Paraquat-Mediated Oxidative Stress in Anopheles gambiae Mosquitoes Is Regulated by An Endoplasmic Reticulum (ER) Stress Response

 ,
, 

Abstract

1. Introduction
2. Materials and Methods
2.1. Mosquito Rearing, Experimental Treatments, and ROS Induction Assays
2.2. Extraction, Solubilization, and Digestion of Proteins
2.3. Online 2D LC-MS/MS Analysis
2.4. Database Searching and Label-Free Quantification Analysis
2.5. qRT-PCR
2.6. Statistical Analyses
3. Results
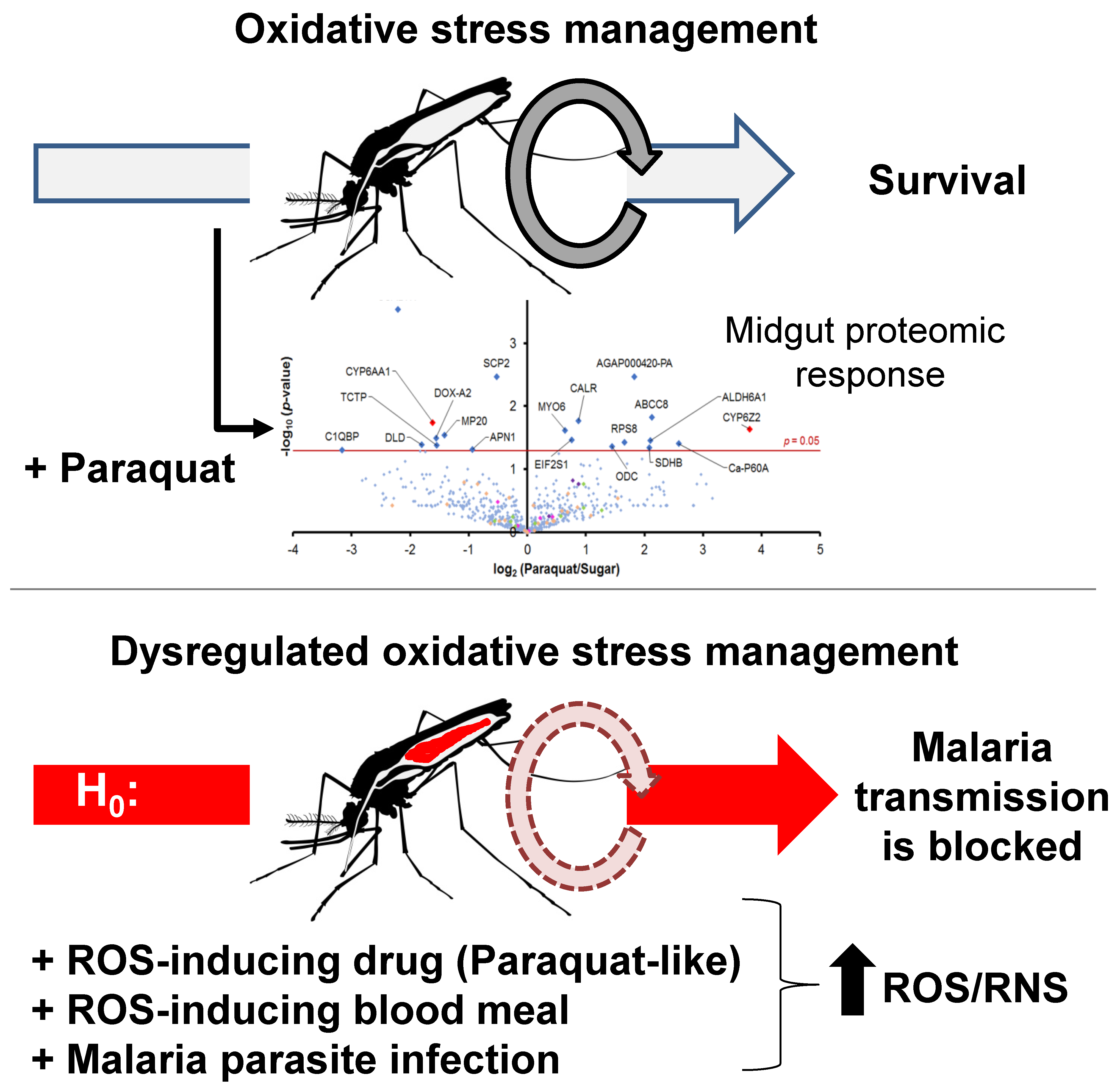
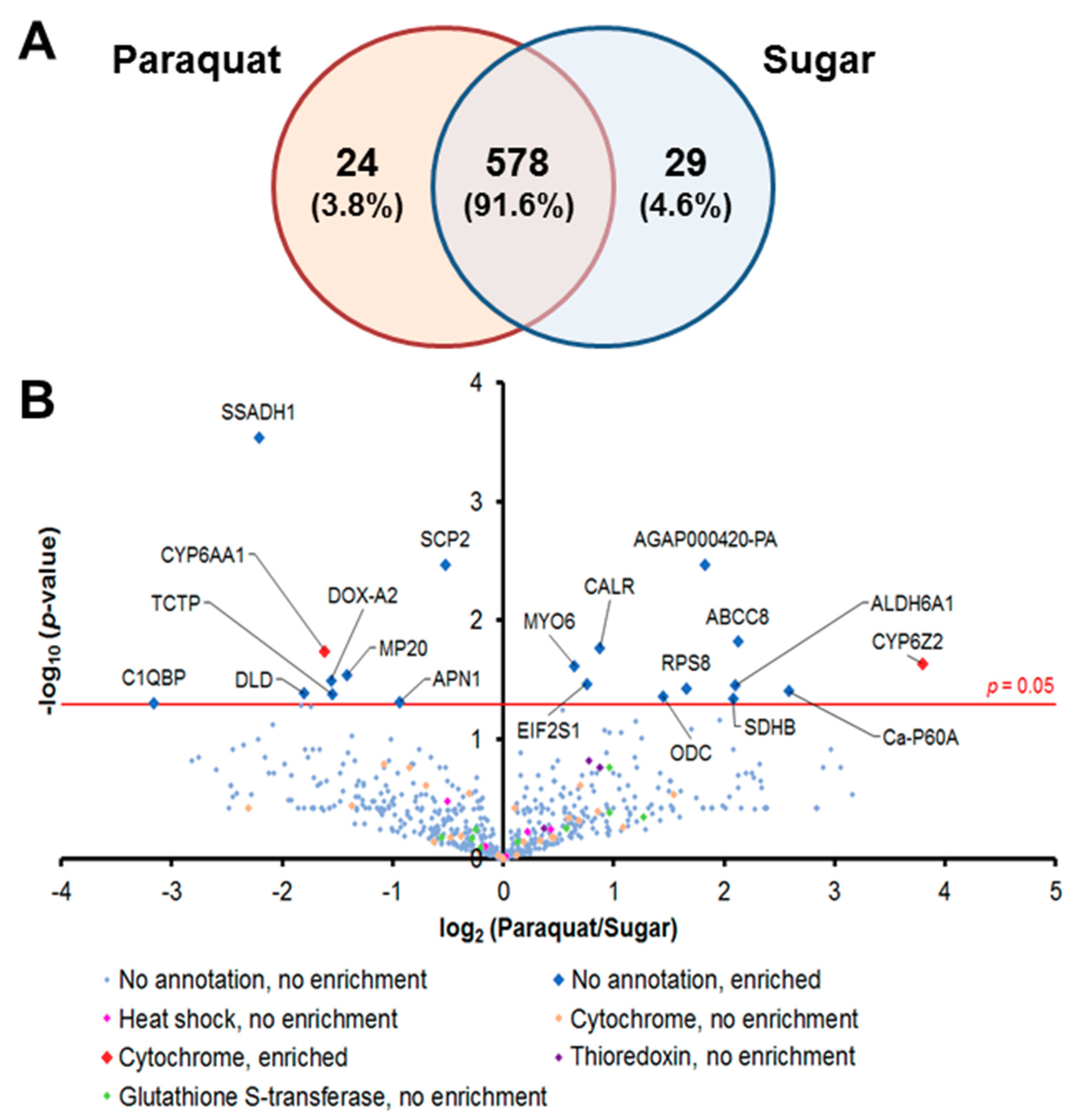
3.1. Global Proteomic Profiles of Midgut Epithelial Cells under Pqt-Induced Oxidative Stress Are Largely Conserved
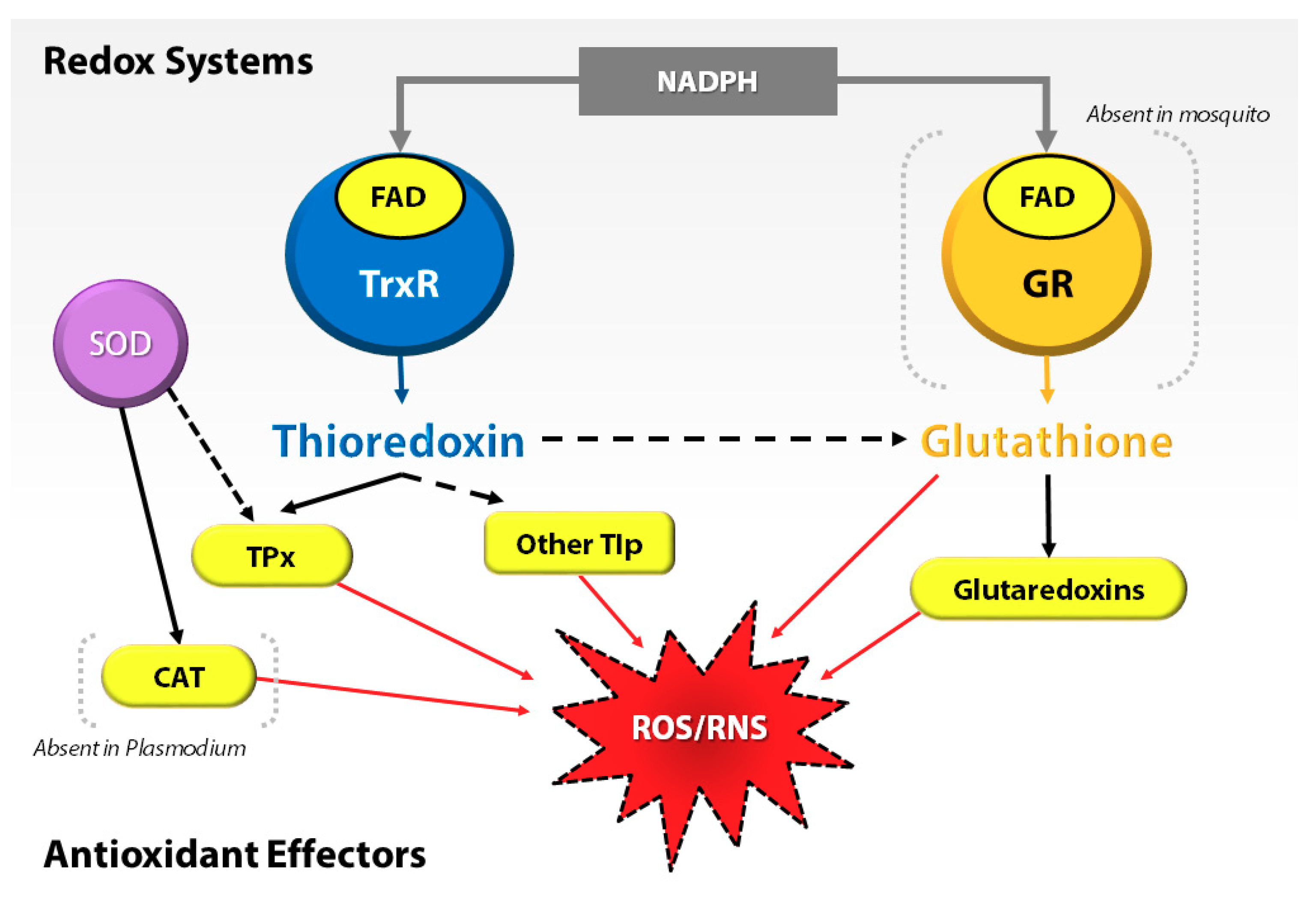
3.2. Antioxidant Proteins Are Not Involved in the Regulation of Pqt-Induced Oxidative Stress in An. Gambiae Midguts
3.3. Evidence of an Endoplasmic Reticulum (ER) Stress Response in Pqt-Treated Midguts
3.4. Proteins Involved in the Detoxification Process Are Enriched in Pqt-Treated Midguts
3.5. P. Falciparum Ookinete Invasion of An. Gambiae Midguts Does Not Upregulate Trx and GSH-Dependent Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Baton, L.A.; Ranford-Cartwright, L.C. Spreading the seeds of million-murdering death: Metamorphoses of malaria in the mosquito. Trends Parasitol. 2005, 21, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Lensen, A.H.W.; Bolmer-Van de Vegte, M.; van Gemert, G.J.; Eling, W.M.; Sauerwein, R.W. Leukocytes in a Plasmodium falciparum-infected blood meal reduce transmission of malaria to anopheles mosquitoes. Infect. Immun. 1997, 65, 3834–3837. [Google Scholar] [PubMed]
- Naotunne, T.S.; Karunaweera, N.D.; Mendis, K.N.; Carter, R. Cytokine-mediated inactivation of malarial gametocytes is dependent on the presence of white blood cells and involves reactive nitrogen intermediates. Immunology 1993, 78, 555–562. [Google Scholar] [PubMed]
- Peterson, T.M.L.; Gow, A.J.; Luckhart, S. Nitric oxide metabolites induced in Anopheles stephensi control malaria parasite infection. Free Radic. Biol. Med. 2007, 42, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Graça-Souza, A.V.; Maya-Monteiro, C.; Paiva-Silva, G.O.; Braz, G.R.C.; Paes, M.C.; Sorgine, M.H.F.; Oliveira, M.F.; Oliveira, P.L. Adaptations against heme toxicity in blood-feeding arthropods. Insect Biochem. Mol. Biol. 2006, 36, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.S.; Thompson, J.; Kafatos, F.C.; Barillas-Mury, C. Molecular interactions between Anopheles stephensi midgut cells and Plasmodium berghei: The time bomb theory of ookinete invasion of mosquitoes. EMBO J. 2000, 19, 6030–6040. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Barillas-Mury, C. Implications of time bomb model of ookinete invasion of midgut cells. Insect Biochem. Mol. Biol. 2002, 32, 1311–1316. [Google Scholar] [CrossRef]
