1. Introduction
Network theory has a vast literature. In the book of Barabási [
1], the general aspects can be found, while the book of van der Hofstad [
2] is devoted to the mathematical models. Any network can be considered as a graph. The nodes of the network are the vertices, and the connections are the edges of the graph. A most famous model is the preferential attachment model proposed by Albert and Barabási [
3]. It is a discrete time network evolution model and it describes connections of two nodes. In real life, the meaning of connection can be any interaction or any cooperation.
There are models for cooperation more than two units. For example, Backhausz and Móri studied three-interactions in [
4]. Their model is generalised for
N-interactions by Fazekas and Porvázsnyik in [
5]. Both of these papers consider cliques where, inside a team, all members cooperate. In some sense, the opposite of the cliques, i.e., star-like connections were considered by Fazekas and Perecsényi [
6]. In [
6], there is no cooperation between two peripheral members of the team but all of them cooperate with the central member of the team. Despite [
3], in papers [
4,
5,
6] the preferential attachment rule is used for certain subgraphs and not for vertices.
We mention that in [
7] the Erdős–Rényi graph, the configuration model and the preferential attachment graph were studied when the population was split into two types. The mathematical tool of the analysis in [
7] is the theory of multi-type branching processes.
There are several continuous-time network evolution models. Here, we list only some papers using continuous time branching processes. Early works in this direction are [
8,
9]. Recently, in [
10], multi-type preferential attachment trees were studied. In [
10], the results of [
11] on multi-type continuous time branching processes were applied to describe the evolution of the network.
In this paper, we study a new network evolution model. The structure and the rules of the evolution of our model were inspired both by some everyday experiences and deep scientific results on motifs. On the one hand, we had in our mind activities and structures based on personal connections of the actors and where teams of some persons are important. Thus, we considered the friendship, the recruitment of party members and cooperation among party members, the recruitment and cooperation of volunteers, cooperation among scientists, informal connections among the employees of a company, etc. In these cases, the network consists of relatively small teams, a person can be a member of several teams at the same time, new teams can be born, and they can die, a newcomer can join the network if he/she joins an existing team.
On the second hand, our model is supported by the theory of motifs and their applications for real life networks. Here, we list only a few papers on this topic.
In [
12], the authors used network motifs: ‘patterns of interconnections occurring in complex networks at numbers that are significantly higher than those in randomized networks’. They developed an algorithm for detecting network motifs and found motifs with three or four vertices in biological and technological networks.
In [
13], the authors analyse the local structure of several networks such as protein signaling, developmental genetic networks, power grids, protein-structure networks, World Wide Web links, social networks, and word-adjacency networks. For the study, they used motifs on three or four vertices. In [
14], the authors found the numbers of all 3- and 4-node subgraphs, in both directed and non-directed geometric networks. In [
15], a method for the identification of all ordered 3-node substructures and the visualization of their significance profile are offered.
Therefore, we wanted to study a network that consists of small substructures, a node can be a member of several substructures at the same time, new substructures can be born and they can die, a new node can join to the network if it joins to an existing substructure.
Concerning the mathematical tools, we follow the line of Móri and Rokob [
16], where connections of two units were described by edges and the evolution of the edges was governed by a continuous-time branching process. In [
17] we extended the model of [
16] to 3-interactions. In this paper, our aim is to study networks containing groups of different sizes. For the sake of simplicity, we consider only groups of sizes 2 and 3. In this case, we can obtain explicit formulae for some quantities and our implicit formulae will be also transparent. The extension of our results to larger groups is possible. We emphasize that in our model all the nodes have the same type. Thus, despite [
7,
10], the theory of multi-type branching processes will be applied for groups of nodes and not just for the nodes.
In our model, when a new member joins to the network, it joins directly to an existing team. If that team consists of two members, then either a new team of two members or a new team of three members is produced. Similarly, if the new member joins to a team of three members, then new teams having two or three members can emerge. Thus, we obtain a two-type continuous time branching process, in which an individual can be either an edge or a triangle of the network.
The starting individual (that is the ancestor) can be either an edge or a triangle. It produces offspring at each time given by the driving branching process. These offspring can be edges or triangles, and after their birth they also start their own reproduction processes. An evolution step of the generic triangle is the following. Always one vertex is born, but it is connected to the triangle with random number of edges. The new vertex can be connected to or 3 vertices of the triangle. Therefore, the offspring of a triangle can be a new edge, or one new triangle or three new triangles. The lifetime of a triangle is determined by the number of its offspring. The reproduction process of an edge is similar.
The structure of this paper is the following. In
Section 2, a detailed description of our model is given. In
Section 3, the general results are presented. These are the survival functions of an edge and of a triangle (Theorem 1), the mean offspring number of an edge and of a triangle (Corollary 1), the Perron root and the Malthusian parameter. As usual, we obtain only implicit expression for the Malthusian parameter, but our expression is simple and numerically tractable.
In
Section 4, asymptotic theorems on the number of edges and triangles (Theorem 2) are proved. Both of them have magnitude
on the event of non-extinction, where
is the Malthusian parameter. To prove Theorem 2, we use the underlying branching process counted with certain random characteristics and apply the asymptotic theorems of [
11].
In
Section 5, the generating functions are calculated. Using the generating functions, the probability of extinction are studied. In
Section 6, the asymptotic behaviour of the degree of a fixed vertex is considered. Here, we again apply the asymptotic theorems of [
11] but with other characteristics than in
Section 4. In
Section 7, we present some simulation results supporting our theorems. Our figures and tables show that the values obtained by simulation fit well to the theoretical results.
The proofs are based on known general results of multi-type continuous-time branching processes. Therefore, for the reader’s convenience, in
Section 8, we list several results on multi-type Crump–Mode–Jagers processes.
