Potential Transient Response of Terrestrial Vegetation and Carbon in Northern North America from Climate Change

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Model
2.2. Climate Data
3. Results
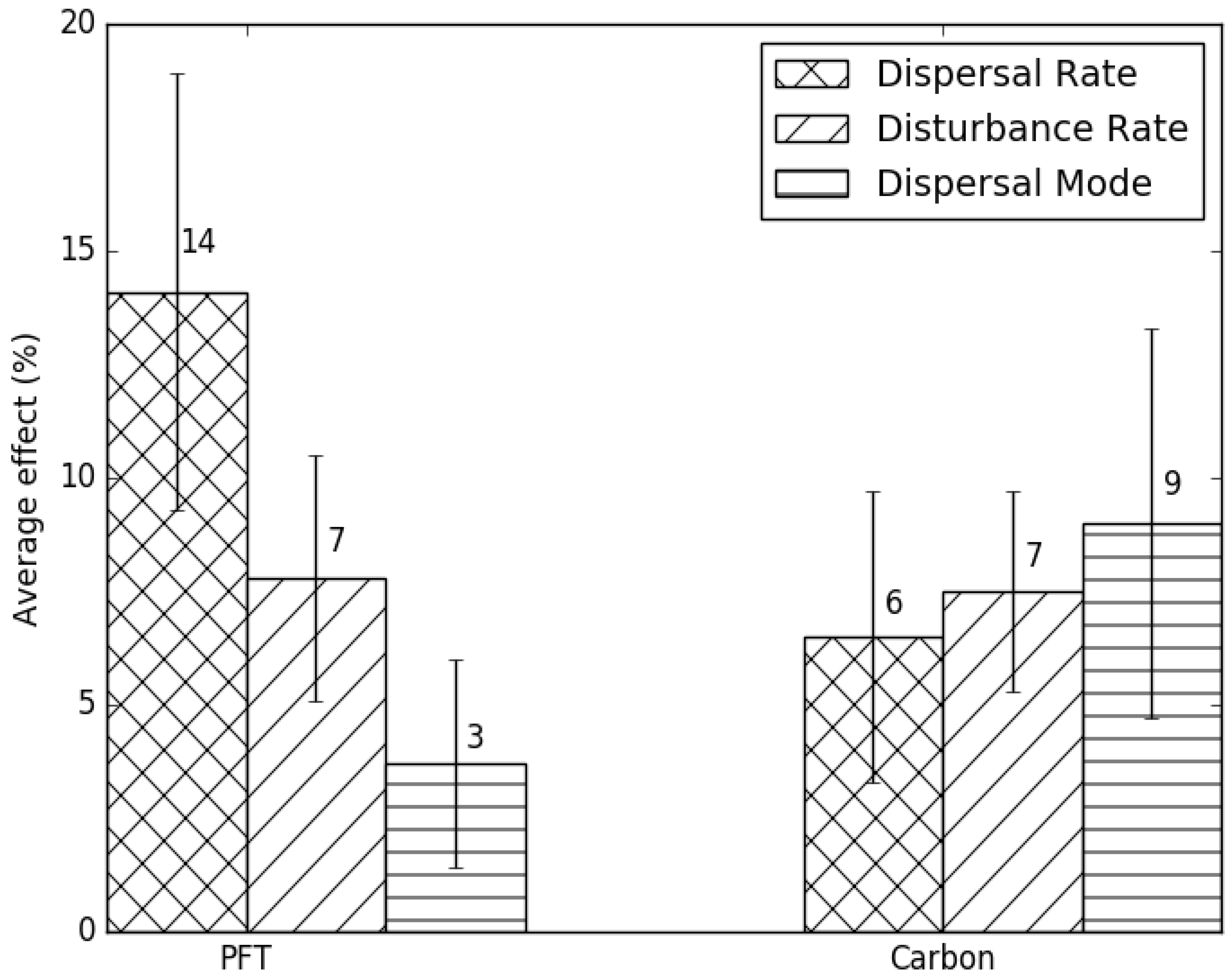
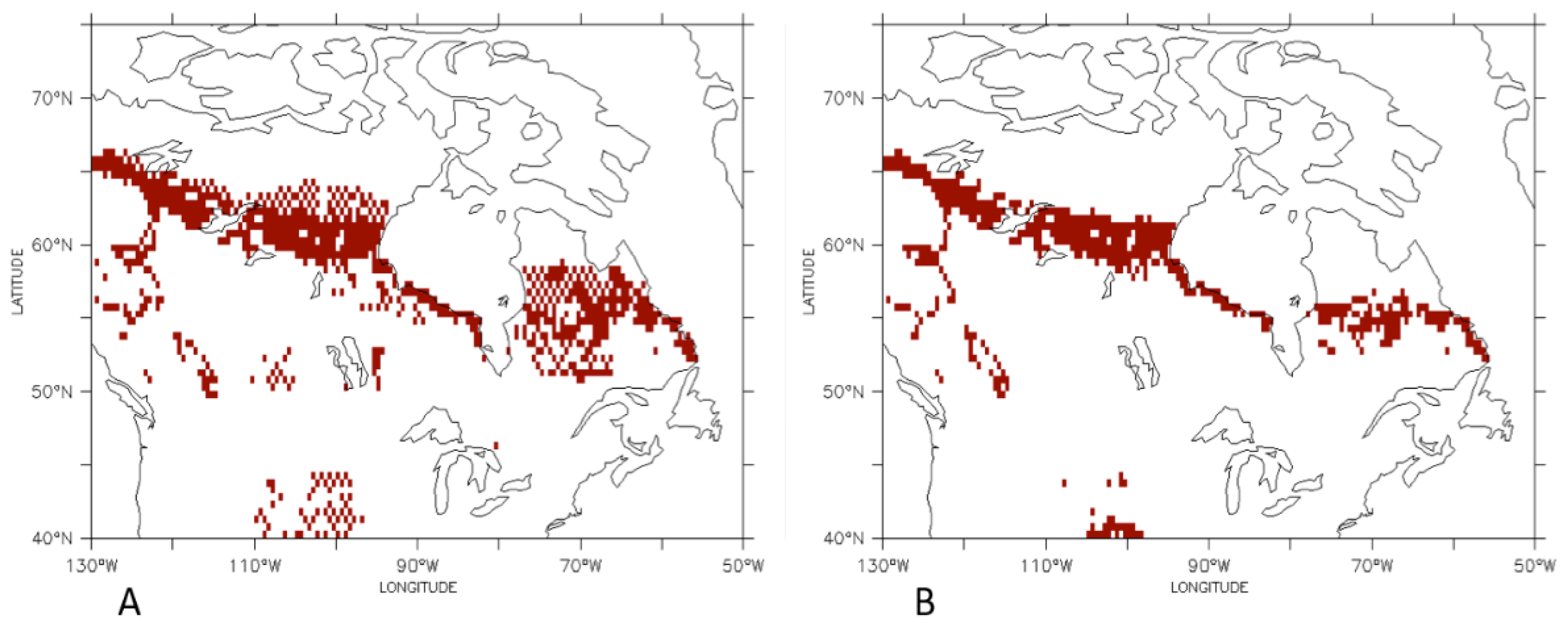
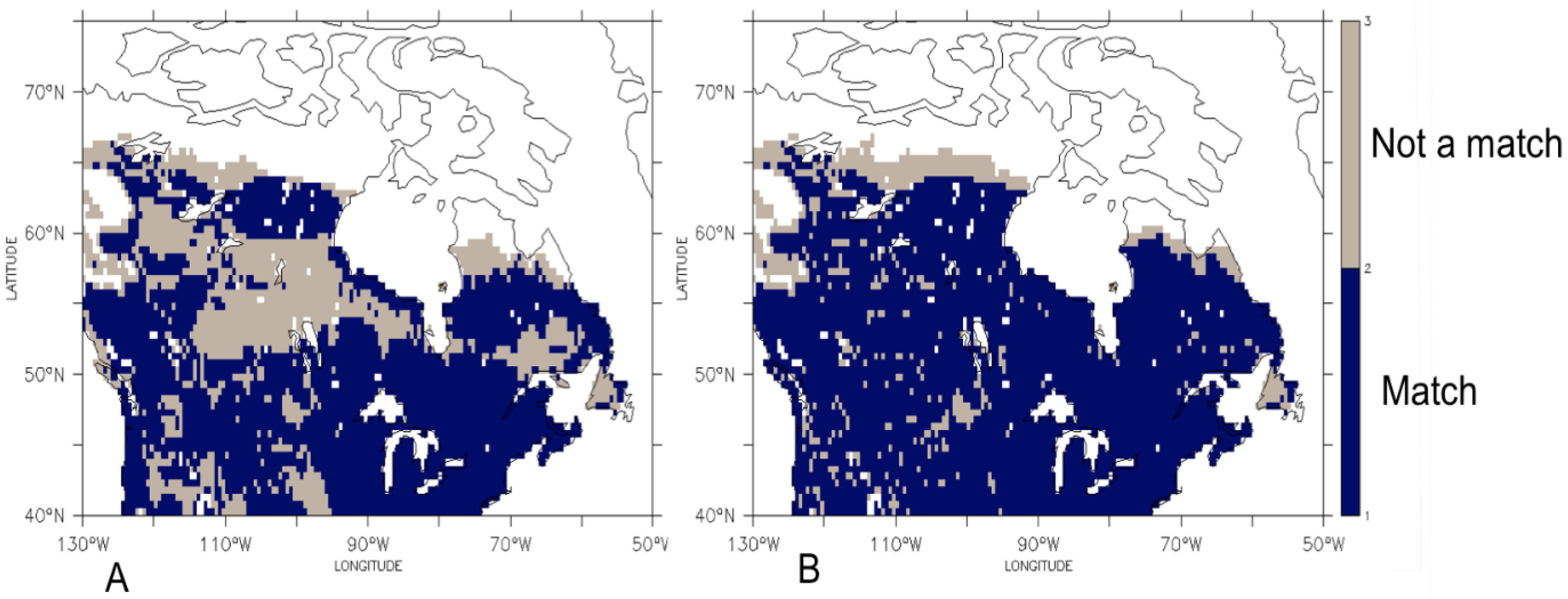
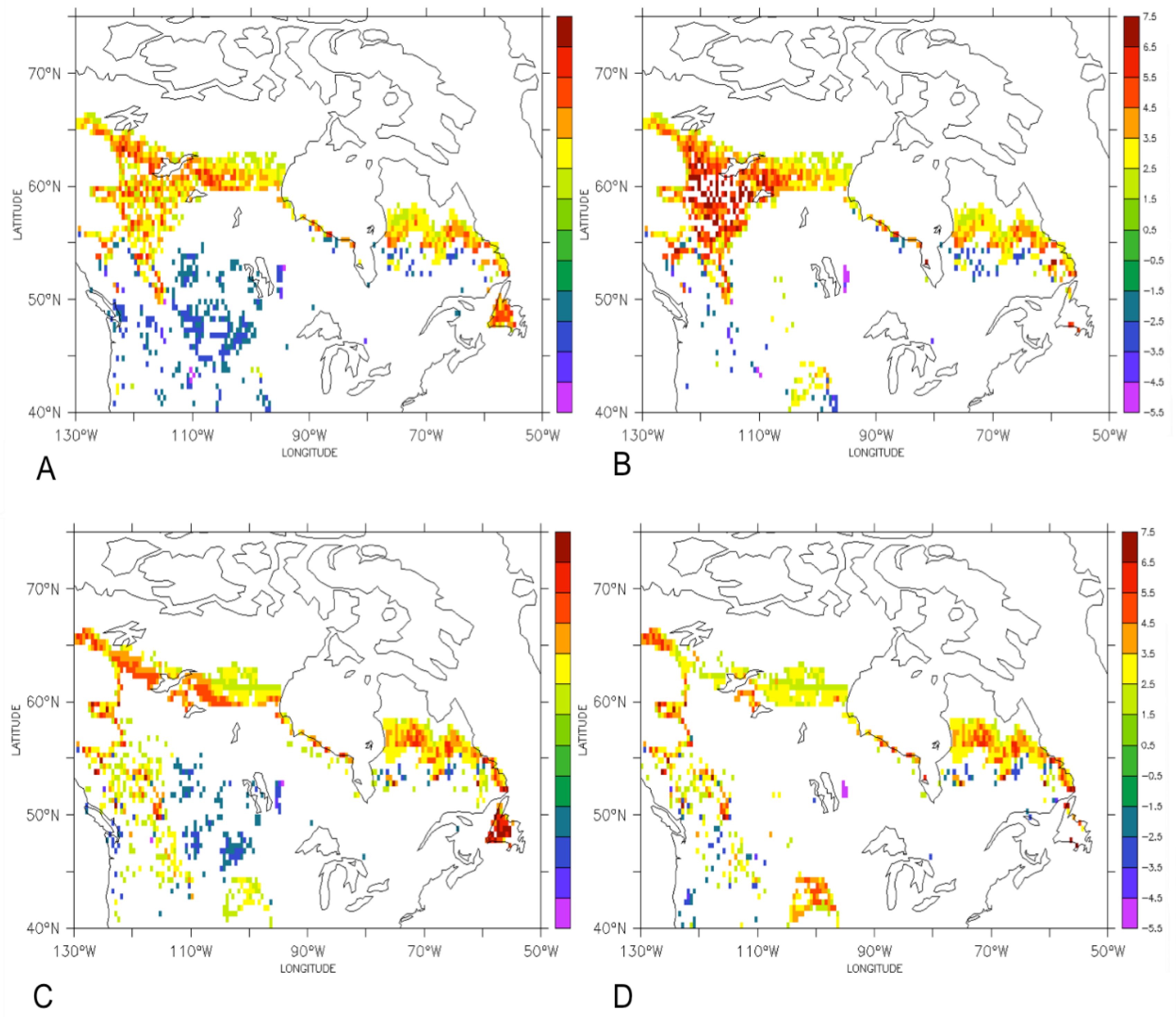
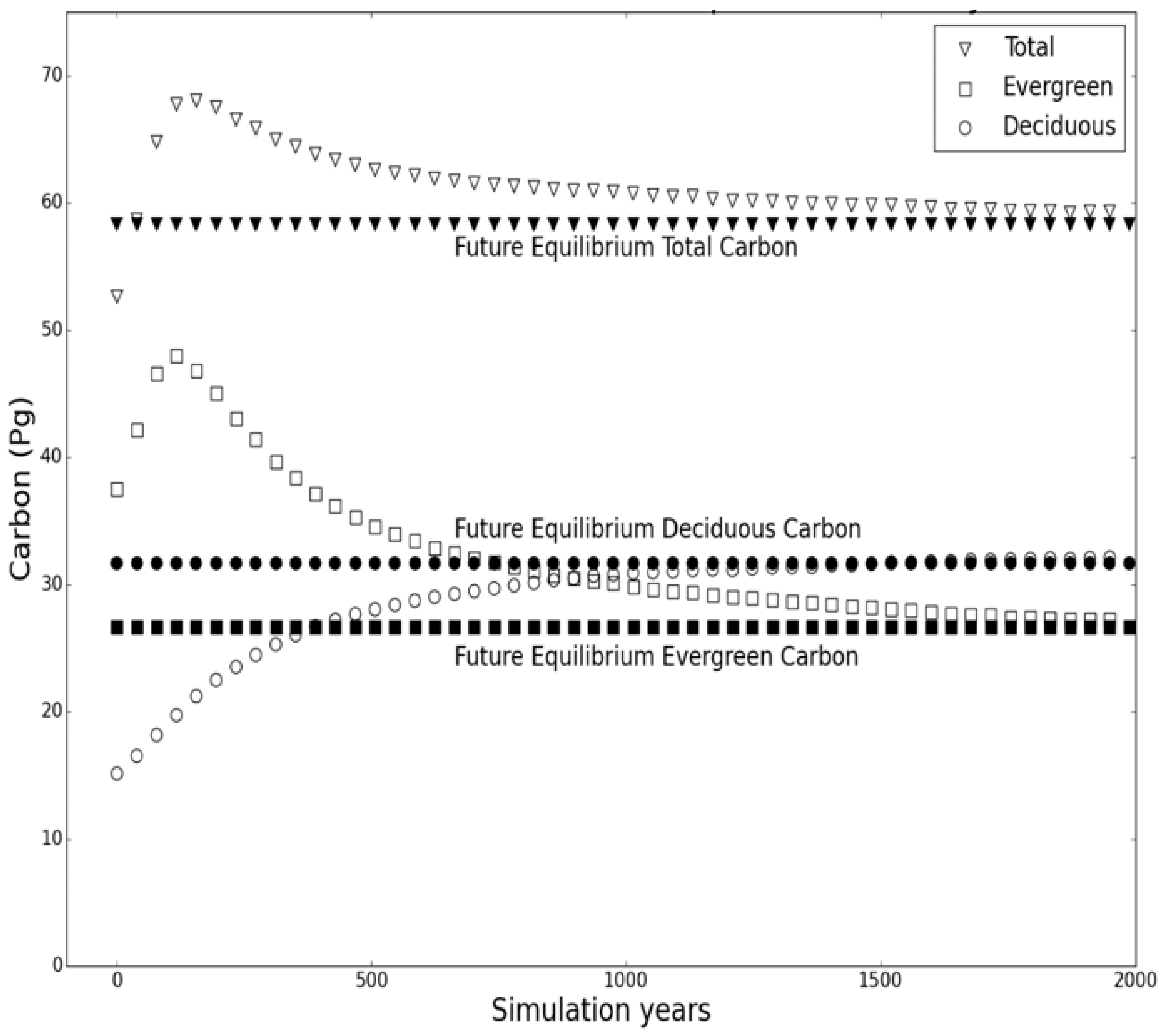
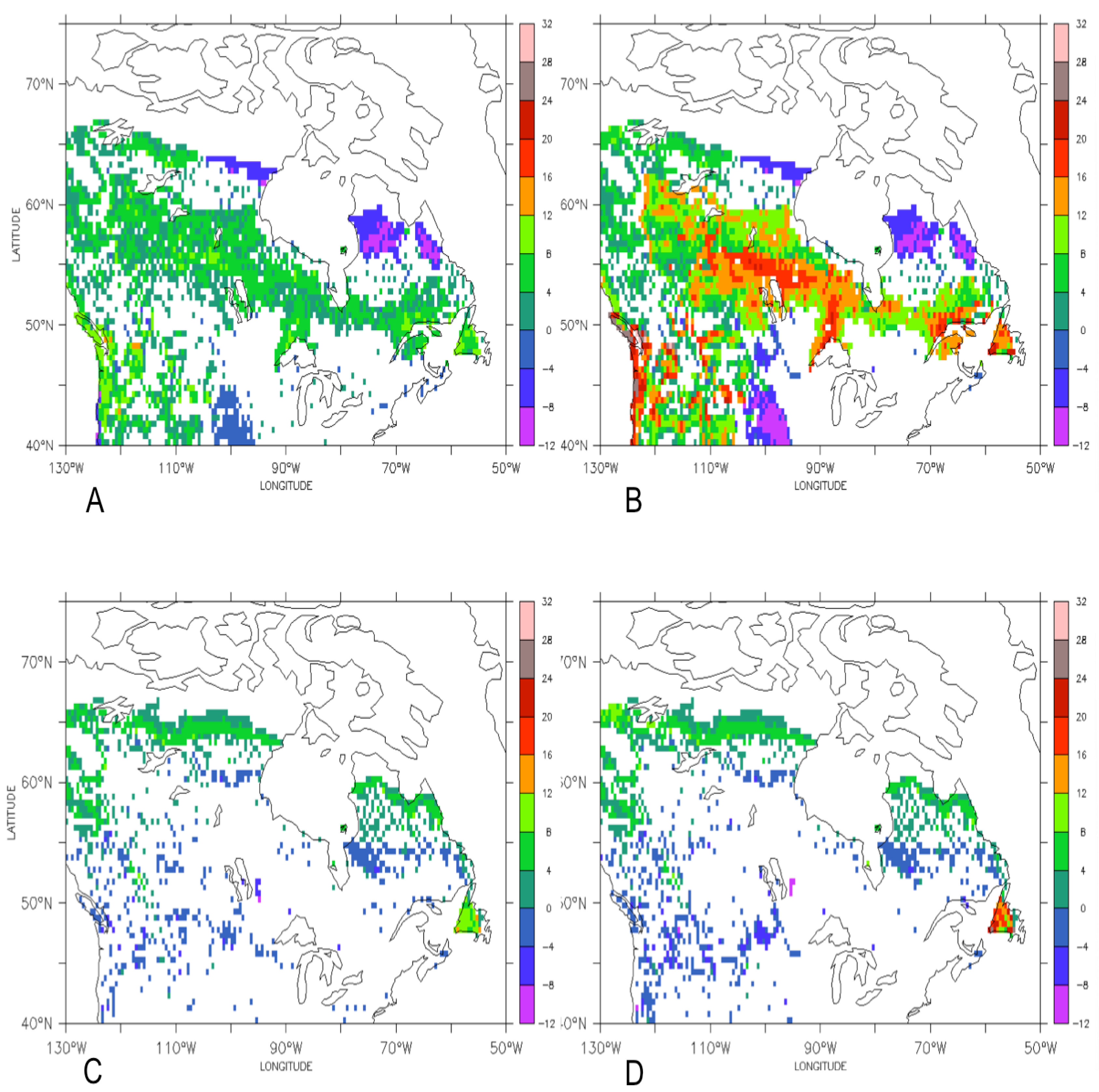
3.1. Comparison of the Transient Response of Migration to the Equilibrium Response
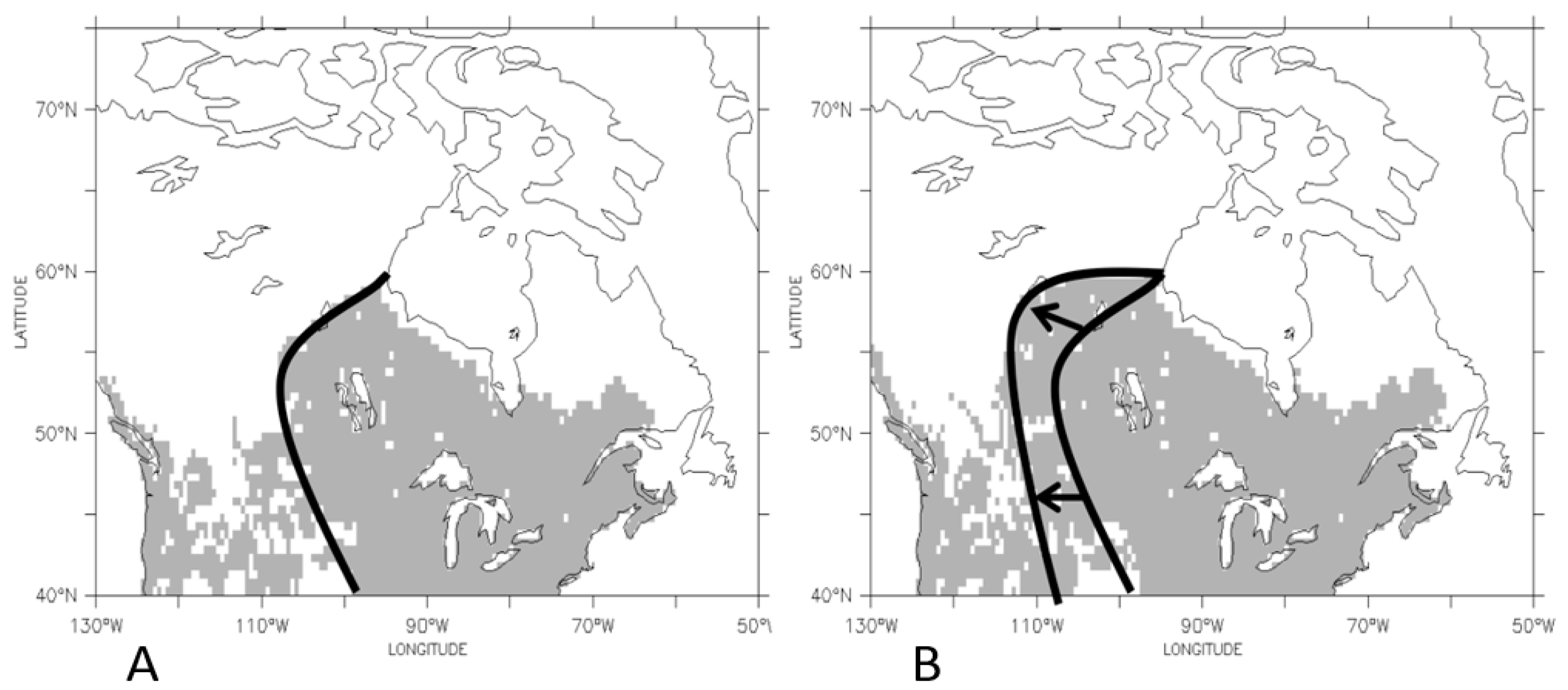
3.2. Temporal Patterns of Migration
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Houghton, R.A. Aboveground Forest Biomass and the Global Carbon Balance. Glob. Chang. Biol. 2005, 11, 945–958. [Google Scholar] [CrossRef]
- Goodale, C.L.; Apps, M.J.; Birdsey, R.A.; Field, C.B.; Heath, L.S.; Houghton, R.A.; Jenkins, J.C.; Kohlmaier, G.H.; Kurz, W.; Liu, S.; et al. Forest Carbon Sink in the Northern Hemisphere. Ecol. Appl. 2002, 12, 891–899. [Google Scholar] [CrossRef]
- Koppen, W. Versuch einer Klassifikation der Klimate, vorzugsweise nach ihren Beziehungen zur Pflanzenwelt. Geogr. Z. 1900, 12, 657–679. [Google Scholar]
- Thornthwaite, C.W. The Climates of North America: According to a New Classification. Geogr. Rev. 1931, 21, 633–655. [Google Scholar] [CrossRef]
- Thornthwaite, C.W. American Geographical Society an Approach toward a Rational Classification of Climate. Source Geogr. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]