- Kumar, S.; Gupta, L.; Han, Y.S.; Barillas-Mury, C. Inducible peroxidases mediate nitration of Anopheles midgut cells undergoing apoptosis in response to Plasmodium invasion. J. Biol. Chem. 2004, 279, 53475–53482. [Google Scholar] [CrossRef] [PubMed]
- Avery, S.V. Molecular targets of oxidative stress. Biochem. J. 2011, 434, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Vega-Rodríguez, J.; Franke-Fayard, B.; Dinglasan, R.R.; Janse, C.J.; Pastrana-Mena, R.; Waters, A.P.; Coppens, I.; Rodríguez-Orengo, J.F.; Srinivasan, P.; Jacobs-Lorena, M.; et al. The glutathione biosynthetic pathway of Plasmodium is essential for mosquito transmission. PLoS Pathog. 2009, 5, e1000302. [Google Scholar] [CrossRef]
- Pastrana-Mena, R.; Dinglasan, R.R.; Franke-Fayard, B.; Vega-Rodríguez, J.; Fuentes-Caraballo, M.; Baerga-Ortiz, A.; Coppens, I.; Jacobs-Lorena, M.; Janse, C.J.; Serrano, A.E. Glutathione reductase-null malaria parasites have normal blood stage growth but arrest during development in the mosquito. J. Biol. Chem. 2010, 285, 27045–27056. [Google Scholar] [CrossRef] [PubMed]
- Kanzok, S.M.; Fechner, A.; Bauer, H.; Ulschmid, J.K.; Müller, H.M.; Botella-Munoz, J.; Schneuwly, S.; Schirmer, R.; Becker, K. Substitution of the thioredoxin system for glutathione reductase in Drosophila melanogaster. Science 2001, 291, 643–646. [Google Scholar] [CrossRef] [PubMed]
- Peterson, T.M.L.; Luckhart, S. A Mosquito 2-cys peroxiredoxin protects against nitrosative and oxidative stresses associated with malaria parasite infection. Free Radic. Biol. Med. 2006, 40, 1067–1082. [Google Scholar] [CrossRef] [PubMed]
- Molina-Cruz, A.; DeJong, R.J.; Charles, B.; Gupta, L.; Kumar, S.; Jaramillo-Cutierrez, G.; Barillas-Mury, C. Reactive oxygen species modulate Anopheles gambiae immunity against bacteria and Plasmodium. J. Biol. Chem. 2008, 283, 3217–3223. [Google Scholar] [CrossRef] [PubMed]
- Turturice, B.A.; Lamm, M.A.; Tasch, J.J.; Zalewski, A.; Kooistra, R.; Schroeter, E.H.; Sharma, S.; Kawazu, S.-I.; Kanzok, S.M. Expression of cytosolic peroxiredoxins in Plasmodium berghei ookinetes is regulated by environmental factors in the mosquito bloodmeal. PLoS Pathog. 2013, 9, e1003136. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Molina-cruz, A.; Gupta, L.; Gupta, J.; Barillas-Mury, C. A Peroxidase/dual oxidase system modulates midgut epithelial immunity in Anopheles gambiae. Science 2010, 327, 1644–1648. [Google Scholar] [CrossRef] [PubMed]
