Contributions
Conceptualization, writing, reviewing and editing: R.K.d.P., V.R.O. and N.C.; supervision: R.K.d.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gibson, G.J.; Loddenkemper, R.; Lundbäck, B.; Sibille, Y. Respiratory health and disease in Europe: The new European Lung White Book. Eur. Respir. J. 2013, 42, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Forum of International Respiratory Societies. The Global Impact of Respiratory Disease, 2nd ed.; European Respiratory Society: Sheffield, UK, 2017. [Google Scholar]
- World Health Organization (WHO). The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 30 June 2021).
- World Health Organization (WHO). Chronic Respiratory Diseases: Asthma. Available online: https://www.who.int/news-room/q-a-detail/chronic-respiratory-diseases-asthma (accessed on 30 June 2021).
- Thorburn, A.N.; McKenzie, C.I.; Shen, S.; Stanley, D.; Macia, L.; Mason, L.J.; Roberts, L.K.; Wong, C.H.; Shim, R.; Robert, R.; et al. Evidence that asthma is a developmental origin disease influenced by maternal diet and bacterial metabolites. Nat. Commun. 2015, 6, 7320. [Google Scholar] [CrossRef] [PubMed]
- Cereta, A.D.; Oliveira, V.R.; Costa, I.P.; Guimarães, L.L.; Afonso, J.P.; Fonseca, A.L.; Sousa, A.R.; Silva, G.A.; Mello, D.A.; Oliveira, L.V.; et al. Early Life Microbial Exposure and Immunity Training Effects on Asthma Development and Progression. Front. Med. 2021, 8, 662262. [Google Scholar] [CrossRef]
- Montefusco-Pereira, C.V.; Carvalho-Wodarz, C.S.; Seeger, J.; Kloft, C.; Michelet, R.; Lehr, C. Decoding (patho-)physiology of the lung by advanced in vitro models for developing novel anti-infectives therapies. Drug Discov. Today 2020, 26, 148–163. [Google Scholar] [CrossRef]
- Patella, V.; Florio, G.; Magliacane, D.; Giuliano, A.; Crivellaro, M.A.; Di Bartolomeo, D.; Genovese, A.; Palmieri, M.; Postiglione, A.; Ridolo, E.; et al. Air Pollution and Climate Change Task Force of the Italian Society of Allergology, Asthma and Clinical Immunology (SIAAIC). Urban air pollution and climate change: “The Decalogue: Allergy Safe Tree” for allergic and respiratory diseases care. Clin. Mol. Allergy 2018, 16, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO). Tuberculosis. Available online: https://www.who.int/news-room/fact-sheets/detail/tuberculosis (accessed on 30 June 2021).
- Ravimohan, S.; Kornfeld, H.; Weissman, D.; Bisson, G.P. Tuberculosis and lung damage: From epidemiology to pathophysiology. Eur. Respir. Rev. 2018, 27, 170077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO). Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 30 June 2021).
- Zheng, M. Classification and Pathology of Lung Cancer. Surg. Oncol. Clin. N. Am. 2016, 25, 447–468. [Google Scholar] [CrossRef]
- Cavalheri, V.; Jenkins, S.; Cecins, N.; Gain, K.; Phillips, M.; Sanders, L.H.; Hill, K. Impairments after curative intent treatment for non-small cell lung cancer: A comparison with age and gender-matched healthy controls. Respir. Med. 2015, 109, 1332–1339. [Google Scholar] [CrossRef] [Green Version]
- Hoeper, M.M.; Humbert, M.; Souza, R.; Idrees, M.; Kawut, S.M.; Aliwa-Hahnle, K.; Jing, Z.; Gibbs, J.S. A global view of pulmonary hypertension. Lancet Respir. Med. 2016, 4, 306–322. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 30 June 2021).