- Holdridge, L.R. Determination of world plant formation from simple climate data. Science 1947, 105, 367–368. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.B. Climatic Changes in Southern Connecticut Recorded by Pollen Deposition at Rogers Lake. Ecology 1969, 3, 409–422. [Google Scholar] [CrossRef]
- Davis, M.B.; Botkin, D.B. Sensitivity of cool-temperate forests and their fossil pollen record to rapid temperature change. Quat. Res. 1985, 3, 327–340. [Google Scholar] [CrossRef]
- Skellam, J.G. Random dispersal in theoretical populations. Biometrika 1951, 38, 196–218. [Google Scholar] [CrossRef]
- Davis, M.B.; Shaw, R.G. Range shifts and adaptive responses to Quaternary climate change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef]
- Lischke, H.; Lotter, A.F.; Fischlin, A. Untangling a Holocene pollen record with forest model simulations and independent climate data. Ecol. Model. 2002, 150, 1–21. [Google Scholar] [CrossRef][Green Version]
- Clark, J.S.; Fastie, C.; Hurtt, G.; Jackson, S.T.; Johnson, C.; King, A.; Lewis, M.; Lynch, J.; Prentice, C.; Wyckoff, P.; et al. Reid’s Paradox of Plant Rapid Migration Dispersal theory and interpretation of paleoecological records. Bioscience 1998, 48, 13–24. [Google Scholar] [CrossRef]
- Clark, J.S. Why trees migrate so fast: Confronting theory with dispersal biology and the paleorecord. Am. Nat. 1998, 152, 204–224. [Google Scholar] [CrossRef] [PubMed]
- Phillips, B.L.; Brown, G.P.; Travis, J.M.J.; Shine, R. Reid’s paradox revisited: The evolution of dispersal kernels during range expansion. Am. Nat. 2008, 172, S34–S48. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A.; Koven, C.D.; Anderegg, W.R.L.; Christoffersen, B.O.; Dietze, M.C.; Farrior, C.E.; Holm, J.A.; Hurtt, G.C.; Knox, R.G.; Lawrence, P.J.; et al. Vegetation demographics in Earth System Models: A review of progress and priorities. Glob. Chang. Biol. 2018, 24, 35–54. [Google Scholar] [CrossRef] [PubMed]
