
Microbial Metalloproteomics

Abstract
:1. Introduction
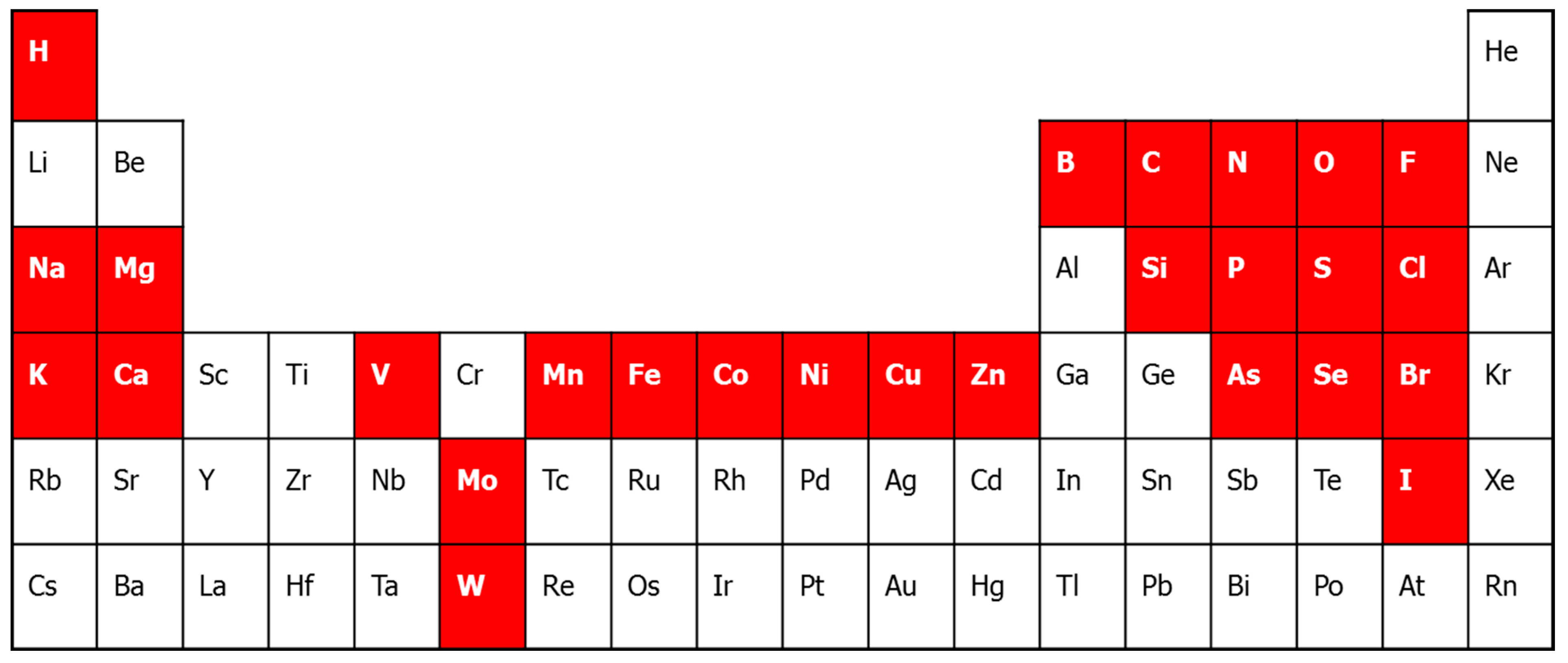
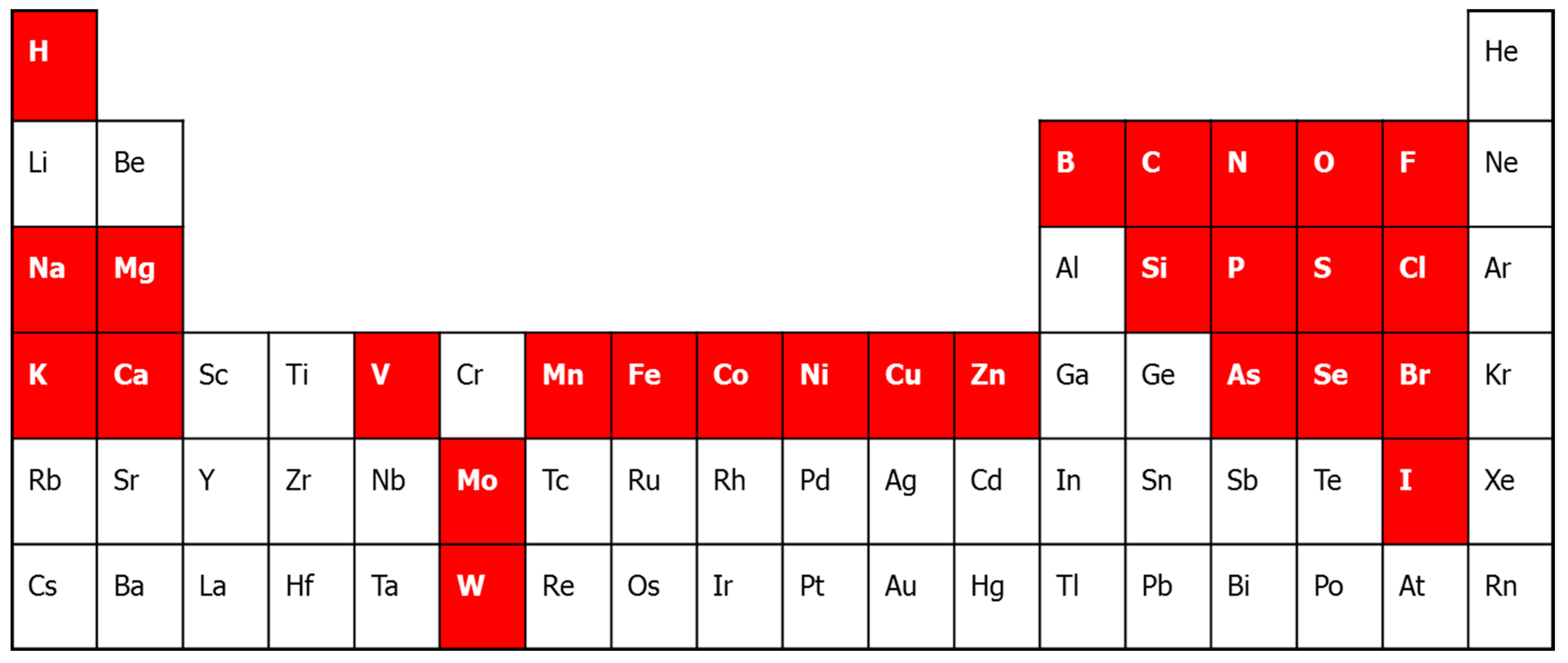
2. Metalloproteomic Approaches

2.1. ICP-MS Based Methods
2.2. X-Ray Absorption/Fluorescence Based Methods
2.3. Radionuclide Based Methods

{kind=link}
{kind=link}
{kind=link}
| Radionuclide | Half-Life Time (h) |
|---|---|
| 56Mn | 2.6 |
| 65Ni | 2.5 |
| 64Cu | 12.7 |
| 67Cu | 61.8 |
| 69Zn | 13.8 |
| 99Mo | 66.0 |
| 187W | 23.8 |
2.4. Bioinformatics Approaches
3. Microbial Metalloproteomics State of the Art
3.1. The Microbial Metalloproteome Has Been Largely Unexplored
3.2. Protein Folding Location is a Way to Select the Metal for a Particular Protein
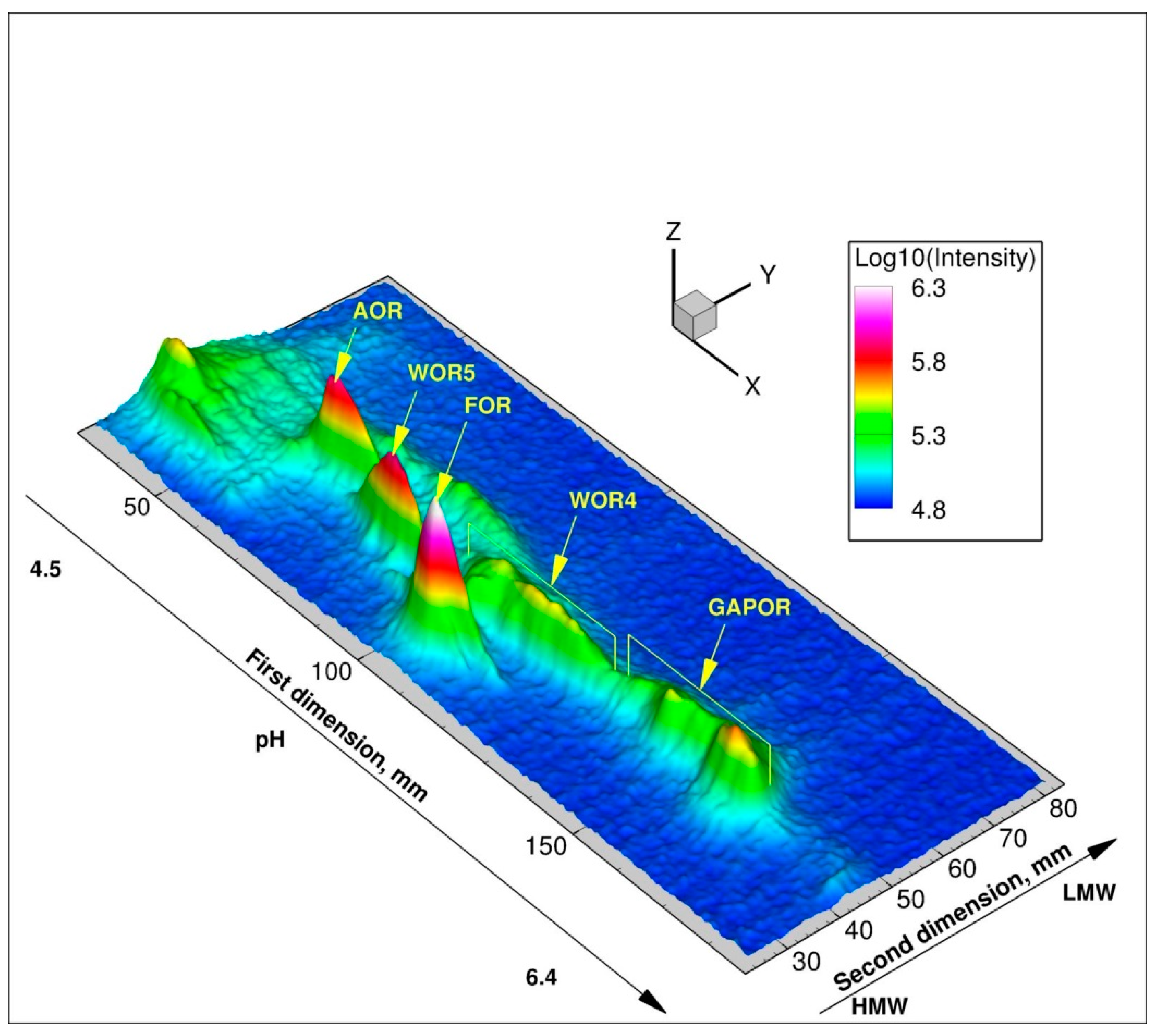
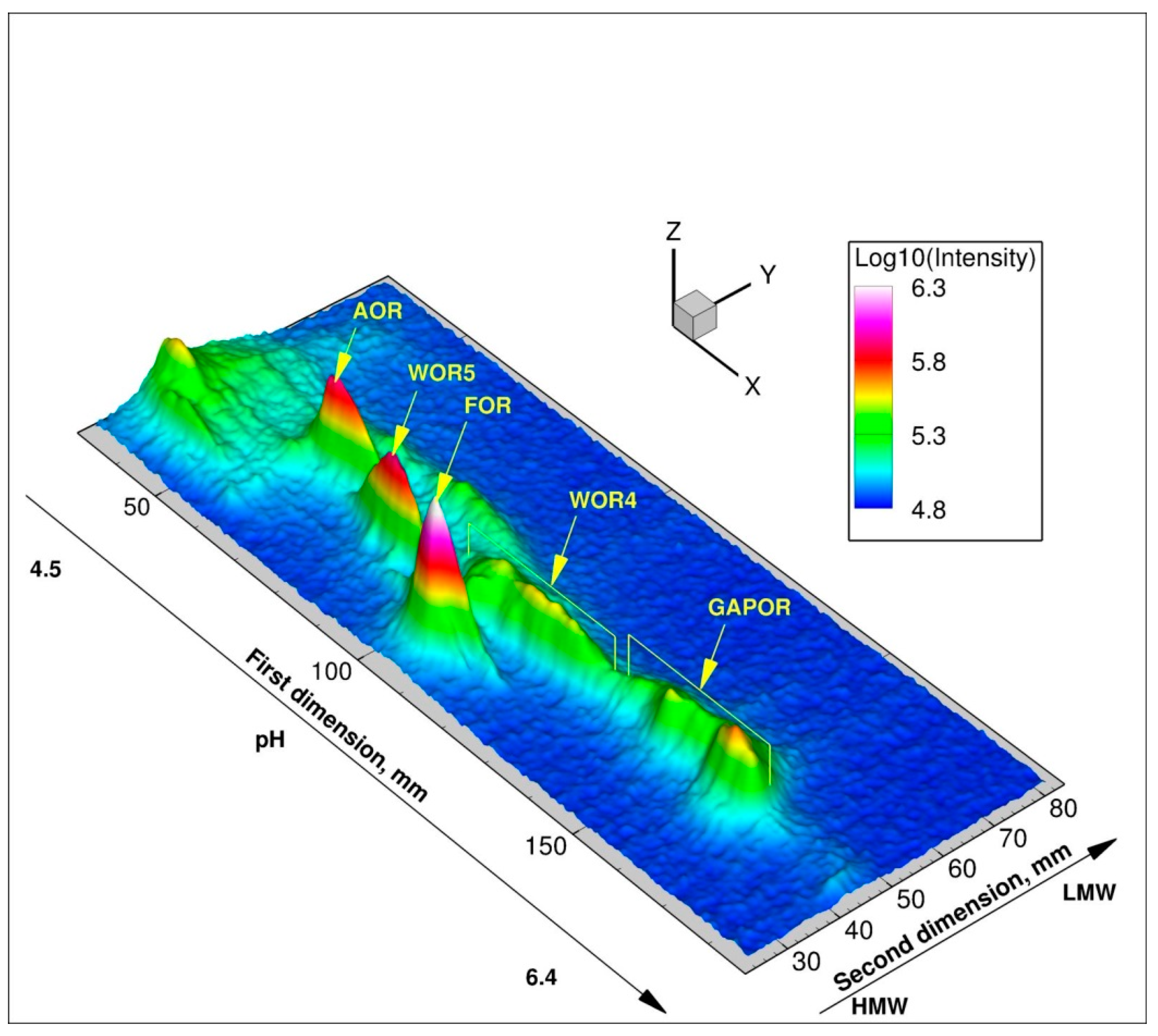
3.3. Pyrococcus Furiosus Can Select between Twin-Elements Mo and W Intracellularly
3.4. Computational Analysis of Metal Usage among the Proteomes of the Three Kingdoms of Life
3.5. Metalloproteomic Identification of Bi-binding Proteins in Helicobacter Pylori
4. Emerging Approaches and Research Areas
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Tainer, J.A.; Roberts, V.A.; Getzoff, E.D. Metal-binding sites in proteins. Curr. Opin. Biotechnol. 1991, 2, 582–591. [Google Scholar] [CrossRef]
- Kretsinger, R.H.; Uversky, V.M.; Permyakov, E.A. Encyclopedia of Metalloproteins; Springer: New York, NY, USA, 2013. [Google Scholar]
- Williams, R.J.P. Chemical selection of elements by cells. Coord. Chem. Rev. 2001, 216–217, 583–595. [Google Scholar] [CrossRef]
- Mounicou, S.; Szpunar, J.; Lobinski, R. Metallomics: The concept and methodology. Chem. Soc. Rev. 2009, 38, 1119–1138. [Google Scholar] [CrossRef] [PubMed]