- Parasher, A. COVID-19: Current understanding of its pathophysiology, clinical presentation and treatment. Postgrad. Med. J. 2021, 97, 312–320. [Google Scholar] [CrossRef]
- Duval, K.; Grover, H.; Han, L.H.; Mou, Y.; Pegoraro, A.F.; Fredberg, J.; Chen, Z. Modeling Physiological Events in 2D vs.3D Cell Culture. Physiology 2017, 32, 266–277. [Google Scholar] [CrossRef]
- Asmani, M.; Velumani, S.; Li, Y.; Wawrzyniak, N.; Hsia, I.; Chen, Z.; Hinz, B.; Zhao, R. Fibrotic microtissue array to predict anti-fibrosis drug efficacy. Nat. Commun. 2018, 9, 2066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bluhmki, T.; Bitzer, S.; Gindele, J.A.; Schruf, E.; Kiechle, T.; Webster, M.; Schymeinsky, J.; Ries, R.; Gantner, F.; Bischoff, D.; et al. Development of a miniaturized 96-Transwell air-liquid interface human small airway epithelial model. Sci. Rep. 2020, 10, 13022. [Google Scholar] [CrossRef]
- Rayner, R.E.; Makena, P.; Prasad, G.L.; Cormet-Boyaka, E. Optimization of Normal Human Bronchial Epithelial (NHBE) Cell 3D Cultures for in vitro Lung Model Studies. Sci. Rep. 2019, 9, 500. [Google Scholar] [CrossRef] [PubMed]
- Katsura, H.; Sontake, V.; Tata, A.; Kobayashi, Y.; Edwards, C.E.; Heaton, B.E.; Konkimalla, A.; Asakura, T.; Mikami, Y.; Fritch, E.J.; et al. Human Lung Stem Cell-Based Alveolospheres Provide Insights into SARS-CoV-2-Mediated Interferon Responses and Pneumocyte Dysfunction. Cell Stem Cell 2020, 27, 890–904.e8. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.R.; Abdullatif, M.B.; Burnett, E.C.; Kempsell, K.E.; Conforti, F.; Tolley, H.; Collins, J.E.; Davies, D.E. Long Term Culture of the A549 Cancer Cell Line Promotes Multilamellar Body Formation and Differentiation towards an Alveolar Type II Pneumocyte Phenotype. PLoS ONE 2016, 11, e0164438. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Jamal, M.; Guo, P.; Jin, Z.; Zheng, F.; Song, X.; Zhan, J.; Wu, H. Irisin alleviates pulmonary epithelial barrier dysfunction in sepsis-induced acute lung injury via activation of AMPK/SIRT1 pathways. Biomed. Pharmacother. 2019, 118, 109363. [Google Scholar] [CrossRef]
- Del Mar Cendra, M.; Torrents, E. Differential adaptability between reference strains and clinical isolates of Pseudomonas aeruginosa into the lung epithelium intracellular lifestyle. Virulence 2020, 11, 862–876. [Google Scholar] [CrossRef]
- Selo, M.A.; Delmas, A.S.; Springer, L.; Zoufal, V.; Sake, J.A.; Clerkin, C.G.; Huwer, H.; Schneider-Daum, N.; Lehr, C.M.; Nickel, S.; et al. Tobacco Smoke and Inhaled Drugs Alter Expression and Activity of Multidrug Resistance-Associated Protein-1 (MRP1) in Human Distal Lung Epithelial Cells in vitro. Front. Bioeng. Biotechnol. 2020, 8, 1030. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, Y.; Xu, M.; Zhang, H.; Chen, Y.; Chung, K.F.; Adcock, I.M.; Li, F. Roles of TRPA1 and TRPV1 in cigarette smoke-induced airway epithelial cell injury model. Free Radic. Biol. Med. 2019, 134, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Tajima, H.; Tajiki-Nishino, R.; Watanabe, Y.; Kurata, K.; Fukuyama, T. Activation of aryl hydrocarbon receptor by benzo[a]pyrene increases interleukin 33 expression and eosinophil infiltration in a mouse model of allergic airway inflammation. J. Appl. Toxicol. 2020, 40, 1545–1553. [Google Scholar] [CrossRef]