- Sitch, S.; Huntingford, C.; Gedney, N.; Levy, P.E.; Lomas, M.; Piao, S.L.; Betts, R.; Cias, P.; Cox, P.; Friendlingstein, P.; et al. Evaluation of the terrestrial carbon cycle, future plant geography and climate-carbon cycle feedbacks using five Dynamic Global Vegetation Models (DGVMs). Glob. Chang. Biol. 2008, 14, 2015–2039. [Google Scholar] [CrossRef]
- Cox, P.M. Description of the TRIFFID Dynamic Global Vegetation Model; Met Office: Exeter, UK, 2001. [Google Scholar]
- Woodward, F.I.; Smith, T.M.; Emanuel, W.R. A global land primary productivity and phytogeography model. Glob. Biogeochem. Cycles 1995, 9, 471–490. [Google Scholar] [CrossRef]
- Woodward, F.I.; Lomas, M.R. Vegetation dynamics—Simulating responses to climatic change. Biol. Rev. 2004, 79, 643–670. [Google Scholar] [CrossRef]
- Krinner, G.; Viovy, N.; de Noblet-Ducoudré, N.; Ogée, J.; Polcher, J.; Friedlingstein, P.; Ciais, P.; Sitch, S.; Prentice, I.C. A dynamic global vegetation model for studies of the coupled atmosphere-biosphere system. Glob. Biogeochem. Cycles 2005, 19, 19. [Google Scholar] [CrossRef]
- Sitch, S.; Smith, B.; Prentice, I.C.; Arneth, A.; Bondeau, A.; Cramer, W.; Kaplan, J.O.; Levis, S.; Lucht, W.; Sykes, M.T.; et al. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model. Glob. Chang. Biol. 2003, 9, 161–185. [Google Scholar] [CrossRef]