We mention that our model was presented in our conference paper [
18]. In that paper some preliminary theoretical results were announced together with some numerical evidence but without mathematical proofs.
2. The Model
We study the following network evolution model. At the initial time the network consists of one single object, this object can be either an edge or a triangle. This object is called the ancestor. During the evolution, this ancestor object produces offspring objects, which can be either edges or triangles. Then, these offspring objects produce their offspring objects and so on. The reproduction times of any fixed object, including the ancestor, are the occurrences in its own Poisson process with rate 1.
From the theory of branching processes, we apply the following usual assumptions. That is we suppose that the reproduction processes of different objects are independent. Moreover, we assume that the reproduction processes of the edges are independent copies of the reproduction process of the generic edge. Similarly, the reproduction processes of the triangles are independent copies of the reproduction process of the generic triangle.
First, we explain the evolution of the generic edge. A Poisson process with parameter 1 gives its reproduction times. At any jumping time of this Poisson process, a new vertex appears and it is connected to the generic edge with one or two edges. The probability that this new vertex is connected to the generic edge by one new edge is , where . The other end point of this new edge is chosen from the two vertices of the generic edge uniformly at random. We see that in this case the generic edge produces always one new edge. The other case is that when the new vertex is connected to both vertices of the generic edge. Its probability is . In this second case the offspring of the generic edge is a triangle consisting of the generic edge and the two new edges. We emphasize that in this last case the generic edge itself and the new triangle will produce offspring, but the two new edges are not substantive parts of the reproduction process, so they alone will not produce offspring.
The reproduction process of the generic triangle is similar. The Poisson process with rate 1 corresponding to the generic triangle is denoted by . The jumping times of are the birth times of the generic triangle. At every birth time a new vertex is born and it joins to the existing graph so that it is connected to our generic triangle with 1, 2 or 3 edges. Denote by () the probability that the new vertex is connected to j vertices of our generic triangle. The vertices of the generic triangle to be connected to the new vertex are chosen uniformly at random.
By the above definition of the evolution process, at each birth step we add precisely 1 new vertex. When the new vertex is connected to one vertex of the generic triangle, the generic triangle gives birth to one new edge. This event has probability . However, in the remaining two cases we count only the new triangles and not the new edges. When the new edge is connected to the generic triangle by two edges, these two edges and one edge of the generic triangle form a new triangle. Therefore, with probability , the generic triangle produces one child triangle. When the new edge is connected to the generic triangle by three edges, these edges and the edges of the generic triangle form three new triangles. Thus, with probability , the generic triangle produces three children triangles.
Any edge is called a type 2 object, and any triangle is called a type 3 object. We use subscript 2 for edges and subscript 3 for triangles. Thus, we denote by
the number of type
j offspring of the type
i generic object up to time
t (
). Recall that
,
, are point processes. Then
gives the total number of offspring (that is both edges and triangles) of the generic edge up to time
t. We can also see that
is the number of all offspring (edges or triangles) of the generic triangle up to time
t.
We denote by
the birth times of the generic triangle, and we denote by
the corresponding total litter sizes. That is, at the
ith birth event, the generic triangle bears
children being either triangles or edges. The discrete random variables
are independent and identically distributed having distribution
. By the above evolution process, we have
We assume that the litter sizes are independent of the birth times.
Let
be the life-length of the generic triangle. It is a finite, non-negative random variable. We assume that the reproduction terminates at the death of the individual. Therefore,
for
. Then, the reproduction process of a triangle can be formulated as
where
is the Poisson process,
gives the total number of offspring of the generic triangle before the
th birth event and by
we denote the minimum of
.
The survival function of the life-length. Let
denote the distribution function of the triangle’s life-length
. Then, the survival function of
is
where
is the hazard rate of the life-length
. We suppose that the hazard rate depends on the total number of offspring, so that
with fixed positive constants
b and
c.
Let
be the life-length of the generic edge. Then,
for
. As the edge always gives birth to one offspring (which can be an edge or a triangle); therefore,
is the total number of offspring of the generic edge, where
is the Poisson process.
We denote by
the distribution function of
. Then, the survival function of the life-length of an edge is
where
is the hazard rate of the life-length
. We suppose that
is of the form
.
We emphasize that we do not delete any edge or any triangle when it dies, because its ingredients can belong to other triangles or edges, too. Thus, dead triangles and edges will be considered as inactive objects not producing new offspring.