- Pooladanda, V.; Thatikonda, S.; Bale, S.; Pattnaik, B.; Sigalapalli, D.K.; Bathini, N.B.; Singh, S.B.; Godugu, C. Nimbolide protects against endotoxin-induced acute respiratory distress syndrome by inhibiting TNF-α mediated NF-κB and HDAC-3 nuclear translocation. Cell Death Dis. 2019, 10, 81. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, W.L.; Yang, J.Y.; Huang, J.; Kang, G.; Hu, H.B.; Xie, S. Downregulation of lysyl oxidase-like 4 LOXL4 by miR-135a-5p promotes lung cancer progression in vitro and in vivo. J. Cell Physiol. 2019, 234, 18679–18687. [Google Scholar] [CrossRef]
- Nichane, M.; Javed, A.; Sivakamasundari, V.; Ganesan, M.; Ang, L.T.; Kraus, P.; Lufkin, T.; Loh, K.M.; Lim, B. Isolation and 3D expansion of multipotent Sox9(+) mouse lung progenitors. Nat. Methods 2017, 14, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Jacob, A.; Morley, M.; Hawkins, F.; McCauley, K.B.; Jean, J.C.; Heins, H.; Na, C.L.; Weaver, T.E.; Vedaie, M.; Hurley, K.; et al. Differentiation of Human Pluripotent Stem Cells into Functional Lung Alveolar Epithelial Cells. Cell Stem Cell 2017, 21, 472–488.e10. [Google Scholar] [CrossRef]
- Hawkins, F.; Kotton, D.N. Embryonic and induced pluripotent stem cells for lung regeneration. Ann. Am. Thorac. Soc. 2015, 1, S50–S53. [Google Scholar] [CrossRef] [PubMed]
- Humayun, M.; Chow, C.W.; Young, E.W. Microfluidic lung airway-on-a-chip with arrayable suspended gels for studying epithelial and smooth muscle cell interactions. Lab Chip 2018, 18, 1298–1309. [Google Scholar] [CrossRef]
- Zhang, M.; Xu, C.; Jiang, L.; Qin, J. A 3D human lung-on-a-chip model for nanotoxicity testing. Toxicol. Res. 2018, 7, 1048–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, J.B.; Luks, A.M. West’s Respiratory Physiology: The Essentials, 10th ed.; Wolters Kluwer: Philadelphia, PA, USA, 2016. [Google Scholar]
- Hsia, C.C.; Ravikumar, P. Role of Mechanical Stress in Lung Repair and Regeneration; Stem Cells in the Lung, Stem Cell Biology and Regenerative Medicine; Bertoncello, I., Ed.; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar] [CrossRef]
- Humphrey, J.D.; Dufresne, E.R.; Schwartz, M.A. Mechanotransduction and extracellular matrix homeostasis. Nat. Rev. Mol. Cell Biol. 2014, 15, 802–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, B. Biology of the extracellular matrix: An overview. J. Glaucoma. 2014, 23, S20–S23. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Takai, K.; Weaver, V.M.; Werb, Z. Extracellular matrix degradation and remodeling in development and disease. Cold Spring Harb. Perspect. Biol. 2011, 3, a005058. [Google Scholar] [CrossRef] [PubMed]
- Baptista, P.M.; Orlando, G.; Mirmalek-Sani, S.H.; Siddiqui, M.; Atala, A.; Soker, S. Whole organ decellularization–a tool for bioscaffold fabrication and organ bioengineering. In Proceedings of the 2009 Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Minneapolis, Minnesota, 3–6 September 2009; pp. 6526–6529. [Google Scholar] [CrossRef]
- Ott, H.C.; Clippinger, B.; Conrad, C.; Schuetz, C.; Pomerantseva, I.; Ikonomou, L.; Kotton, D.; Vacanti, J.P. Regeneration and orthotopic transplantation of a bioartificial lung. Nat. Med. 2020, 16, 927–933. [Google Scholar] [CrossRef]
