
Dynamics in a Predator–Prey Model with Cooperative Hunting and Allee Effect


Abstract
:1. Introduction
2. The Existence of Constant Steady States
3. Bifurcation and Global Dynamics of Kinetic System
3.1. Bifurcation and Global Dynamics of Kinetic System with Weak Cooperative Hunting
3.1.1. Stability of All Equilibria and Hopf Bifurcation at
- (i)
- is a stable node;
- (ii)
- is an unstable node if , and it is a saddle if ; and
- (iii)
- is a stable node if , and it is a saddle if .
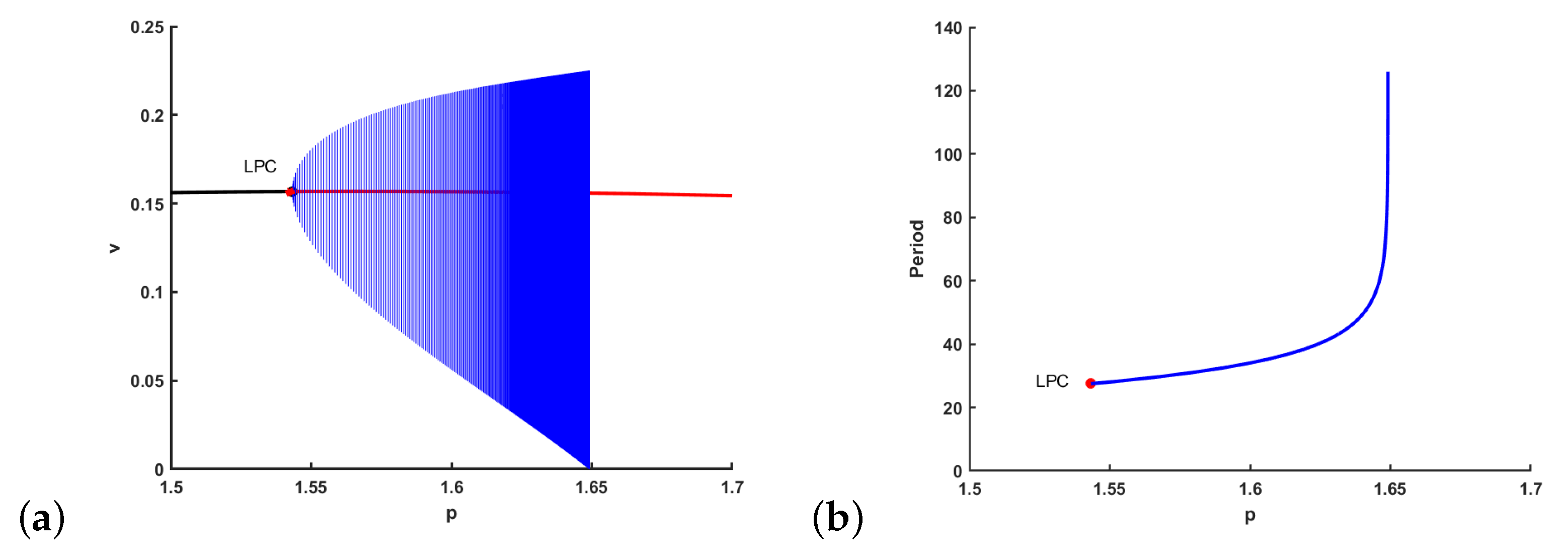
- (i)
- If , the bifurcating periodic solution is orbitally asymptotically stable, and it is bifurcating from as p increases and passes .
- (ii)
- If , the bifurcating periodic solution is unstable, and it is bifurcating from as p decreases and passes .
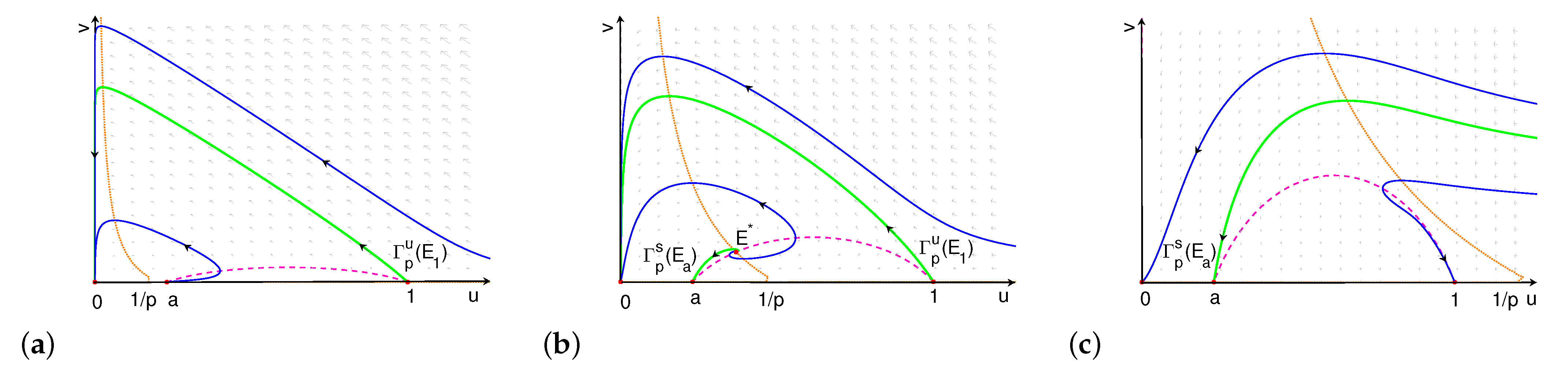
3.1.2. The Global Dynamics of Kinetic System with Weak Cooperative Hunting
- (i)
- If , is globally asymptotically stable (see Figure 3a).
- (ii)
- There is an such that if , connects to , and the extinction equilibrium is globally asymptotically stable (see Figure 3b).
- (iii)
- There is an such that if , then connects to . The orbits through any point above converge to , and the orbits through any point below converge to (see Figure 3c).
- (iv)
- If , the orbits through any point above converge to , and the orbits through any point below converge to (see Figure 3d).
- (i)
- If , connects to . The orbits through any point above converge to , and the orbits through any point below converge to .
- (ii)
- If , is a repellor, and there is a unique limit cycle under . The orbits through any point below converge to the limit cycle. (see Figure 4a,b).
- (iii)
- If , , there are two heteroclinic orbits forming a loop of heteroclinic orbits from to and back to (see Figure 4c). The orbits through any point exterior to the loop converge to , and the orbits through any point interior to the cycle converge to the loop.
- (iv)
- If , connects to , and the extinction equilibrium is globally asymptotically stable (see Figure 4d).