- Banci, L. Metallomics and the Cell, 1st ed.; Springer: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Roberts, E.A.; Sarkar, B. Metalloproteomics: Focus on metabolic issues relating to metals. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Hare, D.J.; Rembach, A.; Roberts, B.R. The Emerging Role of Metalloproteomics in Alzheimer’s Disease Research. In Systems Biology of Alzheimer’s Disease; Castrillo, J., Oliver, S.G., Eds.; Springer: New York, NY, USA, 2015; pp. 379–389. [Google Scholar]
- Haley, K.P.; Skaar, E.P. A battle for iron: Host sequestration and Staphylococcus aureus acquisition. Microb. Infect. Inst. Pasteur 2012, 14, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Hood, M.I.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen-host interface. Nat. Rev. Microbiol. 2012, 10, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Sevcenco, A.M.; Pinkse, M.W.; Bol, E.; Krijger, G.C.; Wolterbeek, H.T.; Verhaert, P.D.E.M.; Hagedoorn, P.L.; Hagen, W.R. The tungsten metallome of Pyrococcus furiosus. Metallomics 2009, 1, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Cvetkovic, A.; Menon, A.L.; Thorgersen, M.P.; Scott, J.W.; Poole, F.L.; Jenney, F.E.J.; Lancaster, A.; Praissman, J.L.; Shanmukh, S.; Vaccaro, B.J.; et al. Microbial metalloproteomes are largely uncharacterized. Nature 2010, 466, 779–782. [Google Scholar] [CrossRef] [PubMed]
- Sevcenco, A.M.; Pinkse, M.W.H.; Wolterbeek, H.T.; Verhaert, P.D.E.M.; Hagen, W.R.; Hagedoorn, P.L. Exploring the microbial metalloproteome using MIRAGE. Metallomics 2011, 3, 1324–1330. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Peng, X.; Zhang, J.; Zhao, J.; Li, Y.; Li, Y.; Li, B.; Hu, Y.; Chai, Z. Cellular response of E. coli upon Hg2+ exposure—A case study of advanced nuclear analytical approach to metalloproteomics. Metallomics 2013, 5, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.A.; Shokes, J.E.; Cosper, N.J.; Jenney, F.E.; Adams, M.W.W. Bottlenecks and roadblocks in high-throughput XAS for structural genomics. J. Synchrotron. Rad. 2005, 12, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Barnett, J.P.; Scanlan, D.J.; Blindauer, C.A. Identification of major zinc-binding proteins from a marine cyanobacterium: Insight into metal uptake in oligotrophic environments. Metallomics 2014, 6, 1254–1268. [Google Scholar] [CrossRef] [PubMed]