- Wagner, D.E.; Bonenfant, N.R.; Sokocevic, D.; DeSarno, M.J.; Borg, Z.D.; Parsons, C.S.; Brooks, E.M.; Platz, J.J.; Khalpey, Z.I.; Hoganson, D.M.; et al. Three-dimensional scaffolds of acellular human and porcine lungs for high throughput studies of lung disease and regeneration. Biomaterials 2014, 35, 2664–2679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo, E.; Kasper, J.Y.; Unger, R.E.; Farré, R.; Kirkpatrick, C.J. Development of a bronchial wall model: Triple culture on a decellularized porcine trachea. Tissue Eng. Part C Methods 2015, 21, 909–921. [Google Scholar] [CrossRef]
- Nonaka, P.N.; Uriarte, J.J.; Campillo, N.; Melo, E.; Navajas, D.; Farré, R.; Oliveira, L.V. Mechanical properties of mouse lungs along organ decellularization by sodium dodecyl sulfate. Respir. Physiol. Neurobiol. 2014, 200, 1–5. [Google Scholar] [CrossRef]
- Jensen, T.; Roszell, B.; Zang, F.; Girard, E.; Matson, A.; Thrall, R.; Jaworski, D.M.; Hatton, C.; Weiss, D.J.; Finck, C. A rapid lung de-cellularization protocol supports embryonic stem cell differentiation in vitro and following implantation. Tissue Eng. Part C Methods 2012, 18, 632–646. [Google Scholar] [CrossRef] [Green Version]
- Gilpin, S.E.; Ren, X.; Okamoto, T.; Guyette, J.P.; Mou, H.; Rajagopal, J.; Mathisen, D.J.; Vacanti, J.P.; Ott, H.C. Enhanced lung epithelial specification of human induced pluripotent stem cells on decellularized lung matrix. Ann. Thorac. Surg. 2014, 98, 1721–1729. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Moser, P.T.; Gilpin, S.E.; Okamoto, T.; Wu, T.; Tapias, L.F.; Mercier, F.E.; Linjie, X.; Ghawi, R.; Scadden, D.T.; et al. Engineering pulmonary vasculature in decellularized rat and human lungs. Nat. Biotechnol. 2015, 33, 1097–1102. [Google Scholar] [CrossRef]
- Barkauskas, C.E.; Chung, M.; Fiortet, B.; Gao, X.; Katsura, H.; Hogan, B.L. Lung organoids: Current uses and future promise. Development 2017, 144, 986–997. [Google Scholar] [CrossRef] [Green Version]
- Tas, S.; Bölükbas, D.A.; Alsafadi, H.N.; Da Silva, I.A.; De Santis, M.M.; Rehnberg, E.; Tamargo, I.; Mohlin, S.; Lindstedt, S.; Wagner, D.E. Decellularized extracellular matrix hydrogels for human airway organoid culture. ERJ Open Res. 2021, 7, 101. [Google Scholar] [CrossRef]
- Skardal, A.; Murphy, S.V.; Devarasetty, M.; Mead, I.; Kang, H.W.; Seol, Y.J.; Zhang, Y.S.; Shin, S.R.; Zhao, L.; Aleman, J.; et al. Multi-tissue interactions in an integrated three-tissue organ-on-a-chip platform. Sci. Rep. 2017, 7, 8837. [Google Scholar] [CrossRef]
- Bishop, E.S.; Mostafa, S.; Pakvasa, M.; Luu, H.H.; Lee, M.J.; Wolf, J.M.; Ameer, G.A.; He, T.C.; Reid, R.R. 3-D bioprinting technologies in tissue engineering and regenerative medicine: Current and future trends. Genes Dis. 2017, 4, 185–195. [Google Scholar] [CrossRef]
- Park, J.Y.; Ryu, H.; Lee, B.; Ha, D.H.; Ahn, M.; Kim, S.; Kim, J.Y.; Jeon, N.L.; Cho, D.W. Development of a functional airway-on-a-chip by 3D cell printing. Biofabrication 2018, 11, 015002. [Google Scholar] [CrossRef] [PubMed]
- Rehmani, S.S.; Al-Ayoubi, A.M.; Ayub, A.; Barsky, M.; Lewis, E.; Flores, R.; Lebovics, R.; Bhora, F.Y. Three-dimensional-printed bioengineered tracheal grafts: Preclinical results and potential for human use. Ann. Thorac. Surg. 2017, 104, 998–1004. [Google Scholar] [CrossRef] [Green Version]
- Kang, D.; Park, J.A.; Kim, W.; Kim, S.; Lee, H.R.; Kim, W.J.; Yoo, J.Y.; Jung, S. All-inject-printed 3D alveolar barrier model with physiologically relevant microarchitecture. Adv. Sci. 2021, 8, 2004990. [Google Scholar] [CrossRef]