3.2. Bifurcation and Global Dynamics of Kinetic System with Strong Cooperative Hunting
3.2.1. Stability and Hopf Bifurcation of Interior Equilibria
3.2.2. The Global Dynamics of Kinetic System with Strong Cooperative Hunting
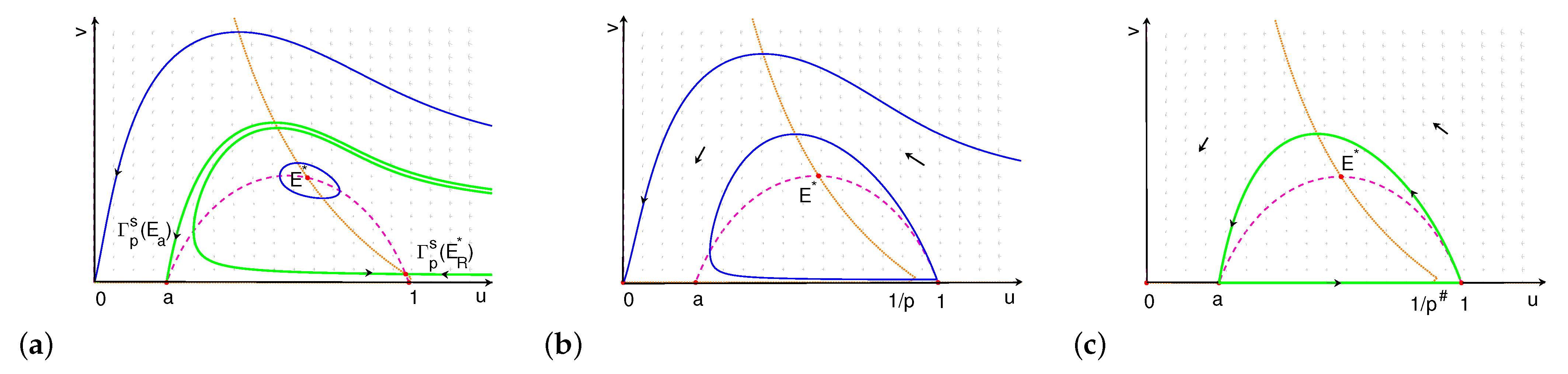
- (i)
- If , the upward connects to . The orbits through any point above converge to , and the orbits through any point inside the stable manifold converge to . The orbits through any point below and exterior to converge to (see Figure 7a).
- (ii)
- If , connects to . The orbits through any point above converge to , and the orbits through any point below converge to .
- (iii)
- When , is a repellor, and there is a unique limit cycle under . The orbits through any point below converge to the limit cycle (see Figure 7b,c).
- (iv)
- When , , and there are two heteroclinic orbits forming a loop of heteroclinic orbits between and (see Figure 7d). The orbits through any point exterior to the cycle converge to , and the orbits through any point interior to the cycle converge to the cycle.
- (v)
- If , connects to , and the extinction equilibrium is globally asymptotically stable (phase portrait is similar as Figure 6b).
- (i)
- If , the upward connects to , and the downward connects to . The orbits through any point above converge to , and the orbits through any point inside the stable manifold converge to . The orbits through any point below and exterior to converge to (phase portrait is similar as Figure 7a).
- (ii)
- When , there is a unique limit cycle inside the stable manifold of (see Figure 8a). The orbits through any point interior to converge to the limit cycle.
- (iii)
- When , there is a unique limit cycle under . The orbits through any point below converge to the limit cycle (see Figure 8b).
- (iv)
- When , , there are two heteroclinic orbits forming a loop of heteroclinic orbits from to and back to (see Figure 8c). The orbits through any point exterior to the cycle converge to , and the orbits through any point interior to the cycle converge to the cycle.