- Cirimotich, C.M.; Dong, Y.; Clayton, A.M.; Sandiford, S.L.; Souza-Neto, J.A.; Mulenga, M.; Dimopoulos, G. Natural microbe-mediated refractoriness to Plasmodium infection in Anopheles gambiae. Science 2011, 332, 855–858. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, R.L.S.; Oliveira, J.H.M.; Oliveira, G.A.; Andersen, J.F.; Oliveira, M.F.; Oliveira, P.L.; Barillas-Mury, C. Mitochondrial reactive oxygen species modulate mosquito susceptibility to Plasmodium infection. PLoS ONE 2012. [Google Scholar] [CrossRef] [PubMed]
- Bahia, A.C.; Oliveira, J.H.M.; Kubota, M.S.; Araújo, H.R.C.; Lima, J.B.P.; Ríos-Velásquez, C.M.; Lacerda, M.V.G.; Oliveira, P.L.; Traub-Csekö, Y.M.; Pimenta, P.F.P. The role of reactive oxygen species in Anopheles aquasalis response to Plasmodium vivax infection. PLoS ONE 2013, 8, e57014. [Google Scholar] [CrossRef] [PubMed]
- Moskalyk, L.A.; Oo, M.M.; Jacobs-Lorena, M. Peritrophic matrix proteins of Anopheles gambiae and Aedes aegypti. Insect Mol. Biol. 1996, 5, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Galun, R.; Avi-Dor, Y.; Bar-Zeev, M. Feeding response in Aedes aegypti: Stimulation by adenosine triphosphate. Science 1963, 142, 1674–1675. [Google Scholar] [CrossRef] [PubMed]
- Billingsley, P.F.; Rudin, W. The role of the mosquito peritrophic membrane in bloodmeal digestion and infectivity of Plasmodium species. J. Parasitol. 1992, 78, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Drexler, A.L.; Pietri, J.E.; Pakpour, N.; Hauck, E.; Wang, B.; Glennon, E.K.K.; Georgis, M.; Riehle, M.A.; Luckhart, S. Human IGF1 regulates midgut oxidative stress and epithelial homeostasis to balance lifespan and Plasmodium falciparum resistance in Anopheles stephensi. PLoS Pathog. 2014, 10, e1004231. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Tao, D.; King, J.G.; Tweedell, R.E.; Jost, P.J.; Boddey, J.A.; Dinglasan, R.R. The acute transcriptomic and proteomic response of HC-04 hepatoma cells to hepatocyte growth factor and its implications for Plasmodium falciparum sporozoite invasion. Mol. Cell. Proteom. 2014, 13, 1153–1164. [Google Scholar] [CrossRef] [PubMed]
- Tweedell, R.; Tao, D.; Dinglasan, R.R. The cellular and proteomic response of primary and immortalized murine kupffer cells following immune stimulation diverges from that of monocyte-derived macrophages. Proteomics 2015, 15, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Tao, D.; Ubaida-Mohien, C.; Mathias, D.K.; King, J.G.; Pastrana-Mena, R.; Tripathi, A.; Goldowitz, I.; Graham, D.R.; Moss, E.; Marti, M.; et al. Sex-partitioning of the Plasmodium falciparum stage V gametocyte proteome provides insight into falciparum-specific cell biology. Mol. Cell. Proteom. 2014, 13, 2705–2724. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.F.; Paton, N.W.; Lilley, K.S.; Binz, P.-A.; Jr, R.K.J.; Jones, A.R. The minimum information about a proteomics experiment (MIAPE). Nat. Biotechnol. 2007, 25, 887–893. [Google Scholar] [PubMed]