- Grigoryan, B.; Paulsen, S.J.; Corbett, D.C.; Sazer, D.W.; Fortin, C.L.; Zaita, A.J.; Greenfield, P.T.; Calafat, N.J.; Gounley, J.P.; Ta, A.H.; et al. Multivascular networks and functional intravascular topologies within biocompatible hydrogels. Science 2019, 364, 458–464. [Google Scholar] [CrossRef]
- Burgstaller, G.; Oehrle, B.; Gerckens, M.; White, E.S.; Schiller, H.B.; Eickelberg, O. The instructive extracellular matrix of the lung: Basic composition and alterations in chronic lung disease. Eur. Respir. J. 2017, 50, 1601805. [Google Scholar] [CrossRef] [Green Version]
- De Santis, M.M.; Alsafadi, H.N.; Tas, S.; Bölükbas, D.A.; Prithiviraj, S.; Da Silva, I.A.N.; Mittendorfer, M.; Ota, C.; Stegmayr, J.; Daoud, F.; et al. Extracellular-matrix-reinforced bioinks for 3D bioprinting human tissue. Adv. Mater. 2021, 33, 2005476. [Google Scholar] [CrossRef]
- Petrou, C.L.; D’ovidio, T.J.; Bölükbas, D.A.; Tas, S.; Brown, R.D.; Allawzi, A.; Lindstedt, S.; Nozik-Grayck, E.; Stenmark, K.R.; Wagner, D.E.; et al. Clickable decellularized extracellular matrix as a new tool for building hybrid-hydrogels to model chronic fibrotic diseases in vitro. J. Mater. Chem. B 2020, 8, 6814–6826. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Liu, T.; Liao, J.; Maharjan, S.; Xie, X.; Pérez, M.; Anaya, I.; Wang, S.; Mayer, A.T.; Kang, Z.; et al. Reversed-engineered human alveolar lung-on-a-chip model. Proc. Natl. Acad. Sci. USA 2021, 118, e2016146118. [Google Scholar] [CrossRef]
- Bessler, N.; Ogiermann, D.; Buchholz, M.B.; Santel, A.; Heidenreich, J.; Ahmmed, R.; Zaehres, H.; Brand-Saberi, B. Nydus One Syringe Extruder (NOSE): A Prusa i3 3D printer conversion for bioprinting applications utilizing the FRESH-method. HardwareX 2019, 6, e00069. [Google Scholar] [CrossRef]
- Low, L.A.; Mummery, C.; Berridge, B.R.; Austin, C.P.; Tagle, D.A. Organs-on-chips: Into the next decade. Nat. Rev. Drug Discov. 2021, 20, 345–361. [Google Scholar] [CrossRef]
- Huh, D.; Matthews, B.D.; Mammoto, A.; Montoya-Zavala, M.; Hsin, H.Y.; Ingber, D.E. Reconstituting organ-level lung functions on a chip. Science 2010, 328, 1662–1668. [Google Scholar] [CrossRef] [Green Version]
- Huh, D.; Leslie, D.C.; Matthews, B.D.; Fraser, J.P.; Jurek, S.; Hamilton, G.A.; Thorneloe, K.S.; McAlexander, M.A.; Ingber, D.E. A human disease model of drug toxicity-induced pulmonary edema in a lung-on-a-chip microdevice. Sci. Transl. Med. 2012, 4, 159ra147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stucki, A.O.; Stucki, J.D.; Hall, S.R.; Felder, M.; Mermoud, Y.; Schmid, R.A.; Geiser, T.; Guenat, O.T. A lung-on-a-chip array with an integrated bio-inspired respiration mechanism. Lab Chip 2015, 15, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- Jorba, I.; Beltrán, G.; Falcones, B.; Suki, B.; Farré, B.; García-Aznar, J.M.; Navajas, D. Nonlinear elasticity of the lung extracellular microenvironment is regulated by macroscale tissue strain. Acta Biomater. 2019, 92, 265–276. [Google Scholar] [CrossRef]
- Zamprogno, P.; Thoma, G.; Cencen, V.; Ferrari, D.; Putz, B.; Michler, J.; Fantner, G.E.; Guenat, O.T. Mechanical Properties of Soft Biological Membranes for Organ-on-a-Chip Assessed by Bulge Test and AFM. ACS Biomater. Sci. Eng. 2021, 7, 2990–2997. [Google Scholar] [CrossRef]