- (v)
- If , connects to , and the extinction equilibrium is globally asymptotically stable (phase portrait is similar as Figure 6b).
- (i)
- Loop of heteroclinic orbits. The downward branch of the unstable manifold of connects to , and the upward connects to , which collides with the the stable manifold of . The upward and downward unstable manifold of , together with the unstable manifold of on the axis, forms a loop of heteroclinic orbits among , and when (see Figure 9d).
- (ii)
- Homoclinic cycle. The upward unstable manifold and the left stable manifold of collide, which forms a homoclinic cycle. Denote the parameter p as (see Figure 9c).
- (iii)
- Limit cycle induced by Hopf bifurcation (see Figure 9a,b).
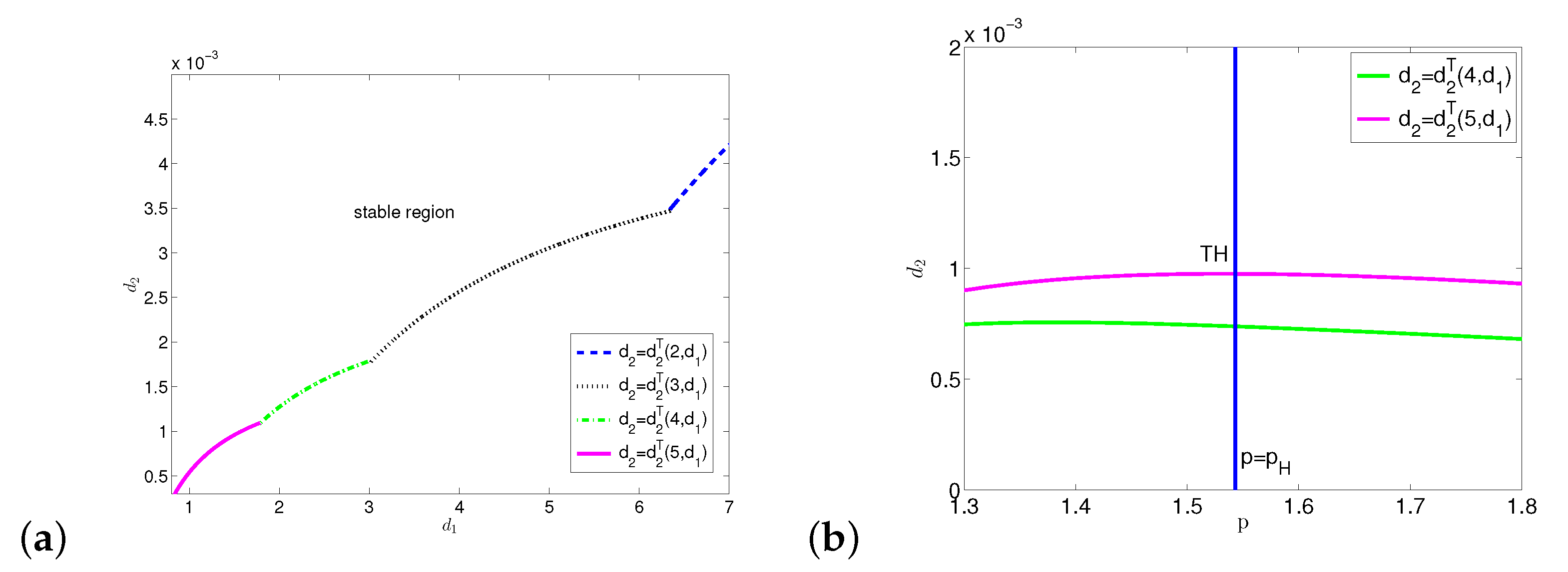
4. Diffusion-Driven Turing Instability and Turing–Hopf Bifurcation
5. The Dynamics of Diffusive System with Two Delays
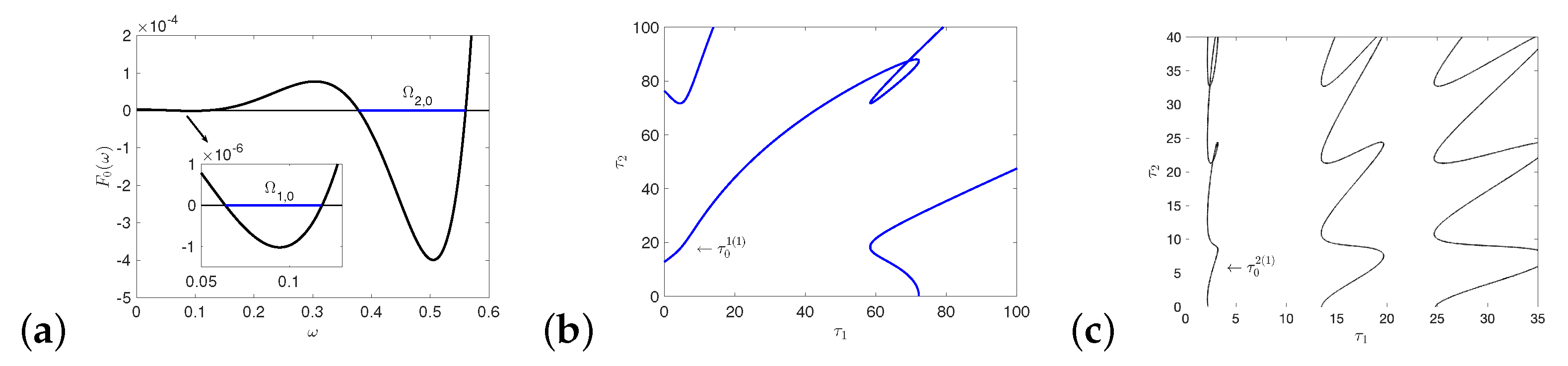
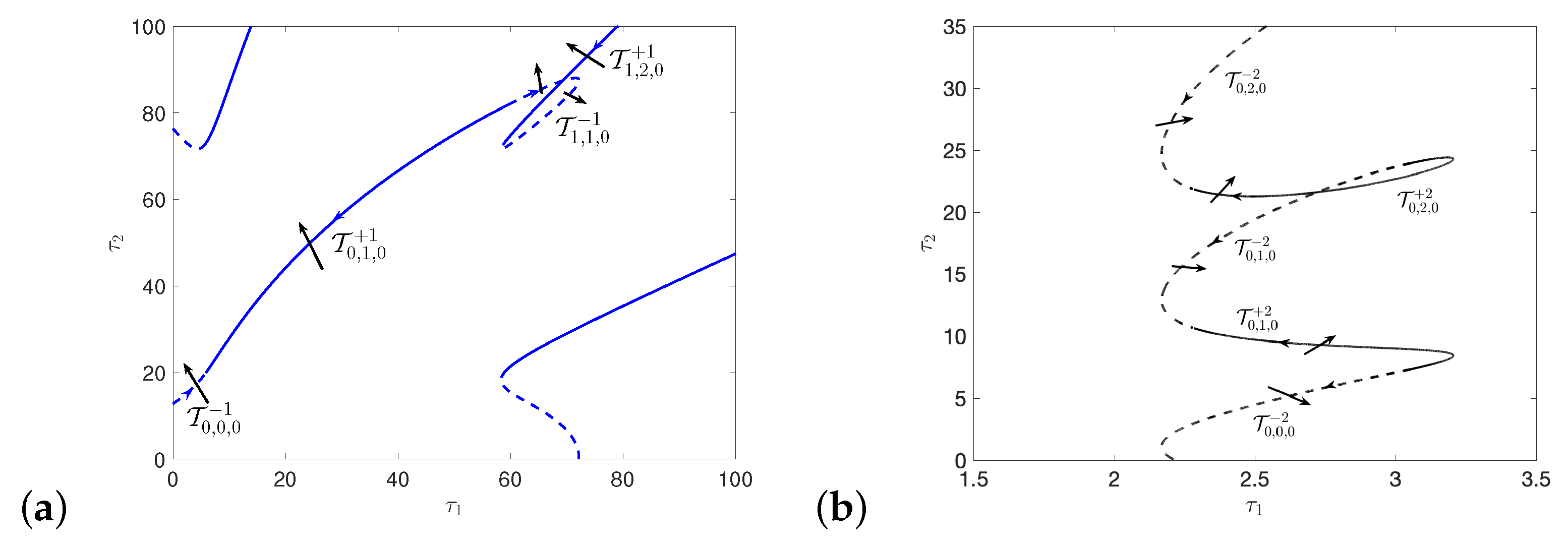
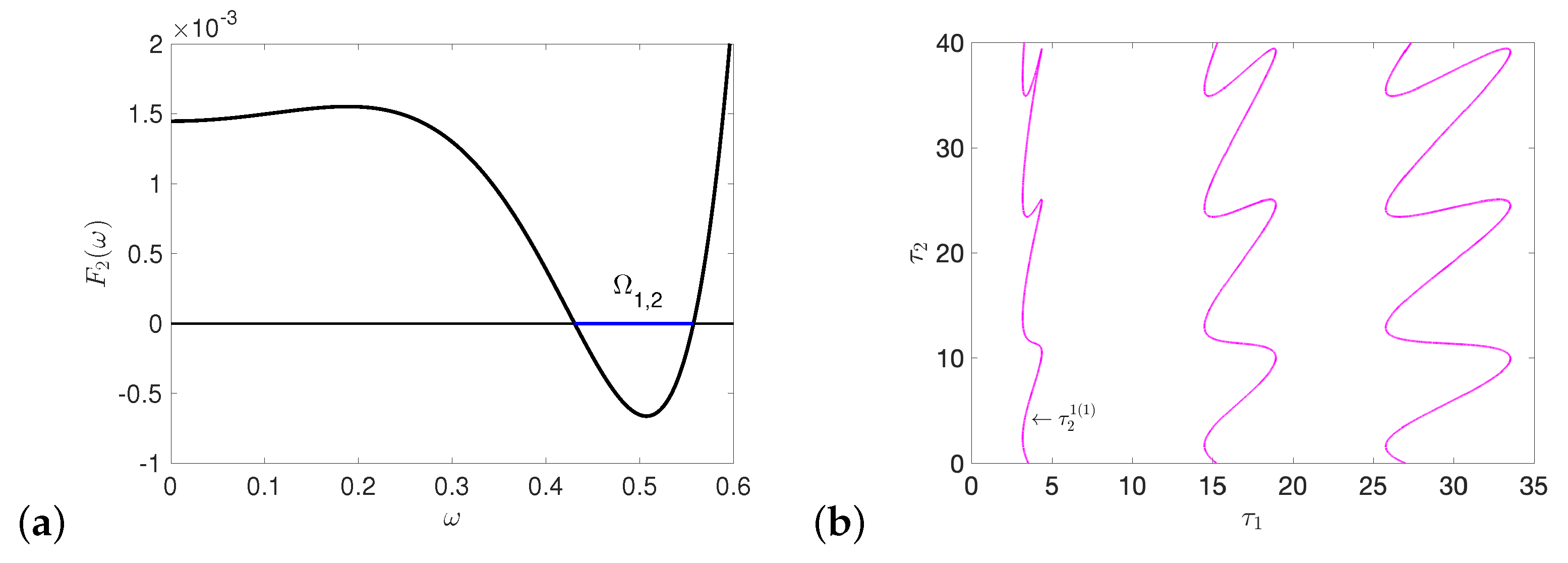
5.1. Hopf and Double Hopf Bifurcation Induced by Two Delays
5.2. Normal Form on the Center Manifold for Double Hopf Bifurcation
6. Numerical Simulations
6.1. Numerical Simulations for Kinetic System
6.1.1. Numerical Simulations for Kinetic System with Weak Cooperative Hunting
6.1.2. Numerical Simulations for Kinetic System with Strong Cooperative Hunting
6.2. Numerical Simulations for Diffusive System without Delays
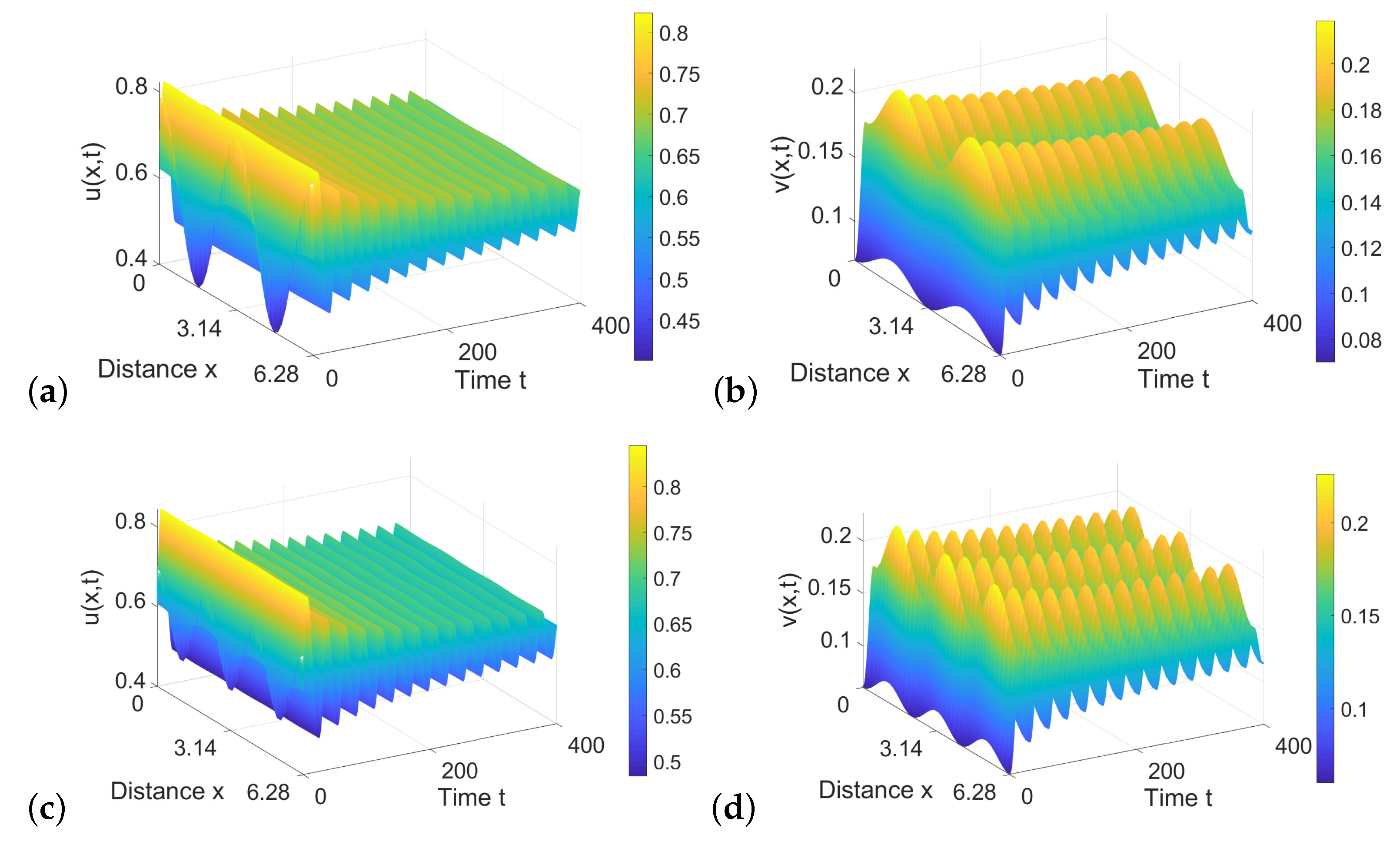
6.3. Numerical Simulations for Diffusive System with Two Delays
7. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. Some Results and Proofs for the Case of Weak Cooperation
Appendix A.1. The Properties of Stable and Unstable Manifold and the Existence or Nonexistence of Periodic Orbits
- (i)
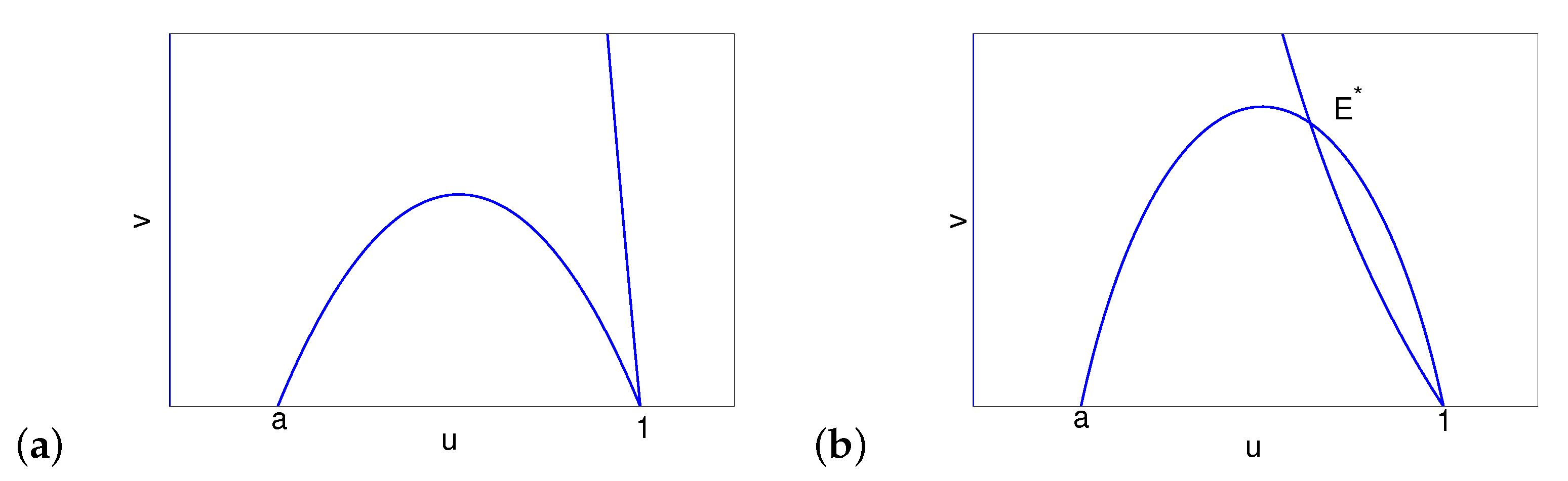
- The orbit meets the v-nullcline at a point , where , and is a monotone decreasing function for .
- (ii)