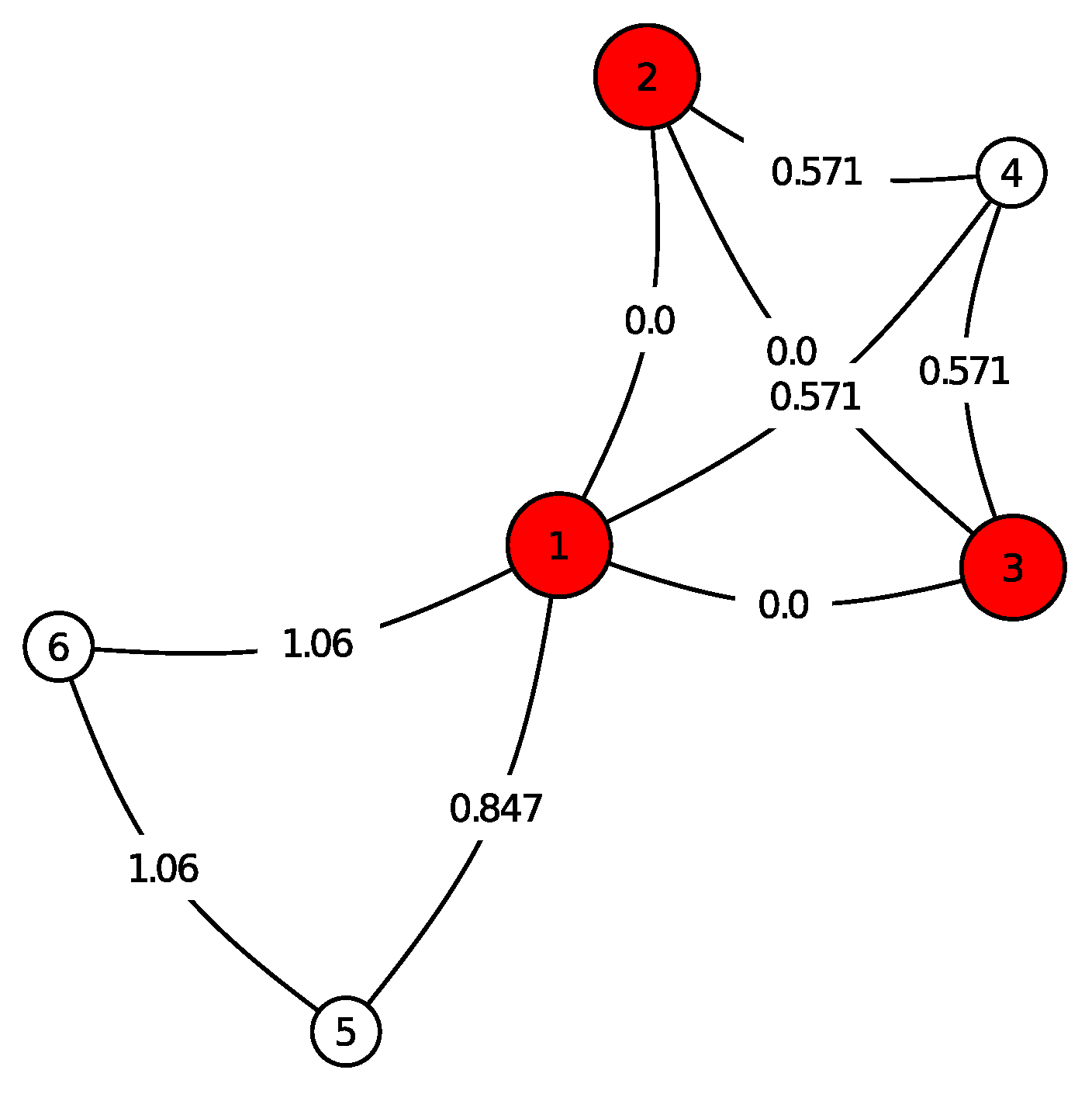
In
Figure 1, an example is shown for our graph evolution model. For a clear view it contains only three birth steps after the initial time
. The nodes of the ancestor are highlighted by red. The edges are labelled with the birth times
t. The following objects appear in
Figure 1, which are described by the labels of their nodes:
(1-2-3): is a triangle, the ancestor with birth time ,
(1-2-3-4): represents three triangles, i.e., the offspring of (1-2-3) at its first reproduction time ,
(1-5): an edge, offspring of (1-2-3) with birth time ,
(1-5-6): a triangle, offspring of (1-5) with birth time .
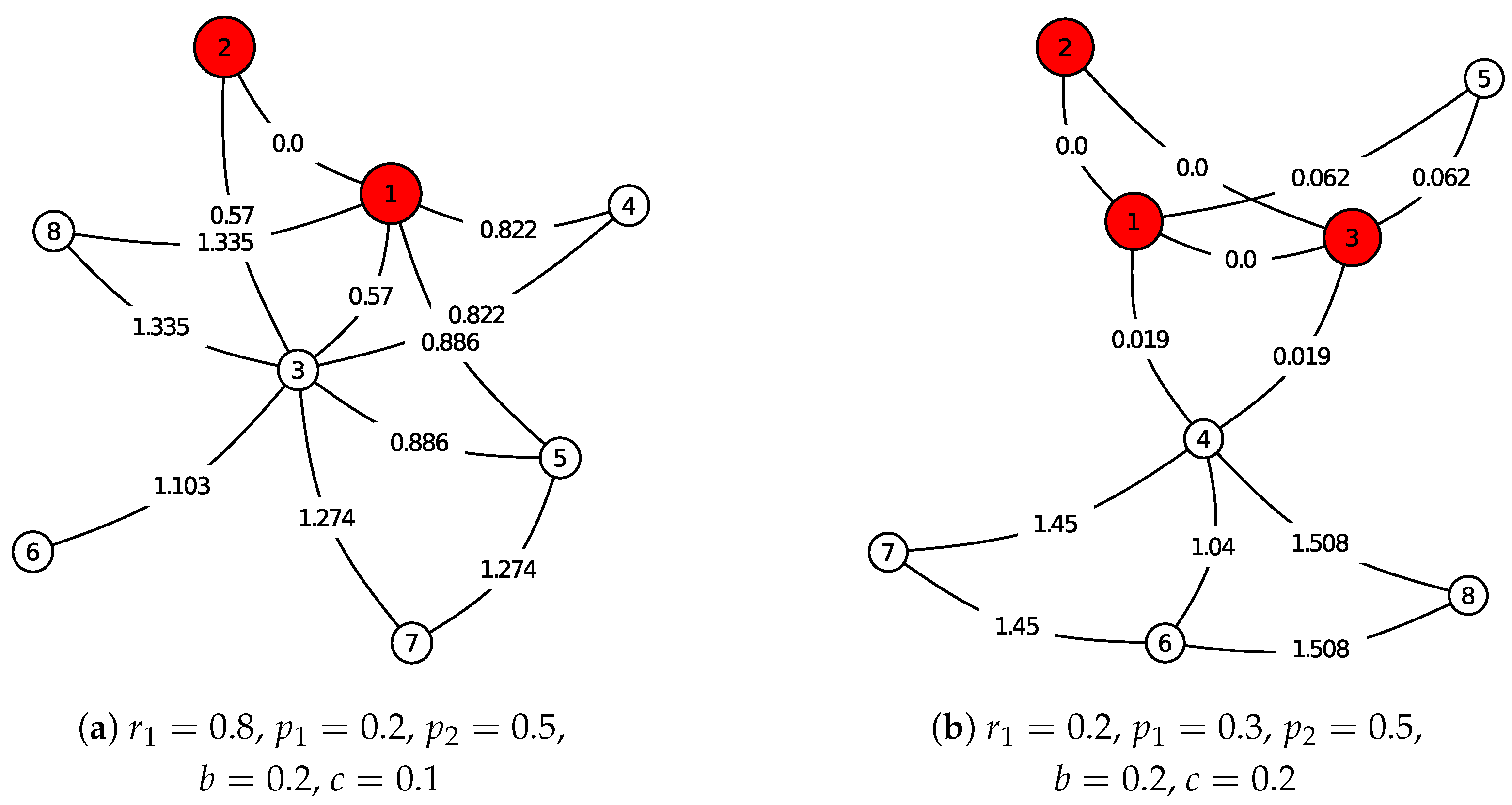
Two more examples are shown in
Figure 2 with different parameters. In
Figure 2a the ancestor is an edge, while in
Figure 2b the ancestor is a triangle.