- Sellgren, K.L.; Butala, E.J.; Gilmour, B.P.; Randell, S.H.; Grego, S. A biomimetic multicellular model of the airways using primary human cells. Lab Chip 2014, 14, 3349–3358. [Google Scholar] [CrossRef]
- Benam, K.H.; Villenave, R.; Lucchesi, C.; Varone, A.; Hubeau, C.; Lee, H.; Alves, S.E.; Salmon, M.; Ferrante, T.C.; Weaver, J.C.; et al. Small airway-on-a-chip enables analysis of human lung inflammation and drug responses in vitro. Nat. Methods 2016, 13, 151–157. [Google Scholar] [CrossRef]
- Nesmith, A.P.; Agarwal, A.; McCain, M.L.; Parker, K.K. Human airway musculature on a chip: An in vitro model of allergic asthmatic bronchoconstriction and bronchodilation. Lab Chip 2014, 14, 3925–3936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tas, S.; Renhberg, E.; Bölükbas, D.; Beech, J.; Kazado, L.N.; Sveningsson, I.; Arvidsson, M.; Sandberg, A.; Dahlgren, K.A.; Edthofer, A.; et al. 3D printed lung-on-a-chip device with a stretchable nanofibrous membrane for modeling ventilator induced lung injury. bioRxiv 2021. [Google Scholar] [CrossRef]
- Tavana, H.; Kuo, C.H.; Lee, Q.Y.; Mosadegh, B.; Huh, D.; Christensen, P.J.; Grotberg, J.B.; Takayama, S. Dynamics of liquid plugs of buffer and surfactant solutions in a microengineered pulmonary airway model. Langmuir 2010, 26, 3744–3752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonaka, P.N.; Falcones, B.; Farré, A.; Artigas, A.; Almendros, I.; Navajas, D. Biophysical preconditioning mesenchymal stem cells improves treatment of ventilator-induced lung injury. Arch. Bronconeumol. 2020, 56, 176–189. [Google Scholar] [CrossRef]
- Grist, S.M.; Chrostowski, L.; Cheung, K.C. Optical oxygen sensors for applications in microfluidic cell culture. Sensors 2010, 10, 9286–9316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campillo, N.; Jorba, I.; Schaedel, L.; Casals, B.; Gozal, D.; Farré, R.; Almendros, I.; Navajas, D. A novel chip for cyclic stretch and intermittent hypoxia cell exposures mimicking obstructive sleep apnea. Front. Physiol. 2016, 7, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campillo, N.; Torres, M.; Vilaseca, A.; Nonaka, P.N.; Gozal, D.; Roca-Ferrer, J.; Picado, C.; Montserrat, J.M.; Farré, R.; Navajas, D.; et al. Role of cyclooxygenase-2 on intermittent hypoxia-induced lung tumor malignancy in a mouse model of sleep apnea. Sci. Rep. 2017, 7, 44693. [Google Scholar] [CrossRef]
- Campillo, N.; Falcones, B.; Otero, J.; Colina, R.; Gozal, D.; Navajas, D.; Farré, R.; Almendros, I. Differential oxygenation in tumor microenvironment modulates macrophage and cancer cell crosstalk: Novel experimental setting and proof of concept. Front. Oncol. 2019, 9, 43. [Google Scholar] [CrossRef] [Green Version]
- Benam, K.H.; Novak, R.; Nawroth, J.; Hirano-Kobayashi, M.; Ferrante, T.C.; Choe, Y.; Prantil-Baun, R.; Weaver, J.C.; Bahinski, A.; Parker, K.K.; et al. Matched-Comparative Modeling of Normal and Diseased Human Airway Responses Using a Microengineered Breathing Lung Chip. Cell Syst. 2016, 3, 456–466.e4. [Google Scholar] [CrossRef]
- Yang, X.; Li, K.; Zhang, X.; Liu, C.; Guo, B.; Wen, W.; Gao, X. Nanofiber membrane supported lung-on-a-chip microdevice for anti-cancer drug testing. Lab Chip 2018, 18, 486–495. [Google Scholar] [CrossRef]
- Park, S.E.; Georgescu, A.; Huh, D. Organoids-on-a-chip. Science 2019, 364, 960–965. [Google Scholar] [CrossRef]
Figure 1.