- The orbit meets the v-nullcline at a point , where , and is a monotone increasing function for .

- (i)
- If , then . Moreover, all orbits in the positive quadrant above converge to , and all orbits below have their limit sets in Ω, which is a positive invariant set.
- (ii)
- If , then , and tends to . Moreover, all orbits in the positive quadrant above converge to , and all orbits below have their limit sets in Ω, which is a negative invariant set.
- (i)
- If , then either is stable or Ω contains a periodic orbits which is stable from the outside (both may be true).
- (ii)
- If , then either is unstable or Ω contains a periodic orbits which is unstable from the outside (both may be true).
- (i)
- There is an such that if , there is no periodic orbits, and connects to , which is a heteroclinic orbit.
- (ii)
- There is an such that if , there is no periodic orbits, and connects to .
- (i)
- If , then
- (a)
- for , there is at least one periodic solution in Ω which is stable from the outside and one (perhaps the same) stable from inside;
- (b)
- if , then there is an such that if , there are at least two distinct periodic solutions, the inner of which is unstable while the outer is stable from the outside.
- (ii)
- If , then
- (a)
- for , there is at least one periodic solution in Ω which is unstable from the outside and one (not necessarily distinct) unstable from inside;
- (b)
- if , then there is an such that if , there are at least two distinct periodic solutions, the inner is stable while the outer is unstable from the outside.
Appendix A.2. The Proof of Theorem 3
Appendix A.3. Proof of Theorem 4
Appendix B. Some Results and Proofs for the Case of Strong Cooperation
- (i)
- The orbit meets the v-nullcline at a point , where , and is a monotone decreasing function for .
- (ii)
- The orbit meets the v-nullcline at a point , where , and is a monotone increasing function for .
- (i)
- If , there exists a unique , such that , forming a heteroclinic orbit from to ;
- (ii)
- If , for any .