- Ascone, I.; Strange, R. Biological X-ray absorption spectroscopy and metalloproteomics. J. Synchrotron. Rad. 2009, 16, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Di Bona, K.R.; Love, S.; Rhodes, N.R.; McAdory, D.; Sinha, S.H.; Kern, N.; Kent, J.; Strickland, J.; Wilson, A.; Beaird, J.; et al. Chromium is not an essential trace element for mammals: Effects of a “low-chromium” diet. J. Biol. Inorg. Chem. 2011, 16, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Sevcenco, A.M.; Krijger, G.C.; Pinkse, M.W.; Verhaert, P.D.E.M.; Hagen, W.R.; Hagedoorn, P.L. Development of a generic approach to native metalloproteomics: Application to the quantitative identification of soluble copper proteins in Escherichia coli. J. Biol. Inorg. Chem. 2009, 14, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Thierse, H.J.; Helm, S.; Pankert, P. Metalloproteomics in the Molecular Study of Cell Physiology and Disease. In 2D-PAGE: Sample Preparation and Fractionation; Posch, A., Ed.; Humana Press: Totowa, NJ, USA, 2008; pp. 139–147. [Google Scholar]
- Lobinski, R.; Moulin, C.; Ortega, R. Imaging and speciation of trace elements in biological environment. Biochimie 2006, 88, 1591–1604. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.S.; Zoriy, M.; Krause-Buchholz, U.; Becker, J.S.; Pickhardt, C.; Przybylski, M.; Pompe, W.; Rödel, G. In-gel screening of phosphorus and copper. zinc and iron in proteins of yeast mitochondria by LA-ICP-MS and identification of phosphorylated protein structures by MALDI-FT-ICR-MS after seperation with two-dimensional gel-electrophoresis. J. Anal. Atom. Spectrom. 2004, 19, 1236–1243. [Google Scholar] [CrossRef]