- Sato, H.; Itoh, A.; Kohyama, T. SEIB–DGVM: A new Dynamic Global Vegetation Model using a spatially explicit individual-based approach. Ecol. Model. 2007, 200, 279–307. [Google Scholar] [CrossRef]
- Sato, H.; Ise, T. Effect of plant dynamic processes on African vegetation responses to climate change: Analysis using the spatially explicit individual-based dynamic global vegetation model (SEIB-DGVM). J. Geophys. Res. Biogeosci. 2012, 117, G03017. [Google Scholar] [CrossRef]
- Snell, R.S. Simulating long-distance seed dispersal in a dynamic vegetation model. Glob. Ecol. Biogeogr. 2014, 23, 89–98. [Google Scholar] [CrossRef]
- Snell, R.S.; Huth, A.; Nabel, J.E.M.S.; Bocedi, G.; Travis, J.M.J.; Gravel, D.; Bugmann, H.; Gutiérrez, A.G.; Hickler, T.; Higgins, S.I.; et al. Using dynamic vegetation models to simulate plant range shifts. Ecography 2014, 37, 1184–1197. [Google Scholar] [CrossRef]
- Lischke, H.; Zimmermann, N.E.; Bolliger, J.; Rickebusch, S.; Löffler, T.J. TreeMig: A forest-landscape model for simulating spatio-temporal patterns from stand to landscape scale. Ecol. Model. 2006, 199, 409–420. [Google Scholar] [CrossRef]
- Jin, W.; He, H.S.; Thompson, F.R., III; Wang, W.J.; Fraser, J.S.; Shifley, S.R.; Hanberry, B.B.; Dijak, W.D. Future forest aboveground carbon dynamics in the central United States: The importance of forest demographic processes. Sci. Rep. 2017, 7, 41821. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.J.; He, H.S.; Thompson, F.R.; Fraser, J.S.; Dijak, W.D. Changes in forest biomass and tree species distribution under climate change in the northeastern United States. Landsc. Ecol. 2017, 32, 1399–1413. [Google Scholar] [CrossRef]