- Vizcaíno, J.A.; Csordas, A.; Del-Toro, N.; Dianes, J.A.; Griss, J.; Lavidas, I.; Mayer, G.; Perez-Riverol, Y.; Reisinger, F.; Ternent, T.; et al. 2016 update of the PRIDE database and its related tools. Nucleic Acids Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−∆∆CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Antonio-Nkondjio, C.; Poupardin, R.; Tene, B.F.; Kopya, E.; Costantini, C.; Awono-Ambene, P.; Wondji, C.S. Investigation of mechanisms of bendiocarb resistance in Anopheles gambiae populations from the city of Yaoundé, Cameroon. Malar. J. 2016, 15, 424. [Google Scholar] [CrossRef] [PubMed]
- Sonenshine, D.E.; Bissinger, B.W.; Egekwu, N.; Donohue, K.V.; Khalil, S.M.; Roe, R.M. First transcriptome of the testis-vas deferens-male accessory gland and proteome of the spermatophore from Dermacentor variabilis (Acari: Ixodidae). PLoS ONE 2011, 6, e24711. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Kono, T.; Evans-Molina, C. Nitric oxide stress and activation of AMP-activated protein kinase impair β-cell sarcoendoplasmic reticulum calcium ATPase 2b activity and protein stability. Cell Death Dis. 2015, 6, e1790. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Evangelista, A.; Cohen, R.A. Targeting the redox regulation of SERCA in vascular physiology and disease. Curr. Opin. Pharmacol. 2010, 10, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Park, S.W.; Zhou, Y.; Lee, J.; Lee, J.; Ozcan, U. Sarco(endo)plasmic reticulum Ca2+-ATPase 2b is a major regulator of endoplasmic reticulum stress and glucose homeostasis in obesity. Proc. Natl. Acad. Sci. USA 2010, 107, 19320–19325. [Google Scholar] [CrossRef] [PubMed]
- Lafleur, M.A.; Stevens, J.L.; Lawrence, J.W. Xenobiotic perturbation of ER stress and the unfolded protein response. Toxicol. Pathol. 2013, 41, 235–262. [Google Scholar] [CrossRef] [PubMed]
- Ihara, Y.; Kageyama, K.; Kondo, T. Overexpression of calreticulin sensitizes SERCA2a to oxidative stress. Biochem. Biophys. Res. Commun. 2005, 329, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Bowes, R.C.; Van De Water, B.; Sillence, C.; Nagelkerke, J.F.; Stevens, J.L. Endoplasmic reticulum chaperones GRP78 and calreticulin prevent oxidative stress, Ca2+ disturbances, and cell death in renal epithelial cells. J. Biol. Chem. 1997, 272, 21751–21759. [Google Scholar] [CrossRef] [PubMed]