- (i)
- Assume .
- (a)
- If , . All orbits in the positive quadrant above converge to . All orbits below have their limit sets in , which is a positive invariant set.
- (b)
- If , , and enters . All orbits in the positive quadrant above converge to . All orbits below have their limit sets in , which is a negative invariant set.
- (ii)
- Assume . for all , and enters . All orbits in the positive quadrant above converge to . All orbits below have their limit sets in , which is a negative invariant set.
- (i)
- The orbit of left meets the v-nullcline at a point , where .
- (ii)
- The orbit of upward meets the v-nullcline at a point , where .
- (ii)
- If . All orbits below the left have their limit sets in , which is a negative invariant set.
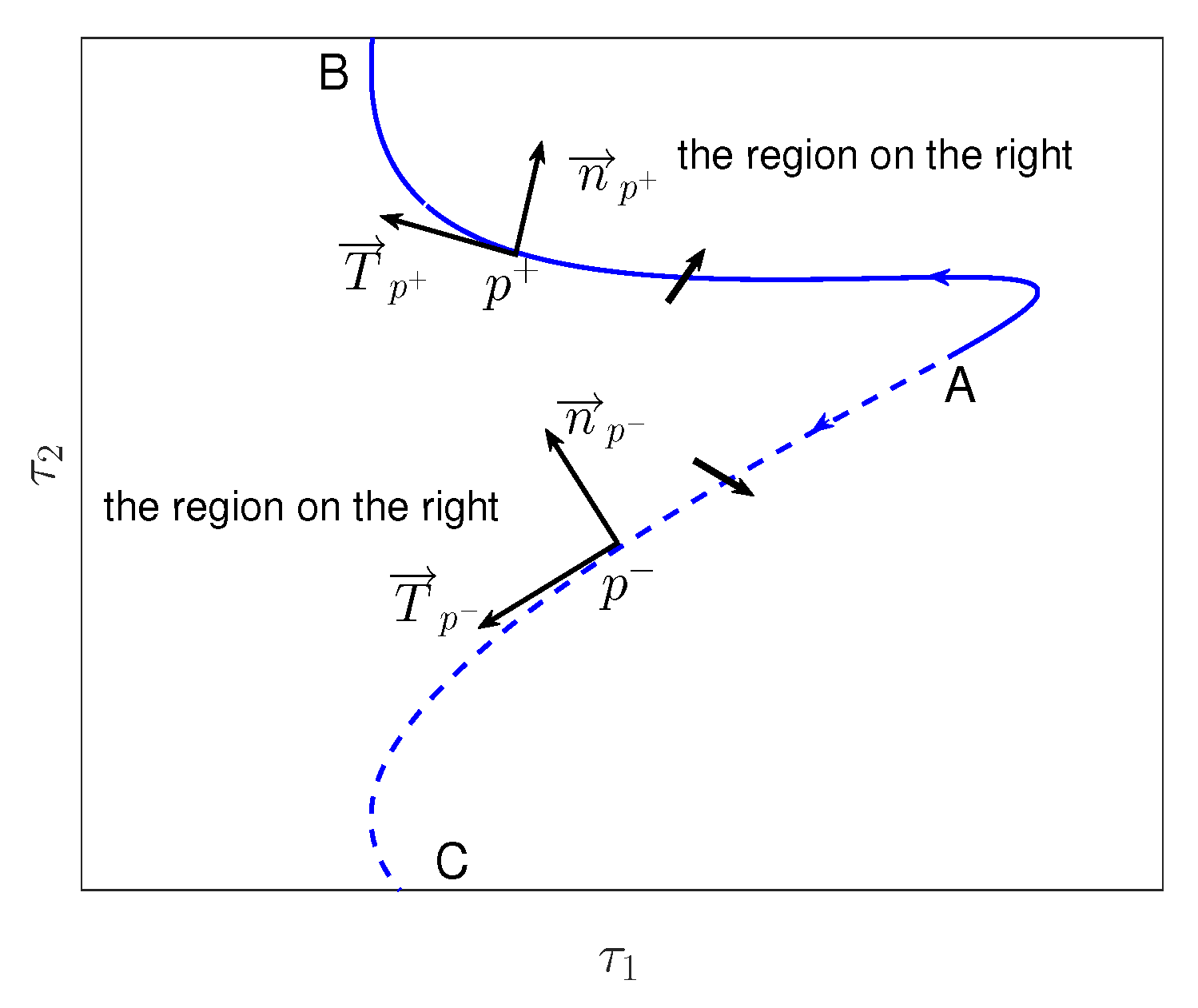
Appendix C. Stability Switching Curves
- (i)
- Finite number of characteristic roots on under the condition
- (ii)
- .
- (iii)
- are coprime polynomials.
- (iv)
- .
Appendix D. Crossing Directions
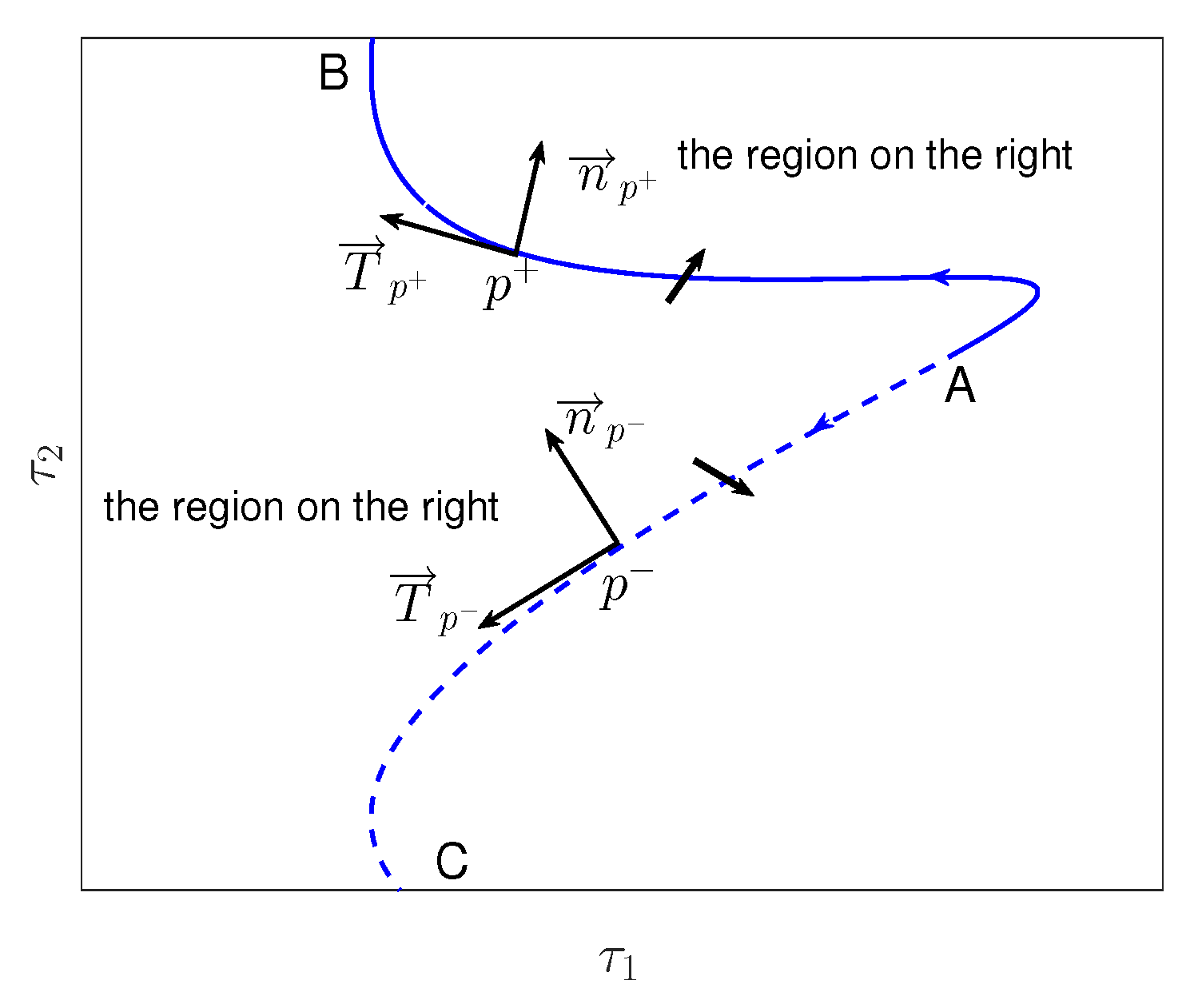
Appendix E. The Figures of Stability Switching Curves



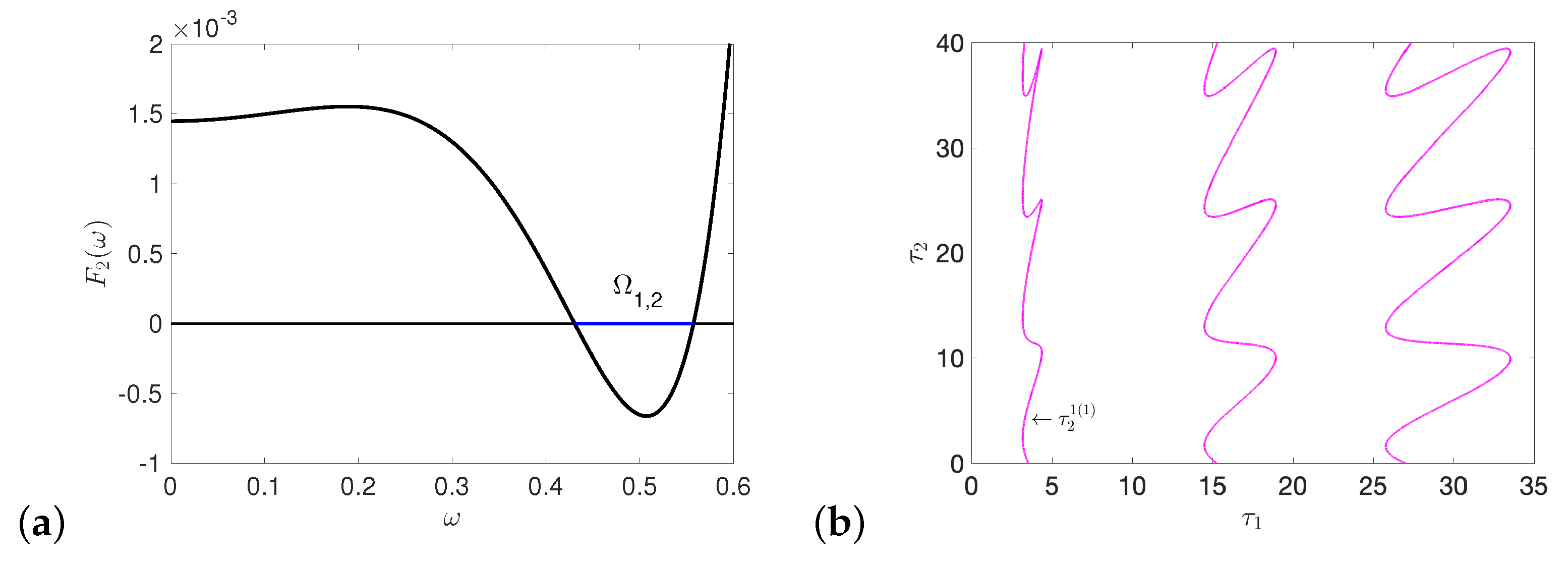
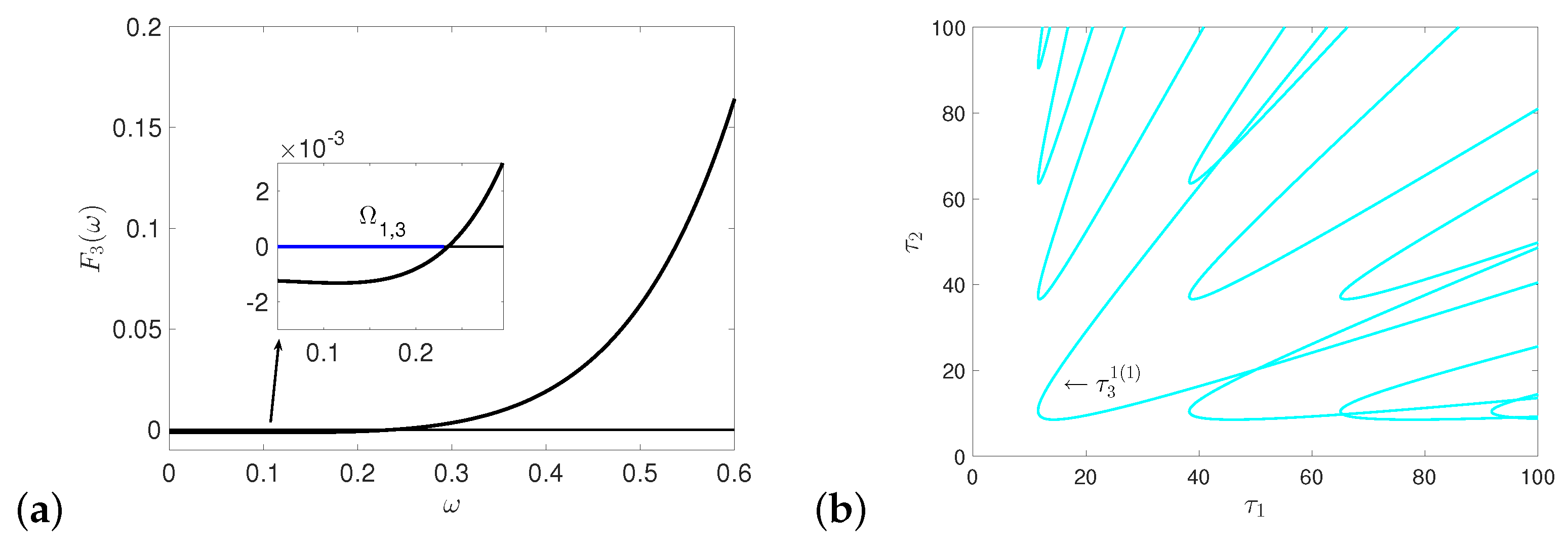
References
- Cosner, C.; DeAngelis, D.L.; Ault, J.S.; Olson, D.B. Effects of spatial grouping on the functional response of predators. Theor. Popul. Biol. 1999, 56, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Jeschke, J.M.; Kopp, M.; Tollrian, R. Predator functional responses: Discriminating between handling and digesting prey. Ecol. Monogr. 2002, 72, 95–112. [Google Scholar] [CrossRef]
- Holling, C.S. The components of predation as revealed by a study of small-mammal predation of the European pine sawfly. Can. Entomol. 1959, 91, 293–320. [Google Scholar] [CrossRef]
- Kazarinov, N.D.; Driessche, P.V.D. A model predator-prey system with functional response. Math. Biosci. 1978, 39, 125–134. [Google Scholar] [CrossRef]
- Turchin, P. Complex Population Dynamics: A Theoretical/Empirical Synthesis; Princeton University Press: Princeton, NJ, USA, 2003. [Google Scholar]
- Song, Y.; Peng, Y.; Zhang, T. The spatially inhomogeneous Hopf bifurcation induced by memory delay in a memory-based diffusion system. J. Differ. Equ. 2021, 300, 597–624. [Google Scholar] [CrossRef]
- Schmidt, P.A.; Mech, L.D. Wolf pack size and food acquisition. Am. Nat. 1997, 150, 513–517. [Google Scholar] [CrossRef]
- Courchamp, F.; Macdonald, D.W. Crucial importance of pack size in the African wild dog Lycaon pictus. Anim. Conserv. 2001, 4, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Scheel, D.; Packer, C. Group hunting behavioir of lions: A search for cooperation. Anim. Behav. 1991, 41, 697–709. [Google Scholar] [CrossRef]
- Boesch, C. Cooperative hunting in wild chimpanzees. Animal. Behav. 1994, 48, 653–667. [Google Scholar] [CrossRef] [Green Version]
- Bshary, R.; Hohner, A.; Ait-el-Djoudi, K.; Fricke, H. Interspecific communicative and coordinated hunting between groupers and giant moray eels in the Red Sea. PLoS Biol. 2006, 4, e431. [Google Scholar] [CrossRef]
- Uetz, G.W. Foraging stratigies of spiders. Trends Ecol. Evol. 1992, 7, 155–159. [Google Scholar] [CrossRef]
- Hector, D.P. Cooperative hunting and its relationship to foraging success and prey size in an avian predator. Ethology 1986, 73, 247–257. [Google Scholar] [CrossRef]
- Moffett, M.W. Foraging dynamics in the group-hunting myrmicine ant, pheidologeton diversus. J. Insect Behav. 1988, 1, 309–331. [Google Scholar] [CrossRef]
- Berec, L. Impacts of foraging facilitation among predators on predator-prey dynamics. Bull. Math. Biol. 2010, 72, 94–121. [Google Scholar] [CrossRef]