- Hurtt, G.C.; Moorcroft, P.R.; Pacala, S.W.; Levin, S.A. Terrestrial models and global change: Challenges for the future. Glob. Chang. Biol. 1998, 4, 581–590. [Google Scholar] [CrossRef]
- Moorcroft, P.R.; Hurtt, G.C.; Pacala, S.W. A Method for Scaling Vegetation Dynamics: The Ecosystem Demography Model (ED). Ecol. Monogr. 2001, 71, 557–586. [Google Scholar] [CrossRef]
- Flanagan, S.A.; Hurtt, G.C.; Fisk, J.P.; Sahajpal, R.; Dubayah, R.O.; Hansen, M.; Sullivan, J.; Collatz, G.J. A Pseduo-spatial Method for Simulating Continental Scale Plant Migration. Working Paper, 2019. [Google Scholar]
- Hurtt, G.C.; Pacala, S.W.; Moorcroft, P.R.; Caspersen, J.; Shevliakova, E.; Houghton, R.A.; Moore, B. Projecting the future of the U.S. carbon sink. Proc. Natl. Acad. Sci. USA 2002, 99, 1389–1394. [Google Scholar] [CrossRef]
- Flanagan, S.; Hurtt, G.; Fisk, J.; Sahajpal, R.; Hansen, M.; Dolan, K.; Sullivan, J.; Zhao, M. Potential Vegetation and Carbon Redistribution in Northern North America from Climate Change. Climate 2016, 4, 2. [Google Scholar] [CrossRef]
- Dolan, K.A.; Hurtt, G.C.; Chambers, J.Q.; Dubayah, R.O.; Frolking, S.; Masek, J.G. Using ICESat’s Geoscience Laser Altimeter System (GLAS) to assess large-scale forest disturbance caused by hurricane Katrina. Remote Sens. Environ. 2011, 115, 86–96. [Google Scholar] [CrossRef]
- Hurtt, G.C.; Dubayah, R.; Drake, J.; Moorcroft, P.R.; Pacala, S.W.; Blair, J.B.; Fearon, M.G. Beyond potential vegetation:combining LIDAR data and a height-structured model for carbon studies. Ecol. Appl. 2004, 14, 873–883. [Google Scholar] [CrossRef]