- Ruddock, L.W.; Molinari, M. N-glycan processing in ER quality control. J. Cell Sci. 2006, 119, 4373–4380. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, L.; Jin, L.; Yi, X.; Dang, E.; Yang, Y.; Li, C.; Gao, T. Oxidative stress-induced calreticulin expression and translocation: New insights into the destruction of melanocytes. J. Investig. Dermatol. 2014, 134, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, J.H.J.; Rodland, G.E.; Boe, C.A.; Haland, T.W.; Sunnerhagen, P.; Grallert, B.; Boye, E. Stress-induced inhibition of translation independently of EIF2 phosphorylation. J. Cell Sci. 2015, 128, 4420–4427. [Google Scholar] [CrossRef] [PubMed]
- Back, S.H.; Scheuner, D.; Han, J.; Song, B.; Ribick, M.; Wang, J.; Gildersleeve, R.D.; Pennathur, S.; Kaufman, R.J. Translation attenuation through EIF2α phosphorylation prevents oxidative stress and maintains the differentiated state in β cells. Cell Metab. 2009, 10, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Harding, H.P.; Zhang, Y.; Ron, D. Protein translation and folding are coupled by an endoplasmic-reticulum-resident kinase. Nature 1999, 397, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Seaman, J.A.; Alout, H.; Meyers, J.I.; Stenglein, M.D.; Dabiré, R.K.; Lozano-Fuentes, S.; Burton, T.A.; Kuklinski, W.S.; Black, W.C.; Foy, B.D. Age and prior blood feeding of Anopheles gambiae influences their susceptibility and gene expression patterns to ivermectin-containing blood meals. BMC Genom. 2015, 16, 797. [Google Scholar] [CrossRef] [PubMed]
- ter Beek, J.; Guskov, A.; Slotboom, D.J. Structural diversity of ABC transporters. J. Gen. Physiol. 2014, 143, 419–435. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.M.; George, A.M. The ABC Transporter structure and mechanism: Perspectives on recent Research. Cell. Mol. Life Sci. 2004, 61, 682–699. [Google Scholar] [CrossRef] [PubMed]
- Ackrell, B.A. Progress in understanding structure-function relationships in respiratory chain complex II. FEBS Lett 2000, 466, 1–5. [Google Scholar] [CrossRef]
- Oyedotun, K.S.; Lemire, B.D. The Quaternary structure of the Saccharomyces cerevisiae succinate dehydrogenase. homology modeling, cofactor docking, and molecular dynamics simulation studies. J. Biol. Chem. 2004, 279, 9424–9431. [Google Scholar] [CrossRef] [PubMed]
- Bauer, H.; Kanzok, S.M.; Schirmer, R.H. Thioredoxin-2 but not thioredoxin-1 is a substrate of thioredoxin peroxidase-1 from Drosophila melanogaster: Isolation and characterization of A second thioredoxin in D. melanogaster and evidence for distinct biological functions of Trx-1 and Trx-2. J. Biol. Chem. 2002, 277, 17457–17463. [Google Scholar] [CrossRef] [PubMed]
- Bauer, H.; Gromer, S.; Urbani, A.; Schnolzer, M.; Schirmer, R.H.; Muller, H.-M. Thioredoxin reductase from the malaria mosquito Anopheles gambiae. Eur. J. Biochem. 2003, 270, 4272–4281. [Google Scholar] [CrossRef] [PubMed]