3. General Results
The survival functions.
Theorem 1. The survival function for a triangle is The survival function for an edge is Proof. At the first part of the proof we omit subscripts 2 and 3, because the calculations are the same for edges and triangles. Let
and assume that
. Then, the first
k birth events happened before time
t. Thus, the birth times
and the corresponding litter sizes
are known. Therefore, the reproduction process
is also known for
. By (
5), a simple calculation shows that the survival function of an object is
Let
be an ordered sample of size
k from uniform distribution on
. Then, the joint conditional distribution of the birth times
given
, coincides with the distribution of
. Therefore
because
. The litter sizes
are independent identically distributed random variables, which are independent also of
. Hence
where we applied that
is uniformly distributed. Using this and the total probability theorem, we find
Therefore, the survival function for a triangle is
Finally, the survival function for an edge is
□
The mean offspring number. Let us denote by the expectation of the number of type j offspring of a type i mother until time t.
Corollary 1. For any , we havewhere For any , we havewhere because . Proof. We have
where
is the number of type
j offspring of a type
i mother at her
kth birth event. Using Wald’s identity, the average number of children is
Using that
is a Poisson process with rate 1, and
is bounded for any
t, from (
14), we obtain that the average number of children is
Now, consider
. Applying (
9) and using the substitution
, we obtain
If we write
instead of
, then we obtain
. Thus, we obtained (
10). Moreover, with
, we have
. Thus, (
11) follows from (
16).
Now, we turn to
. Applying (
8), and using the substitution
, we obtain,
As
, so from (
15) we obtain
. Using that
, we obtain
. Thus, we obtained (
12). Moreover, we have
. Thus, (
13) follows from (
17) with
. □
Let
be the Laplace transform of
.
Proposition 1. For any , we havewhere For any , we havewhere Proof. Apply the definition of , Corollary 1 and substitution . □
The Perron root and the Malthusian parameter. Let
be the matrix of the Laplace transforms. Direct calculation gives that the characteristic roots of
are
The greater of the values
and
is called the Perron root, so
is the Perron root.
We assume that our process is supercritical; that is,
For supercriticality, condition
is sufficient.
That value of
for which the Perron root is equal to 1 is called the Malthusian parameter. Thus, using the usual notation in the theory of branching processes,
is the Malthusian parameter if
. In this paper, we assume the existence of the Malthusian parameter. From relation
and (
23), we obtain that the Malthusian
satisfies the equation
Later, we use the eigenvectors of
. To this end, let
be the Malthusian parameter, and let
be the right eigenvector of
corresponding to eigenvalue 1 and satisfying condition
. Then, direct calculation shows that
Again, let
be the Malthusian parameter and let
be the left eigenvector of
satisfying condition
. Direct calculation shows that
4. Asymptotic Theorems on the Number of Triangles and Edges
In this section, we use Proposition 4 from
Section 8. So we should check the conditions given in
Section 8. For condition (a) from
Section 8, we should guarantee that not all measures
are concentrated on a lattice. By Corollary 1, these measures are absolutely continuous, and thus it is satisfied.
Concerning condition (b1), we underline that we suppose the existence of a positive Malthusian parameter
. To this end, in this section, we assume that (
25) has a finite positive solution
. We can check numerically the existence of this value. For (b2), we assume (
24). Condition (c) from
Section 8 will be checked later in the proofs of the results together with other conditions related to it.
Now, we analyse condition (d). We can see from Corollary 1 that
and
are positive. Thus, we can concentrate on parameters
and
. If
, then (d) is not satisfied; however, in this case, one can study separately the process of edges (it grows at any birth time by 1), and the process of triangles (this is described in [
17]). If
and
, then (d) is not satisfied, and the evolution process is an alternating one. If either
or
, then (d) is not satisfied.
To guarantee condition (d), in this section, we assume that
,
, and it is excluded that both
and
are satisfied at the same time. In this case, condition (d) from
Section 8 is satisfied.
The denominator in the limit theorem. In the following theorem, we need the next formulae. In
Section 8, we see that the denominator of
in the limiting expression is independent of
, and it is
It can be written in the form (and considering our two-dimensional case)
Here,
and
are from Equations (
26) and (
27). Moreover, by Corollary 1 or by Proposition 1, we have that
where
Now, we turn to the number of edges and triangles. Recall that an edge is a type 2, and a triangle is a type 3 object.