Overview of the current strategies available for in-lab lung models.

Figure 2.
Fabrication of functional lung in vitro models. (A) Overall design and implementation strategy for the 3-tissue-representative organ-on-a-chip system using a variety of biofabrication approaches (organoids bioengineering, 3D bioprinting and microfluidic systems). Copyright, 2019, reproduced with permission from Reference [50]. (B) Schematic diagram of inkjet-printing process for the generation of a 3-layered alveolar barrier (from bottom to top: endothelial cells, collagen I and fibroblasts, and type I and II alveolar cells, representing the endothelium, the basement membrane and the epithelium, respectively). Copyright, 2021, reproduced with permission from Reference [54]. (C) Schematic of the lung-mimetic design (left) and the distal lung subunit (right) of hydrogels with vascularized alveolar model topologies. Copyright, 2019, reproduced with permission from Reference [55]. (D) 3D view showing the working principle of the alveolar lung-on-a-chip model with GelMa-based alveolar sac-like 3D structures. Copyright, 2021, reproduced with permission from Reference [59], CC BY-NC-ND 4.0. (E) Schematic depicting FRESH technology employed for the fabrication of small airways. Copyright, 2019, reproduced with permission from Reference [60].
Figure 2.
Fabrication of functional lung in vitro models. (A) Overall design and implementation strategy for the 3-tissue-representative organ-on-a-chip system using a variety of biofabrication approaches (organoids bioengineering, 3D bioprinting and microfluidic systems). Copyright, 2019, reproduced with permission from Reference [50]. (B) Schematic diagram of inkjet-printing process for the generation of a 3-layered alveolar barrier (from bottom to top: endothelial cells, collagen I and fibroblasts, and type I and II alveolar cells, representing the endothelium, the basement membrane and the epithelium, respectively). Copyright, 2021, reproduced with permission from Reference [54]. (C) Schematic of the lung-mimetic design (left) and the distal lung subunit (right) of hydrogels with vascularized alveolar model topologies. Copyright, 2019, reproduced with permission from Reference [55]. (D) 3D view showing the working principle of the alveolar lung-on-a-chip model with GelMa-based alveolar sac-like 3D structures. Copyright, 2021, reproduced with permission from Reference [59], CC BY-NC-ND 4.0. (E) Schematic depicting FRESH technology employed for the fabrication of small airways. Copyright, 2019, reproduced with permission from Reference [60].