- Sussulini, A.; Becker, J.S. Combination of PAGE and LA-ICP-MS as an analytical workflow in metallomics: State of the art. new quantification stratagies, advantages and limitations. Metallomics 2011, 3, 1271–1279. [Google Scholar] [CrossRef] [PubMed]
- Tottey, S.; Waldron, K.J.; Firbank, S.J.; Reale, B.; Bessant, C.; Sato, K.; Cheek, T.R.; Gray, J.; Banfield, M.J.; Dennison, C.; et al. Protein-folding location can regulate manganese-binding versus copper- or zinc-binding. Nature 2008, 455, 1138–1142. [Google Scholar] [CrossRef] [PubMed]
- Katayama, A.; Tsujii, A.; Wada, A.; Nishino, T.; Ishihama, A. Systematic search for zinc-binding proteins in Escherichia coli. Eur. J. Biochem. 2002, 269, 2403–2413. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Yu, G.; Xu, Q.; Li, N.; Xiao, C.; Yin, X.; Cao, K.; Han, J.; He, Q.Y. Putative cobalt- and nickel-binding proteins and motifs in Streptococcus pneumoniae. Metallomics 2013, 5, 928–935. [Google Scholar] [CrossRef] [PubMed]
- Ge, R.; Sun, X.; Gu, Q.; Watt, R.; Tanner, J.; Wong, B.; Xia, H.; Huang, J.D.; He, Q.Y.; Sun, H. A proteomic approach for the identification of bismuth-binding proteins in Helicobacter pylori. JBIC J. Biol. Inorg. Chem. 2007, 12, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Tsang, C.N.; Bianga, J.; Sun, H.; Szpunar, J.; Lobinski, R. Probing of bismuth antiulcer drug targets in H. pylori by laser ablation-inductively coupled plasma mass spectrometry. Metallomics 2012, 4, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Xiao, C.L.; Ge, R.; Yin, X.; Li, H.; Li, N.; Yang, X.; Zhu, Y.; He, X.; He, Q.Y. Putative copper- and zinc-binding motifs in Streptococcus pneumoniae identified by immobilized metal affinity chromatography and mass spectrometry. Proteomics 2011, 11, 3288–3298. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Zhang, L.; Zhang, X.; Lu, H. Selective enrichment of metal-binding proteins based on magnetic core/shell microspheres functionalized with metal cations. Analyst 2015, 140, 4197–4205. [Google Scholar] [CrossRef] [PubMed]