- Dolan, K.A.; Hurtt, G.C.; Flanagan, S.A.; Fisk, J.P.; Sahajpal, R.; Huang, C.; Page, Y.L.; Dubayah, R.; Masek, J.G. Disturbance Distance: Quantifying forests’ vulnerability to disturbance under current and future conditions. Environ. Res. Lett. 2017, 12, 114015. [Google Scholar] [CrossRef]
- Silva, J.A.; Sedano, F.; Flanagan, S.; Ombe, Z.A.; Machoco, R.; Meque, C.H.; Sitoe, A.; Ribeiro, N.; Anderson, K.; Baule, S.; et al. Charcoal-related forest degradation dynamics in dry African woodlands: Evidence from Mozambique. Appl. Geogr. 2019, 107, 72–81. [Google Scholar] [CrossRef]
- Fisk, J.P.; Hurtt, G.C.; Chambers, J.Q.; Zeng, H.; Dolan, K.A.; Negrón-Juárez, R.I. The impacts of tropical cyclones on the net carbon balance of eastern US forests (1851–2000). Environ. Res. Lett. 2013, 8, 045017. [Google Scholar] [CrossRef]
- Hurtt, G.; Zhao, M.; Sahajpal, R.; Armstrong, A.; Birdsey, R.; Campbell, E.; Dolan, K.; Dubayah, R.; Fisk, J.P.; Flanagan, S.; et al. Beyond MRV: High-resolution forest carbon modeling for climate mitigation planning over Maryland, USA. Environ. Res. Lett. 2019, 14, 045013. [Google Scholar] [CrossRef]
- Hurtt, G.C.; Thomas, R.Q.; Fisk, J.P.; Dubayah, R.O.; Sheldon, S.L. The impact of fine-scale disturbances on the predictability of vegetation dynamics and carbon flux. PLoS ONE 2016, 11, e0152883. [Google Scholar] [CrossRef]
- Hurtt, G.; Wickland, D.; Jucks, K.; Bowman, K.; Brown, M.; Duren, R.; Hagen, S.; Verdy, A. NASA Carbon Monitoring System: Prototype Monitoring, Reporting, and Verification Progress Report and Future Plans, 2014. Available online: http://carbon.nasa.gov/pdfs/2014_CarbonMonitoringSystem_ProgressReport.pdf (accessed on 1 November 2015).