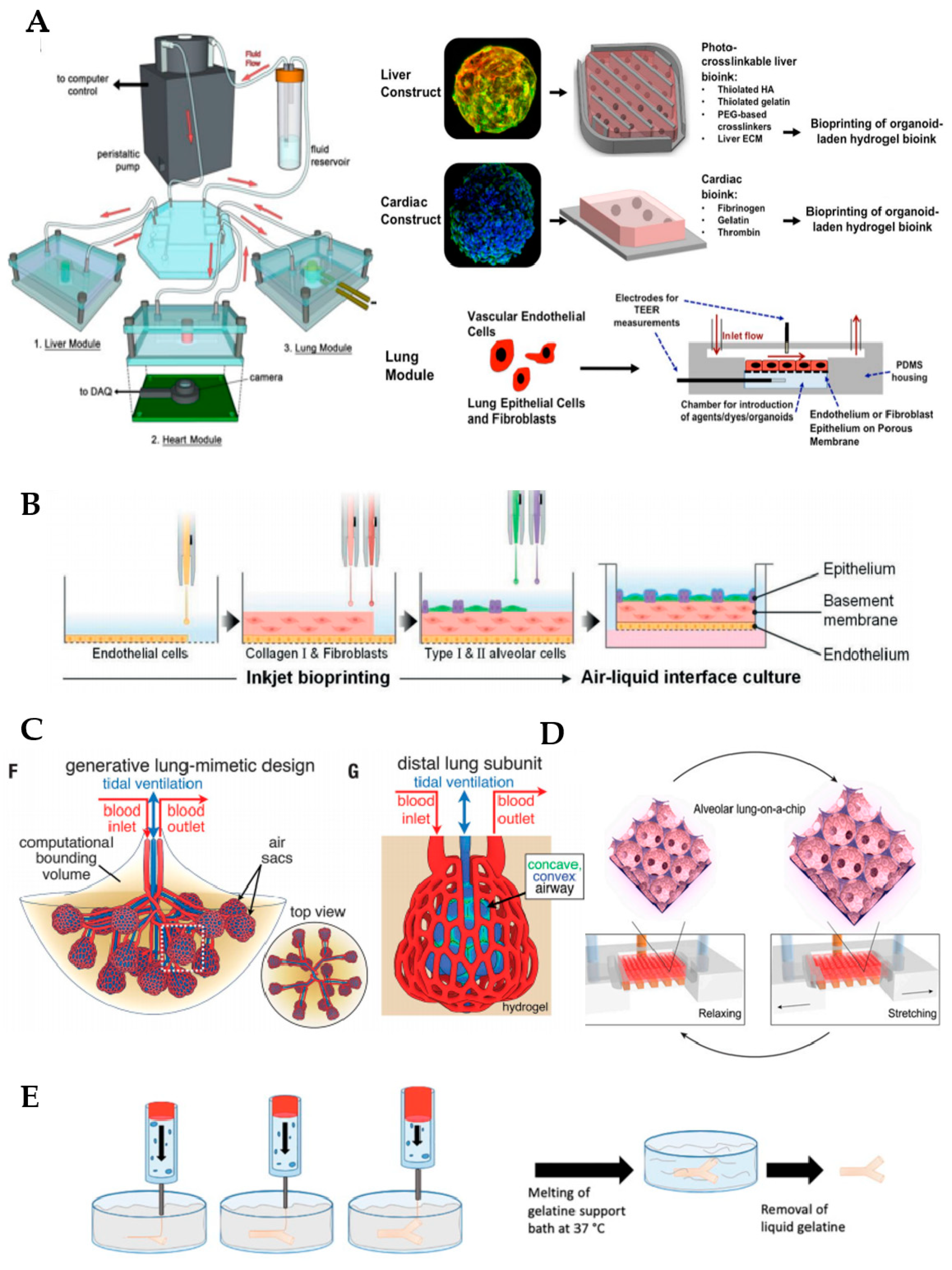
Figure 3.
Alveolar lung-on-chip platform.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Campillo, N.; Oliveira, V.R.; da Palma, R.K. Alveolus Lung-on-a-Chip Platform: A Proposal. Chemosensors 2021, 9, 248. https://doi.org/10.3390/chemosensors9090248
AMA Style
Campillo N, Oliveira VR, da Palma RK. Alveolus Lung-on-a-Chip Platform: A Proposal. Chemosensors. 2021; 9(9):248. https://doi.org/10.3390/chemosensors9090248
Chicago/Turabian StyleCampillo, Noelia, Vinicius Rosa Oliveira, and Renata Kelly da Palma. 2021. "Alveolus Lung-on-a-Chip Platform: A Proposal" Chemosensors 9, no. 9: 248. https://doi.org/10.3390/chemosensors9090248
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.