- Prapanthadara, L.A.; Hemingway, J.; Ketterman, A.J. Partial purification and characterization of glutathione S-transferases involved in DDT resistance from the mosquito Anopheles gambiae. Pestic. Biochem. Physiol. 1993, 47, 119–133. [Google Scholar] [CrossRef]
- Ranson, H.; Rossiter, L.; Ortelli, F.; Jensen, B.; Wang, X.; Roth, C.W.; Collins, F.H.; Hemingway, J. Identification of a novel class of insect glutathione S-transferases involved in resistance to DDT in the malaria vector Anopheles gambiae. Biochem. J. 2001, 359, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Giordano, E.; Peluso, I.; Rendina, R.; Digilio, A.; Furia, M. The clot gene of Drosophila melanogaster encodes a conserved member of the thioredoxin-like protein superfamily. Mol. Genet. Genomics 2003, 268, 692–697. [Google Scholar] [PubMed]
- Mercer, S.W.; Burke, R. Evidence for a role for the putative drosophila HGRX1 orthologue in copper homeostasis. BioMetals 2016, 29, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Hosamani, R.; Muralidhara. Acute exposure of Drosophila melanogaster to paraquat causes oxidative stress and mitochondrial dysfunction. Arch. Insect Biochem. Physiol. 2013, 83, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Dinglasan, R.R.; Devenport, M.; Florens, L.; Johnson, J.R.; Mchugh, C.A.; Carucci, D.J.; Yates, J.R.; Jacobs-Lorena, M. The Anopheles gambiae adult midgut peritrophic matrix proteome. Insect Biochem. Mol. Biol. 2009, 39, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Brüne, B.; von Knethen, A.; Sandau, K.B. Nitric oxide and its role in apoptosis. Eur. J. Pharmacol. 1998, 351, 261–272. [Google Scholar] [CrossRef]
- Gouagna, L.C.; Mulder, B.; Noubissi, E.; Tchuinkam, T.; Verhave, J.P.; Boudin, C. The early sporogonic cycle of Plasmodium falciparum in laboratory-infected Anopheles gambiae: An estimation of parasite efficacy. Trop. Med. Int. Health 1998, 3, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Sinden, R.E.; Billingsley, P.F. Plasmodium invasion of mosquito cells: Hawk or dove? Trends Parasitol. 2001, 17, 209–212. [Google Scholar] [CrossRef]
- Whitten, M.M.A.; Shiao, S.H.; Levashina, E.A. Mosquito midguts and malaria: Cell biology, compartmentalization and immunology. Parasite Immunol. 2006, 28, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.C.; Vega-Rodríguez, J.; Jacobs-Lorena, M. The Plasmodium bottleneck: malaria parasite losses in the mosquito vector. Mem. Inst. Oswaldo Cruz 2014, 1–18. [Google Scholar] [CrossRef]