- Alves, M.T.; Hilker, F.M. Hunting cooperation and Allee effects in predators. J. Theor. Biol. 2017, 419, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.R.J.; Zhang, W.J.; Larriva, V. Cooperative hunting in a predator-prey system with Allee effects in the prey. Nat. Resour. Model. 2018, 31, e12194. [Google Scholar] [CrossRef]
- Pal, S.; Pal, N.; Samanta, S.; Chattopadhyay, J. Effect of hunting cooperation and fear in a predator-prey model. Ecol. Complex. 2019, 39, 100770. [Google Scholar] [CrossRef]
- Sen, D.; Ghorai, S.; Banerjee, S.M. Allee effect in prey versus hunting cooperation on predator-enhancement of stable coexistence. Int. J. Bifurc. Chaos 2019, 29, 1950081. [Google Scholar] [CrossRef]
- Yan, S.; Jia, D.; Zhang, T.; Yuan, S. Pattern dynamics in a diffusive predator-prey model with hunting cooperations. Chaos Solitons Fract. 2020, 130, 109428. [Google Scholar] [CrossRef]
- Wu, D.Y.; Zhao, M. Qualitative analysis for a diffusive predator-prey model with hunting cooperative. Phys. A 2019, 515, 299–309. [Google Scholar] [CrossRef]
- Song, D.X.; Li, C.; Song, Y.L. Stability and cross-diffusion-driven instability in a diffusive predator-prey system with hunting cooperation functional response. Nonlinear Anal. RWA 2020, 54, 103106. [Google Scholar] [CrossRef]
- Pal, S.; Pal, N.; Chattopadhyay, J. Hunting cooperation in a discrete-time predator-prey system. Int. J. Bifurc. Chaos 2018, 28, 1850083. [Google Scholar] [CrossRef]
- Pati, N.C.; Layek, G.C.; Pal, N. Bifurcations and organized structures in a predator-prey model with hunting cooperation. Chaos Soliton Fract. 2020, 140, 110184. [Google Scholar] [CrossRef]
- Chou, Y.H.; Chow, Y.S.; Hu, X.C.; Jang, S.R.J. A Ricker-type predator-prey system with hunting cooperation in discrete time. Math. Comput. Simulat. 2021, 190, 570–586. [Google Scholar] [CrossRef]
- Chow, Y.S.; Jang, S.R.J.; Wang, H.M. Cooperative hunting in a discrete predator–prey system. Int. J. Biomath. 2000, 13, 2050063. [Google Scholar] [CrossRef]
- Allee, W.C. The Social Life of Animals; William Heinemann: London, UK, 1938. [Google Scholar]
- Courchamp, F.; Berec, L.; Gascoigne, J. Allee Effects in Ecology and Conservation; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Wang, J.F.; Shi, J.P.; Wei, J.J. Predator-prey system with strong Allee effect in prey. J. Math. Biol. 2010, 62, 291–331. [Google Scholar] [CrossRef] [PubMed]
- Conway, E.D.; Smoller, J.A. Global analysis of a system of predator-prey equations. SIAM J. Appl. Math. 1986, 46, 630–642. [Google Scholar] [CrossRef]
- Song, Y.; Jiang, H.; Yuan, Y. Turing-Hopf bifurcation in the reaction-diffusion system with delay and application to a diffusive predator-prey model. J. Appl. Anal. Comput. 2019, 9, 1132–1164. [Google Scholar] [CrossRef]
- Xu, X.; Wei, J. Turing-Hopf bifurcation of a class of modified Leslie-Gower model with diffusion. Disc. Contin. Dyn. Sys. B 2018, 23, 765–783. [Google Scholar] [CrossRef] [Green Version]
- An, Q.; Jiang, W. Spatiotemporal attractors generated by the Turing-Hopf bifurcation in a time-delayed reaction-diffusion system. Disc. Contin. Dyn. Syst. B 2019, 24, 487–510. [Google Scholar] [CrossRef] [Green Version]
- Guo, G.; Liu, L.; Li, B.; Li, J. Qualitative analysis on positive steady-state solutions for an autocatalysis model with high order. Nonlinear Anal. RWA 2018, 41, 665–691. [Google Scholar] [CrossRef]
- Kuang, Y. Delay Differential Equations: With Applications in Population Dynamics; Academic Press: Boston, MA, USA, 1993. [Google Scholar]
- Gopalsamy, K. Stability and Oscillations in Delay Differential Equations of Population Dynamics; Springer: Dordrecht, The Netherlands, 1992. [Google Scholar]
- Martin, A.; Ruan, S.G. Predator-prey models with delay and prey harvesting. J. Math. Biol. 2001, 43, 247–267. [Google Scholar] [CrossRef]