- Wittig, I.; Braun, H.P.; Schägger, H. Blue native PAGE. Nat. Protoc. 2006, 1, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.; Golyshina, O.V.; Beloqui, A.; Golyshin, P.N.; Timmis, K.N. The cellular machinery of Ferroplasma acidiphilum is iron-protein-dominated. Nature 2007, 445, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Högbom, M.; Ericsson, U.B.; Lam, R.; Bakali, H.M.A.; Kuznetsova, E.; Nordlund, P.; Zamble, D.B. A high throughput method for the detection of metalloproteins on a microgram scale. Mol. Cell Proteomics 2005, 4, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Maass, S.; Sievers, S.; Zühlke, D.; Kuzinski, J.; Sappa, P.K.; Muntel, J.; Hessling, B.; Bernhardt, J.; Sietmann, R.; Völker, U.; et al. Efficient. Global-Scale Quantification of Absolute Protein Amounts by Integration of Targeted Mass Spectrometry and Two-Dimensional Gel-Based Proteomics. Anal. Chem. 2011, 83, 2677–2684. [Google Scholar] [CrossRef] [PubMed]
- Bronstrup, M. Absolute quantification strategies in proteomics based on mass spectrometry. Expert Rev. Proteomics 2004, 1, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Ishihama, Y.; Oda, Y.; Tabata, T.; Sato, T.; Nagasu, T.; Rappsilber, J.; Mann, M. Exponentially Modified Protein Abundance Index (emPAI) for Estimation of Absolute Protein Amount in Proteomics by the Number of Sequenced Peptides per Protein. Mol. Cell Proteomics 2005, 4, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Chahrour, O.; Cobice, D.; Malone, J. Stable isotope labelling methods in mass spectrometry-based quantitative proteomics. J. Pharm. Biomed. Anal. 2015, 113, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Noble, J.E.; Bailey, M.J.A. Quantitation of Protein. In Methods in Enzymology; Richard, R., Murray, P.D., Eds.; Elsevier: New York, NY, USA, 2009; Chapter 8; pp. 73–95. [Google Scholar]