- Dubayah, R.; Goetz, S.J.; Blair, J.B.; Fatoyinbo, T.E.; Hansen, M.; Healey, S.P.; Hofton, M.A.; Hurtt, G.C.; Kellner, J.; Luthcke, S.B.; et al. The Global Ecosystem Dynamics Investigation. In Proceedings of the American Geophysical Union Fall Meeting 2014, San Francisco, CA, USA, 15–19 December 2014. [Google Scholar]
- Woodall, C.W.; Oswalt, C.M.; Westfall, J.A.; Perry, C.H.; Nelson, M.D.; Finley, A.O. An indicator of tree migration in forests of the eastern United States. For. Ecol. Manag. 2009, 5, 1434–1444. [Google Scholar] [CrossRef]
- Vogelmann, J.E.; Howard, S.M.; Yang, L.; Larson, C.R.; Wylie, B.K.; Driel, J.N. Van Completion of the 1990s National Land Cover Data set for the conterminous United States from Landsat Thematic Mapper data and ancillary data sources. Photogramm. Eng. Remote Sens. 2001, 67, 650–662. [Google Scholar]
- Wei, Y.; Liu, S.; Huntzinger, D.N.; Michalak, A.M.; Viovy, N.; Post, W.M.; Schwalm, C.R.; Schaefer, K.; Jacobson, A.R.; Lu, C.; et al. The North American Carbon Program Multi-scale Synthesis and Terrestrial Model Intercomparison Project—Part 2: Environmental driver data. Geosci. Model Dev. 2014, 7, 2875–2893. [Google Scholar] [CrossRef]
- Huntzinger, D.N.; Schwalm, C.; Michalak, A.M.; Schaefer, K.; King, A.W.; Wei, Y.; Jacobson, A.; Liu, S.; Cook, R.B.; Post, W.M.; et al. The North American Carbon Program Multi-Scale Synthesis and Terrestrial Model Intercomparison Project—Part 1: Overview and experimental design. Geosci. Model Dev. 2013, 6, 2121–2133. [Google Scholar] [CrossRef]
- Mearns, L.O.; Gutowski, W.; Jones, R.; Leung, R.; McGinnis, S.; Nunes, A.; Qian, Y. A Regional Climate Change Assessment Program for North America. Eos Trans. Am. Geophys. Union 2009, 90, 311. [Google Scholar] [CrossRef]
- Wang, W.J.; He, H.S.; Fraser, J.S.; Thompson III, F.R.; Shifley, S.R.; Spetich, M.A. LANDIS PRO: a landscape model that predicts forest composition and structure changes at regional scales. Ecography 2014, 37, 225–229. [Google Scholar] [CrossRef]
- Medvigy, D.; Wofsy, S.C.; Munger, J.W.; Hollinger, D.Y.; Moorcroft, P.R. Mechanistic scaling of ecosystem function and dynamics in space and time: Ecosystem Demography model version 2. J. Geophys. Res. 2009, 114, G01002. [Google Scholar] [CrossRef]
- Xu, X.; Medvigy, D.; Powers, J.S.; Becknell, J.M.; Guan, K. Diversity in plant hydraulic traits explains seasonal and inter-annual variations of vegetation dynamics in seasonally dry tropical forests. New Phytol. 2016, 212, 80–95. [Google Scholar] [CrossRef] [PubMed]
- Schaphoff, S.; Lucht, W.; Gerten, D.; Sitch, S.; Cramer, W.; Prentice, I.C. Terrestrial biosphere carbon storage under alternative climate projections. Clim. Chang. 2006, 74, 97–122. [Google Scholar] [CrossRef]
- Bachelet, D.M.; Lenihan, J.M.; Daly, C.; Neilson, R.P.; Ojima, D.S.; Parton, W.J. MC1: A Dynamic Vegetation Model for Estimating the Distribution of Vegetation and Associated Ecosystem Fluxes of Carbon, Nutrients, and Water; DIANE Publishing: Collingdale, PA, USA, 2001. [Google Scholar]
- Solomon, A.M.; Kirilenko, A.P. Climate Change and Terrestrial Biomass: What if Trees do not Migrate? Glob. Ecol. Biogeogr. Lett. 1997, 6, 139–148. [Google Scholar] [CrossRef]
- Brandt, L.; He, H.; Iverson, L.; Thompson, F.R.; Butler, P.; Handler, S.; Janowiak, M.; Shannon, P.D.; Swanston, C.; Albrecht, M.; et al. Central Hardwoods ecosystem vulnerability assessment and synthesis: A report from the Central Hardwoods Climate Change Response Framework project. Gen. Tech. Rep. 2014, 124, 1–254. [Google Scholar]
- Morin, X.; Viner, D.; Chuine, I. Tree species range shifts at a continental scale: New predictive insights from a process-based model. J. Ecol. 2008, 96, 784–794. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M.; Matthews, S.N.; Peters, M.P. Lessons Learned While Integrating Habitat, Dispersal, Disturbance, and Life-History Traits into Species Habitat Models under Climate Change. Ecosystems 2011, 14, 1005–1020. [Google Scholar] [CrossRef]
- Iverson, L.; Prasad, A.; Matthews, S. Modeling potential climate change impacts on the trees of the northeastern United States. Mitig. Adapt. Strateg. Glob. Chang. 2008, 13, 517–540. [Google Scholar] [CrossRef]
- Ranson, K.J.; Sun, G.; Kharuk, V.I.; Kovacs, K. Assessing tundra–taiga boundary with multi-sensor satellite data. Remote Sens. Environ. 2004, 93, 283–295. [Google Scholar] [CrossRef]
- Goldblum, D.; Rigg, L.S. Tree growth response to climate change at the deciduous–boreal forest ecotone, Ontario, Canada. Can. J. For. Res. 2005, 35, 2709–2718. [Google Scholar] [CrossRef]