- Xia, J.; Liu, Z. H; Yuan, R. The effects of harvesting and time delay on predator-prey systems with Holling type II functional response. SIAM J. Appl. Math. 2009, 70, 1178–1200. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.P.; Wang, C.C.; Wang, H. Diffusive spatial movement with memory and maturation delays. Nonlinearity 2019, 32, 3188–3208. [Google Scholar] [CrossRef] [Green Version]
- Wright, E. A functional equation in the heuristic theory of primes. Math. Gaze. 1961, 45, 15–16. [Google Scholar] [CrossRef]
- May, R.M. Time delay versus stability in population models with two and three trophic levels. Ecology 1973, 54, 315–325. [Google Scholar] [CrossRef]
- Xu, C.; Tang, X.; Liao, M.; He, X. Bifurcation analysis in a delayed Lokta-Volterra predator-prey model with two delays. Nonlinear Dyn. 2011, 66, 169–183. [Google Scholar] [CrossRef]
- Yan, X.P.; Zhang, C.H. Hopf bifurcation in a delayed Lotka-Volterra predator-prey system. Nonlinear Anal. RWA 2008, 9, 114–127. [Google Scholar] [CrossRef]
- Jankovic, M.; Petrovskii, S. Are time delay always destabilizing? Revisiting the role of time delays and the Allee effect. Theor. Ecol. 2014, 7, 335–349. [Google Scholar] [CrossRef] [Green Version]
- Cooke, K.L.; Driessche, P.V.D. Analysis of an SEIRS epidemic model with two delays. J. Math. Biol. 1996, 35, 240–260. [Google Scholar] [CrossRef]
- Freedman, H.I.; Rao, V.S.H. Stability criteria for a system involving two time delays. SIAM J. Appl. Math. 1986, 46, 552–560. [Google Scholar] [CrossRef]
- Wei, J. J, Ruan, S.G. Stability and bifurcation in a neural network model with two delays. Physica D 1999, 130, 255–272. [Google Scholar] [CrossRef]
- Xu, J.; Chung, K.W.; Chan, C.L. An efficient method for studying weak resonant double Hopf bifurcation in nonlinear systems with delayed feedbacks. SIAM J. Appl. Dyn. Syst. 2007, 6, 29–60. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.F.; Niu, B.; Wei, J.J. Two delays induce Hopf bifurcation and double Hopf bifurcation in a diffusive Leslie-Gower predator-prey system. Chaos 2019, 29, 013101. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.X.; Yang, Z.C.; Yi, T.S. On the basins of attraction for a class of delay differential equations with non-monotone bistable nonlinearities. J. Differ. Equ. 2014, 256, 2101–2114. [Google Scholar] [CrossRef]
- Huang, C.X.; Zhao, X.; Cao, J.D.; Alsaadi, F.E. Global dynamics of neoclassical growth model with multiple pairs of variable delays. Nonlinearity 2020, 33, 6819–6834. [Google Scholar] [CrossRef]
- Yu, P.; Yuan, Y.; Xu, J. Study of double Hopf bifurcation and chaos for oscillator with time delay feedback. Commun. Nonlinear Sci. Numer. Simul. 2002, 7, 69–91. [Google Scholar] [CrossRef]
- Du, Y.F.; Niu, B.; Guo, Y.X.; Wei, J.J. Double Hopf bifurcation in delayed reaction-diffusion systems. J. Dynam. Differ. Equ. 2020, 32, 313–358. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Lu, Q.; Feng, Z. Double Hopf bifurcation for van der Pol-Duffing oscillator with parametric delay feedback control. J. Math. Anal. Appl. 2008, 338, 993–1007. [Google Scholar] [CrossRef] [Green Version]
- Campell, S.A.; Bélair, J.; Ohira, T.; Milton, J. Limit cycles, tori, and complex dynamics in a second-order differential equations with delayed negative feedback. J. Dynam. Differ. Equ. 1995, 7, 213–236. [Google Scholar] [CrossRef]
- Acosta-Humánez1, P.B.; Yagasaki, K. Nonintegrability of the unfoldings of codimension-two bifurcations. Nonlinearity 2020, 33, 1366–1387. [Google Scholar] [CrossRef] [Green Version]
- Ge, J.H.; Xu, J. An analytical method for studying double Hopf bifurcations induced by two delays in nonlinear differential systems. Sci. China-Technol. Sci. 2019, 63, 597–602. [Google Scholar] [CrossRef]