- Malhotra, J.D.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress: A vicious cycle or a double-edged sword? Antioxid. Redox Signal. 2007, 9, 2277–2293. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, J.J.M.; Brenner, M.B.; Thomas, D.Y.; Williams, D.B. Calnexin: A membrane-bound chaperone of the endoplasmic reticulum. Trends Biochem. Sci. 1994, 19, 124–128. [Google Scholar] [CrossRef]
- Helenius, A.; Trombetta, E.S.; Hebert, D.N.; Simons, J.F. Calnexin, calreticulin and the folding of glycoproteins. Trends Cell Biol. 1997, 7, 193–200. [Google Scholar]
- Miyawaki, A.; Llopis, J.; Heim, R.; JM, M.; Adams, J.A.; Ikura, M.; Tsien, R.Y. Fluorescent indicators for Ca2+ based on green fluorescent proteins and calmodulin. Nature 1997, 388, 882–887. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, D.H.; Rice, W.J.; Green, N.M. The mechanism of Ca2+ transport by sarco(endo) plasmic reticulum Ca2+-ATPases. J. Biol. Chem. 1997, 272, 28815–28818. [Google Scholar] [CrossRef] [PubMed]
- Pozzan, T.; Rizzuto, R.; Volpe, P.; Meldolesi, J. Molecular and cellular physiology of intracellular calcium stores. Physiol. Rev. 1994, 74, 595–636. [Google Scholar] [CrossRef] [PubMed]
- Ostwald, T.J.; MacLennan, D.H. Isolation of a high affinity calcium-binding protein from sarcoplasmic reticulum. J. Biol. Chem. 1974, 249, 974–979. [Google Scholar] [PubMed]
- Baksh, S.; Michalak, M. Expression of calreticulin in Escherichia coli and identification of its Ca2+ binding domains. J. Biol. Chem. 1991, 266, 21458–21465. [Google Scholar] [PubMed]
- Marva, E.; Chevion, M.; Golenser, J. The effect of free radicals induced by paraquat and copper on the in vitro development of Plasmodium falciparum. Free Radic. Res. Commun. 1991, 12–13, 137–146. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Protein Description | Fold Change | p-Value | Function |
|---|---|---|---|
| ABCC8 (AGAP008437) ATP-binding cassette transporter (ABC transporter) family C member 8 | 4.37 | 0.015 | Upregulated in bendiocarb resistance Anopheles gambiae, a detoxification gene [31] |
| ALDH6A1 (AGAP002499) Methylmalonate-semialdehyde dehydrogenase (acylating), mitochondrial | 4.28 | 0.035 | Classified as environmental and oxidative stress proteins [32] |
| Ca-P60A (AGAP006186) Calcium-transporting ATPase sarcoplasmic/endoplasmic reticulum type | 5.99 | 0.039 | Function impaired by oxidative stress [33,34,35,36] |
| CRT (AGAP004212) Calreticulin | 1.64 | 0.017 | Ca2+ homeostasis [37,38,39] and pro-apoptotic protein [40] |
| EIF2S1 (AGAP011190) Eukaryotic translation initiation factor 2 subunit alpha | 1.69 | 0.034 | Conserved in eukaryotes, the phosphorylation form of this protein serves as a signal of cell survival by attenuating the translation of mRNA [41,42,43] |
| ODC (AGAP011806) Ornithine decarboxylase | 2.72 | 0.043 | Upregulated after ivermectin-containing blood meals [44] |