Theorem 2. Assume that (
24)
is satisfied and (
25)
has a finite positive solution α. Assume that , and it is excluded that both and are satisfied at the same time. Let denote the number of all edges being born up to time t if the ancestor of the population was a type i object, . Thenalmost surely for . Let denote the number of all edges present at time t if the ancestor of the population was a type i object, . Thenalmost surely for . Let denote the number of all triangles being born up to time t if the ancestor of the population was a type i object, . Thenalmost surely for . Let denote the number of all triangles present at time t if the ancestor of the population was a type i object, . Then,almost surely for . The quantities and are a.s. non-negative, , and are a.s. positive on the event of survival.
Proof. We apply Proposition 4. To obtain condition (
71), it is enough to show that
where
and
If
, then
is the birth process of an edge, and the children can be both edges and triangles. Therefore, at each birth, there is one child. Therefore,
where
are the jumps of the Poisson process
. In the Poisson process
the distribution of the interarrival time (
) is exponential with rate 1. Therefore,
has
-distribution
. Using this, we have
Let us denote by
the interarrival time
. Let
be an exponentially distributed random variable with rate 1 that is independent of
M. Then,
Therefore, the distribution of
coincides with the distribution of
M. Therefore, using (
40), we have
Thus, (
37) is true for
.
If
, then
is the birth process of a triangle and the children can be both edges and triangles. Therefore, at each birth there are at most three children. Therefore,
where
are the jumps of the Poisson process
. By the above calculation
, so (
37) is true for
.
If we show that
, for
, then conditions (c) and
of
Section 8 will be proved. Now, for
and
, we have from Corollary 1
because
.
For
and
, we have from Corollary 1
Thus, conditions (c) and
of
Section 8 are proved.
Now, turn to the number of edges.
To obtain (
33), let
if
x is an edge, and
if
x is a triangle. Therefore,
and
. Conditions
and
of
Section 8 are satisfied. Thus, (
69) and (
70) imply (
33).
To obtain (
34), let
if
x is an edge and it is present at
t, and
if
x is a triangle. Therefore,
and
. Conditions
and
of
Section 8 are satisfied. Now,
Thus, (
69) and (
70) imply (
34).
Now, we turn to the number of triangles.
To obtain (
35), let
if
x is an edge, and
if
x is a triangle. Therefore,
and
. Conditions
and
of
Section 8 are satisfied. Thus, (
69) and (
70) imply (
35).
To obtain (
36), let
if
x is an edge, and
if
x is a triangle, and it is present at
t. Therefore,
and
. Conditions
and
of
Section 8 are satisfied. Now,
Thus, (
69) and (
70) imply (
36). □
5. Generating Functions and the Probability of Extinction
The joint generating function of , and . Recall that
is the Poisson process describing the reproduction times of the generic edge and
is its life length. Thus,
is the joint distribution of the offspring size of the generic edge during its whole life and its last reproduction time. We have
where
is the
ith jumping time of the Poisson process
. Thus, it again shows that
is the probability that the
ith birth event is the last one that occurred before death, and the total numbers of the two types of offspring up to time
are equal to
j and
k, respectively.
Now, consider the sequence
Let
and assume for a while that
and
are fixed. Then, using (
4) and (
5) for the hazard rate, we can calculate that, for fixed
and
,
We know that the increment
is exponential with parameter 1; therefore,
At each birth step, the new individual can be either an edge or a triangle. Therefore, using the above calculations, the total probability theorem, and the independence of the type of the newly born individual and
, we have the following recursion for
.
Now, by the definition of
, we can see that
where by (
41),
is the probability that the generic individual dies before the next birth event.
Let
. Then, from (
42), we obtain the following recursion for the sequence
where the initial values are
Now, we calculate the generating function
of the sequence
. We have
First, multiplying with
and then taking the sum of both sides of (
43), we obtain
where
is given by (
44), and we define
if
or
. From this equation, we find
Let
. Now, substituting
y with
,
z with
in (
45), we can obtain the following linear differential equation.
with the initial value condition
Now, we can use the well-known method for linear differential equations. We obtain that the solution of the initial value problem (
46) and (
47) is
With
, we obtain that
We need the generating function of
. It is
From here, we obtain
Proposition 2. The joint generating function of , and iswhere . Corollary 2. The generating function of the total offspring distribution of the generic edge is The joint generating function of , and . Here, we study the offspring of a triangle. To distinguish the notation of this subsection and the previous subsection, but avoid too many subscripts, we use bar. Thus, here
,
,
,
and
denote quantities relating offspring of the generic triangle. Recall that
is the Poisson process describing the reproduction times of the generic triangle and
is the life length of the triangle. Thus,
is the joint distribution of the offspring size of the generic triangle during its whole life and its last reproduction time. We have
where
is the
ith jumping time of the Poisson process
. Thus, we again show that
is the probability that the
ith birth event is the last one that happened before death, and the total numbers of the two types of offspring up to time
are equal to
j and
k, respectively.
Let
, and assume for a while that
and
are fixed. Then, using (
4) and (
5) for the hazard rate, we can calculate that, for fixed
and
,
We know that the increment
is exponential with parameter 1; therefore,
At each birth step, the new individual can be either an edge or a triangle. Therefore, using the above calculations, the total probability theorem, and the independence of the type of the newly born individual and
, we have the following recursion for
.
Now, by the definition of
, we can see that
where by (
51),
is the probability that the generic individual dies before the next birth event.