- Ahrends, R.; Pieper, S.; Kuhn, A.; Weisshoff, H.; Hamester, M.; Lindemann, T.; Scheler, C.; Lehmann, K.; Taubner, K.; Linscheid, M.W. A metal-coded affinity tag approach to quantitative proteomics. Mol. Cell Proteomics 2007, 6, 1907–1916. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.S.; Matusch, A.; Palm, C.; Salber, D.; Morton, K.A.; Becker, J.S. Bioimaging of metals in brain tissue by laser ablation inductively coupled plasma mass spectrometry (LA-ICP-MS) and metallomics. Metallomics 2010, 2, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.S.; Zoriy, M.V.; Pickhardt, C.; Palomero-Gallagher, N.; Zilles, K. Imaging of Copper. Zinc and Other Elements in Thin Section of Human Brain Samples (Hippocampus) by Laser Ablation Inductively Coupled Plasma Mass Spectrometry. Anal. Chem. 2005, 77, 3208–3216. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Chance, M.R. Metalloproteomics: Forward and reverse approaches in metalloprotein structural and functional characterization. Curr. Opin. Chem. Biol. 2011, 15, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Zhan, C.; Ignatov, A.; Manjasetty, B.A.; Marinkovic, N.; Sullivan, M.; Huang, R.; Chance, M.R. Metalloproteomics: High-Througput Structural and Functional Annotation of Proteins in Structural Genomics. Structure 2005, 13, 1473–1486. [Google Scholar] [CrossRef] [PubMed]
- McRae, R.; Bagchi, P.; Sumalekshmy, S.; Fahrni, C.J. In Situ Imaging of Metals in Cells and Tissues. Chem. Rev. 2009, 109, 4780–4827. [Google Scholar] [CrossRef] [PubMed]