| SDHB (AGAP007309) Succinate dehydrogenase (ubiquinone) iron-sulfur subunit | 4.23 | 0.045 | Ferredoxin balance system |
| Transcript/Accession ID | Function/Annotation | Response to P. Falciparum Blood Meal Ingestion. |
|---|---|---|
| Thioredoxin-1 (Trx-1; AGAP009584) | Dithiol–disulfide exchange reaction with GSSG to produce GSH [12] | None (p-value = 0.3088) |
| Thioredoxin-2 (Trx-2; AGAP007201) | Antioxidative function as electron donor to TPx [49] | None (p-value = 0.7309) |
| Thioredoxin reductase (TrxR; AGAP000565) | Key enzyme of the Trx system responsible for replenishing Trx-1 [50] | None (p-value = 0.8806) |
| Thioredoxin peroxidase-1 (TPx-1; AGAP000396) | Antioxidant enzyme that catalyzes peroxides [49] | None (p-value = 0.7976) |
| Atypical 2-Cys peroxiredoxin (Peroxiredoxin V; PrxV; AGAP001325) | Antioxidant enzyme that protects against ROS/RNS [13] | None (p-value = 0.8736) |
| Glutathione synthase (GS; AGAP000534) | Involved in the GSH biosynthesis pathway | None (p-value = 0.8515) |
| Glutathione peroxidase (GPx; AGAP004247) | Antioxidant enzyme that catalyzes peroxides [14] | None (p-value = 0.8998) |
| Glutathione S-transferase delta class 1 (GSTD1; AGAP004164) | Implicated in insecticide resistance and detoxifies xenobiotic compounds [51,52] | None (p-value = 0.9195) |
| Glutaredoxin-1 (Grx-1; AGAP011107) | Essential component of the GSH system [53,54] | None (p-value = 0.4838) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarimo, B.B.; Law, H.C.H.; Tao, D.; Pastrana-Mena, R.; Kanzok, S.M.; Buza, J.J.; Dinglasan, R.R. Paraquat-Mediated Oxidative Stress in Anopheles gambiae Mosquitoes Is Regulated by An Endoplasmic Reticulum (ER) Stress Response. Proteomes 2018, 6, 47. https://doi.org/10.3390/proteomes6040047
Tarimo BB, Law HCH, Tao D, Pastrana-Mena R, Kanzok SM, Buza JJ, Dinglasan RR. Paraquat-Mediated Oxidative Stress in Anopheles gambiae Mosquitoes Is Regulated by An Endoplasmic Reticulum (ER) Stress Response. Proteomes. 2018; 6(4):47. https://doi.org/10.3390/proteomes6040047
Chicago/Turabian StyleTarimo, Brian B., Henry Chun Hin Law, Dingyin Tao, Rebecca Pastrana-Mena, Stefan M. Kanzok, Joram J. Buza, and Rhoel R. Dinglasan. 2018. "Paraquat-Mediated Oxidative Stress in Anopheles gambiae Mosquitoes Is Regulated by An Endoplasmic Reticulum (ER) Stress Response" Proteomes 6, no. 4: 47. https://doi.org/10.3390/proteomes6040047
APA StyleTarimo, B. B., Law, H. C. H., Tao, D., Pastrana-Mena, R., Kanzok, S. M., Buza, J. J., & Dinglasan, R. R. (2018). Paraquat-Mediated Oxidative Stress in Anopheles gambiae Mosquitoes Is Regulated by An Endoplasmic Reticulum (ER) Stress Response. Proteomes, 6(4), 47. https://doi.org/10.3390/proteomes6040047