Now, let
. Then, from (
52), we obtain the following recursion for the sequence
where the initial values are
Now, we calculate the generating function
of the sequence
. We have
First, multiplying with
and then taking the sum of both sides of (
53), we obtain
where
is given by (
54) and we define
if
or
. From this equation, we find
Let
. Now, substituting
y with
,
z with
in (
55), we can obtain the following linear differential equation.
with the initial value condition
One can see that the solution of the initial value problem (
56) and (
57) is
With
, we obtain that
Therefore, the generating function of
is
From here, we obtain
Proposition 3. The joint generating function of , and iswhere . Corollary 3. The generating function of the total offspring distribution of the generic triangle is The probability of extinction. In Theorem 3, we give the probability of extinction. To determine the extinction probability of the process, we consider the well-known embedded multi type Galton–Watson process. At time
, the 0th generation of the Galton–Watson process consists of a single individual, i.e., the ancestor. The first generation consists of all offspring of the ancestor. The offspring of the individuals of the
nth generation form the
th generation. Under some assumptions, the extinction of our original process has the same probability as the extinction of this embedded Galton–Watson process. The reproduction process
gives the number of type
j offspring of an ancestor of type
i up to time
t. With
, we obtain that the total number of offspring is
. Therefore, Corollary 1 gives us the
matrix of the expected total offspring number as
Actually, is the expected offspring number of the embedded Galton–Watson process.
Let and denote the probability of extinction of our process when the ancestor is an edge, resp. triangle.
Theorem 3. Assume that , and it is excluded that both and are satisfied at the same time. Let be the Perron–Frobenius root of . If , then . If , then and . In any case, is the smallest non-negative solution of the vector equationwhere and are given in Corollaries 2 and 3. Proof. We apply Theorem 7.1 in Chapter 1 of [
19]. By Corollary 1,
and
is finite for any
. Therefore, by Theorem 7.1 in Chapter 3 of [
19], the extinction of our original process has the same probability as the extinction of the embedded Galton–Watson process. Thus, we can apply Theorem 7.1 in Chapter 1 of [
19]. Here,
is the matrix of the expected offspring numbers of the embedded Galton–Watson process. Now,
is positively regular because we assume that
,
and it is excluded, that both
and
are satisfied at the same time. Thus, our result follows from Theorem 7.1 in Chapter 1 of [
19]. □
6. The Asymptotic Behaviour of the Degree of a Fixed Vertex
The process of the ‘good children’. To describe the degree of a fixed vertex, we introduce a new branching process that we call the process of ‘good children’. This process contains those objects that contribute to the degree of the fixed vertex. We can see that a newly born vertex can have 1 or 2 edges if its parent is an edge object and 1, 2 or 3 edges if its parent is a triangle object.
First, we consider the case when the newly born vertex has one edge, and thus, at the beginning, it belongs to an edge object. In this paragraph, we call this edge the ‘parent’ edge. We fix the newly born vertex. Then, we distinguish those children objects of the ‘parent’ edge, which contribute to the degree of our fixed vertex. We call a child object of the ‘parent’ edge a ‘good child’ if it contains our fixed vertex. We can see that only the ‘good children’ and their ‘good children’ offspring can contribute to the degree of the fixed vertex. Then, the distribution of the number of ‘good children’ at a reproduction event of the ‘parent’ edge is
where
denotes the number of edge type ‘good children’ and
denotes the triangle type ‘good children’. We have to consider the reproduction process of the ‘good child’, which is the following
where
denotes the number of all edge type ‘good children’, and
denotes the number of all triangle type ‘good children’ born by the ‘parent’ edge,
are i.i.d. copies of
and
are i.i.d. copies of
. Using Corollary 1, we see that the mean values of the number of edge type and triangle type ‘good children’ are
Now, consider the second case where the newly born vertex has two edges, and thus the ‘parent’ object is a single triangle. Let
and
denote the number of edge, resp. triangle type ‘good children’ of the ‘parent’ triangle. The distribution of the number of ‘good children’ will be the following
Let
denote the number of all edge type ‘good children’, and
denote the number of all triangle type ‘good children’ born by the ‘parent’ triangle. We obtain from Corollary 1 that
Therefore, from Proposition 1, it is easily seen that the Laplace transforms of the average number of offspring are
Let
be the matrix of the previous Laplace transforms. The Perron root that is the largest eigenvalue of
is
In the following, we assume supercriticality of the ‘good children’ process; that is, we suppose that
. We can see that the reproduction process of the ‘good children’ is supercritical if
We assume the existence of finite and positive Malthusian parameter of the ‘good children’ process. Thus, let
be the Malthusian parameter; it satisfies equation
. From this equation and from (
63), we see that
is the solution of
Let
denote the right eigenvector of
corresponding to the eigenvalue 1, and let
be the left eigenvector with the conditions
and
. Direct calculations show that
Limit results for the degree. We have already mentioned that the ‘good children’ and only they can contribute to the degree of the fixed vertex. Thus, its degree is equal to the initial degree plus the number of ‘good children’. Let be the degree of a fixed vertex at time t after its birth in the case when the vertex belongs to an edge at its birth. Similarly, is its degree in the case when the vertex belongs to triangle at its birth. Up to an additive constant, is the number of ‘good children’ offspring of an i type ‘parent’ object at time t. It is the sum of the number of edge type ‘good children’ and the triangle type ‘good children’ . To apply Proposition 4, we can use the same method as in Theorem 2. Thus, for the edges, we can again use the random characteristic if x is an edge and if x is a triangle, but the underlying process is the process of ‘good children’. This is similar for triangles.
Therefore, we have almost surely
for
, where
and
are positive on the event of non-extinction of the ‘good children’.
The last case is when the newly born vertex has three edges. Then, three triangles contribute to the degree of that vertex. Let
be the degree of this vertex. Then,
is the sum if ‘good’ offspring of three triangles. Thus, almost surely,
where
are independent copies of
.
Checking the conditions of Proposition 4 for the ‘good children’ process. To complete the previous reasoning, we should check the conditions of Proposition 4. First, we find the the denominator in the limit theorem that is we calculate
. By
Section 8, we see that
Here,
and
are the eigenvectors. Moreover,
where
is the Malthusian parameter in the process of ‘good children’ and
,
denotes the derivatives given in (
31) and (
32).