- Jenney, F.E.; Brereton, P.S.; Izumi, M.; Poole, F.L.; Shah, C.; Sugar, F.J.; Lee, H.S.; Adams, M.W.W. High-throughput production of Pyrococcus furiosus proteins: Considerations for metalloproteins. J. Synchrotron. Rad. 2005, 12, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Bertini, I.; Rosato, A. Metalloproteomes: A Bioinformatic Approach. Acc. Chem. Res. 2009, 42, 1471–1479. [Google Scholar] [CrossRef] [PubMed]
- Estellon, J.; de Ollagnier Choudens, S.; Smadja, M.; Fontecave, M.; Vandenbrouck, Y. An integrative computational model for large-scale identification of metalloproteins in microbial genomes: A focus on iron-sulfur cluster proteins. Metallomics 2014, 6, 1913–1930. [Google Scholar] [CrossRef] [PubMed]
- Azia, A.; Levy, R.; Unger, R.; Edelman, M.; Sobolev, V. Genome-wide computational determination of the human metalloproteome. Proteins 2015, 83, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.H.; Lin, Y.F.; Lin, J.J.; Yu, C.S. Prediction of metal ion-binding sites in proteins using the fragment transformation method. PLoS ONE 2012, 7, e39252. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Bertini, I.; Cavallaro, G.; Holliday, G.L.; Thornton, J.M. Metal ions in biological catalysis: From enzyme databases to general principles. J. Biol. Inorg. Chem. 2008, 13, 1205–1218. [Google Scholar] [CrossRef] [PubMed]
- Metal MACiE: The database of catalytic metal ions. Available online: https://www.ebi.ac.uk/thornton-srv/databases/Metal_MACiE/homehtml (accessed on 30 November 2015).
- Holliday, G.L.; Andreini, C.; Fischer, J.D.; Rahman, S.A.; Almonacid, D.E.; Williams, S.T.; Pearson, W.R. MACiE: Exploring the diversity of biochemical reactions. Nucleic Acids Res. 2011, 40, D783–D789. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Cavallaro, G.; Lorenzini, S.; Rosato, A. MetalPDB: A database of metal sites in biological macromolecular structures. Nucleic Acids Res. 2013, 41, D312–D319. [Google Scholar] [CrossRef] [PubMed]
- MetalPDB. Available online: http://metalweb.cerm.unifi.it/ (accessed on 30 November 2015).