- Rosenzweig, M.L.; MacArthur, R. Graphical representation and stability conditions of predator-prey interactions. Am. Nat. 1963, 97, 209–223. [Google Scholar] [CrossRef]
- Rosenzweig, M.L. Why the prey curve has a hump? Am. Nat. 1969, 103, 81–87. [Google Scholar] [CrossRef]
- Hsu, S.B. On global stability of a predator-prey system. Math. Biosci. 1978, 39, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wiggins, S. Introduction to Applied Nonlinear Dynamical Systems and Chaos; Springer: New York, NY, USA, 1990. [Google Scholar]
- Guckenheimer, J.; Holmes, P. Nonlinear Oscillations, Dynamical Systems, and Bifurcations of Vector Fields; Springer: New York, NY, USA, 1983. [Google Scholar]
- Lin, X.; Wang, H. Stability analysis of delay differential equations with two discrete delays. Can. Appl. Math. Q. 2012, 20, 519–533. [Google Scholar]
- Wu, J. Theory and Applications of Partial Functional-Differential Equations; Springer: New York, NY, USA, 1996. [Google Scholar]
- Faria, T. Normal forms and Hopf bifurcation for partial differential equations with delays. Trans. Am. Math. Soc. 2000, 352, 2217–2238. [Google Scholar] [CrossRef]
- Polking, J.A.; Arnold, D. Ordinary Differential Equations Using MATLAB; Prentice-Hall: Englewood Cliffs, NJ, USA, 2003. [Google Scholar]
- Dhooge, A.; Govaerts, W.; Kuznetsov, Y.A. MATCONT: A MATLAB package for numerical bifurcation analysis of ODEs. ACM Trans. Math. Softw. 2003, 29, 141–164. [Google Scholar] [CrossRef]
- Alexander, J.C.; Yorke, J.A. Global bifurcations of periodic orbits. Am. J. Math. 1978, 100, 263–292. [Google Scholar] [CrossRef]
- Chow, S.N.; Mallet-Paret, J. The Fuller index and global Hopf bifurcation. J. Differ. Equ. 1978, 29, 66–85. [Google Scholar] [CrossRef] [Green Version]
- Wu, J. Symmetric functional-differential equations and neural networks with memory. Trans. Am. Math. Soc. 1998, 350, 4799–4838. [Google Scholar] [CrossRef]
- Hale, J. Theory of Functional Differential Equations; Springer: New York, NY, USA, 1977. [Google Scholar]
- Cooke, K.L.; Grossman, Z. Discrete delay, distributed delay and stability switches. J. Math. Anal. Appl. 1982, 86, 592–627. [Google Scholar] [CrossRef] [Green Version]
- Ruan, S.G.; Wei, J.J. On the zeros of transcendental functions with applications to stability of delay differential equations with two delays. Dyn. Contin. Discret. Impuls. Syst. Ser. A Math. Anal. 2003, 10, 863–874. [Google Scholar]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Figure | Figure 3a | Figure 3b | Figure 3c | Figure 3d |
|---|---|---|---|---|
| p | 5.1 | 3.2 | 1.2 | 0.9 |
| Figure | Figure 4a | Figure 4b | Figure 4c | Figure 4d |
| p | 1.55 | 1.645 | 1.6491 | 1.95 |
| Figure | Figure 7a | Figure 7b | Figure 7c | Figure 7d |
|---|---|---|---|---|
| p | 0.96 | 1.105 | 1.2 | 1.2068 |
| Figure | Figure 8a | Figure 8b | Figure 8c | |
| p | 0.965 | 1.052 | 1.05665 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Y.; Niu, B.; Wei, J. Dynamics in a Predator–Prey Model with Cooperative Hunting and Allee Effect. Mathematics 2021, 9, 3193. https://doi.org/10.3390/math9243193
Du Y, Niu B, Wei J. Dynamics in a Predator–Prey Model with Cooperative Hunting and Allee Effect. Mathematics. 2021; 9(24):3193. https://doi.org/10.3390/math9243193
Chicago/Turabian StyleDu, Yanfei, Ben Niu, and Junjie Wei. 2021. "Dynamics in a Predator–Prey Model with Cooperative Hunting and Allee Effect" Mathematics 9, no. 24: 3193. https://doi.org/10.3390/math9243193
APA StyleDu, Y., Niu, B., & Wei, J. (2021). Dynamics in a Predator–Prey Model with Cooperative Hunting and Allee Effect. Mathematics, 9(24), 3193. https://doi.org/10.3390/math9243193