Condition (a) of Proposition 4 is true because the measures
are non-lattice as they are absolutely continuous. For condition (b1), we assume the existence of a positive Malthusian parameter. That is, we assume that (
64) has a finite and positive solution
. Condition (b2) is true, because we assume that
. Condition (c) is a consequence of
Section 4, because
has shape
, where
c is positive number.
To guarantee condition (d), in this section, we assume that , , and it is excluded that both and are satisfied at the same time. Conditions (i)-(ii)-(iii) and (v) are true because of the shape of . Conditions (iv) and (vi) are consequences of as one can see from the proof of Theorem 2.
The extinction of the degree process. The extinction of the degree process means that the degree of the vertex does not increase after a certain time, that is, the reproduction process of the ‘good children’ dies out. The probability of this kind of extinction is the smallest non-negative root
of the equation
where
and
are the generating functions of the total ‘good children’ distribution of an edge, resp. a triangle. Now, by (
61) and (
62),
where
is the generating function of
, and
is the joint generating function of
and
. Here, by (
49),
Similarly,
where by (
59), the generating function of
is
Moreover, the joint generating function of
and
is
7. Simulations
In this section, we provide some empirical results for our asymptotic theorems. We generated our process in the programming language Julia. We needed an environment, where the priority queues were highly applicable. Using this structure, the running time was reasonable. A more detailed explanation of the algorithm can be found in [
20].
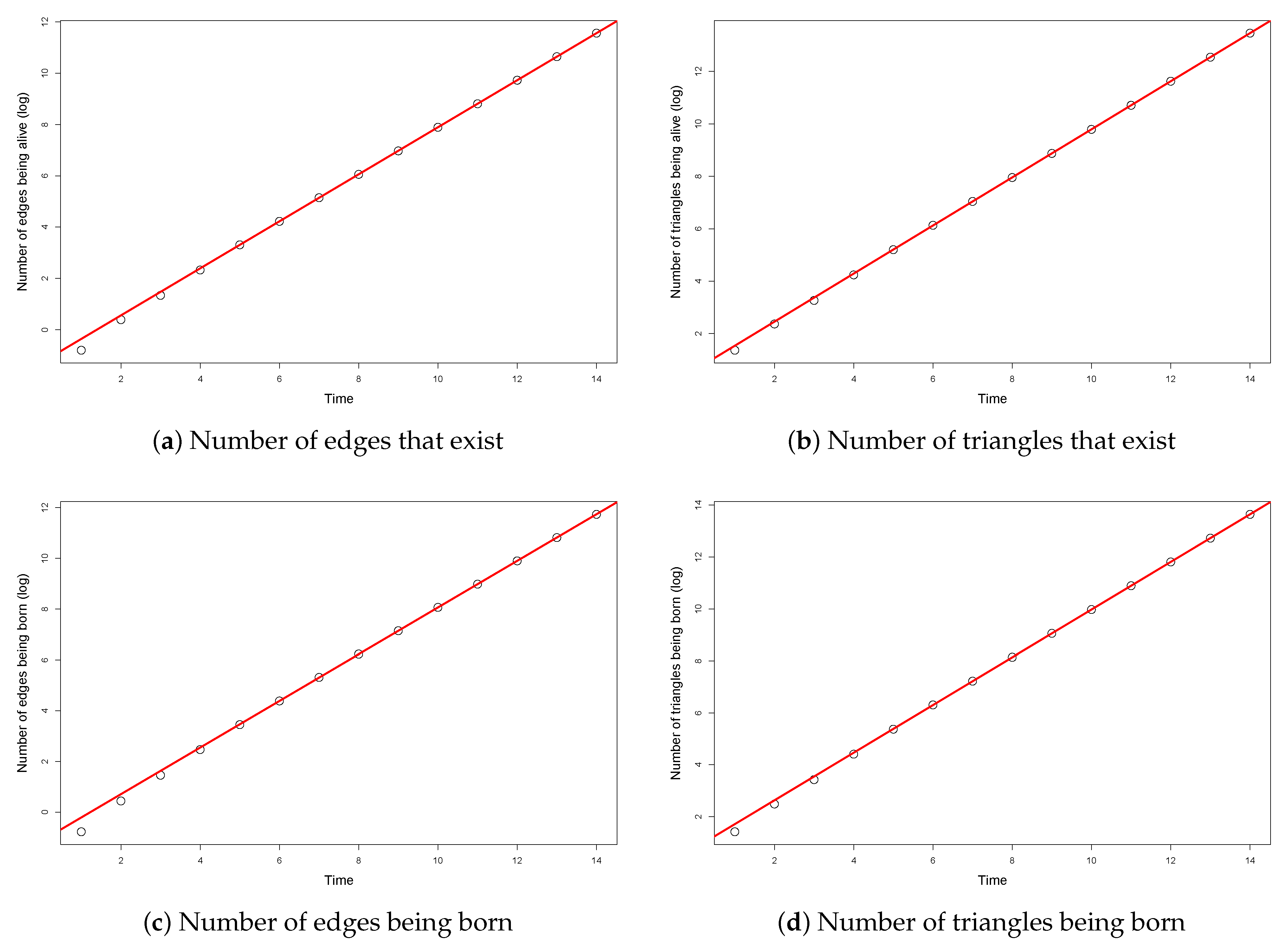
According to Theorem 2, for large
t, the graphs of the numbers of edges and triangles are approximately straight lines on the logarithmic scale. To obtain empirical evidence of our Theorem 2, we investigated the slope of the simulated number of edges and triangles being born and being present up to time
t on the logarithmic scale. The initial instability of the single processes (
Figure 3) motivated us to exclude the first few observations from the calculations, but the lack of them was not relevant, because the asymptotic properties can be observed in the later stage of the processes.