- Falia, G.; Stetter, K.O. Pyrococcus furiosus sp. nov. represents a Novel Genus of Marine Heterotrophic Archaeabacteria growing Optimally at 100 °C. Arch. Microbiol. 1986, 145, 56–61. [Google Scholar] [CrossRef]
- Bevers, L.E.; Bol, E.; Hagedoorn, P.L.; Hagen, W.R. WOR5: A Novel Tungsten Containing Aldehyde Oxidoreductase from Pyrococcus furiosus with a Broad Substrate Specificity. J. Bacteriol. 2005, 187, 7056–7061. [Google Scholar] [CrossRef] [PubMed]
- Bevers, L.E.; Hagedoorn, P.L.; Hagen, W.R. The bioinorganic chemistry of tungsten. Coord. Chem. Rev. 2009, 253, 269–290. [Google Scholar] [CrossRef]
- Kletzin, A.; Adams, M.W.W. Tungsten in Biological Systems. FEMS Microbiol. Rev. 1996, 18, 5–63. [Google Scholar] [CrossRef] [PubMed]
- Sevcenco, A.M.; Bevers, L.E.; Krijger, G.C.; Pinkse, M.W.H.; Wolterbeek, H.T.; Verhaert, P.D.E.M.; Hagen, W.R.; Hagedoorn, P.L. Molybdenum incorporation in tungsten aldehyde oxidoreductase enzymes from Pyrococcus furiosus. J. Bacteriol. 2010, 192, 4143–4152. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Banci, L.; Bertini, I.; Elmi, S.; Rosato, A. Non-heme iron through the three domains of life. Proteins 2007, 67, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Zinc through the Three Domains of Life. J. Proteome Res. 2006, 5, 3173–3178. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, H.; Li, H.; Sun, H. Metallomic and metalloproteomic strategies in elucidating the molecular mechanisms of metallodrugs. Dalton Trans. 2015, 44, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Cheng, T.; He, B.; Li, L.; Wang, Y.; Lai, Y.T.; Jiang, G.; Sun, H. Identification of Metal-Associated Proteins in Cells by Using Continuous-Flow Gel Electrophoresis and Inductively Coupled Plasma Mass Spectrometry. Angew. Chem. Int. Ed. 2013, 52, 4916–4920. [Google Scholar] [CrossRef] [PubMed]
- Duhutrel, P.; Bordat, C.; Wu, T.D.; Zagorec, M.; Guerquin-Kern, J.L.; Champomier-Vergès, M.C. Iron Sources Used by the Nonpathogenic Lactic Acid Bacterium Lactobacillus sakei as Revealed by Electron Energy Loss Spectroscopy and Secondary-Ion Mass Spectrometry. Appl. Environ. Microbiol. 2010, 76, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Sousa, T.; Chung, A.P.; Pereira, A.; Piedade, A.P.; Morais, P.V. Aerobic uranium immobilization by Rhodanobacter A2-61 through formation of intracellular uranium-phosphate complexes. Metallomics 2013, 5, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Carter, K.P.; Young, A.M.; Palmer, A.M. Fluorescent Sensors for Measuring Metal Ions in Living Systems. Chem. Rev. 2014, 114, 4564–4601. [Google Scholar] [CrossRef] [PubMed]
- Hessels, A.M.; Merkx, M. Genetically-encoded FRET-based sensors for monitoring Zn2+ in living cells. Metallomics 2015, 7, 258–266. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hagedoorn, P.-L. Microbial Metalloproteomics. Proteomes 2015, 3, 424-439. https://doi.org/10.3390/proteomes3040424
Hagedoorn P-L. Microbial Metalloproteomics. Proteomes. 2015; 3(4):424-439. https://doi.org/10.3390/proteomes3040424
Chicago/Turabian StyleHagedoorn, Peter-Leon. 2015. "Microbial Metalloproteomics" Proteomes 3, no. 4: 424-439. https://doi.org/10.3390/proteomes3040424
APA StyleHagedoorn, P.-L. (2015). Microbial Metalloproteomics. Proteomes, 3(4), 424-439. https://doi.org/10.3390/proteomes3040424