- Chapin, F.S.; McGuire, A.D.; Ruess, R.W.; Hollingsworth, T.N.; Mack, M.C.; Johnstone, J.F.; Kasischke, E.S.; Euskirchen, E.S.; Jones, J.B.; Jorgenson, M.T.; et al. Resilience of Alaska’s boreal forest to climatic changeThis article is one of a selection of papers from The Dynamics of Change in Alaska’s Boreal Forests: Resilience and Vulnerability in Response to Climate Warming. Can. J. For. Res. 2010, 40, 1360–1370. [Google Scholar] [CrossRef]
- Grace, J.; Berninger, F.; Nagy, L. Impacts of Climate Change on the Tree Line. Ann. Bot. 2002, 90, 537–544. [Google Scholar] [CrossRef]
- Emanuel, W.R.; Shugart, H.H.; Stevenson, M.P. Climatic change and the broad-scale distribution of terrestrial ecosystem complexes. Clim. Chang. 1985, 7, 29–43. [Google Scholar] [CrossRef]
- Denslow, J.S. Patterns of plant species diversity during succession under different disturbance regimes. Oecologia 1980, 46, 18–21. [Google Scholar] [CrossRef]
- Collingham, Y.C.; Huntley, B. Impacts of habitat fragmentation and patch size upon migration rates. Ecol. Appl. 2000, 10, 131–144. [Google Scholar] [CrossRef]
- Sykes, M.T.; Prentice, I.C. Boreal forest futures: Modelling the controls on tree species range limits and transient responses to climate change. Water Air Soil Pollut. 1995, 82, 415–428. [Google Scholar] [CrossRef]
- Renton, M.; Childs, S.; Standish, R.; Shackelford, N. Plant migration and persistence under climate change in fragmented landscapes: Does it depend on the key point of vulnerability within the lifecycle? Ecol. Model. 2013, 249, 50–58. [Google Scholar] [CrossRef]
- Kurz, W.A.; Dymond, C.C.; Stinson, G.; Rampley, G.J.; Neilson, E.T.; Carroll, A.L.; Ebata, T.; Safranyik, L. Mountain pine beetle and forest carbon feedback to climate change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Kasischke, E.S.; Verbyla, D.L.; Rupp, T.S.; McGuire, A.D.; Murphy, K.A.; Jandt, R.; Barnes, J.L.; Hoy, E.E.; Duffy, P.A.; Calef, M.; et al. Alaska’s changing fire regime—Implications for the vulnerability of its boreal forests. Can. J. For. Res. 2010, 40, 1313–1324. [Google Scholar] [CrossRef]
- Zscheischler, J.; Michalak, A.M.; Schwalm, C.; Mahecha, M.D.; Huntzinger, D.N.; Reichstein, M.; Berthier, G.; Ciais, P.; Cook, R.B.; El-Masri, B.; et al. Impact of large-scale climate extremes on biospheric carbon fluxes: An intercomparison based on MsTMIP data. Glob. Biogeochem. Cycles 2014, 28, 585–600. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scenario | Percentage of Equilibrium Response Carbon | Percentage of Equilibrium Response Dominant PFT Sites | ||
|---|---|---|---|---|
| Direct | Even | Direct | Even | |
| 0.1 km distance 1x disturbance | 107 | 110 | 68 | 67 |
| 1.0 km distance 1x disturbance | 104 | 109 | 81 | 74 |
| 10.0 km distance 1x distrubance | 103 | 106 | 86 | 83 |
| 0.1 km distance 2x disturbance | 108 | 116 | 65 | 60 |
| 1.0 km distance 2x disturbance | 103 | 113 | 76 | 69 |
| 10.0 km distance 2x distrubance | 110 | 112 | 80 | 78 |
| 0.1 km distance 3x disturbance | 98 | 114 | 68 | 68 |
| 1.0 km distance 3x disturbance | 94 | 111 | 73 | 75 |
| 10.0 km distance 3x distrubance | 106 | 112 | 74 | 76 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flanagan, S.A.; Hurtt, G.C.; Fisk, J.P.; Sahajpal, R.; Zhao, M.; Dubayah, R.; Hansen, M.C.; Sullivan, J.H.; Collatz, G.J. Potential Transient Response of Terrestrial Vegetation and Carbon in Northern North America from Climate Change. Climate 2019, 7, 113. https://doi.org/10.3390/cli7090113
Flanagan SA, Hurtt GC, Fisk JP, Sahajpal R, Zhao M, Dubayah R, Hansen MC, Sullivan JH, Collatz GJ. Potential Transient Response of Terrestrial Vegetation and Carbon in Northern North America from Climate Change. Climate. 2019; 7(9):113. https://doi.org/10.3390/cli7090113
Chicago/Turabian StyleFlanagan, Steven A., George C. Hurtt, Justin P. Fisk, Ritvik Sahajpal, Maosheng Zhao, Ralph Dubayah, Matthew C. Hansen, Joe H. Sullivan, and G. James Collatz. 2019. "Potential Transient Response of Terrestrial Vegetation and Carbon in Northern North America from Climate Change" Climate 7, no. 9: 113. https://doi.org/10.3390/cli7090113
APA StyleFlanagan, S. A., Hurtt, G. C., Fisk, J. P., Sahajpal, R., Zhao, M., Dubayah, R., Hansen, M. C., Sullivan, J. H., & Collatz, G. J. (2019). Potential Transient Response of Terrestrial Vegetation and Carbon in Northern North America from Climate Change. Climate, 7(9), 113. https://doi.org/10.3390/cli7090113