For each parameter set, we stored the mentioned measurements only in integer time steps, and then we took the average of 100 simulated processes. In
Figure 4, an example is shown for a specific parameter set (
,
,
,
,
). The values of the averages are plotted by dots. In each case, we fitted a regression line (plotted by continuous red line) to the last 9 values. We can see that the fit is perfect, thus, supporting our theorem.
Our main goal was to obtain a 95% confidence interval for the slope of the linear regression line, as that was our simulated approximation of the Malthusian parameter
.
Table 1 contains the boundaries of the 95% confidence intervals for
. The columns labelled with
and
refer to the lower and the upper bounds obtained from simulations, while the column of
refers to the numerical solution of Equation (
25).
For each fixed parameter set , we present the confidence intervals calculated from the number of edges being born (E) resp. being present () and from the number of triangles being born (T) resp. being present () up to time . The confidence intervals containing the numerical Malthusian parameter are highlighted with the * symbol. We see that any confidence interval is narrow, and it either contains , or is very close to the interval. These results show that the approximation is good for moderate values of t.
Finally, we present some simulation results for Theorem 3, that is, for the probability of extinction of the evolution process. We made the following computer experiment for any fixed parameter set and for type 2 and type 3 ancestors. We started to generate the process. If this process reached birth steps, then we stopped it and considered it as a non-extinct process. Otherwise, when the process did not reach birth steps, then the process died out. Applying the above method, we generated processes for each parameter sets and counted the relative frequencies of the processes being extinct.
In
Table 2, we show some of the results. Column Ancestor contains the type of the ancestor. In the column Numeric we show the numeric solution of the non-linear equation in Theorem 3. We used Julia’s trust region method. Column Simulation contains the relative frequencies extracted from the simulations. The simulation results slightly underestimate the numeric values. This is reasonable because we stopped all processes at a fixed time.
8. Basic Facts on Branching Processes
In our paper, we use known results of the theory of continuous-time branching processes. The single type general Crump–Mode–Jagers branching processes have been described e.g., in [
21,
22,
23]. The general multi-type branching processes have been studied, e.g., in [
11,
19,
24].
Here, we give a short description of the general multi-type branching processes based on [
11]. The individuals of this process can be of
p different types, which we denote by
. Any individual
x is described by the quantities
. The quantities
are independent copies of the quantities
. Thus, we should give the definition of
, which we consider as the quantities corresponding to the generic individual.
The lifetime is a non-negative random variable which is not necessarily independent from the reproduction. The lifetime distribution is . The reproduction process is , . Here, the random point process describes the births of type j offspring of a type i mother. gives the number of type j offspring of a type i mother up to time t. is determined by the birth events and the numbers of offspring. The process starts at time with one individual called the ancestor and denoted by . When a child is born, it starts its own reproduction process and so on. The birth time of the individual x is denoted by .
Let be a non-negative random function that describes a certain aspect of the life history of the individual. It is usually assumed that for . Then, is called a random characteristic. Let be another random characteristic. Thus, the behaviour of the individual x is described by .
Let us define the branching process
counted by the characteristic
as
where we summarize for all individuals
x. Here, the left subscript
of
Z and of the birth time
is important, because it denotes that the process starts with ancestor
and the type of
has influence for the evolution of the population.
Let us denote by the reproduction function, which is the expected reproduction number .
The following facts are well-known (see [
11] or [
24]).
We assume the following basic conditions in this section.
(a) Not all of the measures are concentrated on a lattice.
Let
be the Laplace transform of
. Let
be the matrix
(b1) There exists a positive Malthusian parameter that is a finite positive value so that has finite entries only, and the Perron–Frobenius root of is equal to 1. Here, the Perron–Frobenius root is the largest eigenvalue of the matrix. Let be the right positive eigenvector and the left positive eigenvector of corresponding to the Perron–Frobenius root. We normalize them as and .
(b2) The matrix has an infinite entry, or all of them are finite, and its Perron–Frobenius root is greater than 1.
(c) The first moment of
is finite and positive; that is,
(d) There exists a finite positive integer K so that all elements of the Kth power of the matrix are positive.
Proposition 4. Let α be the Malthusian parameter. Assume that the random characteristic Φ satisfies the following conditions:
- (i)
,
- (ii)
The trajectories of Φ belong to the Skorohod space D, i.e., they do not have discontinuities of the second kind,
- (iii)
.
Assume also
- (iv)
- (v)
for some for any ancestor. Then,is likely, where i is the type of , is an a.s. non-negative random variable depending on the type of the ancestor but not depending on the choice of Φ
. If, in addition, we assume that
- (vi)
then , is positive with positive probability, and is a.s. positive on the survival set.
The proof is a simple consequence of Theorem 2.4 and Proposition 4.1 of [
11].
9. Discussion
In this paper, a new network evolution model was introduced. This model was inspired by those networks where small substructures play important role. In social life, such substructures could be a group of friends. In the theory of networks, these substructures are called motifs. In this paper, for the sake of simplicity, we consider only two types of substructures, the edges and the triangles. The novelty of the paper is the usage of a two-type continuous time branching process to describe these two types of interactions. Thus, despite [
7,
10], the theory of multi-type branching processes was applied for certain substructures of the network and not just for the nodes. Our paper extends the former studies of [
16,
17], where only one type of interaction was considered.
In this paper, we proved that the magnitude of the number of triangles on the event of non-extinction is
, where
is the Malthusian parameter. We obtained similar results for the number of edges. We also studied the degree process of a fixed vertex and the probability of the extinction. Our results are similar to the ones obtained for the simpler models in [
16,
17]. In addition to mathematical proofs, the results were illustrated by simulations.
In future extensions of the model, more than two types of substructures can be studied using the theory of multi-type branching processes.







{kind=link}
{kind=link}
{kind=link}
{kind=link}